
Post-Transcriptional and Post-Translational Modifications in Telomerase Biogenesis and Recruitment to Telomeres


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Yeast Telomerase
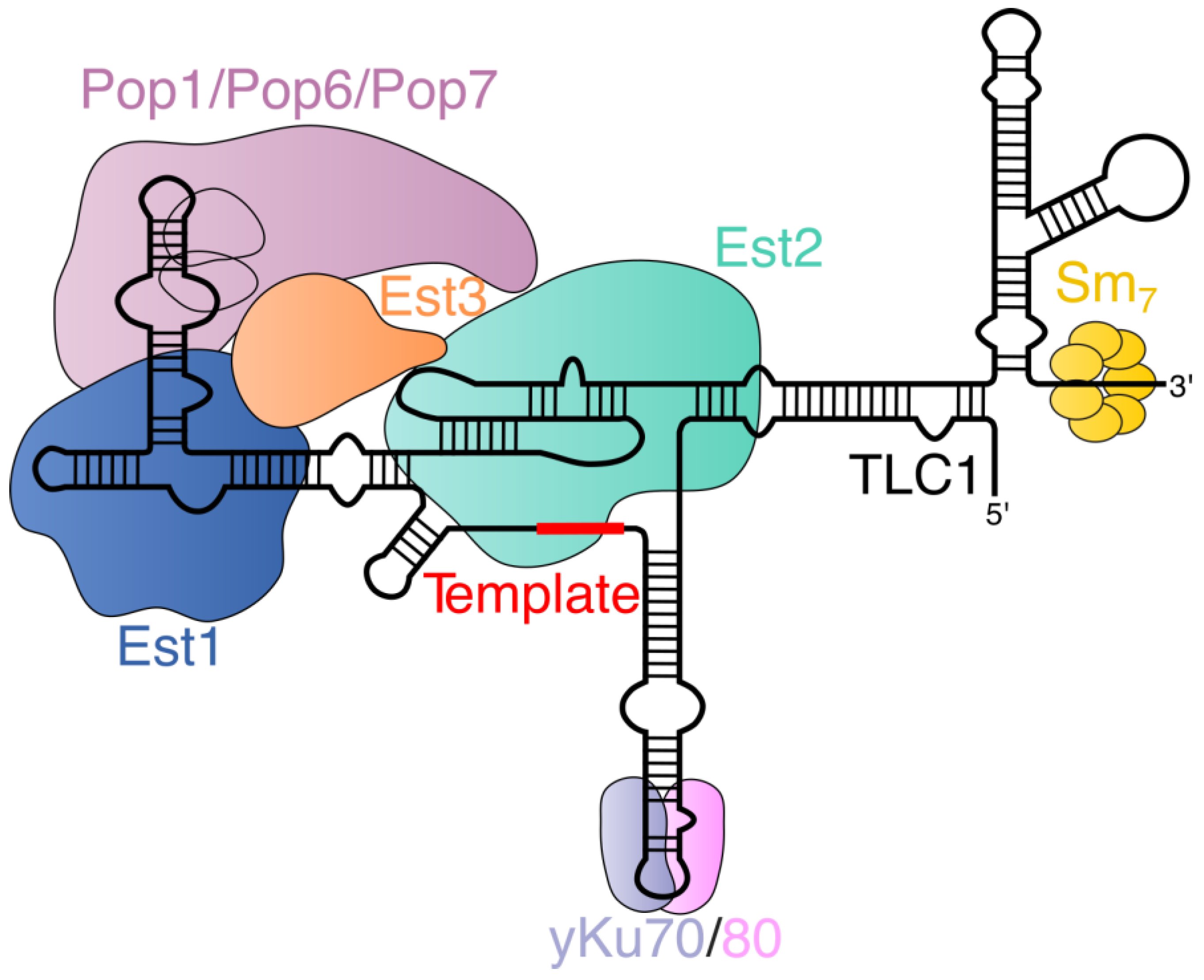
2.1. Yeast Telomerase Holoenzyme Composition
2.2. Telomerase RNA Processing in Yeast
2.3. Assembly of the Yeast Telomerase RNA–Protein Complex
2.4. Telomerase Recruitment to Telomeres in Yeast
2.5. Post-Translational Modifications (PTMs) of the Telomerase Components in Yeast
3. Telomerase in Vertebrates
3.1. Human Telomerase Holoenzyme Composition
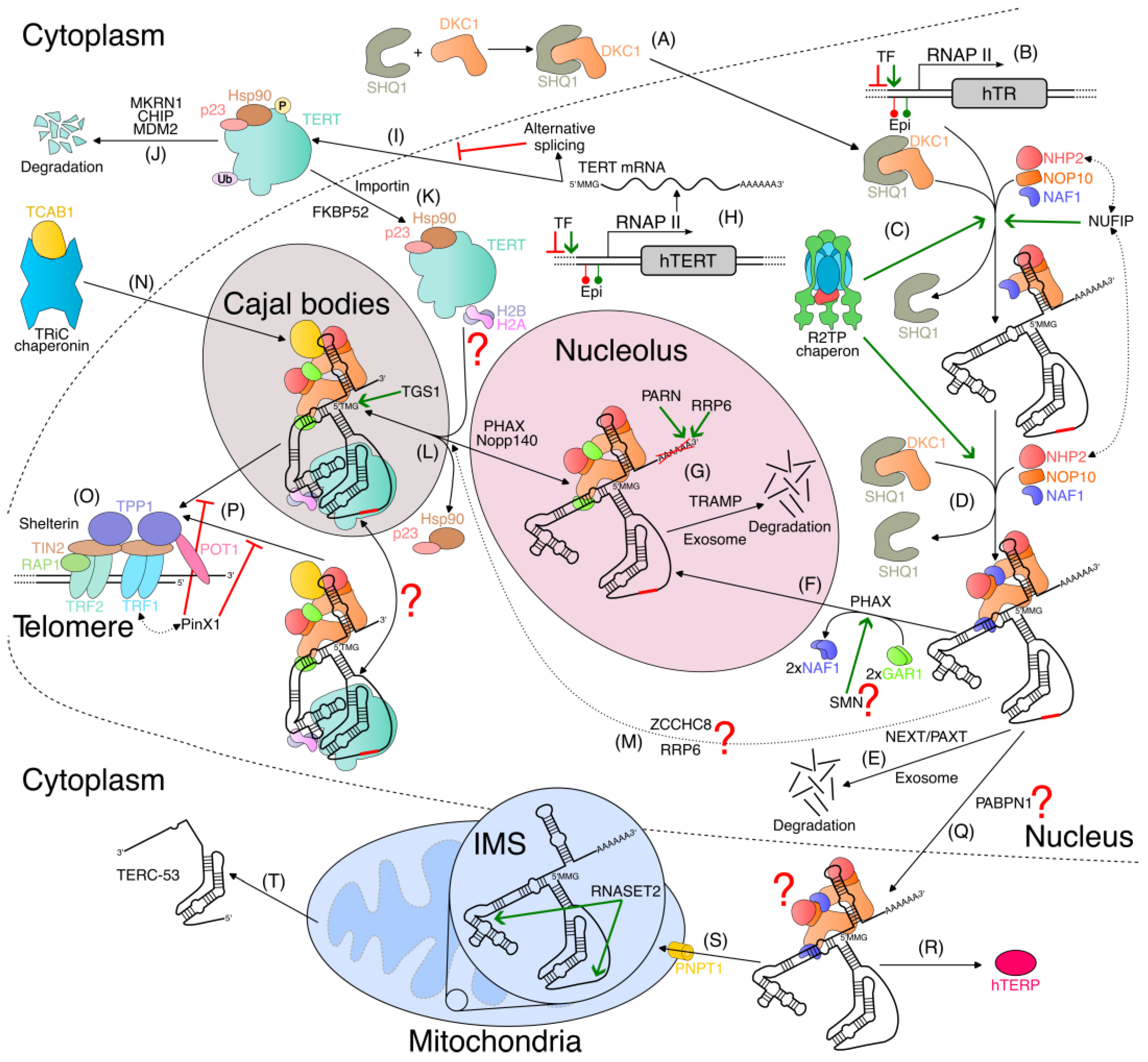
3.2. hTR Biosynthesis and Early Processing
3.3. H/ACA Motif Pre-Assembly of Telomerase RNP
3.4. Posttranslational Modifications in Biogenesis of Telomerase RNP
3.5. Assembly of Active Telomerase Complex
3.6. Transport of Human Telomerase RNP
3.7. hTR Modifications in Telomerase Biogenesis
3.8. hTERT Biosynthesis Regulation
3.8.1. hTERT Alternative Splicing
3.8.2. hTERT Localization
3.8.3. hTERT Phosphorylation
3.8.4. hTERT Ubiquitination and SUMOylation
3.8.5. hTERT Sequestration
3.9. Delivery of Telomerase Holoenzyme to Telomeres in Vertebrates
3.10. Telomerase Recruitment to Telomeres in Vertebrates

4. Common and Specific Motifs of Telomerase Biogenesis in Yeast and Vertebrates
5. Prospects
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Greider, C.W.; Blackburn, E.H. Identification of a Specific Telomere Terminal Transferase Activity in Tetrahymena Extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Morin, G.B. The Human Telomere Terminal Transferase Enzyme Is a Ribonucleoprotein That Synthesizes TTAGGG Repeats. Cell 1989, 59, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Olovnikov, A.M. A Theory of Marginotomy. J. Theor. Biol. 1973, 41, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Chow, T.T.; Zhao, Y.; Mak, S.S.; Shay, J.W.; Wright, W.E. Early and Late Steps in Telomere Overhang Processing in Normal Human Cells: The Position of the Final RNA Primer Drives Telomere Shortening. Genes Dev. 2012, 26, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, V.; Lingner, J. TERRA Promotes Telomere Shortening through Exonuclease 1-Mediated Resection of Chromosome Ends. PLoS Genet. 2012, 8, e1002747. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, M.; Rossiello, F.; Clerici, M.; Barozzi, S.; Cittaro, D.; Kaplunov, J.M.; Bucci, G.; Dobreva, M.; Matti, V.; Beausejour, C.M.; et al. Telomeric DNA Damage Is Irreparable and Causes Persistent DNA-Damage-Response Activation. Nat. Cell Biol. 2012, 14, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G.; Jurk, D.; Marques, F.D.M.; Correia-Melo, C.; Hardy, T.; Gackowska, A.; Anderson, R.; Taschuk, M.; Mann, J.; Passos, J.F. Telomeres Are Favoured Targets of a Persistent DNA Damage Response in Ageing and Stress-Induced Senescence. Nat. Commun. 2012, 3, 708. [Google Scholar] [CrossRef] [Green Version]
- Blasco, M.A. Telomeres and Human Disease: Ageing, Cancer and Beyond. Nat. Rev. Genet. 2005, 6, 611–622. [Google Scholar] [CrossRef]
- Holt, S.E.; Shay, J.W. Role of Telomerase in Cellular Proliferation and Cancer. J. Cell. Physiol. 1999, 180, 10–18. [Google Scholar] [CrossRef]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Weng, N. Telomere and Adaptive Immunity. Mech. Ageing Dev. 2008, 129, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, N.; Levine, B.L.; June, C.H.; Hodes, R.J. Regulation of Telomerase RNA Template Expression in Human T Lymphocyte Development and Activation. J. Immunol. 1997, 158, 3215–3220. [Google Scholar] [CrossRef] [PubMed]
- Weng, N.P.; Levine, B.L.; June, C.H.; Hodes, R.J. Regulated Expression of Telomerase Activity in Human T Lymphocyte Development and Activation. J. Exp. Med. 1996, 183, 2471–2479. [Google Scholar] [CrossRef]
- Lu, W.-Y.; Forbes, S.J. Telomerase Activity Links to Regenerative Capacity of Hepatocytes. Transplantation 2018, 102, 1587–1588. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, S.; Sozen, B.; Demir, N. Telomere Length and Telomerase Activity during Oocyte Maturation and Early Embryo Development in Mammalian Species. Mol. Hum. Reprod. 2014, 20, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlrausch, F.B.; Wang, F.; Chamani, I.; Keefe, D.L. Telomere Shortening and Fusions: A Link to Aneuploidy in Early Human Embryo Development. Obstet. Gynecol. Surv. 2021, 76, 429–436. [Google Scholar] [CrossRef]
- Lupatov, A.Y.; Yarygin, K.N. Telomeres and Telomerase in the Control of Stem Cells. Biomedicines 2022, 10, 2335. [Google Scholar] [CrossRef]
- MacKenzie, D.; Watters, A.K.; To, J.T.; Young, M.W.; Muratori, J.; Wilkoff, M.H.; Abraham, R.G.; Plummer, M.M.; Zhang, D. ALT Positivity in Human Cancers: Prevalence and Clinical Insights. Cancers 2021, 13, 2384. [Google Scholar] [CrossRef]
- Lingner, J.; Cech, T.R.; Hughes, T.R.; Lundblad, V. Three Ever Shorter Telomere (EST) Genes Are Dispensable for In Vitro Yeast Telomerase Activity. Proc. Natl. Acad. Sci. USA 1997, 94, 11190–11195. [Google Scholar] [CrossRef] [Green Version]
- Weinrich, S.L.; Pruzan, R.; Ma, L.; Ouellette, M.; Tesmer, V.M.; Holt, S.E.; Bodnar, A.G.; Lichtsteiner, S.; Kim, N.W.; Trager, J.B.; et al. Reconstitution of Human Telomerase with the Template RNA Component HTR and the Catalytic Protein Subunit HTRT. Nat. Genet. 1997, 17, 498–502. [Google Scholar] [CrossRef]
- Beattie, T.L.; Zhou, W.; Robinson, M.O.; Harrington, L. Reconstitution of Human Telomerase Activity In Vitro. Curr. Biol. 1998, 8, 177–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seto, A.G.; Zaug, A.J.; Sobel, S.G.; Wolin, S.L.; Cech, T.R. Saccharomyces Cerevisiae Telomerase Is an Sm Small Nuclear Ribonucleoprotein Particle. Nature 1999, 401, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Vasianovich, Y.; Bajon, E.; Wellinger, R.J. Telomerase Biogenesis Requires a Novel Mex67 Function and a Cytoplasmic Association with the Sm7 Complex. eLife 2020, 9, e60000. [Google Scholar] [CrossRef]
- Chappell, A.S.; Lundblad, V. Structural Elements Required for Association of the Saccharomyces Cerevisiae Telomerase RNA with the Est2 Reverse Transcriptase. Mol. Cell. Biol. 2004, 24, 7720–7736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubin, J.W.; Tucey, T.M.; Lundblad, V. Using Separation-of-Function Mutagenesis to Define the Full Spectrum of Activities Performed by the Est1 Telomerase Subunit In Vivo. Genetics 2018, 208, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Seto, A.G. A Bulged Stem Tethers Est1p to Telomerase RNA in Budding Yeast. Genes Dev. 2002, 16, 2800–2812. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.K.; Lundblad, V. Est1 and Cdc13 as Comediators of Telomerase Access. Science 1999, 286, 117–120. [Google Scholar] [CrossRef]
- Pennock, E.; Buckley, K.; Lundblad, V. Cdc13 Delivers Separate Complexes to the Telomere for End Protection and Replication. Cell 2001, 104, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Tucey, T.M.; Lundblad, V. A Yeast Telomerase Complex Containing the Est1 Recruitment Protein Is Assembled Early in the Cell Cycle. Biochemistry 2013, 52, 1131–1133. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zakian, V.A. The Telomeric Cdc13 Protein Interacts Directly with the Telomerase Subunit Est1 to Bring It to Telomeric DNA Ends In Vitro. Proc. Natl. Acad. Sci. USA 2011, 108, 20362–20369. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-F.; Lu, C.-Y.; Lin, Y.-C.; Yu, T.-Y.; Chang, C.-P.; Li, J.-R.; Li, H.-W.; Lin, J.-J. Modulation of Yeast Telomerase Activity by Cdc13 and Est1 In Vitro. Sci. Rep. 2016, 6, 34104. [Google Scholar] [CrossRef] [Green Version]
- Lebo, K.J.; Niederer, R.O.; Zappulla, D.C. A Second Essential Function of the Est1-Binding Arm of Yeast Telomerase RNA. RNA 2015, 21, 862–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, S.E.; Stellwagen, A.E.; Diede, S.J.; Singer, M.S.; Haimberger, Z.W.; Johnson, C.O.; Tzoneva, M.; Gottschling, D.E. The Function of a Stem-Loop in Telomerase RNA Is Linked to the DNA Repair Protein Ku. Nat. Genet. 2001, 27, 64–67. [Google Scholar] [CrossRef]
- Pfingsten, J.S.; Goodrich, K.J.; Taabazuing, C.; Ouenzar, F.; Chartrand, P.; Cech, T.R. Mutually Exclusive Binding of Telomerase RNA and DNA by Ku Alters Telomerase Recruitment Model. Cell 2012, 148, 922–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xue, J.; Churikov, D.; Hass, E.P.; Shi, S.; Lemon, L.D.; Luciano, P.; Bertuch, A.A.; Zappulla, D.C.; Géli, V.; et al. Structural Insights into Yeast Telomerase Recruitment to Telomeres. Cell 2018, 172, 331–343.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hass, E.P.; Zappulla, D.C. The Ku Subunit of Telomerase Binds Sir4 to Recruit Telomerase to Lengthen Telomeres in S. cerevisiae. eLife 2015, 4, e07750. [Google Scholar] [CrossRef]
- Tucey, T.M.; Lundblad, V. Regulated Assembly and Disassembly of the Yeast Telomerase Quaternary Complex. Genes Dev. 2014, 28, 2077–2089. [Google Scholar] [CrossRef] [Green Version]
- Bianco, P.R. OB-Fold Families of Genome Guardians: A Universal Theme Constructed from the Small β-Barrel Building Block. Front. Mol. Biosci. 2022, 9, 784451. [Google Scholar] [CrossRef]
- Lemieux, B.; Laterreur, N.; Perederina, A.; Noël, J.-F.; Dubois, M.-L.; Krasilnikov, A.S.; Wellinger, R.J. Active Yeast Telomerase Shares Subunits with Ribonucleoproteins RNase P and RNase MRP. Cell 2016, 165, 1171–1181. [Google Scholar] [CrossRef] [Green Version]
- Laterreur, N.; Lemieux, B.; Neumann, H.; Berger-Dancause, J.-C.; Lafontaine, D.; Wellinger, R.J. The Yeast Telomerase Module for Telomere Recruitment Requires a Specific RNA Architecture. RNA 2018, 24, 1067–1079. [Google Scholar] [CrossRef] [Green Version]
- Garcia, P.D.; Leach, R.W.; Wadsworth, G.M.; Choudhary, K.; Li, H.; Aviran, S.; Kim, H.D.; Zakian, V.A. Stability and Nuclear Localization of Yeast Telomerase Depend on Protein Components of RNase P/MRP. Nat. Commun. 2020, 11, 2173. [Google Scholar] [CrossRef] [PubMed]
- Mozdy, A.D.; Cech, T.R. Low Abundance of Telomerase in Yeast: Implications for Telomerase Haploinsufficiency. RNA 2006, 12, 1721–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.-W.; McDonald, K.R.; Guise, A.J.; Chan, A.; Cristea, I.M.; Zakian, V.A. Proteomics of Yeast Telomerase Identified Cdc48-Npl4-Ufd1 and Ufd4 as Regulators of Est1 and Telomere Length. Nat. Commun. 2015, 6, 8290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dionne, I.; Larose, S.; Dandjinou, A.T.; Abou Elela, S.; Wellinger, R.J. Cell Cycle-Dependent Transcription Factors Control the Expression of Yeast Telomerase RNA. RNA 2013, 19, 992–1002. [Google Scholar] [CrossRef] [Green Version]
- Noël, J.-F.; Larose, S.; Abou Elela, S.; Wellinger, R.J. Budding Yeast Telomerase RNA Transcription Termination Is Dictated by the Nrd1/Nab3 Non-Coding RNA Termination Pathway. Nucleic Acids Res. 2012, 40, 5625–5636. [Google Scholar] [CrossRef] [Green Version]
- Arndt, K.M.; Reines, D. Termination of Transcription of Short Noncoding RNAs by RNA Polymerase II. Annu. Rev. Biochem. 2015, 84, 381–404. [Google Scholar] [CrossRef] [Green Version]
- Chapon, C.; Cech, T.R.; Zaug, A.J. Polyadenylation of Telomerase RNA in Budding Yeast. RNA 1997, 3, 1337–1351. [Google Scholar]
- Lemay, J.-F.; Bachand, F. Fail-Safe Transcription Termination: Because One Is Never Enough. RNA Biol. 2015, 12, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Box, J.A.; Bunch, J.T.; Tang, W.; Baumann, P. Spliceosomal Cleavage Generates the 3′ End of Telomerase RNA. Nature 2008, 456, 910–914. [Google Scholar] [CrossRef]
- Qi, X.; Rand, D.P.; Podlevsky, J.D.; Li, Y.; Mosig, A.; Stadler, P.F.; Chen, J.J.-L. Prevalent and Distinct Spliceosomal 3′-End Processing Mechanisms for Fungal Telomerase RNA. Nat. Commun. 2015, 6, 6105. [Google Scholar] [CrossRef] [Green Version]
- Kannan, R.; Helston, R.M.; Dannebaum, R.O.; Baumann, P. Diverse Mechanisms for Spliceosome-Mediated 3′ End Processing of Telomerase RNA. Nat. Commun. 2015, 6, 6104. [Google Scholar] [CrossRef] [Green Version]
- Gunisova, S.; Elboher, E.; Nosek, J.; Gorkovoy, V.; Brown, Y.; Lucier, J.-F.; Laterreur, N.; Wellinger, R.J.; Tzfati, Y.; Tomaska, L. Identification and Comparative Analysis of Telomerase RNAs from Candida Species Reveal Conservation of Functional Elements. RNA 2009, 15, 546–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smekalova, E.M.; Malyavko, A.N.; Zvereva, M.I.; Mardanov, A.V.; Ravin, N.V.; Skryabin, K.G.; Westhof, E.; Dontsova, O.A. Specific Features of Telomerase RNA from Hansenula Polymorpha. RNA 2013, 19, 1563–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Červenák, F.; Juríková, K.; Devillers, H.; Kaffe, B.; Khatib, A.; Bonnell, E.; Sopkovičová, M.; Wellinger, R.J.; Nosek, J.; Tzfati, Y.; et al. Identification of Telomerase RNAs in Species of the Yarrowia Clade Provides Insights into the Co-Evolution of Telomerase, Telomeric Repeats and Telomere-Binding Proteins. Sci. Rep. 2019, 9, 13365. [Google Scholar] [CrossRef] [Green Version]
- Bartle, L.; Vasianovich, Y.; Wellinger, R.J. Maturation and Shuttling of the Yeast Telomerase RNP: Assembling Something New Using Recycled Parts. Curr. Genet. 2022, 68, 3–14. [Google Scholar] [CrossRef]
- Okamura, M.; Inose, H.; Masuda, S. RNA Export through the NPC in Eukaryotes. Genes 2015, 6, 124–149. [Google Scholar] [CrossRef]
- Gallardo, F.; Olivier, C.; Dandjinou, A.T.; Wellinger, R.J.; Chartrand, P. TLC1 RNA Nucleo-Cytoplasmic Trafficking Links Telomerase Biogenesis to Its Recruitment to Telomeres. EMBO J. 2008, 27, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Becker, D.; Krebber, H. Telomerase RNA TLC1 Shuttling to the Cytoplasm Requires MRNA Export Factors and Is Important for Telomere Maintenance. Cell Rep. 2014, 8, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Kannan, R.; Blanchette, M.; Baumann, P. Telomerase RNA Biogenesis Involves Sequential Binding by Sm and Lsm Complexes. Nature 2012, 484, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Collopy, L.C.; Ware, T.L.; Goncalves, T.; í Kongsstovu, S.; Yang, Q.; Amelina, H.; Pinder, C.; Alenazi, A.; Moiseeva, V.; Pearson, S.R.; et al. LARP7 Family Proteins Have Conserved Function in Telomerase Assembly. Nat. Commun. 2018, 9, 557. [Google Scholar] [CrossRef] [Green Version]
- Páez-Moscoso, D.J.; Ho, D.V.; Pan, L.; Hildebrand, K.; Jensen, K.L.; Levy, M.J.; Florens, L.; Baumann, P. A Putative Cap Binding Protein and the Methyl Phosphate Capping Enzyme Bin3/MePCE Function in Telomerase Biogenesis. Nat. Commun. 2022, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Porat, J.; El Baidouri, M.; Grigull, J.; Deragon, J.-M.; Bayfield, M.A. The Methyl Phosphate Capping Enzyme Bmc1/Bin3 Is a Stable Component of the Fission Yeast Telomerase Holoenzyme. Nat. Commun. 2022, 13, 1277. [Google Scholar] [CrossRef] [PubMed]
- Rhind, N.; Chen, Z.; Yassour, M.; Thompson, D.A.; Haas, B.J.; Habib, N.; Wapinski, I.; Roy, S.; Lin, M.F.; Heiman, D.I.; et al. Comparative Functional Genomics of the Fission Yeasts. Science 2011, 332, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Livengood, A.J.; Zaug, A.J.; Cech, T.R. Essential Regions of Saccharomyces Cerevisiae Telomerase RNA: Separate Elements for Est1p and Est2p Interaction. Mol. Cell. Biol. 2002, 22, 2366–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Blackburn, E.H. Nucleolar Protein PinX1p Regulates Telomerase by Sequestering Its Protein Catalytic Subunit in an Inactive Complex Lacking Telomerase RNA. Genes Dev. 2004, 18, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Yen, W.-F.; Chico, L.; Lei, M.; Lue, N.F. Telomerase Regulatory Subunit Est3 in Two Candida Species Physically Interacts with the TEN Domain of TERT and Telomeric DNA. Proc. Natl. Acad. Sci. USA 2011, 108, 20370–20375. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.; Yu, E.Y.; Singh, S.M.; Lue, N.F. Mutual Dependence of Candida Albicans Est1p and Est3p in Telomerase Assembly and Activation. Eukaryot. Cell 2007, 6, 1330–1338. [Google Scholar] [CrossRef] [Green Version]
- Shepelev, N.M.; Mariasina, S.S.; Mantsyzov, A.B.; Malyavko, A.N.; Efimov, S.V.; Petrova, O.A.; Rodina, E.V.; Zvereva, M.I.; Dontsova, O.A.; Polshakov, V.I. Insights into the Structure and Function of Est3 from the Hansenula Polymorpha Telomerase. Sci. Rep. 2020, 10, 11109. [Google Scholar] [CrossRef]
- Hirsch, A.G.; Becker, D.; Lamping, J.-P.; Krebber, H. Unraveling the Stepwise Maturation of the Yeast Telomerase Including a Cse1 and Mtr10 Mediated Quality Control Checkpoint. Sci. Rep. 2021, 11, 22174. [Google Scholar] [CrossRef]
- Dalby, A.B.; Goodrich, K.J.; Pfingsten, J.S.; Cech, T.R. RNA Recognition by the DNA End-Binding Ku Heterodimer. RNA 2013, 19, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Malyavko, A.N.; Petrova, O.A.; Zvereva, M.I.; Polshakov, V.I.; Dontsova, O.A. Telomere Length Regulation by Rif1 Protein from Hansenula Polymorpha. eLife 2022, 11, e75010. [Google Scholar] [CrossRef] [PubMed]
- Ferrezuelo, F.; Steiner, B.; Aldea, M.; Futcher, B. Biogenesis of Yeast Telomerase Depends on the Importin Mtr10. Mol. Cell. Biol. 2002, 22, 6046–6055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Kanoh, J.; Saito, M.; Ishikawa, F. Fission Yeast Pot1-Tpp1 Protects Telomeres and Regulates Telomere Length. Science 2008, 320, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.A.; Nakamura, T.M. Protection and Replication of Telomeres in Fission Yeast. Biochem. Cell Biol. 2009, 87, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.H.; Zakian, V.A. Protein-DNA Interactions in Soluble Telosomes from Saccharomyces cerevisiae. Nucl. Acids Res. 1995, 23, 1454–1460. [Google Scholar] [CrossRef] [Green Version]
- Hardy, C.F.; Sussel, L.; Shore, D. A RAP1-Interacting Protein Involved in Transcriptional Silencing and Telomere Length Regulation. Genes Dev. 1992, 6, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Wotton, D.; Shore, D. A Novel Rap1p-Interacting Factor, Rif2p, Cooperates with Rif1p to Regulate Telomere Length in Saccharomyces Cerevisiae. Genes Dev. 1997, 11, 748–760. [Google Scholar] [CrossRef] [Green Version]
- Moretti, P.; Freeman, K.; Coodly, L.; Shore, D. Evidence That a Complex of SIR Proteins Interacts with the Silencer and Telomere-Binding Protein RAP1. Genes Dev. 1994, 8, 2257–2269. [Google Scholar] [CrossRef] [Green Version]
- Moretti, P.; Shore, D. Multiple Interactions in Sir Protein Recruitment by Rap1p at Silencers and Telomeres in Yeast. Mol. Cell. Biol. 2001, 21, 8082–8094. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Cervantes, R.B.; Mandell, E.K.; Otero, J.H.; Lundblad, V. RPA-like Proteins Mediate Yeast Telomere Function. Nat. Struct. Mol. Biol. 2007, 14, 208–214. [Google Scholar] [CrossRef]
- Bochman, M.L.; Paeschke, K.; Chan, A.; Zakian, V.A. Hrq1, a Homolog of the Human RecQ4 Helicase, Acts Catalytically and Structurally to Promote Genome Integrity. Cell Rep. 2014, 6, 346–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickens, D.G.; Rogers, C.M.; Bochman, M.L. The Saccharomyces Cerevisiae Hrq1 and Pif1 DNA Helicases Synergistically Modulate Telomerase Activity In Vitro. J. Biol. Chem. 2018, 293, 14481–14496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, A.K.; Raney, K.D. A Parallel Quadruplex DNA Is Bound Tightly but Unfolded Slowly by Pif1 Helicase. J. Biol. Chem. 2015, 290, 6482–6494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, L.R.; Phillips, J.A.; Thornton, B.R.; Benanti, J.A.; Onigbanjo, M.T.; Toczyski, D.P.; Zakian, V.A. Sensitivity of Yeast Strains with Long G-Tails to Levels of Telomere-Bound Telomerase. PLoS Genet. 2007, 3, e105. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Zhang, J.; Bochman, M.L.; Zakian, V.A.; Ha, T. Periodic DNA Patrolling Underlies Diverse Functions of Pif1 on R-Loops and G-Rich DNA. eLife 2014, 3, e02190. [Google Scholar] [CrossRef]
- Chan, A.; Boulé, J.-B.; Zakian, V.A. Two Pathways Recruit Telomerase to Saccharomyces Cerevisiae Telomeres. PLoS Genet. 2008, 4, e1000236. [Google Scholar] [CrossRef] [Green Version]
- Stellwagen, A.E.; Haimberger, Z.W.; Veatch, J.R.; Gottschling, D.E. Ku Interacts with Telomerase RNA to Promote Telomere Addition at Native and Broken Chromosome Ends. Genes Dev. 2003, 17, 2384–2395. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.T.; Arneric, M.; Sperisen, P.; Lingner, J. Telomere Length Homeostasis Is Achieved via a Switch between Telomerase- Extendible and -Nonextendible States. Cell 2004, 117, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Marcand, S.; Gilson, E.; Shore, D. A Protein-Counting Mechanism for Telomere Length Regulation in Yeast. Science 1997, 275, 986–990. [Google Scholar] [CrossRef]
- Levy, D.L.; Blackburn, E.H. Counting of Rif1p and Rif2p on Saccharomyces Cerevisiae Telomeres Regulates Telomere Length. Mol. Cell. Biol. 2004, 24, 10857–10867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellinger, R.J.; Wolf, A.J.; Zakian, V.A. Saccharomyces Telomeres Acquire Single-Strand TG1–3 Tails Late in S Phase. Cell 1993, 72, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Dionne, I.; Wellinger, R.J. Processing of Telomeric DNA Ends Requires the Passage of a Replication Fork. Nucleic Acids Res. 1998, 26, 5365–5371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrivée, M.; LeBel, C.; Wellinger, R.J. The Generation of Proper Constitutive G-Tails on Yeast Telomeres Is Dependent on the MRX Complex. Genes Dev. 2004, 18, 1391–1396. [Google Scholar] [CrossRef] [Green Version]
- Bonetti, D.; Martina, M.; Clerici, M.; Lucchini, G.; Longhese, M.P. Multiple Pathways Regulate 3′ Overhang Generation at S. cerevisiae Telomeres. Mol. Cell 2009, 35, 70–81. [Google Scholar] [CrossRef]
- Lustig, A.J. Towards the Mechanism of Yeast Telomere Dynamics. Trends Cell Biol. 2019, 29, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.; Shore, D. Increased Association of Telomerase with Short Telomeres in Yeast. Genes Dev. 2007, 21, 1726–1730. [Google Scholar] [CrossRef] [Green Version]
- Sabourin, M.; Tuzon, C.T.; Zakian, V.A. Telomerase and Tel1p Preferentially Associate with Short Telomeres in S. cerevisiae. Mol. Cell 2007, 27, 550–561. [Google Scholar] [CrossRef] [Green Version]
- Martina, M.; Clerici, M.; Baldo, V.; Bonetti, D.; Lucchini, G.; Longhese, M.P. A Balance between Tel1 and Rif2 Activities Regulates Nucleolytic Processing and Elongation at Telomeres. Mol. Cell. Biol. 2012, 32, 1604–1617. [Google Scholar] [CrossRef] [Green Version]
- Bonetti, D.; Clerici, M.; Anbalagan, S.; Martina, M.; Lucchini, G.; Longhese, M.P. Shelterin-like Proteins and Yku Inhibit Nucleolytic Processing of Saccharomyces Cerevisiae Telomeres. PLoS Genet. 2010, 6, e1000966. [Google Scholar] [CrossRef] [Green Version]
- Bonetti, D.; Clerici, M.; Manfrini, N.; Lucchini, G.; Longhese, M.P. The MRX Complex Plays Multiple Functions in Resection of Yku- and Rif2-Protected DNA Ends. PLoS ONE 2010, 5, e14142. [Google Scholar] [CrossRef]
- Tseng, S.-F.; Lin, J.-J.; Teng, S.-C. The Telomerase-Recruitment Domain of the Telomere Binding Protein Cdc13 Is Regulated by Mec1p/Tel1p-Dependent Phosphorylation. Nucleic Acids Res. 2006, 34, 6327–6336. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.-J.; Hsu, P.-H.; Su, Y.-T.; Yang, C.-W.; Kao, L.; Tseng, S.-F.; Tsai, M.-D.; Teng, S.-C. PP2A and Aurora Differentially Modify Cdc13 to Promote Telomerase Release from Telomeres at G2/M Phase. Nat. Commun. 2014, 5, 5312. [Google Scholar] [CrossRef] [Green Version]
- Moser, B.A.; Subramanian, L.; Khair, L.; Chang, Y.-T.; Nakamura, T.M. Fission Yeast Tel1ATM and Rad3ATR Promote Telomere Protection and Telomerase Recruitment. PLoS Genet. 2009, 5, e1000622. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, A.; Bianchi, A.; Lemmens, L.; Damay, P.; Shore, D. Distinct Roles for Yeast Stn1 in Telomere Capping and Telomerase Inhibition. EMBO J. 2008, 27, 2328–2339. [Google Scholar] [CrossRef] [Green Version]
- Zalzman, M.; Meltzer, W.A.; Portney, B.A.; Brown, R.A.; Gupta, A. The Role of Ubiquitination and SUMOylation in Telomere Biology. Curr. Issues Mol. Biol. 2020, 35, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Taggart, A.K.P.; Teng, S.-C.; Zakian, V.A. Est1p As a Cell Cycle-Regulated Activator of Telomere-Bound Telomerase. Science 2002, 297, 1023–1026. [Google Scholar] [CrossRef]
- Osterhage, J.L.; Talley, J.M.; Friedman, K.L. Proteasome-Dependent Degradation of Est1p Regulates the Cell Cycle–Restricted Assembly of Telomerase in Saccharomyces Cerevisiae. Nat. Struct. Mol. Biol. 2006, 13, 720–728. [Google Scholar] [CrossRef]
- Ferguson, J.L.; Chao, W.C.H.; Lee, E.; Friedman, K.L. The Anaphase Promoting Complex Contributes to the Degradation of the S. cerevisiae Telomerase Recruitment Subunit Est1p. PLoS ONE 2013, 8, e55055. [Google Scholar] [CrossRef]
- Tuzon, C.T.; Wu, Y.; Chan, A.; Zakian, V.A. The Saccharomyces Cerevisiae Telomerase Subunit Est3 Binds Telomeres in a Cell Cycle– and Est1–Dependent Manner and Interacts Directly with Est1 In Vitro. PLoS Genet. 2011, 7, e1002060. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Blobel, G. A SUMO Ligase Is Part of a Nuclear Multiprotein Complex That Affects DNA Repair and Chromosomal Organization. Proc. Natl. Acad. Sci. USA 2005, 102, 4777–4782. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.C.; Luke, B.; Schober, H.; Kalck, V.; Lingner, J.; Gasser, S.M. The PIAS Homologue Siz2 Regulates Perinuclear Telomere Position and Telomerase Activity in Budding Yeast. Nat. Cell Biol. 2011, 13, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Hang, L.E.; Lopez, C.R.; Liu, X.; Williams, J.M.; Chung, I.; Wei, L.; Bertuch, A.A.; Zhao, X. Regulation of Ku-DNA Association by Yku70 C-Terminal Tail and SUMO Modification. J. Biol. Chem. 2014, 289, 10308–10317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, L.E.; Liu, X.; Cheung, I.; Yang, Y.; Zhao, X. SUMOylation Regulates Telomere Length Homeostasis by Targeting Cdc13. Nat. Struct. Mol. Biol. 2011, 18, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Carballar, R.; Martínez-Láinez, J.M.; Samper, B.; Bru, S.; Bállega, E.; Mirallas, O.; Ricco, N.; Clotet, J.; Jiménez, J. CDK-Mediated Yku80 Phosphorylation Regulates the Balance Between Non-Homologous End Joining (NHEJ) and Homologous Directed Recombination (HDR). J. Mol. Biol. 2020, 432, 166715. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, P.; Wagner, S.A.; Weinert, B.T.; Sharma, S.; Bacinskaja, G.; Rehman, M.; Juffer, A.H.; Walther, T.C.; Lisby, M.; Choudhary, C. Proteome-Wide Analysis of Lysine Acetylation Suggests Its Broad Regulatory Scope in Saccharomyces Cerevisiae. Mol. Cell. Proteom. 2012, 11, 1510–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villén, J. Global Analysis of Phosphorylation and Ubiquitylation Cross-Talk in Protein Degradation. Nat. Methods 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Lanz, M.C.; Yugandhar, K.; Gupta, S.; Sanford, E.J.; Faça, V.M.; Vega, S.; Joiner, A.M.N.; Fromme, J.C.; Yu, H.; Smolka, M.B. In-Depth and 3-Dimensional Exploration of the Budding Yeast Phosphoproteome. EMBO Rep. 2021, 22, e51121. [Google Scholar] [CrossRef]
- Bertuch, A.A. The Molecular Genetics of the Telomere Biology Disorders. RNA Biol. 2016, 13, 696–706. [Google Scholar] [CrossRef] [Green Version]
- Heiss, N.S.; Knight, S.W.; Vulliamy, T.J.; Klauck, S.M.; Wiemann, S.; Mason, P.J.; Poustka, A.; Dokal, I. X-Linked Dyskeratosis Congenita Is Caused by Mutations in a Highly Conserved Gene with Putative Nucleolar Functions. Nat. Genet. 1998, 19, 32–38. [Google Scholar] [CrossRef]
- Armanios, M.; Chen, J.-L.; Chang, Y.-P.C.; Brodsky, R.A.; Hawkins, A.; Griffin, C.A.; Eshleman, J.R.; Cohen, A.R.; Chakravarti, A.; Hamosh, A.; et al. Haploinsufficiency of Telomerase Reverse Transcriptase Leads to Anticipation in Autosomal Dominant Dyskeratosis Congenita. Proc. Natl. Acad. Sci. USA 2005, 102, 15960–15964. [Google Scholar] [CrossRef] [Green Version]
- Vulliamy, T.; Marrone, A.; Goldman, F.; Dearlove, A.; Bessler, M.; Mason, P.J.; Dokal, I. The RNA Component of Telomerase Is Mutated in Autosomal Dominant Dyskeratosis Congenita. Nature 2001, 413, 432–435. [Google Scholar] [CrossRef]
- Savage, S.A.; Giri, N.; Baerlocher, G.M.; Orr, N.; Lansdorp, P.M.; Alter, B.P. TINF2, a Component of the Shelterin Telomere Protection Complex, Is Mutated in Dyskeratosis Congenita. Am. J. Hum. Genet. 2008, 82, 501–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, E.D.; Collins, K. Specificity and Stoichiometry of Subunit Interactions in the Human Telomerase Holoenzyme Assembled In Vivo. Mol. Cell. Biol. 2010, 30, 2775–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.H.D. Structural Biology of Human Telomerase: Progress and Prospects. Biochem. Soc. Trans. 2021, 49, 1927–1939. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-L.; Blasco, M.A.; Greider, C.W. Secondary Structure of Vertebrate Telomerase RNA. Cell 2000, 100, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autexier, C.; Lue, N.F. The Structure and Function of Telomerase Reverse Transcriptase. Annu. Rev. Biochem. 2006, 75, 493–517. [Google Scholar] [CrossRef]
- Wu, R.A.; Dagdas, Y.S.; Yilmaz, S.T.; Yildiz, A.; Collins, K. Single-Molecule Imaging of Telomerase Reverse Transcriptase in Human Telomerase Holoenzyme and Minimal RNP Complexes. eLife 2015, 4, e08363. [Google Scholar] [CrossRef]
- Feng, J.; Funk, W.D.; Wang, S.-S.; Weinrich, S.L.; Avilion, A.A.; Chiu, C.-P.; Adams, R.R.; Chang, E.; Allsopp, R.C.; Yu, J.; et al. The RNA Component of Human Telomerase. Science 1995, 269, 1236–1241. [Google Scholar] [CrossRef]
- Mitchell, J.R.; Cheng, J.; Collins, K. A Box H/ACA Small Nucleolar RNA-Like Domain at the Human Telomerase RNA 3′ End. Mol. Cell. Biol. 1999, 19, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Hinkley, C.S.; Blasco, M.A.; Funk, W.D.; Feng, J.; Villeponteau, B.; Greider, C.W.; Herr, W. The Mouse Telomerase RNA 5′-End Lies Just Upstream of the Telomerase Template Sequence. Nucleic Acids Res. 1998, 26, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Roake, C.M.; Galati, A.; Bavasso, F.; Micheli, E.; Saggio, I.; Schoeftner, S.; Cacchione, S.; Gatti, M.; Artandi, S.E.; et al. Loss of Human TGS1 Hypermethylase Promotes Increased Telomerase RNA and Telomere Elongation. Cell Rep. 2020, 30, 1358–1372.e5. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, K.C.; Cech, T.R. 3′ Terminal Diversity of MRP RNA and Other Human Noncoding RNAs Revealed by Deep Sequencing. BMC Mol. Biol. 2013, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.-K.; Wang, H.-F.; Burns, A.M.; Schroeder, M.R.; Gaspari, M.; Baumann, P. Human Telomerase RNA Processing and Quality Control. Cell Rep. 2015, 13, 2232–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theimer, C.A.; Jády, B.E.; Chim, N.; Richard, P.; Breece, K.E.; Kiss, T.; Feigon, J. Structural and Functional Characterization of Human Telomerase RNA Processing and Cajal Body Localization Signals. Mol. Cell 2007, 27, 869–881. [Google Scholar] [CrossRef]
- Pakhomova, T.; Moshareva, M.; Vasilkova, D.; Zatsepin, T.; Dontsova, O.; Rubtsova, M. Role of RNA Biogenesis Factors in the Processing and Transport of Human Telomerase RNA. Biomedicines 2022, 10, 1275. [Google Scholar] [CrossRef]
- Rubtsova, M.; Naraykina, Y.; Vasilkova, D.; Meerson, M.; Zvereva, M.; Prassolov, V.; Lazarev, V.; Manuvera, V.; Kovalchuk, S.; Anikanov, N.; et al. Protein Encoded in Human Telomerase RNA Is Involved in Cell Protective Pathways. Nucleic Acids Res. 2018, 46, 8966–8977. [Google Scholar] [CrossRef] [Green Version]
- Shliapina, V.; Koriagina, M.; Vasilkova, D.; Govorun, V.; Dontsova, O.; Rubtsova, M. Human Telomerase RNA Protein Encoded by Telomerase RNA Is Involved in Metabolic Responses. Front. Cell Dev. Biol. 2021, 9, 754611. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, P.; Zheng, Q.; Gao, G.; Yuan, J.; Wang, P.; Huang, J.; Xie, L.; Lu, X.; Tong, T.; et al. Mitochondrial Trafficking and Processing of Telomerase RNA TERC. Cell Rep. 2018, 24, 2589–2595. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Liu, P.; Gao, G.; Yuan, J.; Wang, P.; Huang, J.; Xie, L.; Lu, X.; Di, F.; Tong, T.; et al. Mitochondrion-Processed TERC Regulates Senescence without Affecting Telomerase Activities. Protein Cell 2019, 10, 631–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sexton, A.N.; Collins, K. The 5′ Guanosine Tracts of Human Telomerase RNA Are Recognized by the G-Quadruplex Binding Domain of the RNA Helicase DHX36 and Function to Increase RNA Accumulation. Mol. Cell. Biol. 2011, 31, 736–743. [Google Scholar] [CrossRef] [Green Version]
- Lattmann, S.; Stadler, M.B.; Vaughn, J.P.; Akman, S.A.; Nagamine, Y. The DEAH-Box RNA Helicase RHAU Binds an Intramolecular RNA G-quadruplex in TERC and Associates with Telomerase Holoenzyme. Nucleic Acids Res. 2011, 39, 9390–9404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booy, E.P.; Meier, M.; Okun, N.; Novakowski, S.K.; Xiong, S.; Stetefeld, J.; McKenna, S.A. The RNA Helicase RHAU (DHX36) Unwinds a G4-Quadruplex in Human Telomerase RNA and Promotes the Formation of the P1 Helix Template Boundary. Nucleic Acids Res. 2012, 40, 4110–4124. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Liu, L.; Lazarev, D.; Al-Zain, A.; Fomin, V.; Yeung, P.L.; Chambers, S.M.; Lu, C.-W.; Studer, L.; Manley, J.L. TCF3 Alternative Splicing Controlled by HnRNP H/F Regulates E-Cadherin Expression and HESC Pluripotency. Genes Dev. 2018, 32, 1161–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyson-Capper, A.; Gautrey, H. Regulation of Mcl-1 Alternative Splicing by HnRNP F, H1 and K in Breast Cancer Cells. RNA Biol. 2018, 15, 1448–1457. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Xie, N.; Su, Y.; Sun, Z.; Liang, Y.; Zhang, N.; Liu, D.; Jia, S.; Xing, X.; Han, L.; et al. HnRNP F/H Associate with HTERC and Telomerase Holoenzyme to Modulate Telomerase Function and Promote Cell Proliferation. Cell Death Differ 2020, 27, 1998–2013. [Google Scholar] [CrossRef]
- Allen, B.L.; Taatjes, D.J. The Mediator Complex: A Central Integrator of Transcription. Nat. Rev. Mol. Cell. Biol. 2015, 16, 155–166. [Google Scholar] [CrossRef]
- Kirstein, N.; Gomes Dos Santos, H.; Blumenthal, E.; Shiekhattar, R. The Integrator Complex at the Crossroad of Coding and Noncoding RNA. Curr. Opin. Cell Biol. 2021, 70, 37–43. [Google Scholar] [CrossRef]
- Rubtsova, M.P.; Vasilkova, D.P.; Moshareva, M.A.; Malyavko, A.N.; Meerson, M.B.; Zatsepin, T.S.; Naraykina, Y.V.; Beletsky, A.V.; Ravin, N.V.; Dontsova, O.A. Integrator Is a Key Component of Human Telomerase RNA Biogenesis. Sci. Rep. 2019, 9, 1701. [Google Scholar] [CrossRef] [Green Version]
- Rubtsova, M.; Dontsova, O. How Structural Features Define Biogenesis and Function of Human Telomerase RNA Primary Transcript. Biomedicines 2022, 10, 1650. [Google Scholar] [CrossRef]
- Yi, X.; Tesmer, V.M.; Savre-Train, I.; Shay, J.W.; Wright, W.E. Both Transcriptional and Posttranscriptional Mechanisms Regulate Human Telomerase Template RNA Levels. Mol. Cell. Biol. 1999, 19, 3989–3997. [Google Scholar] [CrossRef] [Green Version]
- Cioce, M.; Lamond, A.I. Cajal Bodies: A Long History of Discovery. Annu. Rev. Cell Dev. Biol. 2005, 21, 105–131. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-L. A Critical Stem-Loop Structure in the CR4-CR5 Domain of Mammalian Telomerase RNA. Nucleic Acids Res. 2002, 30, 592–597. [Google Scholar] [CrossRef]
- Egan, E.D.; Collins, K. An Enhanced H/ACA RNP Assembly Mechanism for Human Telomerase RNA. Mol. Cell. Biol. 2012, 32, 2428–2439. [Google Scholar] [CrossRef] [Green Version]
- Richard, P.; Darzacq, X.; Bertrand, E.; Jády, B.E.; Verheggen, C.; Kiss, T. A Common Sequence Motif Determines the Cajal Body-Specific Localization of Box H/ACA ScaRNAs. EMBO J. 2003, 22, 4283–4293. [Google Scholar] [CrossRef]
- Tollervey, D.; Kiss, T. Function and Synthesis of Small Nucleolar RNAs. Curr. Opin. Cell Biol. 1997, 9, 337–342. [Google Scholar] [CrossRef]
- Hargrove, B.W.; Bhattacharyya, A.; Domitrovich, A.M.; Kapler, G.M.; Kirk, K.; Shippen, D.E.; Kunkel, G.R. Identification of an Essential Proximal Sequence Element in the Promoter of the Telomerase RNA Gene of Tetrahymena Thermophila. Nucleic Acids Res. 1999, 27, 4269–4275. [Google Scholar] [CrossRef] [Green Version]
- Kiss, T. Biogenesis of Small Nuclear RNPs. J. Cell Sci. 2004, 117, 5949–5951. [Google Scholar] [CrossRef] [Green Version]
- Kiss, T.; Fayet, E.; Jady, B.E.; Richard, P.; Weber, M. Biogenesis and Intranuclear Trafficking of Human Box C/D and H/ACA RNPs. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Pogacic, V.; Dragon, F.; Filipowicz, W. Human H/ACA Small Nucleolar RNPs and Telomerase Share Evolutionarily Conserved Proteins NHP2 and NOP10. Mol. Cell Biol. 2000, 20, 9028–9040. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Collins, K. Purification of Human Telomerase Complexes Identifies Factors Involved in Telomerase Biogenesis and Telomere Length Regulation. Mol. Cell 2007, 28, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Hoareau-Aveilla, C.; Bonoli, M.; Caizergues-Ferrer, M.; Henry, Y. HNaf1 Is Required for Accumulation of Human Box H/ACA SnoRNPs, ScaRNPs, and Telomerase. RNA 2006, 12, 832–840. [Google Scholar] [CrossRef] [Green Version]
- Machado-Pinilla, R.; Liger, D.; Leulliot, N.; Meier, U.T. Mechanism of the AAA+ ATPases Pontin and Reptin in the Biogenesis of H/ACA RNPs. RNA 2012, 18, 1833–1845. [Google Scholar] [CrossRef] [Green Version]
- Grozdanov, P.N.; Roy, S.; Kittur, N.; Meier, U.T. SHQ1 Is Required Prior to NAF1 for Assembly of H/ACA Small Nucleolar and Telomerase RNPs. RNA 2009, 15, 1188–1197. [Google Scholar] [CrossRef] [Green Version]
- Walbott, H.; Machado-Pinilla, R.; Liger, D.; Blaud, M.; Réty, S.; Grozdanov, P.N.; Godin, K.; van Tilbeurgh, H.; Varani, G.; Meier, U.T.; et al. The H/ACA RNP Assembly Factor SHQ1 Functions as an RNA Mimic. Genes Dev. 2011, 25, 2398–2408. [Google Scholar] [CrossRef] [Green Version]
- Boulon, S.; Marmier-Gourrier, N.; Pradet-Balade, B.; Wurth, L.; Verheggen, C.; Jády, B.E.; Rothé, B.; Pescia, C.; Robert, M.-C.; Kiss, T.; et al. The Hsp90 Chaperone Controls the Biogenesis of L7Ae RNPs through Conserved Machinery. J. Cell Biol. 2008, 180, 579–595. [Google Scholar] [CrossRef]
- Venteicher, A.S.; Meng, Z.; Mason, P.J.; Veenstra, T.D.; Artandi, S.E. Identification of ATPases Pontin and Reptin as Telomerase Components Essential for Holoenzyme Assembly. Cell 2008, 132, 945–957. [Google Scholar] [CrossRef] [Green Version]
- Darzacq, X.; Kittur, N.; Roy, S.; Shav-Tal, Y.; Singer, R.H.; Meier, U.T. Stepwise RNP Assembly at the Site of H/ACA RNA Transcription in Human Cells. J. Cell Biol. 2006, 173, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Ashbridge, B.; Orte, A.; Yeoman, J.A.; Kirwan, M.; Vulliamy, T.; Dokal, I.; Klenerman, D.; Balasubramanian, S. Single-Molecule Analysis of the Human Telomerase RNA·Dyskerin Interaction and the Effect of Dyskeratosis Congenita Mutations. Biochemistry 2009, 48, 10858–10865. [Google Scholar] [CrossRef] [Green Version]
- Leulliot, N.; Godin, K.S.; Hoareau-Aveilla, C.; Quevillon-Cheruel, S.; Varani, G.; Henry, Y.; Van Tilbeurgh, H. The Box H/ACA RNP Assembly Factor Naf1p Contains a Domain Homologous to Gar1p Mediating Its Interaction with Cbf5p. J. Mol. Biol. 2007, 371, 1338–1353. [Google Scholar] [CrossRef]
- Fatica, A.; Dlakić, M.; Tollervey, D. Naf1p Is a Box H/ACA SnoRNP Assembly Factor. RNA 2002, 8, 1502–1514. [Google Scholar] [CrossRef]
- Zeng, X.-L.; Thumati, N.R.; Fleisig, H.B.; Hukezalie, K.R.; Savage, S.A.; Giri, N.; Alter, B.P.; Wong, J.M.Y. The Accumulation and Not the Specific Activity of Telomerase Ribonucleoprotein Determines Telomere Maintenance Deficiency in X-Linked Dyskeratosis Congenita. Hum. Mol. Genet. 2012, 21, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, T.; Beswick, R.; Kirwan, M.; Marrone, A.; Digweed, M.; Walne, A.; Dokal, I. Mutations in the Telomerase Component NHP2 Cause the Premature Ageing Syndrome Dyskeratosis Congenita. Proc. Natl. Acad. Sci. USA 2008, 105, 8073–8078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walne, A.J.; Vulliamy, T.; Marrone, A.; Beswick, R.; Kirwan, M.; Masunari, Y.; Al-Qurashi, F.-H.; Aljurf, M.; Dokal, I. Genetic Heterogeneity in Autosomal Recessive Dyskeratosis Congenita with One Subtype Due to Mutations in the Telomerase-Associated Protein NOP10. Hum. Mol. Genet. 2007, 16, 1619–1629. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.E.; Gable, D.L.; Wagner, C.L.; Carlile, T.M.; Hanumanthu, V.S.; Podlevsky, J.D.; Khalil, S.E.; DeZern, A.E.; Rojas-Duran, M.F.; Applegate, C.D.; et al. Loss-of-Function Mutations in the RNA Biogenesis Factor NAF1 Predispose to Pulmonary Fibrosis-Emphysema. Sci. Transl. Med. 2016, 8, 351ra107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thumati, N.R.; Zeng, X.-L.; Au, H.H.T.; Jang, C.J.; Jan, E.; Wong, J.M.Y. Severity of X-Linked Dyskeratosis Congenita (DKCX) Cellular Defects Is Not Directly Related to Dyskerin (DKC1) Activity in Ribosomal RNA Biogenesis or MRNA Translation. Hum. Mutat. 2013, 34, 1698–1707. [Google Scholar] [CrossRef]
- Savelyev, N.V.; Shepelev, N.M.; Lavrik, O.I.; Rubtsova, M.P.; Dontsova, O.A. PARP1 Regulates the Biogenesis and Activity of Telomerase Complex Through Modification of H/ACA-Proteins. Front. Cell Dev. Biol. 2021, 9, 621134. [Google Scholar] [CrossRef]
- Becker, J.; Barysch, S.V.; Karaca, S.; Dittner, C.; Hsiao, H.-H.; Diaz, M.B.; Herzig, S.; Urlaub, H.; Melchior, F. Detecting Endogenous SUMO Targets in Mammalian Cells and Tissues. Nat. Struct. Mol. Biol. 2013, 20, 525–531. [Google Scholar] [CrossRef]
- Hendriks, I.A.; D’Souza, R.C.J.; Yang, B.; Verlaan-de Vries, M.; Mann, M.; Vertegaal, A.C.O. Uncovering Global SUMOylation Signaling Networks in a Site-Specific Manner. Nat. Struct. Mol. Biol. 2014, 21, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Impens, F.; Radoshevich, L.; Cossart, P.; Ribet, D. Mapping of SUMO Sites and Analysis of SUMOylation Changes Induced by External Stimuli. Proc. Natl. Acad. Sci. USA 2014, 111, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Chang, J.-G.; Hendriks, I.A.; Sigurðsson, J.O.; Olsen, J.V.; Vertegaal, A.C.O. System-Wide Analysis of SUMOylation Dynamics in Response to Replication Stress Reveals Novel Small Ubiquitin-like Modified Target Proteins and Acceptor Lysines Relevant for Genome Stability. Mol. Cell. Proteom. 2015, 14, 1419–1434. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, I.A.; Lyon, D.; Young, C.; Jensen, L.J.; Vertegaal, A.C.O.; Nielsen, M.L. Site-Specific Mapping of the Human SUMO Proteome Reveals Co-Modification with Phosphorylation. Nat. Struct. Mol. Biol. 2017, 24, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Hochstrasser, M. Origin and Function of Ubiquitin-like Proteins. Nature 2009, 458, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacNeil, D.E.; Lambert-Lanteigne, P.; Qin, J.; McManus, F.P.; Bonneil, E.; Thibault, P.; Autexier, C. SUMOylation- and GAR1-Dependent Regulation of Dyskerin Nuclear and Subnuclear Localization. Mol. Cell. Biol. 2021, 41, e00464-20. [Google Scholar] [CrossRef]
- Al-Masraf, B.S.; Perez, G.I.; Adams-Boone, K.; Cohen, S.B.; Han, L.; Yu, K.; Schmidt, J.C. TCAB1 Prevents Nucleolar Accumulation of the Telomerase RNA to Promote Telomerase Assembly. Biorxiv 2021. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.M.; Nam, H.J.; Choi, H.J.; Yang, J.W.; Lee, J.S.; Kim, M.H.; Kim, S.-I.; Chung, C.H.; Kim, K.I.; et al. SUMOylation of Pontin Chromatin-Remodeling Complex Reveals a Signal Integration Code in Prostate Cancer Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 20793–20798. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Choi, H.J.; Kim, B.; Kim, M.H.; Lee, J.M.; Kim, I.S.; Lee, M.H.; Choi, S.J.; Kim, K.I.; Kim, S.-I.; et al. Roles of Sumoylation of a Reptin Chromatin-Remodelling Complex in Cancer Metastasis. Nat. Cell Biol. 2006, 8, 631–639. [Google Scholar] [CrossRef]
- Jobert, L.; Skjeldam, H.K.; Dalhus, B.; Galashevskaya, A.; Vågbø, C.B.; Bjørås, M.; Nilsen, H. The Human Base Excision Repair Enzyme SMUG1 Directly Interacts with DKC1 and Contributes to RNA Quality Control. Mol. Cell 2013, 49, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Kroustallaki, P.; Lirussi, L.; Carracedo, S.; You, P.; Esbensen, Q.Y.; Götz, A.; Jobert, L.; Alsøe, L.; Sætrom, P.; Gagos, S.; et al. SMUG1 Promotes Telomere Maintenance through Telomerase RNA Processing. Cell Rep. 2019, 28, 1690–1702.e10. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.V.; Vermeulen, M.; Santamaria, A.; Kumar, C.; Miller, M.L.; Jensen, L.J.; Gnad, F.; Cox, J.; Jensen, T.S.; Nigg, E.A.; et al. Quantitative Phosphoproteomics Reveals Widespread Full Phosphorylation Site Occupancy during Mitosis. Sci. Signal. 2010, 3, ra3. [Google Scholar] [CrossRef] [PubMed]
- Hebert, M.D.; Poole, A.R. Towards an Understanding of Regulating Cajal Body Activity by Protein Modification. RNA Biol. 2017, 14, 761–778. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Tomlinson, R.L.; Lukowiak, A.A.; Terns, R.M.; Terns, M.P. Telomerase RNA Accumulates in Cajal Bodies in Human Cancer Cells. MBoC 2004, 15, 81–90. [Google Scholar] [CrossRef]
- Jády, B.E.; Bertrand, E.; Kiss, T. Human Telomerase RNA and Box H/ACA ScaRNAs Share a Common Cajal Body-Specific Localization Signal. J. Cell Biol. 2004, 164, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Venteicher, A.S.; Abreu, E.B.; Meng, Z.; McCann, K.E.; Terns, R.M.; Veenstra, T.D.; Terns, M.P.; Artandi, S.E. A Human Telomerase Holoenzyme Protein Required for Cajal Body Localization and Telomere Synthesis. Science 2009, 323, 644–648. [Google Scholar] [CrossRef] [Green Version]
- Zhong, F.; Savage, S.A.; Shkreli, M.; Giri, N.; Jessop, L.; Myers, T.; Chen, R.; Alter, B.P.; Artandi, S.E. Disruption of Telomerase Trafficking by TCAB1 Mutation Causes Dyskeratosis Congenita. Genes Dev. 2011, 25, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Vogan, J.M.; Zhang, X.; Youmans, D.T.; Regalado, S.G.; Johnson, J.Z.; Hockemeyer, D.; Collins, K. Minimized Human Telomerase Maintains Telomeres and Resolves Endogenous Roles of H/ACA Proteins, TCAB1, and Cajal Bodies. eLife 2016, 5, e18221. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Roake, C.M.; Freund, A.; Batista, P.J.; Tian, S.; Yin, Y.A.; Gajera, C.R.; Lin, S.; Lee, B.; Pech, M.F.; et al. An Activity Switch in Human Telomerase Based on RNA Conformation and Shaped by TCAB1. Cell 2018, 174, 218–230.e13. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, G.E.; Fountain, A.J.; van Roon, A.-M.M.; Rangan, R.; Das, R.; Collins, K.; Nguyen, T.H.D. Structure of Human Telomerase Holoenzyme with Bound Telomeric DNA. Nature 2021, 593, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Zhong, F.L.; Venteicher, A.S.; Meng, Z.; Veenstra, T.D.; Frydman, J.; Artandi, S.E. Proteostatic Control of Telomerase Function through TRiC-Mediated Folding of TCAB1. Cell 2014, 159, 1389–1403. [Google Scholar] [CrossRef] [Green Version]
- Vogan, J.M.; Collins, K. Dynamics of Human Telomerase Holoenzyme Assembly and Subunit Exchange across the Cell Cycle. J. Biol. Chem. 2015, 290, 21320–21335. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.D.; Tam, J.; Wu, R.A.; Greber, B.J.; Toso, D.; Nogales, E.; Collins, K. Cryo-EM Structure of Substrate-Bound Human Telomerase Holoenzyme. Nature 2018, 557, 190–195. [Google Scholar] [CrossRef]
- Keppler, B.R.; Grady, A.T.; Jarstfer, M.B. The Biochemical Role of the Heat Shock Protein 90 Chaperone Complex in Establishing Human Telomerase Activity. J. Biol. Chem. 2006, 281, 19840–19848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toogun, O.A.; DeZwaan, D.C.; Freeman, B.C. The Hsp90 Molecular Chaperone Modulates Multiple Telomerase Activities. Mol. Cell. Biol. 2008, 28, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.E. Hsp90: Structure and Function. Top. Curr. Chem. 2013, 328, 155–240. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; An, S.; Lee, H.-C.; Jin, H.-O.; Seo, S.-K.; Yoo, D.-H.; Lee, K.-H.; Rhee, C.H.; Choi, E.-J.; Hong, S.-I.; et al. A Truncated Form of P23 Down-Regulates Telomerase Activity via Disruption of Hsp90 Function. J. Biol. Chem. 2009, 284, 30871–30880. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Chung, I.K. Curcumin Inhibits Nuclear Localization of Telomerase by Dissociating the Hsp90 Co-Chaperone P23 from HTERT. Cancer Lett. 2010, 290, 76–86. [Google Scholar] [CrossRef]
- Holt, S.E.; Aisner, D.L.; Baur, J.; Tesmer, V.M.; Dy, M.; Ouellette, M.; Trager, J.B.; Morin, G.B.; Toft, D.O.; Shay, J.W.; et al. Functional Requirement of P23 and Hsp90 in Telomerase Complexes. Genes Dev. 1999, 13, 817–826. [Google Scholar] [CrossRef]
- Her, J.; Chung, I.K. The AAA-ATPase NVL2 Is a Telomerase Component Essential for Holoenzyme Assembly. Biochem. Biophys. Res. Commun. 2012, 417, 1086–1092. [Google Scholar] [CrossRef]
- Bachand, F.; Boisvert, F.-M.; Côté, J.; Richard, S.; Autexier, C. The Product of the Survival of Motor Neuron (SMN) Gene Is a Human Telomerase-Associated Protein. MBoC 2002, 13, 3192–3202. [Google Scholar] [CrossRef] [Green Version]
- Pellizzoni, L.; Baccon, J.; Charroux, B.; Dreyfuss, G. The Survival of Motor Neurons (SMN) Protein Interacts with the SnoRNP Proteins Fibrillarin and GAR1. Curr. Biol. 2001, 11, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, S.E.; Jones, K.W.; Zhang, X.; Cheng, X.; Terns, R.M.; Terns, M.P. Determinants of the Interaction of the Spinal Muscular Atrophy Disease Protein SMN with the Dimethylarginine-Modified Box H/ACA Small Nucleolar Ribonucleoprotein GAR1. J. Biol. Chem. 2002, 277, 48087–48093. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Benhattar, J.; Seelentag, W.; Stehle, J.-C.; Bosman, F.T. Immunohistochemical Localization of HTERT Protein in Human Tissues. Histochem. Cell Biol. 2004, 121, 391–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Jin, R.; Zhang, B.; Yang, P.X.; Chen, H.; Bai, Y.X.; Xie, Y.; Huang, C.; Huang, J. Characterization of a Novel Effect of HPinX1 on HTERT Nucleolar Localization. Biochem. Biophys. Res. Commun. 2007, 353, 946–952. [Google Scholar] [CrossRef]
- Wong, J.M.Y.; Kusdra, L.; Collins, K. Subnuclear Shuttling of Human Telomerase Induced by Transformation and DNA Damage. Nat. Cell Biol. 2002, 4, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, R.L.; Ziegler, T.D.; Supakorndej, T.; Terns, R.M.; Terns, M.P. Cell Cycle-Regulated Trafficking of Human Telomerase to Telomeres. MBoC 2006, 17, 955–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etheridge, K.T.; Banik, S.S.R.; Armbruster, B.N.; Zhu, Y.; Terns, R.M.; Terns, M.P.; Counter, C.M. The Nucleolar Localization Domain of the Catalytic Subunit of Human Telomerase. J. Biol. Chem. 2002, 277, 24764–24770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Lee, Y.S.; Jeong, S.A.; Khadka, P.; Roth, J.; Chung, I.K. Catalytically Active Telomerase Holoenzyme Is Assembled in the Dense Fibrillar Component of the Nucleolus during S Phase. Histochem. Cell Biol. 2014, 141, 137–152. [Google Scholar] [CrossRef]
- Khurts, S.; Masutomi, K.; Delgermaa, L.; Arai, K.; Oishi, N.; Mizuno, H.; Hayashi, N.; Hahn, W.C.; Murakami, S. Nucleolin Interacts with Telomerase. J. Biol. Chem. 2004, 279, 51508–51515. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Jin, R.; Zhang, B.; Chen, H.; Bai, Y.X.; Yang, P.X.; Han, S.W.; Xie, Y.H.; Huang, P.T.; Huang, C.; et al. Nucleolar Localization of TERT Is Unrelated to Telomerase Function in Human Cells. J. Cell Sci. 2008, 121, 2169–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laprade, H.; Querido, E.; Smith, M.J.; Guérit, D.; Crimmins, H.; Conomos, D.; Pourret, E.; Chartrand, P.; Sfeir, A. Single-Molecule Imaging of Telomerase RNA Reveals a Recruitment-Retention Model for Telomere Elongation. Mol. Cell 2020, 79, 115–126.e6. [Google Scholar] [CrossRef]
- Chiba, K.; Vogan, J.M.; Wu, R.A.; Gill, M.S.; Zhang, X.; Collins, K.; Hockemeyer, D. Endogenous Telomerase Reverse Transcriptase N-Terminal Tagging Affects Human Telomerase Function at Telomeres In Vivo. Mol. Cell. Biol. 2017, 37, e00541-16. [Google Scholar] [CrossRef] [Green Version]
- Strahl, B.D.; Allis, C.D. The Language of Covalent Histone Modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Verheggen, C.; Neel, H.; Cammas, A.; Vagner, S.; Soret, J.; Bertrand, E.; Bordonné, R. Characterization of a Short Isoform of Human Tgs1 Hypermethylase Associating with Small Nucleolar Ribonucleoprotein Core Proteins and Produced by Limited Proteolytic Processing. J. Biol. Chem. 2008, 283, 2060–2069. [Google Scholar] [CrossRef] [Green Version]
- Buemi, V.; Schillaci, O.; Santorsola, M.; Bonazza, D.; Broccia, P.V.; Zappone, A.; Bottin, C.; Dell’Omo, G.; Kengne, S.; Cacchione, S.; et al. TGS1 Mediates 2,2,7-Trimethyl Guanosine Capping of the Human Telomerase RNA to Direct Telomerase Dependent Telomere Maintenance. Nat. Commun. 2022, 13, 2302. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Grenier St-Sauveur, V.; Bergeron, D.; Dupuis-Sandoval, F.; Scott, M.S.; Bachand, F. A Polyadenylation-Dependent 3′ End Maturation Pathway Is Required for the Synthesis of the Human Telomerase RNA. Cell Rep. 2015, 13, 2244–2257. [Google Scholar] [CrossRef] [Green Version]
- Machyna, M.; Kehr, S.; Straube, K.; Kappei, D.; Buchholz, F.; Butter, F.; Ule, J.; Hertel, J.; Stadler, P.F.; Neugebauer, K.M. The Coilin Interactome Identifies Hundreds of Small Noncoding RNAs That Traffic through Cajal Bodies. Mol. Cell 2014, 56, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Boulon, S.; Verheggen, C.; Jady, B.E.; Girard, C.; Pescia, C.; Paul, C.; Ospina, J.K.; Kiss, T.; Matera, A.G.; Bordonné, R.; et al. PHAX and CRM1 Are Required Sequentially to Transport U3 SnoRNA to Nucleoli. Mol. Cell 2004, 16, 777–787. [Google Scholar] [CrossRef]
- Bizarro, J.; Bhardwaj, A.; Smith, S.; Meier, U.T. Nopp140-Mediated Concentration of Telomerase in Cajal Bodies Regulates Telomere Length. Mol. Biol. Cell 2019, 30, 3136–3150. [Google Scholar] [CrossRef]
- Isaac, C.; Yang, Y.; Meier, U.T. Nopp140 Functions as a Molecular Link between the Nucleolus and the Coiled Bodies. J. Cell Biol. 1998, 142, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, R.L.; Abreu, E.B.; Ziegler, T.; Ly, H.; Counter, C.M.; Terns, R.M.; Terns, M.P. Telomerase Reverse Transcriptase Is Required for the Localization of Telomerase RNA to Cajal Bodies and Telomeres in Human Cancer Cells. MBoC 2008, 19, 3793–3800. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.C.; Zaug, A.J.; Cech, T.R. Live Cell Imaging Reveals the Dynamics of Telomerase Recruitment to Telomeres. Cell 2016, 166, 1188–1197.e9. [Google Scholar] [CrossRef] [Green Version]
- Trinkle-Mulcahy, L.; Sleeman, J.E. The Cajal Body and the Nucleolus: “In a Relationship” or “It’s Complicated”? RNA Biol. 2017, 14, 739–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread Occurrence of 5-Methylcytosine in Human Coding and Non-Coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Wang, H.; Cheng, X.; Fan, X.; Yang, F.; Zhang, M.; Chen, Y.; Tian, Y.; Liu, C.; Shao, D.; et al. HuR Regulates Telomerase Activity through TERC Methylation. Nat. Commun. 2018, 9, 2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Gu, X.; Xia, T.; Ma, Z.; Yang, Z.; Feng, H.L.; Zhao, Y.; Ma, W.; Ju, Z.; Gorospe, M.; et al. HuB and HuD Repress Telomerase Activity by Dissociating HuR from TERC. Nucleic Acids Res. 2021, 49, 2848–2858. [Google Scholar] [CrossRef]
- Kim, N.-K.; Theimer, C.A.; Mitchell, J.R.; Collins, K.; Feigon, J. Effect of Pseudouridylation on the Structure and Activity of the Catalytically Essential P6.1 Hairpin in Human Telomerase RNA. Nucleic Acids Res. 2010, 38, 6746–6756. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; León-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-Wide Mapping Reveals Widespread Dynamic-Regulated Pseudouridylation of NcRNA and MRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemora, G.; Handl, S.; Waldsich, C. Human Telomerase Reverse Transcriptase Binds to a Pre-Organized HTR In Vivo Exposing Its Template. Nucleic Acids Res. 2016, 44, 413–425. [Google Scholar] [CrossRef]
- Holt, S.E.; Aisner, D.L.; Shay, J.W.; Wright, W.E. Lack of Cell Cycle Regulation of Telomerase Activity in Human Cells. Proc. Natl. Acad. Sci. USA 1997, 94, 10687–10692. [Google Scholar] [CrossRef] [Green Version]
- Hou, M.; Xu, D.; Björkholm, M.; Gruber, A. Real-Time Quantitative Telomeric Repeat Amplification Protocol Assay for the Detection of Telomerase Activity2. Clin. Chem. 2001, 47, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Rojas, C.; Shippen, D.E. Telomerase Regulation. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2012, 730, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Heidenreich, B.; Kumar, R. TERT Promoter Mutations in Telomere Biology. Mutat. Res./Rev. Mutat. Res. 2017, 771, 15–31. [Google Scholar] [CrossRef]
- Listerman, I.; Sun, J.; Gazzaniga, F.S.; Lukas, J.L.; Blackburn, E.H. The Major Reverse Transcriptase–Incompetent Splice Variant of the Human Telomerase Protein Inhibits Telomerase Activity but Protects from Apoptosis. Cancer Res. 2013, 73, 2817–2828. [Google Scholar] [CrossRef] [Green Version]
- Hrdličková, R.; Nehyba, J.; Bose, H.R. Alternatively Spliced Telomerase Reverse Transcriptase Variants Lacking Telomerase Activity Stimulate Cell Proliferation. Mol. Cell Biol. 2012, 32, 4283–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colgin, L.M.; Wilkinso, C.; Englezou, A.; Kilian, A.; Robinson, M.O.; Reddel, R.R. The HTERTα Splice Variant Is a Dominant Negative Inhibitor of Telomerase Activity. Neoplasia 2000, 2, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludlow, A.T.; Wong, M.S.; Robin, J.D.; Batten, K.; Yuan, L.; Lai, T.-P.; Dahlson, N.; Zhang, L.; Mender, I.; Tedone, E.; et al. NOVA1 Regulates HTERT Splicing and Cell Growth in Non-Small Cell Lung Cancer. Nat. Commun. 2018, 9, 3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penev, A.; Bazley, A.; Shen, M.; Boeke, J.D.; Savage, S.A.; Sfeir, A. Alternative Splicing Is a Developmental Switch for HTERT Expression. Mol. Cell 2021, 81, 2349–2360.e6. [Google Scholar] [CrossRef]
- Chung, J.; Khadka, P.; Chung, I.K. Nuclear Import of HTERT Requires a Bipartite Nuclear Localization Signal and Akt-Mediated Phosphorylation. J. Cell Sci. 2012, 125, 2684–2697. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.A.; Kim, K.; Lee, J.H.; Cha, J.S.; Khadka, P.; Cho, H.-S.; Chung, I.K. Akt-Mediated Phosphorylation Increases the Binding Affinity of HTERT for Importin α to Promote Nuclear Translocation. J. Cell Sci. 2015, 128, 2287–2301. [Google Scholar] [CrossRef] [Green Version]
- Lagadari, M.; Zgajnar, N.R.; Gallo, L.I.; Galigniana, M.D. Hsp90-Binding Immunophilin FKBP51 Forms Complexes with HTERT Enhancing Telomerase Activity. Mol. Oncol. 2016, 10, 1086–1098. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.Y.; Her, J.; Oh, S.-Y.; Chung, I.K. Hsp90-Binding Immunophilin FKBP52 Modulates Telomerase Activity by Promoting the Cytoplasmic Retrotransport of HTERT. Biochem. J. 2016, 473, 3517–3532. [Google Scholar] [CrossRef]
- Kovalenko, O.A.; Caron, M.J.; Ulema, P.; Medrano, C.; Thomas, A.P.; Kimura, M.; Bonini, M.G.; Herbig, U.; Santos, J.H. A Mutant Telomerase Defective in Nuclear-Cytoplasmic Shuttling Fails to Immortalize Cells and Is Associated with Mitochondrial Dysfunction. Aging Cell 2010, 9, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, M.; Ando, Y.; Yamashita, T.; Matsuda, Y.; Shoji, S.; Morioka, M.S.; Kawaji, H.; Shiozawa, K.; Machitani, M.; Abe, T.; et al. CDK1 Dependent Phosphorylation of HTERT Contributes to Cancer Progression. Nat. Commun. 2020, 11, 1557. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.-Y.; Wang, X.; Jun, S.; Park, J.-I. Dyrk2-Associated EDD-DDB1-VprBP E3 Ligase Inhibits Telomerase by TERT Degradation. J. Biol. Chem. 2013, 288, 7252–7262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haendeler, J.; Hoffmann, J.; Brandes, R.P.; Zeiher, A.M.; Dimmeler, S. Hydrogen Peroxide Triggers Nuclear Export of Telomerase Reverse Transcriptase via Src Kinase Family-Dependent Phosphorylation of Tyrosine 707. Mol. Cell. Biol. 2003, 23, 4598–4610. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.S.; Kwon, T.; Kwon, D.Y.; Do, S.I. Akt Protein Kinase Enhances Human Telomerase Activity through Phosphorylation of Telomerase Reverse Transcriptase Subunit. J. Biol. Chem. 1999, 274, 13085–13090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, Y.; Yamashita, T.; Ye, J.; Yasukawa, M.; Yamakawa, K.; Mukai, Y.; Machitani, M.; Daigo, Y.; Miyagi, Y.; Yokose, T.; et al. Phosphorylation of HTERT at Threonine 249 Is a Novel Tumor Biomarker of Aggressive Cancer with Poor Prognosis in Multiple Organs. J. Pathol. 2022, 257, 172–185. [Google Scholar] [CrossRef]
- Jakob, S.; Schroeder, P.; Lukosz, M.; Büchner, N.; Spyridopoulos, I.; Altschmied, J.; Haendeler, J. Nuclear Protein Tyrosine Phosphatase Shp-2 Is One Important Negative Regulator of Nuclear Export of Telomerase Reverse Transcriptase. J. Biol. Chem. 2008, 283, 33155–33161. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhao, L.; Yang, Z.; Funder, J.W.; Liu, J.P. Telomerase Is Controlled by Protein Kinase Calpha in Human Breast Cancer Cells. J. Biol. Chem. 1998, 273, 33436–33442. [Google Scholar] [CrossRef] [Green Version]
- Kharbanda, S.; Kumar, V.; Dhar, S.; Pandey, P.; Chen, C.; Majumder, P.; Yuan, Z.-M.; Whang, Y.; Strauss, W.; Pandita, T.K.; et al. Regulation of the HTERT Telomerase Catalytic Subunit by the C-Abl Tyrosine Kinase. Curr. Biol. 2000, 10, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Xi, P.; Zhou, L.; Wang, M.; Liu, J.-P.; Cong, Y.-S. Serine/Threonine-Protein Phosphatase 2A Physically Interacts with Human Telomerase Reverse Transcriptase HTERT and Regulates Its Subcellular Distribution. J. Cell. Biochem. 2013, 114, 409–417. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.-M.; Kang, M.R.; Oh, S.-Y.; Lee, T.H.; Muller, M.T.; Chung, I.K. Ubiquitin Ligase MKRN1 Modulates Telomere Length Homeostasis through a Proteolysis of HTERT. Genes Dev. 2005, 19, 776–781. [Google Scholar] [CrossRef] [Green Version]
- Salvatico, J.; Kim, J.H.; Chung, I.K.; Muller, M.T. Differentiation Linked Regulation of Telomerase Activity by Makorin-1. Mol. Cell. Biochem. 2010, 342, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Panneer Selvam, S.; De Palma, R.M.; Oaks, J.J.; Oleinik, N.; Peterson, Y.K.; Stahelin, R.V.; Skordalakes, E.; Ponnusamy, S.; Garrett-Mayer, E.; Smith, C.D.; et al. Binding of the Sphingolipid S1P to HTERT Stabilizes Telomerase at the Nuclear Periphery by Allosterically Mimicking Protein Phosphorylation. Sci. Signal. 2015, 8, ra58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, W.; Lee, E.-W.; Lee, D.; Yang, M.-R.; Ko, A.; Yoon, C.-H.; Lee, H.-W.; Bae, Y.-S.; Choi, C.Y.; Song, J. Hdm2 Negatively Regulates Telomerase Activity by Functioning as an E3 Ligase of HTERT. Oncogene 2010, 29, 4101–4112. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wang, W.; Yang, H.; Wu, L.; He, Z.; Zhou, S.; Zhao, H.; Fu, Z.; Zhou, F.; Zhou, Y. UBE2D3 Gene Overexpression Increases Radiosensitivity of EC109 Esophageal Cancer Cells In Vitro and In Vivo. Oncotarget 2016, 7, 32543–32553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Khadka, P.; Baek, S.H.; Chung, I.K. CHIP Promotes Human Telomerase Reverse Transcriptase Degradation and Negatively Regulates Telomerase Activity. J. Biol. Chem. 2010, 285, 42033–42045. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Sun, L.; Liu, N.; Wei, Q.; Jiang, L.; Tong, X.; Ye, X. Polo-like Kinase 1 (Plk1) Up-Regulates Telomerase Activity by Affecting Human Telomerase Reverse Transcriptase (HTERT) Stability. J. Biol. Chem. 2015, 290, 18865–18873. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, S.; Mondal, P.; Sen, S.; Sengupta Bandyopadhyay, S.; Das, C. SUMO E3 Ligase CBX4 Regulates HTERT-Mediated Transcription of CDH1 and Promotes Breast Cancer Cell Migration and Invasion. Biochem. J. 2020, 477, 3803–3818. [Google Scholar] [CrossRef]
- Udroiu, I.; Marinaccio, J.; Sgura, A. Many Functions of Telomerase Components: Certainties, Doubts, and Inconsistencies. Int. J. Mol. Sci. 2022, 23, 15189. [Google Scholar] [CrossRef]
- Soohoo, C.Y.; Shi, R.; Lee, T.H.; Huang, P.; Lu, K.P.; Zhou, X.Z. Telomerase Inhibitor PinX1 Provides a Link between TRF1 and Telomerase to Prevent Telomere Elongation. J. Biol. Chem. 2011, 286, 3894–3906. [Google Scholar] [CrossRef] [Green Version]
- Banik, S.S.R.; Counter, C.M. Characterization of Interactions between PinX1 and Human Telomerase Subunits HTERT and HTR. J. Biol. Chem. 2004, 279, 51745–51748. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.Z.; Lu, K.P. The Pin2/TRF1-Interacting Protein PinX1 Is a Potent Telomerase Inhibitor. Cell 2001, 107, 347–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Da, L.; Xu, Y.; Xu, M.; Song, L.; Li, T.; Zhao, M. C-Terminal Amino Acids 290–328 of LPTS/PinX1 Confer Telomerase Inhibition. Biochem. Biophys. Res. Commun. 2010, 398, 683–689. [Google Scholar] [CrossRef]
- Zhang, B.; Bai, Y.X.; Ma, H.H.; Feng, F.; Jin, R.; Wang, Z.L.; Lin, J.; Sun, S.P.; Yang, P.; Wang, X.X.; et al. Silencing PinX1 Compromises Telomere Length Maintenance As Well As Tumorigenicity in Telomerase-Positive Human Cancer Cells. Cancer Res. 2009, 69, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, D.H.-C.; Kung, H.-F.; Huang, J.-J.; Shaw, P.-C. PinX1 Is Involved in Telomerase Recruitment and Regulates Telomerase Function by Mediating Its Localization. FEBS Lett. 2012, 586, 3166–3171. [Google Scholar] [CrossRef] [Green Version]
- Cheung, D.H.-C.; Ho, S.-T.; Lau, K.-F.; Jin, R.; Wang, Y.-N.; Kung, H.-F.; Huang, J.-J.; Shaw, P.-C. Nucleophosmin Interacts with PIN2/TERF1-Interacting Telomerase Inhibitor 1 (PinX1) and Attenuates the PinX1 Inhibition on Telomerase Activity. Sci. Rep. 2017, 7, 43650. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.-T.; Jin, R.; Cheung, D.H.-C.; Huang, J.-J.; Shaw, P.-C. The PinX1/NPM Interaction Associates with HTERT in Early-S Phase and Facilitates Telomerase Activation. Cell Biosci. 2019, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, Y.; Chen, G.; Xing, Z.; Zhao, J.; Yokoyama, K.K.; Li, T.; Zhao, M. Human MCRS2, a Cell-Cycle-Dependent Protein, Associates with LPTS/PinX1 and Reduces the Telomere Length. Biochem. Biophys. Res. Commun. 2004, 316, 1116–1123. [Google Scholar] [CrossRef]
- Jády, B.E.; Richard, P.; Bertrand, E.; Kiss, T. Cell Cycle-Dependent Recruitment of Telomerase RNA and Cajal Bodies to Human Telomeres. MBoC 2006, 17, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Stern, J.L.; Zyner, K.G.; Pickett, H.A.; Cohen, S.B.; Bryan, T.M. Telomerase Recruitment Requires Both TCAB1 and Cajal Bodies Independently. Mol. Cell. Biol. 2012, 32, 2384–2395. [Google Scholar] [CrossRef] [Green Version]
- Zhong, F.L.; Batista, L.F.Z.; Freund, A.; Pech, M.F.; Venteicher, A.S.; Artandi, S.E. TPP1 OB-Fold Domain Controls Telomere Maintenance by Recruiting Telomerase to Chromosome Ends. Cell 2012, 150, 481–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofari, G.; Adolf, E.; Reichenbach, P.; Sikora, K.; Terns, R.M.; Terns, M.P.; Lingner, J. Human Telomerase RNA Accumulation in Cajal Bodies Facilitates Telomerase Recruitment to Telomeres and Telomere Elongation. Mol. Cell 2007, 27, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Deng, Z.; Jiang, S.; Hu, Q.; Liu, H.; Songyang, Z.; Ma, W.; Chen, S.; Zhao, Y. Human Cells Lacking Coilin and Cajal Bodies Are Proficient in Telomerase Assembly, Trafficking and Telomere Maintenance. Nucleic Acids Res. 2015, 43, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, R.L.; Li, J.; Culp, B.R.; Terns, R.M.; Terns, M.P. A Cajal Body-Independent Pathway for Telomerase Trafficking in Mice. Exp. Cell Res. 2010, 316, 2797–2809. [Google Scholar] [CrossRef] [Green Version]
- de Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, M.S.; Safari, A.; Xin, H.; Liu, D.; Songyang, Z. A Critical Role for TPP1 and TIN2 Interaction in High-Order Telomeric Complex Assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 11874–11879. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, I.; de Lange, T. Shelterin. Curr. Biol. 2016, 26, R397–R399. [Google Scholar] [CrossRef]
- Chong, L.; van Steensel, B.; Broccoli, D.; Erdjument-Bromage, H.; Hanish, J.; Tempst, P.; de Lange, T. A Human Telomeric Protein. Science 1995, 270, 1663–1667. [Google Scholar] [CrossRef]
- Broccoli, D.; Smogorzewska, A.; Chong, L.; de Lange, T. Human Telomeres Contain Two Distinct Myb–Related Proteins, TRF1 and TRF2. Nat. Genet. 1997, 17, 231–235. [Google Scholar] [CrossRef]
- Loayza, D.; de Lange, T. POT1 as a Terminal Transducer of TRF1 Telomere Length Control. Nature 2003, 423, 1013–1018. [Google Scholar] [CrossRef]
- Lei, M.; Podell, E.R.; Cech, T.R. Structure of Human POT1 Bound to Telomeric Single-Stranded DNA Provides a Model for Chromosome End-Protection. Nat. Struct. Mol. Biol. 2004, 11, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Z.-S.; Hockemeyer, D.; Krutchinsky, A.N.; Loayza, D.; Hooper, S.M.; Chait, B.T.; de Lange, T. POT1-Interacting Protein PIP1: A Telomere Length Regulator That Recruits POT1 to the TIN2/TRF1 Complex. Genes Dev. 2004, 18, 1649–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, R.; Chen, Y.; Lei, M.; Chang, S. TRF2-RAP1 Is Required to Protect Telomeres from Engaging in Homologous Recombination-Mediated Deletions and Fusions. Nat. Commun. 2016, 7, 10881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.J.; Cech, T.R. Shaping Human Telomeres: From Shelterin and CST Complexes to Telomeric Chromatin Organization. Nat. Rev. Mol. Cell. Biol. 2021, 22, 283–298. [Google Scholar] [CrossRef]
- Abreu, E.; Aritonovska, E.; Reichenbach, P.; Cristofari, G.; Culp, B.; Terns, R.M.; Lingner, J.; Terns, M.P. TIN2-Tethered TPP1 Recruits Human Telomerase to Telomeres In Vivo. Mol. Cell. Biol. 2010, 30, 2971–2982. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, J.; Bell, C.F.; Weidenfeld, I.; Zaug, A.J.; Leinwand, L.A.; Cech, T.R. The TEL Patch of Telomere Protein TPP1 Mediates Telomerase Recruitment and Processivity. Nature 2012, 492, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Podell, E.R.; Zaug, A.J.; Yang, Y.; Baciu, P.; Cech, T.R.; Lei, M. The POT1–TPP1 Telomere Complex Is a Telomerase Processivity Factor. Nature 2007, 445, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like Mammalian Ctc1-Stn1-Ten1 Complex Binds to Single-Stranded DNA and Protects Telomeres Independently of the Pot1 Pathway. Mol. Cell 2009, 36, 193–206. [Google Scholar] [CrossRef]
- Chen, L.-Y.; Redon, S.; Lingner, J. The Human CST Complex Is a Terminator of Telomerase Activity. Nature 2012, 488, 540–544. [Google Scholar] [CrossRef]
- Lim, C.J.; Barbour, A.T.; Zaug, A.J.; Goodrich, K.J.; McKay, A.E.; Wuttke, D.S.; Cech, T.R. The Structure of Human CST Reveals a Decameric Assembly Bound to Telomeric DNA. Science 2020, 368, 1081–1085. [Google Scholar] [CrossRef]
- Timashev, L.A.; De Lange, T. Characterization of T-Loop Formation by TRF2. Nucleus 2020, 11, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; de Lange, T. Mammalian Telomeres End in a Large Duplex Loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doksani, Y.; Wu, J.Y.; de Lange, T.; Zhuang, X. Super-Resolution Fluorescence Imaging of Telomeres Reveals TRF2-Dependent T-Loop Formation. Cell 2013, 155, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Le Guen, T.; Jullien, L.; Touzot, F.; Schertzer, M.; Gaillard, L.; Perderiset, M.; Carpentier, W.; Nitschke, P.; Picard, C.; Couillault, G.; et al. Human RTEL1 Deficiency Causes Hoyeraal–Hreidarsson Syndrome with Short Telomeres and Genome Instability. Hum. Mol. Genet. 2013, 22, 3239–3249. [Google Scholar] [CrossRef] [Green Version]
- Sfeir, A.; Kosiyatrakul, S.T.; Hockemeyer, D.; MacRae, S.L.; Karlseder, J.; Schildkraut, C.L.; de Lange, T. Mammalian Telomeres Resemble Fragile Sites and Require TRF1 for Efficient Replication. Cell 2009, 138, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Z.; Glousker, G.; Molczan, A.; Fox, A.J.; Lamm, N.; Dheekollu, J.; Weizman, O.-E.; Schertzer, M.; Wang, Z.; Vladimirova, O.; et al. Inherited Mutations in the Helicase RTEL1 Cause Telomere Dysfunction and Hoyeraal–Hreidarsson Syndrome. Proc. Natl. Acad. Sci. USA 2013, 110, E3408–E3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarek, G.; Vannier, J.-B.; Panier, S.; Petrini, J.H.J.; Boulton, S.J. TRF2 Recruits RTEL1 to Telomeres in S Phase to Promote T-Loop Unwinding. Mol. Cell 2015, 57, 622–635. [Google Scholar] [CrossRef] [Green Version]
- Barber, L.J.; Youds, J.L.; Ward, J.D.; McIlwraith, M.J.; O’Neil, N.J.; Petalcorin, M.I.R.; Martin, J.S.; Collis, S.J.; Cantor, S.B.; Auclair, M.; et al. RTEL1 Maintains Genomic Stability by Suppressing Homologous Recombination. Cell 2008, 135, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Vannier, J.-B.; Sandhu, S.; Petalcorin, M.I.R.; Wu, X.; Nabi, Z.; Ding, H.; Boulton, S.J. RTEL1 Is a Replisome-Associated Helicase That Promotes Telomere and Genome-Wide Replication. Science 2013, 342, 239–242. [Google Scholar] [CrossRef]
- Porreca, R.M.; Glousker, G.; Awad, A.; Matilla Fernandez, M.I.; Gibaud, A.; Naucke, C.; Cohen, S.B.; Bryan, T.M.; Tzfati, Y.; Draskovic, I.; et al. Human RTEL1 Stabilizes Long G-Overhangs Allowing Telomerase-Dependent over-Extension. Nucleic Acids Res. 2018, 46, 4533–4545. [Google Scholar] [CrossRef] [Green Version]
- Tong, A.S.; Stern, J.L.; Sfeir, A.; Kartawinata, M.; de Lange, T.; Zhu, X.-D.; Bryan, T.M. ATM and ATR Signaling Regulate the Recruitment of Human Telomerase to Telomeres. Cell Rep. 2015, 13, 1633–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.S.; Bohrson, C.; Pike, A.M.; Wheelan, S.J.; Greider, C.W. ATM Kinase Is Required for Telomere Elongation in Mouse and Human Cells. Cell Rep. 2015, 13, 1623–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKerlie, M.; Lin, S.; Zhu, X.-D. ATM Regulates Proteasome-Dependent Subnuclear Localization of TRF1, Which Is Important for Telomere Maintenance. Nucleic Acids Res. 2012, 40, 3975–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, M.; Zheng, W.; Huang, W.; Wang, Z.; Jin, K.; Liu, L.; Yu, Z. Dynamics of TRF1 Organizing a Single Human Telomere. Nucleic Acids Res. 2021, 49, 760–775. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, S.; Zhu, X.-D. MRE11-RAD50-NBS1 and ATM Function as Co-Mediators of TRF1 in Telomere Length Control. Nat. Struct. Mol. Biol. 2007, 14, 832–840. [Google Scholar] [CrossRef]
- Lee, J.-H.; Mand, M.R.; Deshpande, R.A.; Kinoshita, E.; Yang, S.-H.; Wyman, C.; Paull, T.T. Ataxia Telangiectasia-Mutated (ATM) Kinase Activity Is Regulated by ATP-Driven Conformational Changes in the Mre11/Rad50/Nbs1 (MRN) Complex. J. Biol. Chem. 2013, 288, 12840–12851. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.R.; Zhu, X.-D. Post-Translational Modifications of TRF1 and TRF2 and Their Roles in Telomere Maintenance. Mech. Ageing Dev. 2012, 133, 421–434. [Google Scholar] [CrossRef]
- Imran, S.A.M.; Yazid, M.D.; Cui, W.; Lokanathan, Y. The Intra- and Extra-Telomeric Role of TRF2 in the DNA Damage Response. Int. J. Mol. Sci. 2021, 22, 9900. [Google Scholar] [CrossRef]
- Roake, C.M.; Chen, L.; Chakravarthy, A.L.; Ferrell, J.E.; Raffa, G.D.; Artandi, S.E. Disruption of Telomerase RNA Maturation Kinetics Precipitates Disease. Mol. Cell 2019, 74, 688–700.e3. [Google Scholar] [CrossRef]
- Qin, J.; Autexier, C. Regulation of Human Telomerase RNA Biogenesis and Localization. RNA Biol. 2021, 18, 305–315. [Google Scholar] [CrossRef]
- Grandin, N.; Charbonneau, M. Hsp90 Levels Affect Telomere Length in Yeast. Mol. Genet. Genom. 2001, 265, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Toogun, O.A.; Zeiger, W.; Freeman, B.C. The P23 Molecular Chaperone Promotes Functional Telomerase Complexes through DNA Dissociation. Proc. Natl. Acad. Sci. USA 2007, 104, 5765–5770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, A.A.; Weber, C.A.; Jülicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, J.; Wright, W.E.; Shay, J.W. Clustered Telomeres in Phase-Separated Nuclear Condensates Engage Mitotic DNA Synthesis through BLM and RAD52. Genes Dev. 2019, 33, 814–827. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, R.; Tones, J.; Liu, M.; Dilley, R.L.; Chenoweth, D.M.; Greenberg, R.A.; Lampson, M.A. Nuclear Body Phase Separation Drives Telomere Clustering in ALT Cancer Cells. MBoC 2020, 31, 2048–2056. [Google Scholar] [CrossRef]
- Smith, S.; Giriat, I.; Schmitt, A.; de Lange, T. Tankyrase, a Poly(ADP-Ribose) Polymerase at Human Telomeres. Science 1998, 282, 1484–1487. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shepelev, N.; Dontsova, O.; Rubtsova, M. Post-Transcriptional and Post-Translational Modifications in Telomerase Biogenesis and Recruitment to Telomeres. Int. J. Mol. Sci. 2023, 24, 5027. https://doi.org/10.3390/ijms24055027
Shepelev N, Dontsova O, Rubtsova M. Post-Transcriptional and Post-Translational Modifications in Telomerase Biogenesis and Recruitment to Telomeres. International Journal of Molecular Sciences. 2023; 24(5):5027. https://doi.org/10.3390/ijms24055027
Chicago/Turabian StyleShepelev, Nikita, Olga Dontsova, and Maria Rubtsova. 2023. "Post-Transcriptional and Post-Translational Modifications in Telomerase Biogenesis and Recruitment to Telomeres" International Journal of Molecular Sciences 24, no. 5: 5027. https://doi.org/10.3390/ijms24055027
APA StyleShepelev, N., Dontsova, O., & Rubtsova, M. (2023). Post-Transcriptional and Post-Translational Modifications in Telomerase Biogenesis and Recruitment to Telomeres. International Journal of Molecular Sciences, 24(5), 5027. https://doi.org/10.3390/ijms24055027