
Chronic Pain-Associated Cardiovascular Disease: The Role of Sympathetic Nerve Activity

Abstract
:1. Introduction
2. Sympathetic Nervous System and Cardiovascular Disease
3. Basic Neurocircuitry of the Sympathetic Nervous System
4. Basic Neurocircuitry of Nociception
5. The Relationship between Blood Pressure and Acute Pain Sensitivity
6. Chronic Pain as a Driver of SNS Overactivation
7. Maladaptive Changes in the Neural Circuitry Leading to Sympathetic Overactivation and Chronic Pain
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| α-2 | Alpha-2 adrenergic receptors |
| ACC | Anterior cingulate cortex |
| DH | Dorsal horn |
| dlPFC | Dorsolateral prefrontal cortex |
| DMH | Dorsomedial hypothalamus |
| CVLM | Caudal ventrolateral medulla |
| fMRI | Functional magnetic resonance imaging |
| IC | Insular cortex |
| IML | Intermediolateral cell column |
| LC | Locus coeruleus |
| MRI | Magnetic resonance imaging |
| mPFC | Medial prefrontal cortex |
| GABA | γ- aminobutyric acid |
| NTS | Nucleus of the solitary tract |
| PFC | Prefrontal cortex |
| PVN | Paraventricular nucleus |
| RF | Reticular formation |
| RVLM | Rostral ventrolateral medulla |
| SNA | Sympathetic nerve activity |
| SNS | Sympathetic nerve system |
| SPN | Sympathetic preganglionic neurons |
References
- Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [CrossRef]
- Fayaz, A.; Ayis, S.; Panesar, S.S.; Langford, R.M.; Donaldson, L.J. Assessing the relationship between chronic pain and cardiovascular disease: A systematic review and meta-analysis. Scand. J. Pain 2016, 13, 76–90. [Google Scholar] [CrossRef]
- Oliveira, C.B.; Maher, C.G.; Franco, M.R.; Kamper, S.J.; Williams, C.M.; Silva, F.G.; Pinto, R.Z. Co-occurrence of Chronic Musculoskeletal Pain and Cardiovascular Diseases: A Systematic Review with Meta-analysis. Pain Med. 2020, 21, 1106–1121. [Google Scholar] [CrossRef]
- Macfarlane, G.J.; Barnish, M.S.; Jones, G.T. Persons with chronic widespread pain experience excess mortality: Longitudinal results from UK Biobank and meta-analysis. Ann. Rheum. Dis. 2017, 76, 1815–1822. [Google Scholar] [CrossRef]
- Holmberg, T.; Davidsen, M.; Thygesen, L.C.; Krøll, M.J.; Tolstrup, J.S. Mortality among persons experiencing musculoskeletal pain: A prospective study among Danish men and women. BMC Musculoskelet. Disord. 2020, 21, 666. [Google Scholar] [CrossRef] [PubMed]
- Tesarz, J.; Eich, W.; Baumeister, D.; Kohlmann, T.; D’Agostino, R.; Schuster, A.K. Widespread pain is a risk factor for cardiovascular mortality: Results from the Framingham Heart Study. Eur. Heart J. 2019, 40, 1609–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBeth, J.; Symmons, D.; Silman, A.; Allison, T.; Webb, R.; Brammah, T.; Macfarlane, G. Musculoskeletal pain is associated with a long-term increased risk of cancer and cardiovascular-related mortality. Rheumatology 2009, 48, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Andersson, H.I. Increased mortality among individuals with chronic widespread pain relates to lifestyle factors: A prospective population-based study. Disabil. Rehabil. 2009, 31, 1980–1987. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, I.; Ortolá, R.; Graciani, A.; Martínez-Gómez, D.; Banegas, J.R.; Rodríguez-Artalejo, F.; García-Esquinas, E. Pain characteristics, cardiovascular risk factors, and cardiovascular disease. J. Gerontol. Ser. A 2022, 77, 204–213. [Google Scholar] [CrossRef]
- Lindgren, H.; Bergman, S. Chronic musculoskeletal pain predicted hospitalisation due to serious medical conditions in a 10 year follow up study. BMC Musculoskelet. Disord. 2010, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendzerska, T.; Jüni, P.; King, L.K.; Croxford, R.; Stanaitis, I.; Hawker, G.A. The longitudinal relationship between hand, hip and knee osteoarthritis and cardiovascular events: A population-based cohort study. Osteoarthr. Cartil. 2017, 25, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Atiquzzaman, M.; Karim, M.E.; Kopec, J.; Wong, H.; Anis, A.H. Role of nonsteroidal antiinflammatory drugs in the association between osteoarthritis and cardiovascular diseases: A longitudinal study. Arthritis Rheumatol. 2019, 71, 1835–1843. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kopec, J.A.; Anis, A.H.; Cibere, J.; Goldsmith, C.H. Risk of cardiovascular disease in patients with osteoarthritis: A prospective longitudinal study. Arthritis Care Res. 2013, 65, 1951–1958. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-H.; Chen, J.-H.; Lan, J.-L.; Wang, Y.-C.; Tseng, C.-H.; Hsu, C.-Y.; Huang, L. Increased risk of coronary heart disease in patients with primary fibromyalgia and those with concomitant comorbidity—A Taiwanese population-based cohort study. PLoS ONE 2015, 10, e0137137. [Google Scholar] [CrossRef] [Green Version]
- Kluzek, S.; Sanchez-Santos, M.; Leyland, K.; Judge, A.; Spector, T.; Hart, D.; Cooper, C.; Newton, J.; Arden, N. Painful knee but not hand osteoarthritis is an independent predictor of mortality over 23 years follow-up of a population-based cohort of middle-aged women. Ann. Rheum. Dis. 2016, 75, 1749–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rönnegård, A.-S.; Nowak, C.; Äng, B.; Ärnlöv, J. The association between short-term, chronic localized and chronic widespread pain and risk for cardiovascular disease in the UK Biobank. Eur. J. Prev. Cardiol. 2022, 29, 1994–2002. [Google Scholar] [CrossRef] [PubMed]
- Ha, I.-H.; Lee, J.; Kim, M.-R.; Kim, H.; Shin, J.-S. The association between the history of cardiovascular diseases and chronic low back pain in South Koreans: A cross-sectional study. PLoS ONE 2014, 9, e93671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, S.; McBeth, J.; Macfarlane, G.; Hannaford, P.; Symmons, D. Self-reported pain severity is associated with a history of coronary heart disease. Eur. J. Pain 2015, 19, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Von Korff, M.; Crane, P.; Lane, M.; Miglioretti, D.L.; Simon, G.; Saunders, K.; Stang, P.; Brandenburg, N.; Kessler, R. Chronic spinal pain and physical–mental comorbidity in the United States: Results from the national comorbidity survey replication. Pain 2005, 113, 331–339. [Google Scholar] [CrossRef]
- Keller, J.J.; Chen, Y.K.; Lin, H.C. Comorbidities of bladder pain syndrome/interstitial cystitis: A population-based study. BJU Int. 2012, 110, E903–E909. [Google Scholar] [CrossRef]
- Dreyer, L.; Kendall, S.; Danneskiold-Samsøe, B.; Bartels, E.M.; Bliddal, H. Mortality in a cohort of Danish patients with fibromyalgia: Increased frequency of suicide. Arthritis Rheum. 2010, 62, 3101–3108. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Devine, A.; Dick, I.M.; Prince, R.L. Association of back pain frequency with mortality, coronary heart events, mobility, and quality of life in elderly women. Spine 2007, 32, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- Conway, L.C.; Smith, B.H.; Hocking, L.J.; McGilchrist, M.M.; Dominiczak, A.F.; Morris, A.; Porteous, D.J.; Goebel, A.; Goodson, N.J. The prevalence of rose angina is increased in people reporting chronic pain: Results from a cross-sectional general population study. Rheumatology 2014, 53, i68. [Google Scholar] [CrossRef] [Green Version]
- Svensson, H.-O.; Vedin, A.; Wilhelmsson, C.; Andersson, G.B. Low-back pain in relation to other diseases and cardiovascular risk factors. Spine 1983, 8, 277–285. [Google Scholar] [CrossRef]
- Pontari, M.A.; McNaughton-Collins, M.; O’leary, M.P.; Calhoun, E.A.; Jang, T.; Kusek, J.W.; Landis, J.R.; Knauss, J.; Litwin, M.S.; Group, C.S. A case-control study of risk factors in men with chronic pelvic pain syndrome. BJU Int. 2005, 96, 559–565. [Google Scholar] [CrossRef]
- Ablin, J.N.; Beilinson, N.; Aloush, V.; Elkayam, O.; Finkelstein, A. Association between fibromyalgia and coronary heart disease and coronary catheterization. Clin. Cardiol. Int. Index. Peer-Rev. J. Adv. Treat. Cardiovasc. Dis. 2009, 32, E7–E11. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-S.; Fan, Y.-C.; Huang, C.-J. Fibromyalgia is associated with coronary heart disease: A population-based cohort study. Reg. Anesth. Pain Med. 2015, 40, 37–42. [Google Scholar] [CrossRef]
- Mäkela, M.; Heliövaara, M.; Sievers, K.; Impivaara, O.; Knekt, P.; Aromaa, A. Prevalence, determinants, and consequences of chronic neck pain in Finland. Am. J. Epidemiol. 1991, 134, 1356–1367. [Google Scholar] [CrossRef]
- Ohayon, M.M.; Stingl, J.C. Prevalence and comorbidity of chronic pain in the German general population. J. Psychiatr. Res. 2012, 46, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.; McDonough, S.; Kirwan, J.; Leveille, S.; Martin, D. An investigation of association between chronic musculoskeletal pain and cardiovascular disease in the Health Survey for England (2008). Eur. J. Pain 2014, 18, 740–750. [Google Scholar] [CrossRef]
- Heliövaara, M.; Mäkelä, M.; Aromaa, A.; Impivaara, O.; Knekt, P.; Reunanen, A. Low back pain and subsequent cardiovascular mortality. Spine 1995, 20, 2109–2111. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.H.; Elliott, A.M.; Hannaford, P.C. Pain and subsequent mortality and cancer among women in the Royal College of General Practitioners Oral Contraception Study. Br. J. Gen. Pract. 2003, 53, 45–46. [Google Scholar] [PubMed]
- Torrance, N.; Elliott, A.M.; Lee, A.J.; Smith, B.H. Severe chronic pain is associated with increased 10 year mortality. A cohort record linkage study. Eur. J. Pain 2010, 14, 380–386. [Google Scholar] [CrossRef]
- Sherrington, C.S. Experiments on the value of vascular and visceral factors for the genesis of emotion. Proc. R. Soc. Lond. 1900, 66, 390–403. [Google Scholar]
- Sherrington, C.S. Address on the spinal animal. Med. Chir. Trans. 1899, 82, 449. [Google Scholar] [CrossRef]
- Reis, D.J.; Ruggiero, D.A.; Morrison, S.F. The CI area of the rostral ventrolateral medulla oblongata: A critical brainstem region for control of resting and reflex integration of arterial pressure. Am. J. Hypertens. 1989, 2, 363S–374S. [Google Scholar] [CrossRef] [PubMed]
- Stornetta, R.L.; Morrison, S.F.; Ruggiero, D.A.; Reis, D.J. Neurons of rostral ventrolateral medulla mediate somatic pressor reflex. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1989, 256, R448–R462. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Fields, H.L. Endogenous pain control systems: Brainstem spinal pathways and endorphin circuitry. Annu. Rev. Neurosci. 1984, 7, 309–338. [Google Scholar] [CrossRef]
- Malpas, S.C. Sympathetic nervous system overactivity and its role in the development of cardiovascular disease. Physiol. Rev. 2010, 90, 513–557. [Google Scholar] [CrossRef]
- Rozanski, A.; Blumenthal, J.A.; Kaplan, J. Impact of psychological factors on the pathogenesis of cardiovascular disease and implications for therapy. Circulation 1999, 99, 2192–2217. [Google Scholar] [CrossRef] [Green Version]
- Maixner, W.; Sigurdsson, A.; Fillingim, R.; Lundeen, T.; Booker, D. Regulation of acute and chronic orofacial pain. In Orofacial Pain and Temopromandibular Disorders; Raven Press: New York, NY, USA, 1995; pp. 85–102. [Google Scholar]
- Bruehl, S.; Chung, O.Y.; Ward, P.; Johnson, B.; McCubbin, J.A. The relationship between resting blood pressure and acute pain sensitivity in healthy normotensives and chronic back pain sufferers: The effects of opioid blockade. Pain 2002, 100, 191–201. [Google Scholar] [CrossRef]
- Bruehl, S.; Burns, J.W.; McCubbin, J.A. Altered cardiovascular/pain regulatory relationships in chronic pain. Int. J. Behav. Med. 1998, 5, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Brody, S.; Angrilli, A.; Weiss, U.; Birbaumer, N.; Mini, A.; Veit, R.; Rau, H. Somatotosensory evoked potentials during baroreceptor stimulation in chronic low back pain patients and normal controls. Int. J. Psychophysiol. 1997, 25, 201–210. [Google Scholar] [CrossRef]
- Perry, F.; Heller, P.H.; Kamiya, J.; Levine, J.D. Altered autonomic function in patients with arthritis or with chronic myofascial pain. Pain 1989, 39, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Hjemdahl, P.; Larsson, P.; Wallen, N. Effects of stress and beta-blockade on platelet function. Circulation 1991, 84, VI44–VI61. [Google Scholar] [PubMed]
- Ardlie, N.; Glew, G.; Schwartz, C. Influence of catecholamines on nucleotide-induced platelet aggregation. Nature 1966, 212, 415–417. [Google Scholar] [CrossRef]
- O’brien, J. Some effects of adrenaline and anti-adrenaline compounds on platelets in vitro and in vivo. Nature 1963, 200, 763–764. [Google Scholar] [CrossRef]
- Remme, W. The sympathetic nervous system and ischaemic heart disease. Eur. Heart J. 1998, 19, F62–F71. [Google Scholar]
- Benarroch, E.E. The central autonomic network: Functional organization, dysfunction, and perspective. In Proceedings of Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 1993; pp. 988–1001. [Google Scholar]
- Calaresu, F.; Yardley, C. Medullary basal sympathetic tone. Annu. Rev. Physiol. 1988, 50, 511–524. [Google Scholar] [CrossRef]
- Guertzenstein, P.; Silver, A. Fall in blood pressure produced from discrete regions of the ventral surface of the medulla by glycine and lesions. J. Physiol. 1974, 242, 489–503. [Google Scholar] [CrossRef]
- Stein, R.; Weaver, L.; Yardley, C. Ventrolateral medullary neurones: Effects on magnitude and rhythm of discharge of mesenteric and renal nerves in cats. J. Physiol. 1989, 408, 571–586. [Google Scholar] [CrossRef] [Green Version]
- Guyenet, P.G.; Haselton, J.R.; Sun, M.-K. Sympathoexcitatory neurons of the rostroventrolateral medulla and the origin of the sympathetic vasomotor tone. Prog. Brain Res. 1989, 81, 105–116. [Google Scholar] [PubMed]
- Sakima, A.; Yamazato, M.; Sesoko, S.; Muratani, H.; Fukiyama, K. Cardiovascular and sympathetic effects of L-glutamate and glycine injected into the rostral ventrolateral medulla of conscious rats. Hypertens. Res. 2000, 23, 633–641. [Google Scholar] [CrossRef]
- Willette, R.; Barcas, P.; Krieger, A.; Sapru, H.N. Vasopressor and depressor areas in the rat medulla: Identification by microinjection of L-glutamate. Neuropharmacology 1983, 22, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Dampney, R.; Moon, E.A. Role of ventrolateral medulla in vasomotor response to cerebral ischemia. Am. J. Physiol.-Heart Circ. Physiol. 1980, 239, H349–H358. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-K.; Reis, D.J. Central neural mechanisms mediating excitation of sympathetic neurons by hypoxia. Prog. Neurobiol. 1994, 44, 197–219. [Google Scholar] [CrossRef]
- Kubo, T.; Goshima, Y.; Hata, H.; Misu, Y. Evidence that endogenous catecholamines are involved inα2-adrenoceptor-mediated modulation of the aortic baroreceptor reflex in the nucleus tractus solitarii of the rat. Brain Res. 1990, 526, 313–317. [Google Scholar] [CrossRef]
- Lawrence, A.J.; Jarrott, B. Neurochemical modulation of cardiovascular control in the nucleus tractus solitarius. Prog. Neurobiol. 1996, 48, 21–53. [Google Scholar] [CrossRef]
- Murase, S.; Inui, K.; Nosaka, S. Baroreceptor inhibition of the locus coeruleus noradrenergic neurons. Neuroscience 1994, 61, 635–643. [Google Scholar] [CrossRef]
- Morrison, S.F. RVLM and raphe differentially regulate sympathetic outflows to splanchnic and brown adipose tissue. Am. J. Physiol. 1999, 276, R962–R973. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.A.; O’Leary, D.S.; Ly, C.; Smith, S.A.; Minic, Z. Development of a decerebrate model for investigating mechanisms mediating viscero-sympathetic reflexes in the spinalized rat. Am. J. Physiol.-Heart Circ. Physiol. 2019, 316, H1332–H1340. [Google Scholar] [CrossRef] [PubMed]
- Mayorov, D.N.; Adams, M.A.; Krassioukov, A.V. Telemetric blood pressure monitoring in conscious rats before and after compression injury of spinal cord. J. Neurotrauma 2001, 18, 727. [Google Scholar] [CrossRef] [PubMed]
- Osborn, J.W.; Taylor, R.F.; Schramm, L.P. Determinants of arterial pressure after chronic spinal transection in rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1989, 256, R666–R673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, K.; Yardley, C.; Weaver, L. Evidence for descending tonic inhibition specifically affecting sympathetic pathways to the kidney in rats. J. Physiol. 1991, 434, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, M.; Tagawa, T.; Polson, J.; Cavanagh, S.-J.; Dampney, R. Descending pathways mediating cardiovascular response from dorsomedial hypothalamic nucleus. Am. J. Physiol.-Heart Circ. Physiol. 2001, 280, H2891–H2901. [Google Scholar] [CrossRef]
- Cao, W.H.; Fan, W.; Morrison, S.F. Medullary pathways mediating specific sympathetic responses to activation of dorsomedial hypothalamus. Neuroscience 2004, 126, 229–240. [Google Scholar] [CrossRef]
- Martin, D.S.; Segura, T.; Haywood, J.R. Cardiovascular responses to bicuculline in the paraventricular nucleus of the rat. Hypertension 1991, 18, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.R.; Kupiers, N.T.; Ray, C.A. Neurovascular responses to mental stress. J. Physiol. 2005, 564, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.R.; Ray, C.A. Sympathetic neural responses to mental stress: Responders, nonresponders and sex differences. Am. J. Physiol.-Heart Circ. Physiol. 2009, 296, H847–H853. [Google Scholar] [CrossRef] [Green Version]
- Coote, J.H.; Dodds, W.N. The baroreceptor reflex and the cardiovascular changes associated with sustained muscular contraction in the cat. Pflügers Arch. 1976, 363, 167–173. [Google Scholar] [CrossRef]
- Sheriff, D.; O’Leary, D.S.; Scher, A.M.; Rowell, L.B. Baroreflex attenuates pressor response to graded muscle ischemia in exercising dogs. Am. J. Physiol.-Heart Circ. Physiol. 1990, 258, H305–H310. [Google Scholar] [CrossRef] [PubMed]
- Hatton, D.C.; Brooks, V.; Qi, Y.; McCarron, D.A. Cardiovascular response to stress: Baroreflex resetting and hemodynamics. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 1997, 272, R1588–R1594. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.B.; Young, B.E.; Fadel, P.J. Arterial baroreflex resetting during exercise in humans: Underlying signaling mechanisms. Exerc. Sport Sci. Rev. 2019, 47, 129–141. [Google Scholar] [CrossRef]
- Durocher, J.J.; Klein, J.C.; Carter, J.R. Attenuation of sympathetic baroreflex sensitivity during the onset of acute mental stress in humans. Am. J. Physiol.-Heart Circ. Physiol. 2011, 300, H1788–H1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terreberry, R.R.; Neafsey, E.J. Rat medial frontal cortex: A visceral motor region with a direct projection to the solitary nucleus. Brain Res. 1983, 278, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Terreberry, R.R.; Neafsey, E.J. The rat medial frontal cortex projects directly to autonomic regions of the brainstem. Brain Res. Bull. 1987, 19, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Macefield, V.G.; Henderson, L.A. “Real-time” imaging of cortical and subcortical sites of cardiovascular control: Concurrent recordings of sympathetic nerve activity and fMRI in awake subjects. J. Neurophysiol. 2016, 116, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, J.K.; Wong, S.W.; Cechetto, D.F. Cortical circuitry associated with reflex cardiovascular control in humans: Does the cortical autonomic network “speak” or “listen” during cardiovascular arousal. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 1375–1384. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, A.W.; Cohen, J.D.; Stenger, V.A.; Carter, C.S. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 2000, 288, 1835–1838. [Google Scholar] [CrossRef] [Green Version]
- Fechir, M.; Gamer, M.; Blasius, I.; Bauermann, T.; Breimhorst, M.; Schlindwein, P.; Schlereth, T.; Birklein, F. Functional imaging of sympathetic activation during mental stress. Neuroimage 2010, 50, 847–854. [Google Scholar] [CrossRef]
- Ruggiero, D.A.; Mraovitch, S.; Granata, A.R.; Anwar, M.; Reis, D.J. A role of insular cortex in cardiovascular function. J. Comp. Neurol. 1987, 257, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Saha, S. Role of the central nucleus of the amygdala in the control of blood pressure: Descending pathways to medullary cardiovascular nuclei. Clin. Exp. Pharmacol. Physiol. 2005, 32, 450–456. [Google Scholar] [CrossRef]
- Chau, D.; Johns, D.G.; Schramm, L.P. Ongoing and stimulus-evoked activity of sympathetically correlated neurons in the intermediate zone and dorsal horn of acutely spinalized rats. J. Neurophysiol. 2000, 83, 2699–2707. [Google Scholar] [CrossRef]
- Barman, S.M.; Gebber, G.L. Spinal interneurons with sympathetic nerve-related activity. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1984, 247, R761–R767. [Google Scholar] [CrossRef] [PubMed]
- Sherrington, C.; Laslett, E. Observations on some spinal reflexes and the interconnection of spinal segments. J. Physiol. 1903, 29, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, F.R. Viscero-motor reflexes. II. Am. J. Physiol.-Leg. Content 1924, 71, 84–89. [Google Scholar] [CrossRef]
- Miller, F.R.; Waud, R. Viscero-motor reflexes. IV. Am. J. Physiol.-Leg. Content 1925, 73, 329–340. [Google Scholar] [CrossRef]
- Downman, C.; McSwiney, B. Reflexes elicited by visceral stimulation in the acute spinal animal. J. Physiol. 1946, 105, 80. [Google Scholar] [CrossRef] [Green Version]
- Minic, Z.; O’Leary, D.S.; Reynolds, C.A. Spinal reflex control of arterial blood pressure: The role of TRP channels and their endogenous eicosanoid modulators. Front. Physiol. 2022, 13, 838175. [Google Scholar] [CrossRef]
- Jankowska, E.; Padel, Y.; Tanaka, R. Disynaptic inhibition of spinal motoneurones from the motor cortex in the monkey. J. Physiol. 1976, 258, 467–487. [Google Scholar] [CrossRef]
- Triggs, W.J.; Macdonell, R.A.; Cros, D.; Chiappa, K.H.; Shahani, B.T.; Day, B.J. Motor inhibition and excitation are independent effects of magnetic cortical stimulation. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1992, 32, 345–351. [Google Scholar] [CrossRef]
- Agarwal, S.; Gelsema, A.; Calaresu, F. Neurons in rostral VLM are inhibited by chemical stimulation of caudal VLM in rats. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 1989, 257, R265–R270. [Google Scholar] [CrossRef]
- Willette, R.; Punnen, S.; Krieger, A.; Sapru, H. Interdependence of rostral and caudal ventrolateral medullary areas in the control of blood pressure. Brain Res. 1984, 321, 169–174. [Google Scholar] [CrossRef]
- McCall, R.B. GABA-mediated inhibition of sympathoexcitatory neurons by midline medullary stimulation. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1988, 255, R605–R615. [Google Scholar] [CrossRef]
- Llewellyn-Smith, I.J. GABA in the control of sympathetic preganglionic neurons. Clin. Exp. Pharmacol. Physiol. 2002, 29, 507–513. [Google Scholar] [CrossRef]
- McCall, R.B.; Harris, L.T. Sympathetic alterations after midline medullary raphe lesions. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1987, 253, R91–R100. [Google Scholar] [CrossRef]
- Pilowsky, P.; West, M.; Chalmers, J. Renal sympathetic nerve responses to stimulation, inhibition and destruction of the ventrolateral medulla in the rabbit. Neurosci. Lett. 1985, 60, 51–55. [Google Scholar] [CrossRef]
- Loewy, A. Raphe pallidus and raphe obscurus projections to the intermediolateral cell column in the rat. Brain Res. 1981, 222, 129–133. [Google Scholar] [CrossRef]
- Loewy, A.; McKellar, S. Serotonergic projections from the ventral medulla to the intermediolateral cell column in the rat. Brain Res. 1981, 211, 146–152. [Google Scholar] [CrossRef]
- Fleetwood-Walker, S.M.; Coote, J.H.; Gilbey, M.P. Identification of spinally projecting neurones in the A1 catecholamine cell group of the ventrolateral medulla. Brain Res. 1983, 273, 25–33. [Google Scholar] [CrossRef]
- Dembowsky, K.; Czachurski, J.; Amendt, K.; Seller, H. Tonic descending inhibition of the spinal somato-sympathetic reflex from the lower brain stem. J. Auton. Nerv. Syst. 1980, 2, 157–182. [Google Scholar] [CrossRef]
- Müller, U.W.; Dembowsky, K.; Czachurski, J.; Seller, H. Tonic descending inhibition of the spinal cardio-sympathetic reflex in the cat. J. Auton. Nerv. Syst. 1988, 23, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Deuchars, S.A.; Milligan, C.J.; Stornetta, R.L.; Deuchars, J. GABAergic neurons in the central region of the spinal cord: A novel substrate for sympathetic inhibition. J. Neurosci. 2005, 25, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Bennett, G.J. Update on the neurophysiology of pain transmission and modulation: Focus on the NMDA-receptor. J. Pain Symptom Manag. 2000, 19, 2–6. [Google Scholar] [CrossRef]
- Sandkühler, J.; Bromm, B.; Gebhart, G.F. Nervous System Plasticity and Chronic Pain; Elsevier Science Limited: Amsterdam, The Netherlands, 2000; Volume 129. [Google Scholar]
- Craig, A.D. Pain mechanisms: Labeled lines versus convergence in central processing. Annu. Rev. Neurosci. 2003, 26, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, J.; Tracey, I. From nociception to pain perception: Imaging the spinal and supraspinal pathways. J. Anat. 2005, 207, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, M.C.; Duncan, G.H.; Hofbauer, R.K.; Ha, B.; Chen, J.I.; Carrier, B. Pain perception: Is there a role for primary somatosensory cortex? Proc. Natl. Acad. Sci. USA 1999, 96, 7705–7709. [Google Scholar] [CrossRef] [Green Version]
- Rainville, P.; Duncan, G.H.; Price, D.D.; Carrier, B.; Bushnell, M.C. Pain affect encoded in human anterior cingulate but not somatosensory cortex. Science 1997, 277, 968–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coghill, R.C.; Sang, C.N.; Maisog, J.M.; Iadarola, M.J. Pain intensity processing within the human brain: A bilateral, distributed mechanism. J. Neurophysiol. 1999, 82, 1934–1943. [Google Scholar] [CrossRef]
- Strack, A.; Sawyer, W.; Hughes, J.; Platt, K.; Loewy, A. A general pattern of CNS innervation of the sympathetic outflow demonstrated by transneuronal pseudorabies viral infections. Brain Res. 1989, 491, 156–162. [Google Scholar] [CrossRef]
- Yamashita, H.; Inenaga, K.; Koizumi, K. Possible projections from regions of paraventricular and supraoptic nuclei to the spinal cord: Electrophysiological studies. Brain Res. 1984, 296, 373–378. [Google Scholar] [CrossRef]
- Badoer, E. Hypothalamic paraventricular nucleus and cardiovascular regulation. Clin. Exp. Pharmacol. Physiol. 2001, 28, 95–99. [Google Scholar] [CrossRef]
- Ramchandra, R.; Hood, S.G.; Frithiof, R.; McKinley, M.J.; May, C.N. The role of the paraventricular nucleus of the hypothalamus in the regulation of cardiac and renal sympathetic nerve activity in conscious normal and heart failure sheep. J. Physiol. 2013, 591, 93–107. [Google Scholar] [CrossRef]
- Cechetto, D.F.; Saper, C.B. Neurochemical organization of the hypothalamic projection to the spinal cord in the rat. J. Comp. Neurol. 1988, 272, 579–604. [Google Scholar] [CrossRef]
- Elias, C.F.; Lee, C.; Kelly, J.; Aschkenasi, C.; Ahima, R.S.; Couceyro, P.R.; Kuhar, M.J.; Saper, C.B.; Elmquist, J.K. Leptin activates hypothalamic CART neurons projecting to the spinal cord. Neuron 1998, 21, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Sapru, H.N. Role of the hypothalamic arcuate nucleus in cardiovascular regulation. Auton. Neurosci. 2013, 175, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Fulwiler, C.E.; Saper, C.B. Subnuclear organization of the efferent connections of the parabrachial nucleus in the rat. Brain Res. Rev. 1984, 7, 229–259. [Google Scholar] [CrossRef]
- Saleh, T.M.; Connell, B.J.; Cribb, A.E. Estrogen in the parabrachial nucleus attenuates the sympathoexcitation following MCAO in male rats. Brain Res. 2005, 1066, 187–195. [Google Scholar] [CrossRef]
- Hayward, L.F.; Felder, R.B. Lateral parabrachial nucleus modulates baroreflex regulation of sympathetic nerve activity. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1998, 274, R1274–R1282. [Google Scholar] [CrossRef]
- Mtui, E.P.; Anwar, M.; Gomez, R.; Reis, D.J.; Ruggiero, D.A. Projections from the nucleus tractus solitarii to the spinal cord. J. Comp. Neurol. 1993, 337, 231–252. [Google Scholar] [CrossRef]
- Stornetta, R.L.; Guyenet, P.G. Distribution of glutamic acid decarboxylase mRNA-containing neurons in rat medulla projecting to thoracic spinal cord in relation to monoaminergic brainstem neurons. J. Comp. Neurol. 1999, 407, 367–380. [Google Scholar] [CrossRef]
- Loewy, A.; Burton, H. Nuclei of the solitary tract: Efferent projections to the lower brain stem and spinal cord of the cat. J. Comp. Neurol. 1978, 181, 421–449. [Google Scholar] [CrossRef] [PubMed]
- Madden, C. Consumption of a high fat diet inhibits sympathetic outflow to brown adipose tissue (BAT) via vagal afferent activation of neurons in the Nucleus Tractus Solitarius (NTS). Auton. Neurosci. Basic Clin. 2015, 192, 13. [Google Scholar] [CrossRef]
- Haselton, J.R.; Winters, R.W.; Liskowsky, D.R.; Haselton, C.L.; McCabe, P.M.; Schneiderman, N. Anatomical and functional connections of neurons of the rostral medullary raphe of the rabbit. Brain Res. 1988, 453, 176–182. [Google Scholar] [CrossRef]
- Arami, M.K.; Komaki, A.; Gharibzadeh, S. Contribution of nucleus raphe magnus to thermoregulation. Physiol. Pharmacol. 2020, 24, 165–173. [Google Scholar] [CrossRef]
- Leung, C.G.; Mason, P. Spectral analysis of arterial blood pressure and raphe magnus neuronal activity in anesthetized rats. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 1996, 271, R483–R489. [Google Scholar] [CrossRef]
- Blessing, W.; Nalivaiko, E. Raphe magnus/pallidus neurons regulate tail but not mesenteric arterial blood flow in rats. Neuroscience 2001, 105, 923–929. [Google Scholar] [CrossRef]
- Babic, T.; Ciriello, J. Medullary and spinal cord projections from cardiovascular responsive sites in the rostral ventromedial medulla. J. Comp. Neurol. 2004, 469, 391–412. [Google Scholar] [CrossRef]
- Elam, M.; Svensson, T.H.; Thoren, P. Differentiated cardiovascular afferent regulation of locus coeruleus neurons and sympathetic nerves. Brain Res. 1985, 358, 77–84. [Google Scholar] [CrossRef]
- Bruinstroop, E.; Cano, G.; Vanderhorst, V.G.; Cavalcante, J.C.; Wirth, J.; Sena-Esteves, M.; Saper, C.B. Spinal projections of the A5, A6 (locus coeruleus), and A7 noradrenergic cell groups in rats. J. Comp. Neurol. 2012, 520, 1985–2001. [Google Scholar] [CrossRef] [Green Version]
- Miyawaki, T.; Kawamura, H.; Komatsu, K.; Yasugi, T. Chemical stimulation of the locus coeruleus: Inhibitory effects on hemodynamics and renal sympathetic nerve activity. Brain Res. 1991, 568, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Aicher, S.A.; Reis, D.J.; Nicolae, R.; Milner, T.A. Monosynaptic projections from the medullary gigantocellular reticular formation to sympathetic preganglionic neurons in the thoracic spinal cord. J. Comp. Neurol. 1995, 363, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Korkola, M.L.; Weaver, L.C. Role of dorsal medullary reticular formation in maintenance of vasomotor tone in rats. J. Auton. Nerv. Syst. 1994, 46, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Sesa-Ashton, G.; Wong, R.; McCarthy, B.; Datta, S.; Henderson, L.A.; Dawood, T.; Macefield, V.G. Stimulation of the dorsolateral prefrontal cortex modulates muscle sympathetic nerve activity and blood pressure in humans. Cereb. Cortex Commun. 2022, 3, tgac017. [Google Scholar] [CrossRef]
- James, C.; Macefield, V.G.; Henderson, L.A. Real-time imaging of cortical and subcortical control of muscle sympathetic nerve activity in awake human subjects. Neuroimage 2013, 70, 59–65. [Google Scholar] [CrossRef]
- Van Eden, C.G.; Buijs, R.M. Functional neuroanatomy of the prefrontal cortex: Autonomic interactions. Prog. Brain Res. 2000, 126, 49–62. [Google Scholar]
- Miller, M. The origin of corticospinal projection neurons in rat. Exp. Brain Res. 1987, 67, 339–351. [Google Scholar] [CrossRef]
- Hurley, K.M.; Herbert, H.; Moga, M.M.; Saper, C.B. Efferent projections of the infralimbic cortex of the rat. J. Comp. Neurol. 1991, 308, 249–276. [Google Scholar] [CrossRef]
- Kuroda, R.; Kawao, N.; Yoshimura, H.; Umeda, W.; Takemura, M.; Shigenaga, Y.; Kawabata, A. Secondary somatosensory cortex stimulation facilitates the antinociceptive effect of the NO synthase inhibitor through suppression of spinal nociceptive neurons in the rat. Brain Res. 2001, 903, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Bruehl, S.; Carlson, C.R.; McCubbin, J.A. The relationship between pain sensitivity and blood pressure in normotensives. Pain 1992, 48, 463–467. [Google Scholar] [CrossRef]
- Bruehl, S.; Chung, O.Y. Interactions between the cardiovascular and pain regulatory systems: An updated review of mechanisms and possible alterations in chronic pain. Neurosci. Biobehav. Rev. 2004, 28, 395–414. [Google Scholar] [CrossRef]
- McCubbin, J.A.; Bruehl, S. Do endogenous opioids mediate the relationship between blood pressure and pain sensitivity in normotensives? Pain 1994, 57, 63–67. [Google Scholar] [CrossRef]
- Fillingim, R.B.; Maixner, W. The influence of resting blood pressure and gender on pain responses. Psychosom. Med. 1996, 58, 326–332. [Google Scholar] [CrossRef]
- Pfleeger, M.; Straneva, P.A.; Fillingim, R.B.; Maixner, W.; Girdler, S.S. Menstrual cycle, blood pressure and ischemic pain sensitivity in women: A preliminary investigation. Int. J. Psychophysiol. 1997, 27, 161–166. [Google Scholar] [CrossRef]
- Fillingim, R.B.; Maixner, W.; Bunting, S.; Silva, S. Resting blood pressure and thermal pain responses among females: Effects on pain unpleasantness but not pain intensity. Int. J. Psychophysiol. 1998, 30, 313–318. [Google Scholar] [CrossRef]
- Myers, C.D.; Robinson, M.E.; Riley III, J.L.; Sheffield, D. Sex, gender, and blood pressure: Contributions to experimental pain report. Psychosom. Med. 2001, 63, 545–550. [Google Scholar] [CrossRef]
- Al’Absi, M.; Buchanan, T.; Lovallo, W.R. Pain perception and cardiovascular responses in men with positive parental history for hypertension. Psychophysiology 1996, 33, 655–661. [Google Scholar] [CrossRef]
- Al’Absi, M.; Buchanan, T.W.; Marrero, A.; Lovallo, W.R. Sex differences in pain perception and cardiovascular responses in persons with parental history for hypertension. Pain 1999, 83, 331–338. [Google Scholar] [CrossRef]
- al’Absi, M.; Petersen, K.L.; Wittmers, L.E. Blood pressure but not parental history for hypertension predicts pain perception in women. Pain 2000, 88, 61–68. [Google Scholar] [CrossRef]
- al’Absi, M.; Petersen, K.L.; Wittmers, L.E. Adrenocortical and hemodynamic predictors of pain perception in men and women. Pain 2002, 96, 197–204. [Google Scholar] [CrossRef]
- Randich, A.; Maixner, W. Interactions between cardiovascular and pain regulatory systems. Neurosci. Biobehav. Rev. 1984, 8, 343–367. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanimoto, T.; Ojima, K.; Matsumoto, S. Suppressive effect of vagal afferents on the activity of the trigeminal spinal neurons related to the jaw-opening reflex in rats: Involvement of the endogenous opioid system. Brain Res. Bull. 1998, 47, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Bossut, D.; Maixner, W. Effects of cardiac vagal afferent electrostimulation on the responses of trigerninal and trigeminothalamic neurons to noxious orofacial stimulation. Pain 1996, 65, 101–109. [Google Scholar] [CrossRef]
- Dworkin, B.; Filewich, R.; Miller, N.; Craigmyle, N.; Pickering, T. Baroreceptor activation reduces reactivity to noxious stimulation: Implications for hypertension. Science 1979, 205, 1299–1301. [Google Scholar] [CrossRef]
- Thurston, C.L.; Randich, A. Acute increases in arterial blood pressure produced by occlusion of the abdominal aorta induces antinociception: Peripheral and central substrates. Brain Res. 1990, 519, 12–22. [Google Scholar] [CrossRef]
- Maixner, W.; Touw, K.B.; Brody, M.J.; Gebhart, G.F. Factors influencing the altered pain perception in the spontaneously hypertensive rat. Brain Res. 1982, 237, 137–145. [Google Scholar] [CrossRef]
- D’Antono, B.; Ditto, B.; Sita, A.; Miller, S.B. Cardiopulmonary baroreflex stimulation and blood pressure-related hypoalgesia. Biol. Psychol. 2000, 53, 217–231. [Google Scholar] [CrossRef]
- Dworkin, B.R.; Elbert, T.; Rau, H.; Birbaumer, N.; Pauli, P.; Droste, C.; Brunia, C. Central effects of baroreceptor activation in humans: Attenuation of skeletal reflexes and pain perception. Proc. Natl. Acad. Sci. USA 1994, 91, 6329–6333. [Google Scholar] [CrossRef] [Green Version]
- Rau, H.; Brody, S.; Larbig, W.; Pauli, P.; Vöhringer, M.; Harsch, B.; Kröling, P.; Birbaumer, N. Effects of PRES baroreceptor stimulation on thermal and mechanical pain threshold in borderline hypertensives and normotensives. Psychophysiology 1994, 31, 480–485. [Google Scholar] [CrossRef]
- Angrilli, A.; Mini, A.; Mucha, R.F.; Rau, H. The influence of low blood pressure and baroreceptor activity on pain responses. Physiol. Behav. 1997, 62, 391–397. [Google Scholar] [CrossRef]
- Edwards, L.; McIntyre, D.; Carroll, D.; Ring, C.; France, C.R.; Martin, U. Effects of artificial and natural baroreceptor stimulation on nociceptive responding and pain. Psychophysiology 2003, 40, 762–769. [Google Scholar] [CrossRef]
- Edwards, L.; Ring, C.; McIntyre, D.; Carroll, D. Modulation of the human nociceptive flexion reflex across the cardiac cycle. Psychophysiology 2001, 38, 712–718. [Google Scholar] [CrossRef]
- Bruehl, S.; Chung, O.Y.; Jirjis, J.N.; Biridepalli, S. Prevalence of clinical hypertension in patients with chronic pain compared to nonpain general medical patients. Clin. J. Pain 2005, 21, 147–153. [Google Scholar] [CrossRef]
- Browne, T.J.; Hughes, D.I.; Dayas, C.V.; Callister, R.J.; Graham, B.A. Projection neuron axon collaterals in the dorsal horn: Placing a new player in spinal cord pain processing. Front. Physiol. 2020, 11, 560802. [Google Scholar] [CrossRef]
- Schramm, L.P. Spinal sympathetic interneurons: Their identification and roles after spinal cord injury. Prog. Brain Res. 2006, 152, 27–37. [Google Scholar]
- Clifton, G.L.; Ziegler, M.G.; Grossman, R.G. Circulating catecholamines and sympathetic activity after head injury. Neurosurgery 1981, 8, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Park, C.I.; Cho, S.R.; Lee, J.W.; Kim, E.J. Assessment of autonomic nervous system with analysis of heart rate variability in children with spastic cerebral palsy. Yonsei Med. J. 2002, 43, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kushki, A.; Drumm, E.; Mobarak, M.P.; Tanel, N.; Dupuis, A.; Chau, T.; Anagnostou, E. Investigating the autonomic nervous system response to anxiety in children with autism spectrum disorders. PLoS ONE 2013, 8, e59730. [Google Scholar] [CrossRef] [Green Version]
- Appelhans, B.M.; Luecken, L.J. Heart rate variability as an index of regulated emotional responding. Rev. Gen. Psychol. 2006, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Webb, S.; Adgey, A.; Pantridge, J. Autonomic disturbance at onset of acute myocardial infarction. Br. Med. J. 1972, 3, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, P.J.; Vanoli, E. Cardiac arrhythmias elicited by interaction between acute myocardial ischemia and sympathetic hyperactivity: A new experimental model for the study of antiarrhythmic drugs. J. Cardiovasc. Pharmacol. 1981, 3, 1251–1259. [Google Scholar] [CrossRef]
- Lyon, A.R.; Rees, P.S.; Prasad, S.; Poole-Wilson, P.A.; Harding, S.E. Stress (Takotsubo) cardiomyopathy—A novel pathophysiological hypothesis to explain catecholamine-induced acute myocardial stunning. Nat. Rev. Cardiol. 2008, 5, 22. [Google Scholar] [CrossRef]
- Esler, M.; Lambert, G.; Brunner-La Rocca, H.; Vaddadi, G.; Kaye, D. Sympathetic nerve activity and neurotransmitter release in humans: Translation from pathophysiology into clinical practice. Acta Physiol. Scand. 2003, 177, 275–284. [Google Scholar] [CrossRef]
- Schadt, J.C.; Ludbrook, J. Hemodynamic and neurohumoral responses to acute hypovolemia in conscious mammals. Am. J. Physiol. -Heart Circ. Physiol. 1991, 260, H305–H318. [Google Scholar] [CrossRef]
- Evans, R.G.; Ventura, S.; Dampney, R.A.; Ludbrook, J. John Ludbrook APPS Symposium Neural Mechanisms In The Cardiovascular Responses To Acute Central Hypovolaemia. Clin. Exp. Pharmacol. Physiol. 2001, 28, 479–487. [Google Scholar] [CrossRef]
- Carlson, C.R.; Okeson, J.P.; Falace, D.A.; Nitz, A.J.; Curran, S.L.; Anderson, D. Comparison of psychologic and physiologic functioning between patients with masticatory muscle pain and matched controls. J. Orofac. Pain 1993, 7. [Google Scholar]
- Peters, M.L.; Schmidt, A.J. Psychophysiological responses to repeated acute pain stimulation in chronic low back pain patients. J. Psychosom. Res. 1991, 35, 59–74. [Google Scholar] [CrossRef]
- Collins, G.; Cohen, M.; Naliboff, B.; Schandler, S. Comparative analysis of paraspinal and frontalis EMG, heart rate and skin conductance in chronic low back pain patients and normals to various postures and stress. Scand. J. Rehabil. Med. 1982, 14, 39–46. [Google Scholar]
- Schweinhardt, P.; Bushnell, M.C. Pain imaging in health and disease—How far have we come? J. Clin. Investig. 2010, 120, 3788–3797. [Google Scholar] [CrossRef]
- Baliki, M.N.; Chialvo, D.R.; Geha, P.Y.; Levy, R.M.; Harden, R.N.; Parrish, T.B.; Apkarian, A.V. Chronic pain and the emotional brain: Specific brain activity associated with spontaneous fluctuations of intensity of chronic back pain. J. Neurosci. 2006, 26, 12165–12173. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, J.A.; Baliki, M.N.; Huang, L.; Baria, A.T.; Torbey, S.; Hermann, K.M.; Schnitzer, T.J.; Apkarian, A.V. Shape shifting pain: Chronification of back pain shifts brain representation from nociceptive to emotional circuits. Brain 2013, 136, 2751–2768. [Google Scholar] [CrossRef] [Green Version]
- May, A. Chronic pain may change the structure of the brain. PAIN® 2008, 137, 7–15. [Google Scholar] [CrossRef]
- Hubbard, C.S.; Khan, S.A.; Keaser, M.L.; Mathur, V.A.; Goyal, M.; Seminowicz, D.A. Altered brain structure and function correlate with disease severity and pain catastrophizing in migraine patients. Eneuro 2014, 1. [Google Scholar] [CrossRef]
- Ihara, N.; Wakaizumi, K.; Nishimura, D.; Kato, J.; Yamada, T.; Suzuki, T.; Hashiguchi, S.; Terasawa, Y.; Kosugi, S.; Morisaki, H. Aberrant resting-state functional connectivity of the dorsolateral prefrontal cortex to the anterior insula and its association with fear avoidance belief in chronic neck pain patients. PLoS ONE 2019, 14, e0221023. [Google Scholar] [CrossRef] [Green Version]
- Seminowicz, D.A.; Moayedi, M. The dorsolateral prefrontal cortex in acute and chronic pain. J. Pain 2017, 18, 1027–1035. [Google Scholar] [CrossRef]
- Qi, X.; Cui, K.; Zhang, Y.; Wang, L.; Tong, J.; Sun, W.; Shao, S.; Wang, J.; Wang, C.; Sun, X.; et al. A nociceptive neuronal ensemble in the dorsomedial prefrontal cortex underlies pain chronicity. Cell Rep. 2022, 41, 111833. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- Holden, J.E.; Naleway, E. Microinjection of carbachol in the lateral hypothalamus produces opposing actions on nociception mediated by α1-and α2-adrenoceptors. Brain Res. 2001, 911, 27–36. [Google Scholar] [CrossRef]
- Karim, F.; Roerig, S.C. Differential effects of antisense oligodeoxynucleotides directed against Gzα and Goα on antinociception produced by spinal opioid and α2 adrenergic receptor agonists. Pain 2000, 87, 181–191. [Google Scholar] [CrossRef]
- Singewald, N.; Philippu, A. Involvement of biogenic amines and amino acids in the central regulation of cardiovascular homeostasis. Trends Pharmacol. Sci. 1996, 17, 356–363. [Google Scholar] [CrossRef]
- Eisenach, J.C.; De Kock, M.; Klimscha, W. α2-Adrenergic agonists for regional anesthesia: A clinical review of clonidine (1984–1995). J. Am. Soc. Anesthesiol. 1996, 85, 655–674. [Google Scholar] [CrossRef]
- Maze, M.; Tranquilli, W. Alpha-2 adrenoceptor agonists: Defining the role in clinical anesthesia. J. Am. Soc. Anesthesiol. 1991, 74, 581–605. [Google Scholar] [CrossRef]
- Pertovaara, A.; Auppila, T.; Jyväsjärvi, E.; Kalso, E. Involvement of supraspinal and spinal segmental alpha-2-adrenergic mechanisms in the medetomidine-induced antinociception. Neuroscience 1991, 44, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Ghelardini, C.; Galeotti, N.; Bartolini, A. Antinociception induced by amitriptyline and imipramine is mediated by α2A-adrenoceptors. Jpn. J. Pharmacol. 2000, 82, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, G.M.; Lyons, L.; Dickenson, A.H. α2-adrenoceptor antagonists enhance responses of dorsal horn neurones to formalin induced inflammation. Eur. J. Pharmacol. 1998, 347, 201–204. [Google Scholar] [CrossRef]
- Xu, M.; Kontinen, V.K.; Kalso, E. Endogenous noradrenergic tone controls symptoms of allodynia in the spinal nerve ligation model of neuropathic pain. Eur. J. Pharmacol. 1999, 366, 41–45. [Google Scholar] [CrossRef]
- Feldman, P.; Felder, R. α-Adrenergic influences on neuronal responses to visceral afferent input in the nucleus tractus solitarius. Neuropharmacology 1989, 28, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Brodie, M.S.; Proudfit, H.K. Antinociception induced by local injections of carbachol into the nucleus raphe magnus in rats: Alteration by intrathecal injection of monoaminergic antagonists. Brain Res. 1986, 371, 70–79. [Google Scholar] [CrossRef]
- Budai, D.; Harasawa, I.; Fields, H.L. Midbrain periaqueductal gray (PAG) inhibits nociceptive inputs to sacral dorsal horn nociceptive neurons through α2-adrenergic receptors. J. Neurophysiol. 1998, 80, 2244–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Feng, Y.; McAdoo, D.; Willis, W. Periaqueductal gray stimulation-induced inhibition of nociceptive dorsal horn neurons in rats is associated with the release of norepinephrine, serotonin, and amino acids. J. Pharmacol. Exp. Ther. 1999, 289, 868–876. [Google Scholar]
- Adair, J.R.; Hamilton, B.L.; Scappaticci, K.A.; Helke, C.J.; Gillis, R.A. Cardiovascular responses to electrical stimulation of the medullary raphe area of the cat. Brain Res. 1977, 128, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.C.; Shin, J.W.; Chimenti, P.C.; Loewy, A.D. Paraventricular hypothalamic nucleus: Axonal projections to the brainstem. J. Comp. Neurol. 2010, 518, 1460–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Kooy, D.; Koda, L.Y.; McGinty, J.F.; Gerfen, C.R.; Bloom, F.E. The organization of projections from the cortes, amygdala, and hypothalamus to the nucleus of the solitary tract in rat. J. Comp. Neurol. 1984, 224, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Michelini, L.C. The NTS and integration of cardiovascular control during exercise in normotensive and hypertensive individuals. Curr. Hypertens. Rep. 2007, 9, 214–221. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
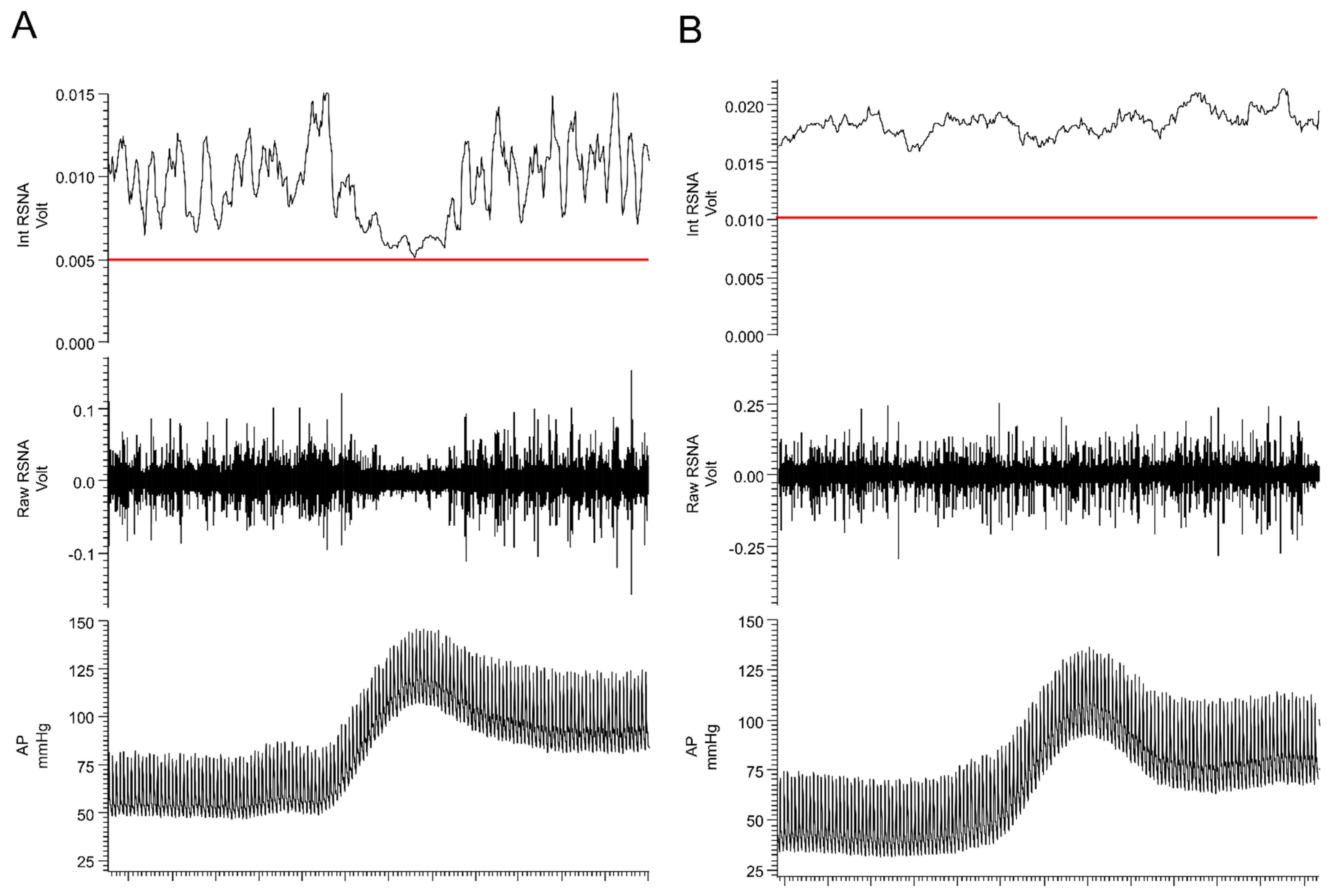{kind=link}
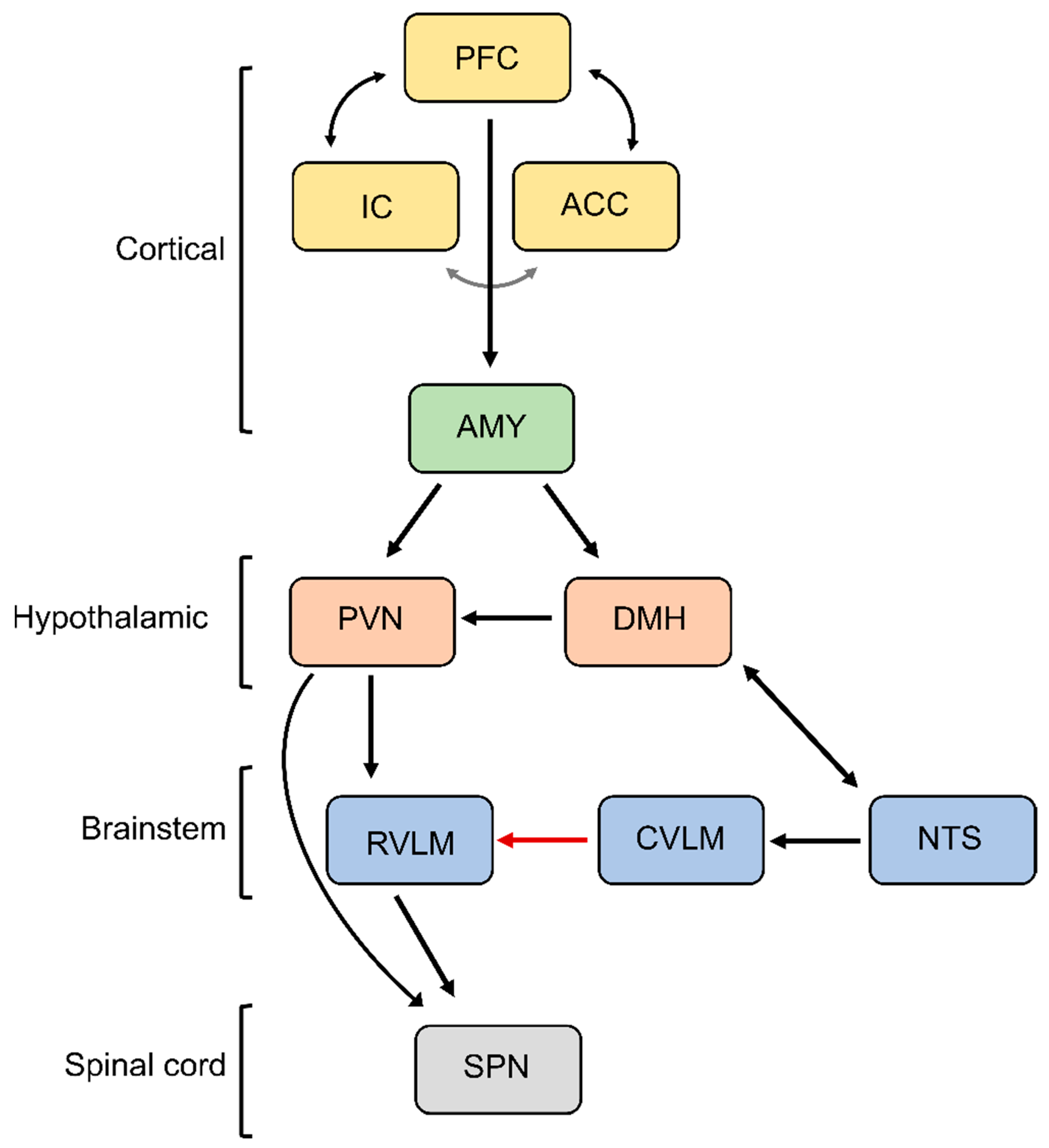{kind=link}
| Cardiovascular Outcome | References |
|---|---|
| Myocardial Infarction | [10,14,16,17,18,19,20,21,22] |
| Angina | [14,17,18,22,23,24] |
| Arrhythmia | [20,25] |
| Coronary Artery Disease | [14,22,25,26,27] |
| Hypertension | [20,25,28,29,30] |
| Stroke | [10,16,17,19,20,21,29] |
| Heart failure | [16,20] |
| Cardiovascular mortality | [8,16,21,22,30,31,32,33] |
| Structure | Effect(s) on DH Neurons | Effect(s) on IML Neurons | References |
|---|---|---|---|
| Hypothalamus | |||
| Paraventricular nucleus | Inhibition | Inhibition/Excitation | [113,114,115,116] |
| Arcuate nucleus | Inhibition/Facilitation | Inhibition/Excitation | [117,118,119] |
| Parabrachial nucleus | Inhibition | Excitation | [120,121,122] |
| Brainstem | |||
| Nucleus of the solitary tract | Inhibition | Inhibition | [123,124,125,126] |
| Raphe magnus/pallidus | Inhibition/Facilitation | Inhibition/Excitation | [127,128,129,130] |
| Rostroventromedial medulla | Facilitation | Excitation | [131] |
| Locus coeruleus | Inhibition | Inhibition | [132,133,134] |
| Medullary reticular formation | Inhibition/Facilitation | Inhibition/Excitation | [135,136] |
| Cerebral cortex | |||
| Frontal/parietal | Inhibition | Inhibition/Excitation | [137,138,139,140,141,142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reynolds, C.A.; Minic, Z. Chronic Pain-Associated Cardiovascular Disease: The Role of Sympathetic Nerve Activity. Int. J. Mol. Sci. 2023, 24, 5378. https://doi.org/10.3390/ijms24065378
Reynolds CA, Minic Z. Chronic Pain-Associated Cardiovascular Disease: The Role of Sympathetic Nerve Activity. International Journal of Molecular Sciences. 2023; 24(6):5378. https://doi.org/10.3390/ijms24065378
Chicago/Turabian StyleReynolds, Christian A., and Zeljka Minic. 2023. "Chronic Pain-Associated Cardiovascular Disease: The Role of Sympathetic Nerve Activity" International Journal of Molecular Sciences 24, no. 6: 5378. https://doi.org/10.3390/ijms24065378
APA StyleReynolds, C. A., & Minic, Z. (2023). Chronic Pain-Associated Cardiovascular Disease: The Role of Sympathetic Nerve Activity. International Journal of Molecular Sciences, 24(6), 5378. https://doi.org/10.3390/ijms24065378