
Genome-Wide Association Study and Transcriptome of Japanese Patients with Developmental Dysplasia of the Hip Demonstrates an Association with the Ferroptosis Signaling Pathway
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Demographics and Clinical Data
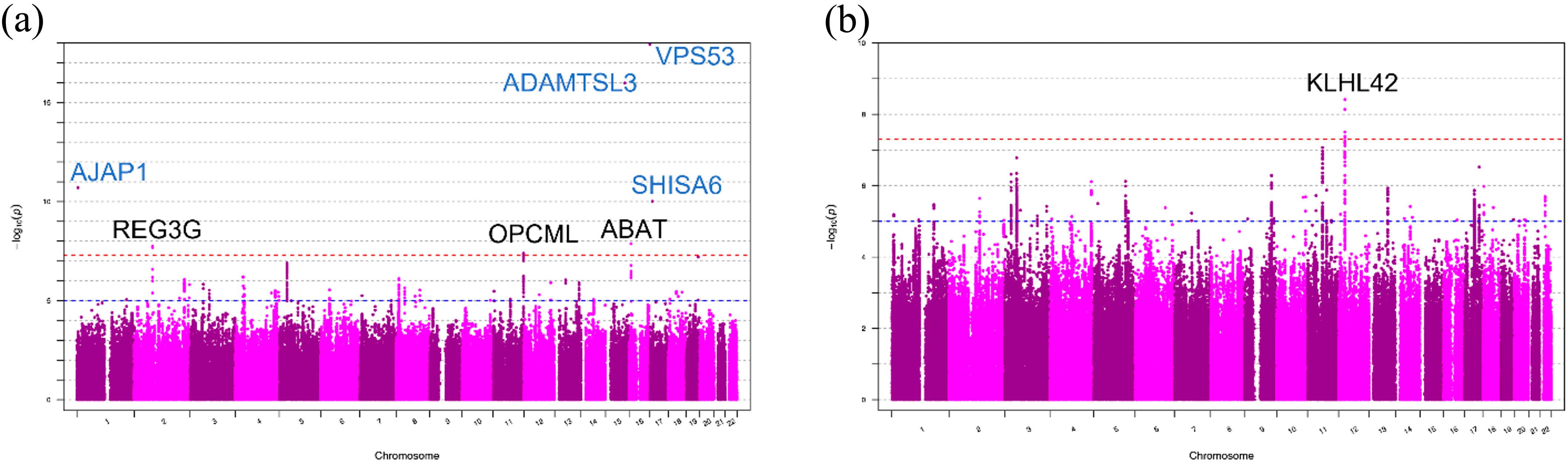
2.2. Genome-Wide Association Analyses of DDH in Japanese Patients
2.3. Replication Study and Genome-Wide Association Analyses of DDH in the UK Biobank Data Set
2.4. Gene Mapping of Disease-Related Variants and Gene Set Enrichment Analysis (GSEA)
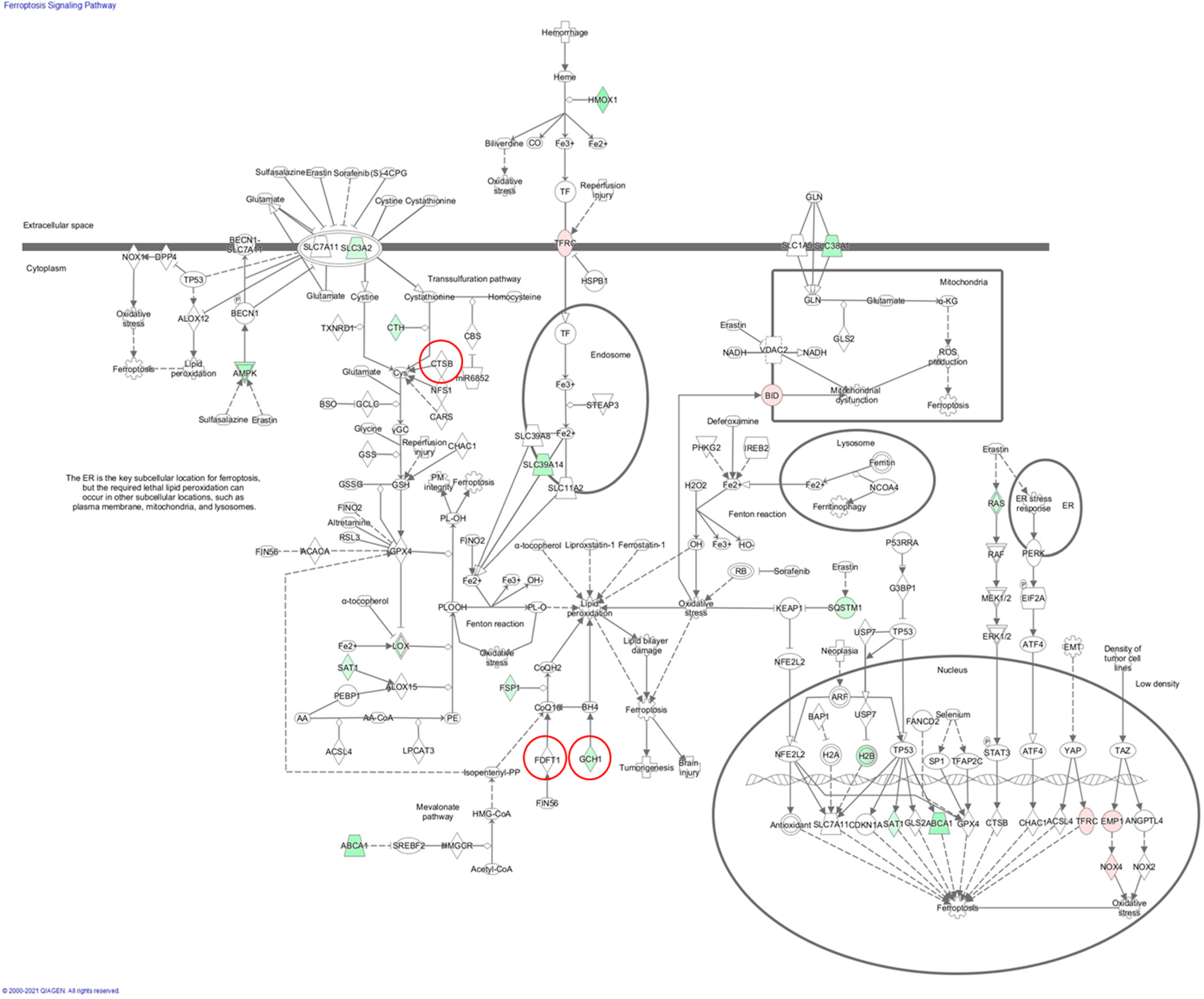
2.5. Pathway Enrichment Analysis for the Merged Gene Set
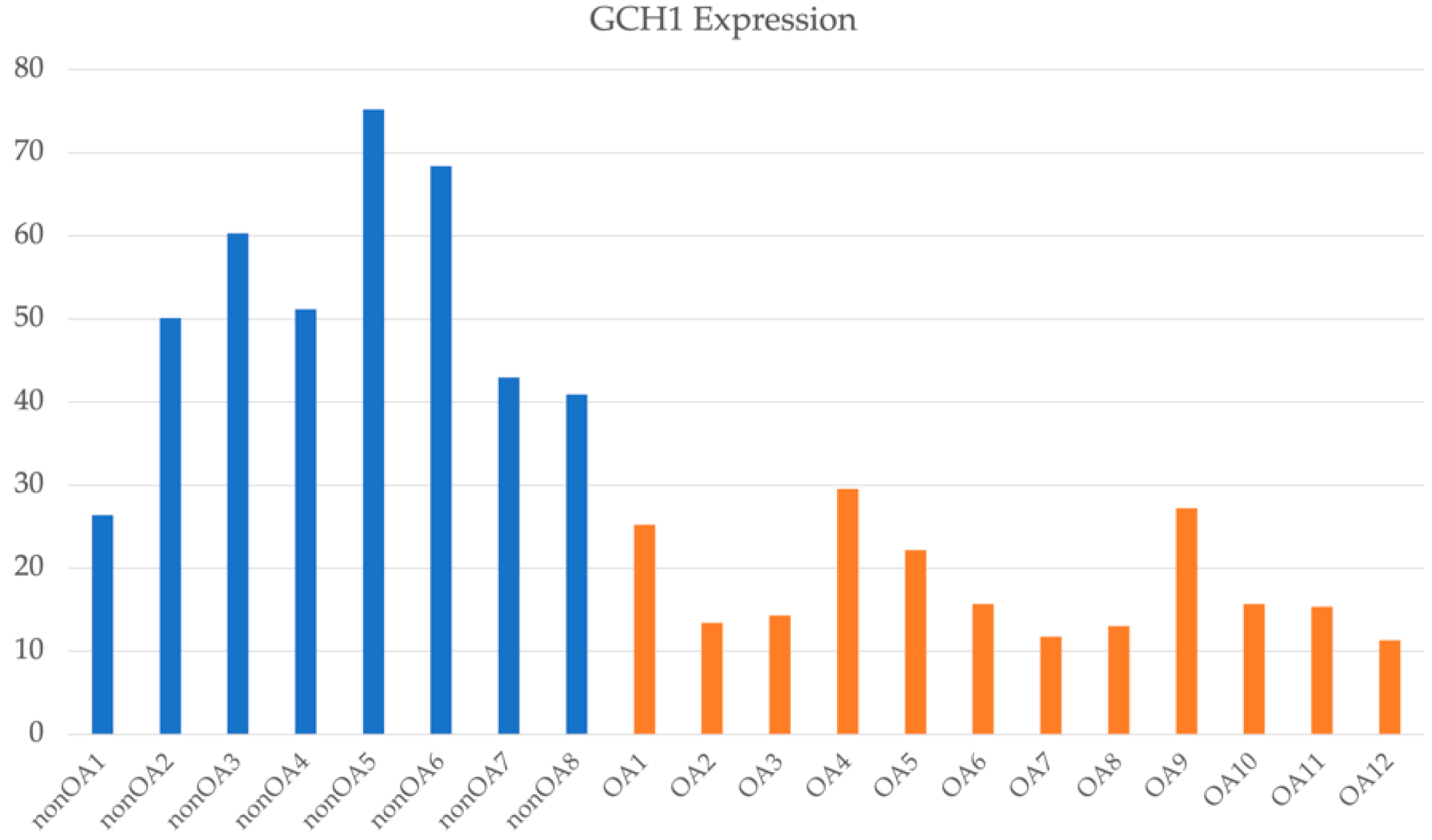
2.6. Transcriptome Analyses of OA Associated with DDH in Japanese Patients
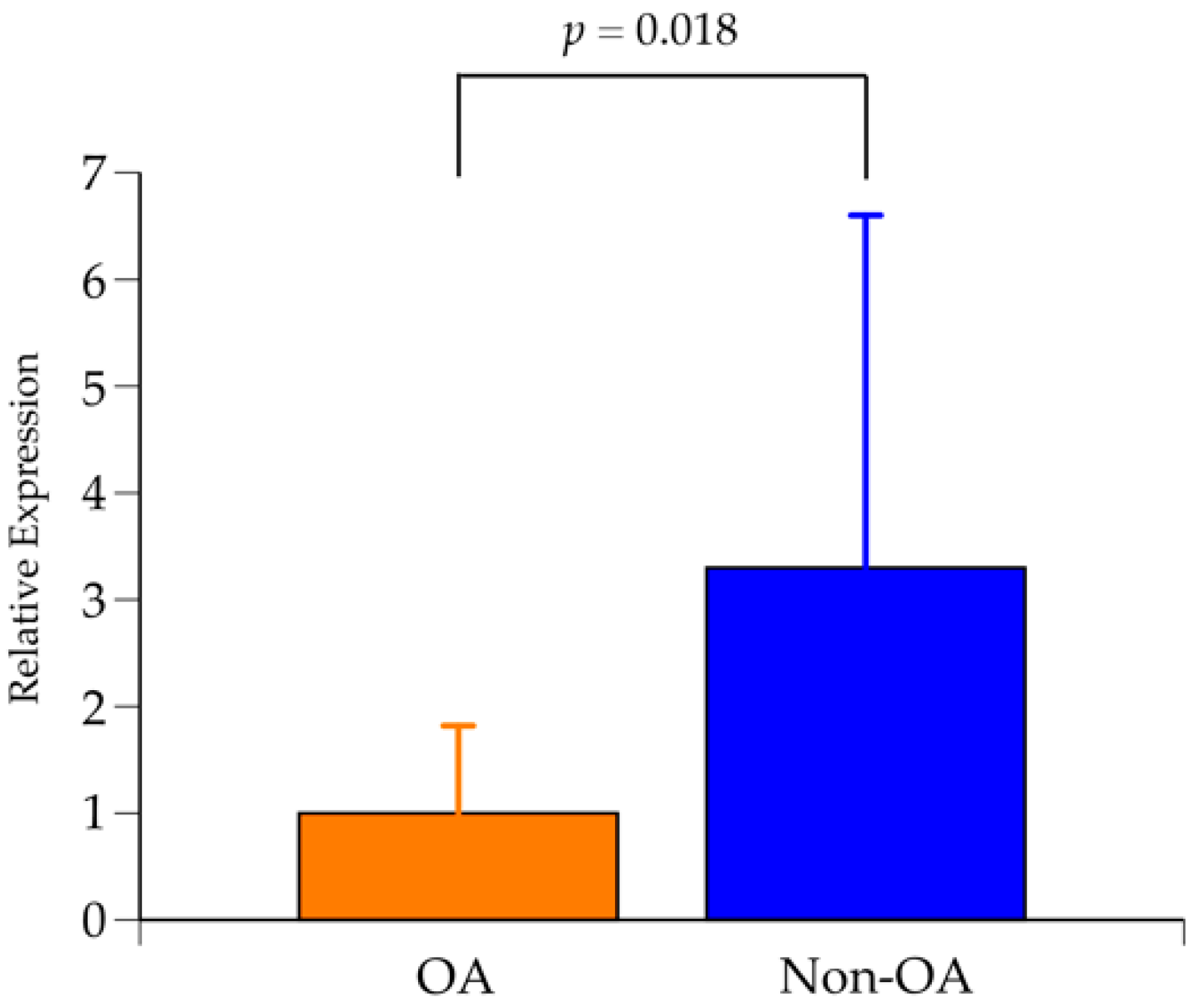
2.7. Expression Analysis of GCH1 Using Qualitative Real-Time Polymerase Chain Reaction (RT-PCR)
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Genotyping, QC, Imputation, and Association Analyses
4.3. Association Analysis and Gene Assignment
4.4. Quantitation of mRNA Expression in Chondrocytes from Patients with OA and Those with Femoral Neck Fracture
4.5. Gene Mapping of Disease-Related Variants and Gene Set Enrichment Analysis (GSEA)
4.6. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jacobsen, S.; Sonne-Holm, S. Hip dysplasia: A significant risk factor for the development of hip osteoarthritis. A cross-sectional survey. Rheumatology 2005, 44, 211–218. [Google Scholar] [CrossRef]
- Reijman, M.; Hazes, J.M.; Pols, H.A.; Koes, B.W.; Bierma-Zeinstra, S.M. Acetabular dysplasia predicts incident osteoarthritis of the hip: The Rotterdam study. Arthritis Rheum. 2005, 52, 787–793. [Google Scholar] [CrossRef]
- Jingushi, S.; Ohfuji, S.; Sofue, M.; Hirota, Y.; Itoman, M.; Matsumoto, T.; Hamada, Y.; Shindo, H.; Takatori, Y.; Yamada, H.; et al. Multiinstitutional epidemiological study regarding osteoarthritis of the hip in Japan. J. Orthop. Sci. 2010, 15, 626–631. [Google Scholar] [CrossRef]
- Jingushi, S.; Ohfuji, S.; Sofue, M.; Hirota, Y.; Itoman, M.; Matsumoto, T.; Hamada, Y.; Shindo, H.; Takatori, Y.; Yamada, H.; et al. Osteoarthritis hip joints in Japan: Involvement of acetabular dysplasia. J. Orthop. Sci. 2011, 16, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Loder, R.T.; Skopelja, E.N. The epidemiology and demographics of hip dysplasia. ISRN Orthop. 2011, 2011, 238607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiberg, G. Shelf operation in congenital dysplasia of the acetabulum and in subluxation and dislocation of the hip. J. Bone Jt. Surg. Am. 1953, 35-A, 65–80. [Google Scholar] [CrossRef]
- Yoshimura, N.; Campbell, L.; Hashimoto, T.; Kinoshita, H.; Okayasu, T.; Wilman, C.; Coggon, D.; Croft, P.; Cooper, C. Acetabular dysplasia and hip osteoarthritis in Britain and Japan. Br. J. Rheumatol. 1998, 37, 1193–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Wicart, P.; Kawasaki, T.; Huang, J.; Ushiyama, T.; Hukuda, S.; Courpied, J. Prevalence of hip osteoarthritis and acetabular dysplasia in french and japanese adults. Rheumatology 2000, 39, 745–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenanidis, E.; Gkekas, N.K.; Karasmani, A.; Anagnostis, P.; Christofilopoulos, P.; Tsiridis, E. Genetic Predisposition to Developmental Dysplasia of the Hip. J. Arthroplast. 2020, 35, 291–300.e1. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, J.J.; Bracha, P.; Liu, S.S.; Piyaworakhun, S.; Goetz, D.D.; Johnston, R.C. Survivorship of a Charnley total hip arthroplasty. A concise follow-up, at a minimum of thirty-five years, of previous reports. J. Bone Jt. Surg. Am. 2009, 91, 2617–2621. [Google Scholar] [CrossRef] [Green Version]
- Klapach, A.S.; Callaghan, J.J.; Goetz, D.D.; Olejniczak, J.P.; Johnston, R.C. Charnley total hip arthroplasty with use of improved cementing techniques: A minimum twenty-year follow-up study. J. Bone Joint. Surg. Am. 2001, 83, 1840–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, D.; Yamada, N.; Mori, Y.; Oyama, M.; Ohtsu, S.; Kuwahara, Y.; Baba, K.; Tanaka, H.; Aizawa, T.; Hanada, S.; et al. Mid-term results of a new femoral prosthesis using Ti-Nb-Sn alloy with low Young’s modulus. BMC Musculoskelet. Disord. 2021, 22, 987. [Google Scholar] [CrossRef] [PubMed]
- Southam, L.; Rodriguez-Lopez, J.; Wilkins, J.M.; Pombo-Suarez, M.; Snelling, S.; Gomez-Reino, J.J.; Chapman, K.; Gonzalez, A.; Loughlin, J. An SNP in the 5’-UTR of GDF5 is associated with osteoarthritis susceptibility in Europeans and with in vivo differences in allelic expression in articular cartilage. Hum. Mol. Genet. 2007, 16, 2226–2232. [Google Scholar] [CrossRef]
- Masuya, H.; Nishida, K.; Furuichi, T.; Toki, H.; Nishimura, G.; Kawabata, H.; Yokoyama, H.; Yoshida, A.; Tominaga, S.; Nagano, J.; et al. A novel dominant-negative mutation in Gdf5 generated by ENU mutagenesis impairs joint formation and causes osteoarthritis in mice. Hum. Mol. Genet. 2007, 16, 2366–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loughlin, J.; Sinsheimer, J.S.; Carr, A.; Chapman, K. The CALM1 core promoter polymorphism is not associated with hip osteoarthritis in a United Kingdom Caucasian population. Osteoarthr. Cartil. 2006, 14, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Mototani, H.; Mabuchi, A.; Saito, S.; Fujioka, M.; Iida, A.; Takatori, Y.; Kotani, A.; Kubo, T.; Nakamura, K.; Sekine, A.; et al. A functional single nucleotide polymorphism in the core promoter region of CALM1 is associated with hip osteoarthritis in Japanese. Hum. Mol. Genet. 2005, 14, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Kizawa, H.; Saitoh, M.; Kou, I.; Miyazono, K.; Ikegawa, S. Mechanisms for asporin function and regulation in articular cartilage. J. Biol. Chem. 2007, 282, 32185–32192. [Google Scholar] [CrossRef] [Green Version]
- Rouault, K.; Scotet, V.; Autret, S.; Gaucher, F.; Dubrana, F.; Tanguy, D.; El Rassi, C.Y.; Fenoll, B.; Ferec, C. Evidence of association between GDF5 polymorphisms and congenital dislocation of the hip in a Caucasian population. Osteoarthr. Cartil. 2010, 18, 1144–1149. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, A.; Nakamura, S.; Takatori, Y.; Ikegawa, S. Familial osteoarthritis of the hip joint associated with acetabular dysplasia maps to chromosome 13q. Am. J. Hum. Genet. 2006, 79, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Feldman, G.; Dalsey, C.; Fertala, K.; Azimi, D.; Fortina, P.; Devoto, M.; Pacifici, M.; Parvizi, J. The Otto Aufranc Award: Identification of a 4 Mb region on chromosome 17q21 linked to developmental dysplasia of the hip in one 18-member, multigeneration family. Clin. Orthop. Relat. Res. 2010, 468, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Hatzikotoulas, K.; Roposch, A.; Consortium, D.D.H.C.C.; Shah, K.M.; Clark, M.J.; Bratherton, S.; Limbani, V.; Steinberg, J.; Zengini, E.; Warsame, K.; et al. Genome-wide association study of developmental dysplasia of the hip identifies an association with GDF5. Commun. Biol. 2018, 1, 56. [Google Scholar] [CrossRef]
- Harsanyi, S.; Zamborsky, R.; Krajciova, L.; Kokavec, M.; Danisovic, L. Genetic Study of IL6, GDF5 and PAPPA2 in Association with Developmental Dysplasia of the Hip. Genes 2021, 12, 986. [Google Scholar] [CrossRef]
- Harsanyi, S.; Zamborsky, R.; Krajciova, L.; Bohmer, D.; Kokavec, M.; Danisovic, L. Association Analysis of GDF5 and Contributing Factors in Developmental Dysplasia of the Hip in Infants. Ortop. Traumatol. Rehabil. 2021, 23, 335–339. [Google Scholar] [CrossRef]
- Feldman, G.J.; Parvizi, J.; Sawan, H.; Erickson, J.A.; Peters, C.L. Linkage mapping and whole exome sequencing identify a shared variant in CX3CR1 in a large multi-generation family. J. Arthroplast. 2014, 29, 238–241. [Google Scholar] [CrossRef]
- Feldman, G.; Offemaria, A.; Sawan, H.; Parvizi, J.; Freeman, T.A. A murine model for developmental dysplasia of the hip: Ablation of CX3CR1 affects acetabular morphology and gait. J. Transl. Med. 2017, 15, 233. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, X.; Zhao, Q.; Wang, E.; Wang, L.; Cheng, J.; Zhang, L.; Wang, B. CX3CR1 polymorphisms associated with an increased risk of developmental dysplasia of the hip in human. J. Orthop. Res. 2017, 35, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wang, C.; Hao, Z.; Dai, J.; Chen, D.; Xu, Z.; Shi, D.; Mao, P.; Teng, H.; Gao, X.; et al. A common variant of ubiquinol-cytochrome c reductase complex is associated with DDH. PLoS ONE 2015, 10, e0120212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.J.; Zeiss, C.; Chew, E.Y.; Tsai, J.Y.; Sackler, R.S.; Haynes, C.; Henning, A.K.; SanGiovanni, J.P.; Mane, S.M.; Mayne, S.T.; et al. Complement factor H polymorphism in age-related macular degeneration. Science 2005, 308, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.; Patel, N.; Turcotte, M.; Bosse, Y.; Pare, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Nagasaki, M.; Yasuda, J.; Katsuoka, F.; Nariai, N.; Kojima, K.; Kawai, Y.; Yamaguchi-Kabata, Y.; Yokozawa, J.; Danjoh, I.; Saito, S.; et al. Rare variant discovery by deep whole-genome sequencing of 1,070 Japanese individuals. Nat. Commun. 2015, 6, 8018. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Mimori, T.; Kojima, K.; Nariai, N.; Danjoh, I.; Saito, R.; Yasuda, J.; Yamamoto, M.; Nagasaki, M. Japonica array: Improved genotype imputation by designing a population-specific SNP array with 1070 Japanese individuals. J. Hum. Genet. 2015, 60, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Shiga, Y.; Nishiguchi, K.M.; Kawai, Y.; Kojima, K.; Sato, K.; Fujita, K.; Takahashi, M.; Omodaka, K.; Araie, M.; Kashiwagi, K.; et al. Genetic analysis of Japanese primary open-angle glaucoma patients and clinical characterization of risk alleles near CDKN2B-AS1, SIX6 and GAS7. PLoS ONE 2017, 12, e0186678. [Google Scholar] [CrossRef] [PubMed]
- Naito, T.; Yokoyama, N.; Kakuta, Y.; Ueno, K.; Kawai, Y.; Onodera, M.; Moroi, R.; Kuroha, M.; Kanazawa, Y.; Kimura, T.; et al. Clinical and genetic risk factors for decreased bone mineral density in Japanese patients with inflammatory bowel disease. J. Gastroenterol. Hepatol. 2018, 33, 1873–1881. [Google Scholar] [CrossRef]
- Hitomi, Y.; Ueno, K.; Kawai, Y.; Nishida, N.; Kojima, K.; Kawashima, M.; Aiba, Y.; Nakamura, H.; Kouno, H.; Kouno, H.; et al. POGLUT1, the putative effector gene driven by rs2293370 in primary biliary cholangitis susceptibility locus chromosome 3q13.33. Sci. Rep. 2019, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Spector, T.D.; Williams, F.M. The UK Adult Twin Registry (TwinsUK). Twin. Res. Hum. Genet. 2006, 9, 899–906. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Sun, K.; Yu, S.; Luo, J.; Guo, J.; Lin, J.; Wang, G.; Guo, Z.; Ye, Y.; Guo, F. Chondrocyte ferroptosis contribute to the progression of osteoarthritis. J. Orthop. Translat. 2021, 27, 33–43. [Google Scholar] [CrossRef]
- Huang, S.W.; Wang, W.T.; Lin, L.F.; Liao, C.D.; Liou, T.H.; Lin, H.W. Association between psychiatric disorders and osteoarthritis: A nationwide longitudinal population-based study. Medicine 2016, 95, e4016. [Google Scholar] [CrossRef]
- Guo, Z.; Lin, J.; Sun, K.; Guo, J.; Yao, X.; Wang, G.; Hou, L.; Xu, J.; Guo, J.; Guo, F. Deferoxamine Alleviates Osteoarthritis by Inhibiting Chondrocyte Ferroptosis and Activating the Nrf2 Pathway. Front. Pharmacol. 2022, 13, 791376. [Google Scholar] [CrossRef]
- Lv, M.; Cai, Y.; Hou, W.; Peng, K.; Xu, K.; Lu, C.; Yu, W.; Zhang, W.; Liu, L. The RNA-binding protein SND1 promotes the degradation of GPX4 by destabilizing the HSPA5 mRNA and suppressing HSPA5 expression, promoting ferroptosis in osteoarthritis chondrocytes. Inflamm. Res. 2022, 71, 461–472. [Google Scholar] [CrossRef]
- Yang, J.; Hu, S.; Bian, Y.; Yao, J.; Wang, D.; Liu, X.; Guo, Z.; Zhang, S.; Peng, L. Targeting Cell Death: Pyroptosis, Ferroptosis, Apoptosis and Necroptosis in Osteoarthritis. Front. Cell Dev. Biol. 2021, 9, 789948. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, Y.; Sun, W.; Zhang, Z.; Liu, J.; Yang, W.; Yuan, W.; Yi, Y.; Wang, J.; Liu, J. D-mannose alleviates osteoarthritis progression by inhibiting chondrocyte ferroptosis in a HIF-2alpha-dependent manner. Cell Prolif. 2021, 54, e13134. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Wei, W.; Wu, D.; Huang, F.; Li, M.; Li, W.; Yin, J.; Peng, Y.; Lu, Y.; Zhao, Q.; et al. Blockade of GCH1/BH4 Axis Activates Ferritinophagy to Mitigate the Resistance of Colorectal Cancer to Erastin-Induced Ferroptosis. Front. Cell Dev. Biol. 2022, 10, 810327. [Google Scholar] [CrossRef]
- Lu, M.; Li, J.; Fan, X.; Xie, F.; Fan, J.; Xiong, Y. Novel Immune-Related Ferroptosis Signature in Esophageal Cancer: An Informatics Exploration of Biological Processes Related to the TMEM161B-AS1/hsa-miR-27a-3p/GCH1 Regulatory Network. Front. Genet. 2022, 13, 829384. [Google Scholar] [CrossRef]
- Kuang, F.; Liu, J.; Li, C.; Kang, R.; Tang, D. Cathepsin B is a mediator of organelle-specific initiation of ferroptosis. Biochem. Biophys. Res. Commun. 2020, 533, 1464–1469. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhang, C.; Wang, X.; Zou, X.; Xiang, Z.; Wang, Z.; Gui, B.; Lin, T.; Hu, H. Identification of FDFT1 as a potential biomarker associated with ferroptosis in ccRCC. Cancer Med. 2022, 11, 3993–4004. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Shi, D.; Zhu, P.; Qin, J.; Ni, H.; Xu, Y.; Yao, C.; Zhu, L.; Zhu, H.; Zhao, B.; et al. Association of a single nucleotide polymorphism in growth differentiate factor 5 with congenital dysplasia of the hip: A case-control study. Arthritis Res. Ther. 2008, 10, R126. [Google Scholar] [CrossRef] [Green Version]
- Hampton, S.N.; Nakonezny, P.A.; Richard, H.M.; Wells, J.E. Pain catastrophizing, anxiety, and depression in hip pathology. Bone Joint. J. 2019, 101-B, 800–807. [Google Scholar] [CrossRef]
- Gudmundsson, P.; Nakonezny, P.A.; Lin, J.; Owhonda, R.; Richard, H.; Wells, J. Functional improvement in hip pathology is related to improvement in anxiety, depression, and pain catastrophizing: An intricate link between physical and mental well-being. BMC Musculoskelet. Disord. 2021, 22, 133. [Google Scholar] [CrossRef]
- Gershlick, D.C.; Ishida, M.; Jones, J.R.; Bellomo, A.; Bonifacino, J.S.; Everman, D.B. A neurodevelopmental disorder caused by mutations in the VPS51 subunit of the GARP and EARP complexes. Hum. Mol. Genet. 2019, 28, 1548–1560. [Google Scholar] [CrossRef] [Green Version]
- Kakuta, Y.; Kawai, Y.; Naito, T.; Hirano, A.; Umeno, J.; Fuyuno, Y.; Liu, Z.; Li, D.; Nakano, T.; Izumiyama, Y.; et al. A Genome-wide Association Study Identifying RAP1A as a Novel Susceptibility Gene for Crohn’s Disease in Japanese Individuals. J. Crohns Colitis 2019, 13, 648–658. [Google Scholar] [CrossRef]
- Jiang, L.; Zheng, Z.; Fang, H.; Yang, J. A generalized linear mixed model association tool for biobank-scale data. Nat. Genet. 2021, 53, 1616–1621. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Kamimura, M.; Mori, Y.; Sugahara-Tobinai, A.; Takai, T.; Itoi, E. Impaired Fracture Healing Caused by Deficiency of the Immunoreceptor Adaptor Protein DAP12. PLoS ONE 2015, 10, e0128210. [Google Scholar] [CrossRef] [PubMed]
- Aki, T.; Hashimoto, K.; Ogasawara, M.; Itoi, E. A whole-genome transcriptome analysis of articular chondrocytes in secondary osteoarthritis of the hip. PLoS ONE 2018, 13, e0199734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, H.; Mori, Y.; Kogure, A.; Tanaka, H.; Kamimura, M.; Masahashi, N.; Hanada, S.; Itoi, E. Effects of intramedullary nails composed of a new beta-type Ti-Nb-Sn alloy with low Young’s modulus on fracture healing in mouse tibiae. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 2841–2848. [Google Scholar] [CrossRef]
- Mori, Y.; Fujisawa, H.; Kamimura, M.; Kogure, A.; Tanaka, H.; Mori, N.; Masahashi, N.; Aizawa, T. Acceleration of Fracture Healing in Mouse Tibiae Using Intramedullary Nails Composed of beta-Type TiNbSn Alloy with Low Young’s Modulus. Tohoku J. Exp. Med. 2021, 255, 135–142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n = 238 | |
|---|---|
| Sex (%) | 206 females (86.5%) |
| Age (years) | 60.0 ± 15.0 |
| Positive family history of DDH (%) | 94 (39.4%) |
| Treatment history in childhood (%) | 110 (46.2%) |
| Surgical treatment (%) | 161 (67.6%) |
| Total hip arthroplasty (%) | 72 (30.0%) |
| (a) Japanese GWAS | ||||||
| CHR | SNP | BP | OR | p | Nearest gene | GTEx eQTL gene |
| 1 | rs11802858 | 4682515 | 0.3631 | 1.98 × 10−11 | AJAP1 | NS |
| 2 | rs149003127 | 79236134 | 0.2296 | 1.82 × 10−8 | REG3G, REG1B | NA |
| 11 | rs55669018 | 132871263 | 1.785 | 4.09 × 10−8 | OPCML | NS |
| 15 | rs2554380 | 84315884 | 4.022 | 1.04 × 10−16 | ADAMTSL3 | ADAMTSL3, CSPG4P11, DNM1P41, DNM1P51, GOLGA2P7, GOLGA6L4, GOLGA6L5P, LINC00933, RP11-182J1.14, RP11-671M22.4, SCAND2P, UBE2Q2L, UBE2Q2P1 |
| 16 | rs77485026 | 8846734 | 0.1884 | 1.34 × 10−8 | ABAT | NA |
| 17 | rs79657649 | 505105 | 0.1957 | 1.19 × 10−18 | VPS53 | NS |
| 17 | rs17699467 | 11359275 | 0.351 | 9.82 × 10−11 | SHISA6 | NS |
| (b) UK Biobank GWAS | ||||||
| CHR | dbSNPID | Position | Beta | p | Nearest gene | GTEx eQTL gene |
| 12 | rs11049197 | 27996732 | 0.203 | 2.62 × 10−9 | KLHL42 | NS |
| (a) | ||||
| Ingenuity canonical pathways | Nominal p-value | BH FDR | Genes | |
| Ferroptosis signaling pathway | 0.0015 | 0.065 | Japan (CTSB, FDFT1, GCH1) | |
| (b) | ||||
| Disease category (ID) | Nominal p-value | BF p-value | Genes | |
| Osteoarthritis of the hip (C0029410; DisGeNET curated) | 1.16 × 10−12 | 2.74 × 10−9 | UK (IL11, MAPT, ITIH1, CRHR1, LMX1B, ASTN2, LTBP3) | |
| Bipolar disorder (C0005586; DisGeNET curated) | 4.05 × 10−6 | 9.57 × 10−3 | UK (ITIH1, CRHR1, ITIH3, PDLIM5, ITIH4, STAB1, NEK4, PBRM1, MTHFD1, DLG2, ASTN2) | |
| (c) | ||||
| Category | Nominal p-value | Adj. p | Genes | |
| Cytoskeleton organization (GO: 0007010) | 5.00 × 10−6 | 0.00296 | Japan (ARHGAP28, DNAH7, DNAH8, FAT1, FLNB, MAP6, SIGLEC15) UK (ABLIM1, ABLIM2, BRSK1, DNAH1, HIP1, LMOD3, MAPT, NISCH, SYNE2, SYNPO, TRIM32, TWF2) | |
| Cell–cell signaling (GO: 0007267) | 1.08 × 10−6 | 0.00296 | Japan (ABAT, C1QTNF3, CELF4, DGKI, FAT1, GLP1R, SHISA6) UK (BRSK1, CRHR1, CTBP2, DLG2, ESR2, HTR3E, IL11, MAPT, NISCH, PSEN1, STAB1, SYNPO, TMF1, WNT3) | |
| (d) | ||||
| Ingenuity canonical pathways | p-value | BH FDR | Genes | |
| Ferroptosis signaling pathway | 0.006026 | 0.477529 | Japan (CTSB, FDFT1, GCH1) UK (BAP1) | |
| (e) | ||||
| Disease category (ID) | Nominal p-value | BF p-value | Genes | |
| Osteoarthritis of the hip (C0029410; DisGeNET curated) | 1.66 × 10−11 | 5.16 × 10−8 | Japan (no hit) UK (IL11, MAPT, ITIH1, CRHR1, LMX1B, ASTN2, LTBP3) | |
| Bipolar disorder (C0005586; DisGeNET curated) | 5.66 × 10−6 | 1.75 × 10−2 | Japan (IMPA1, DNAH8) UK (ITIH1, CRHR1, ITIH3, PDLIM5, ITIH4, STAB1, NEK4, PBRM1, MTHFD1, DLG2, ASTN2) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, Y.; Ueno, K.; Chiba, D.; Hashimoto, K.; Kawai, Y.; Baba, K.; Tanaka, H.; Aki, T.; Ogasawara, M.; Shibasaki, N.; et al. Genome-Wide Association Study and Transcriptome of Japanese Patients with Developmental Dysplasia of the Hip Demonstrates an Association with the Ferroptosis Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 5019. https://doi.org/10.3390/ijms24055019
Mori Y, Ueno K, Chiba D, Hashimoto K, Kawai Y, Baba K, Tanaka H, Aki T, Ogasawara M, Shibasaki N, et al. Genome-Wide Association Study and Transcriptome of Japanese Patients with Developmental Dysplasia of the Hip Demonstrates an Association with the Ferroptosis Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(5):5019. https://doi.org/10.3390/ijms24055019
Chicago/Turabian StyleMori, Yu, Kazuko Ueno, Daisuke Chiba, Ko Hashimoto, Yosuke Kawai, Kazuyoshi Baba, Hidetatsu Tanaka, Takashi Aki, Masanori Ogasawara, Naoto Shibasaki, and et al. 2023. "Genome-Wide Association Study and Transcriptome of Japanese Patients with Developmental Dysplasia of the Hip Demonstrates an Association with the Ferroptosis Signaling Pathway" International Journal of Molecular Sciences 24, no. 5: 5019. https://doi.org/10.3390/ijms24055019
APA StyleMori, Y., Ueno, K., Chiba, D., Hashimoto, K., Kawai, Y., Baba, K., Tanaka, H., Aki, T., Ogasawara, M., Shibasaki, N., Tokunaga, K., Aizawa, T., & Nagasaki, M. (2023). Genome-Wide Association Study and Transcriptome of Japanese Patients with Developmental Dysplasia of the Hip Demonstrates an Association with the Ferroptosis Signaling Pathway. International Journal of Molecular Sciences, 24(5), 5019. https://doi.org/10.3390/ijms24055019