
Intrinsic Allergenicity Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid and Hexaploid Wheats: Validation Using an Adjuvant-Free Mouse Model

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Validation of the Transdermal Sensitization and Oral Elicitation of Disease Mouse Model for Triticum monococcum Using Salt-Soluble Protein Extract
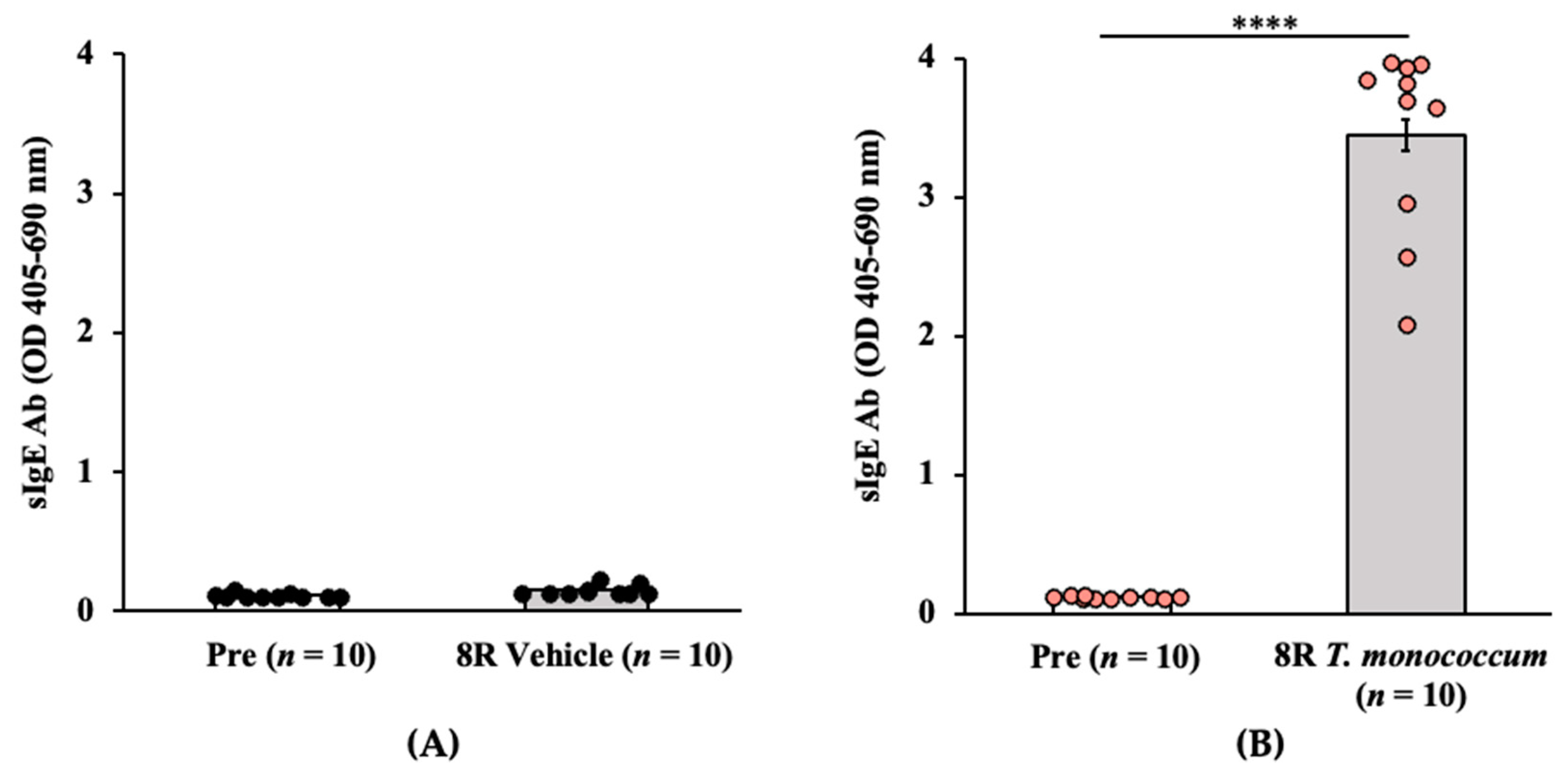
2.1.1. Transdermal Exposure to Salt-Soluble Protein Extract from T. monococcum Elicits Robust Specific-IgE Antibody Response in Balb/c Mice
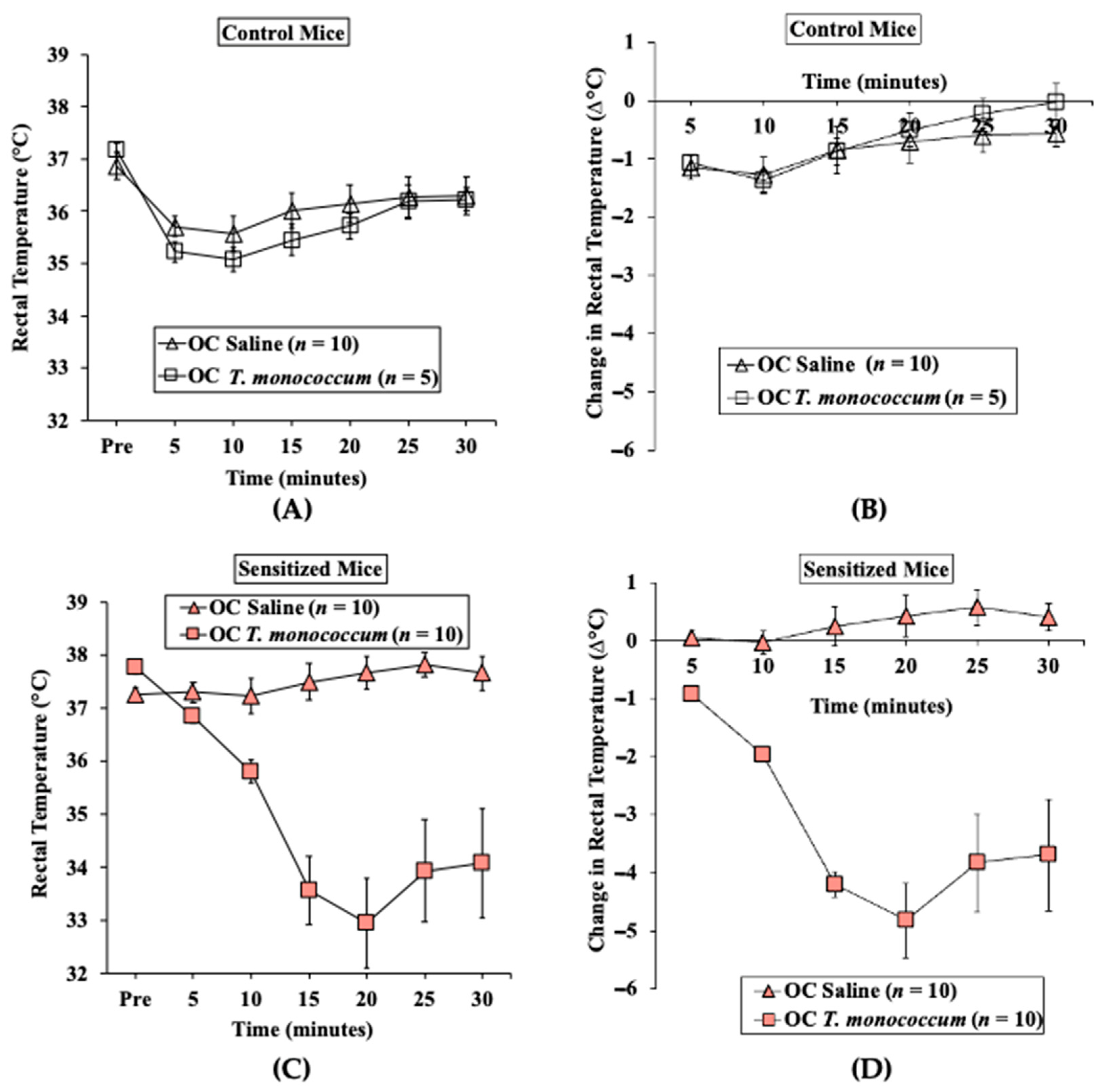
2.1.2. Oral Challenge with T. monococcum SSPE Elicits Hypothermic Shock Responses in Skin-Sensitized Mice
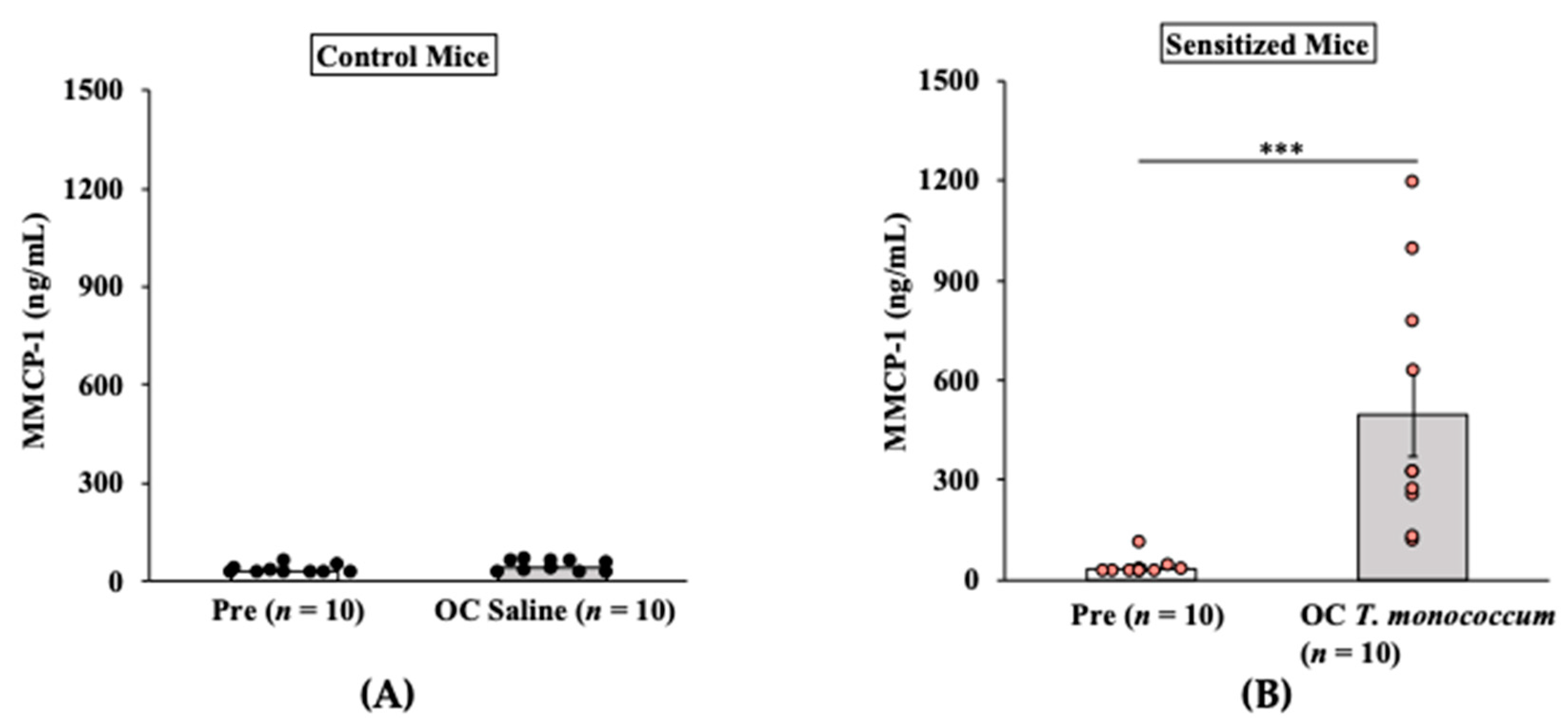
2.1.3. Oral Anaphylaxis Elicited by T. monococcum Is Associated with Robust Mucosal Mast Cell Response (MMCR) in Balb/c Mice
2.2. Validation of Transdermal Sensitization and Oral Elicitation of Disease Model Using Salt-Soluble Protein Extract from Triticum aestivum
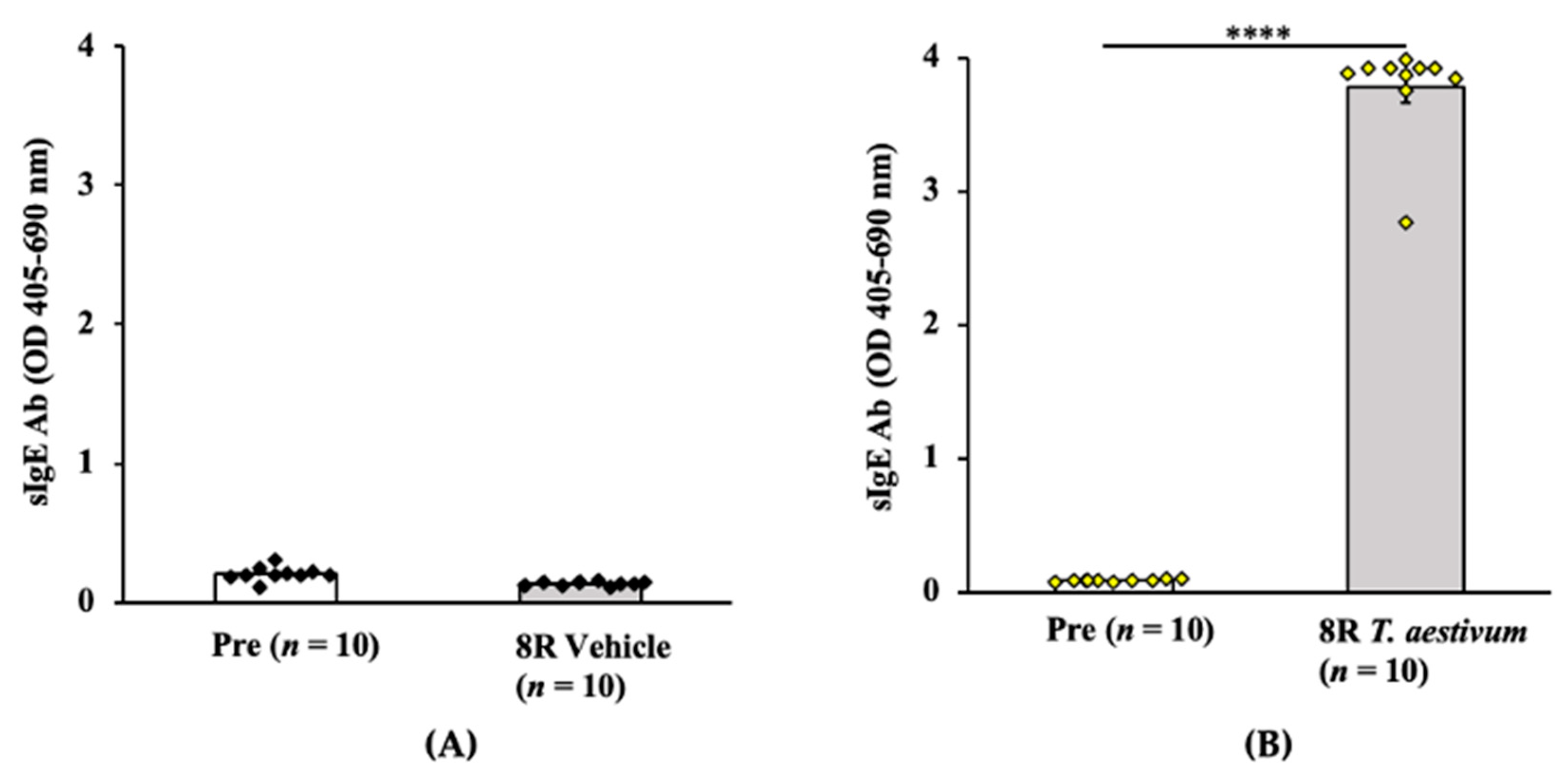
2.2.1. Transdermal Exposure to Salt-Soluble Protein Extract from T. aestivum Elicits Robust Specific-IgE Antibody Response in Balb/c Mice
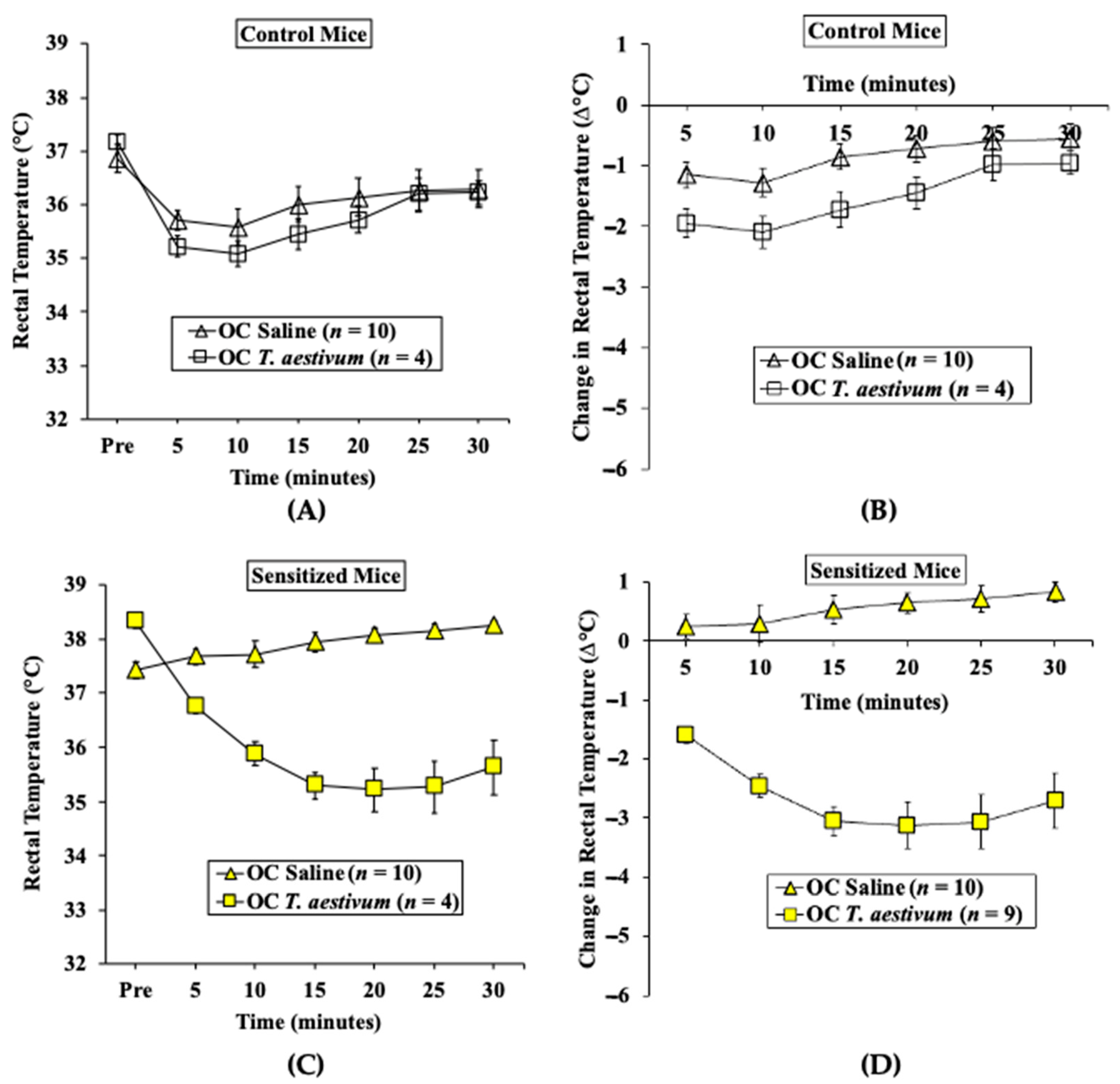
2.2.2. Oral Challenge with T. aestivum SSPE Elicits Hypothermic Shock Responses in Skin-Sensitized Mice
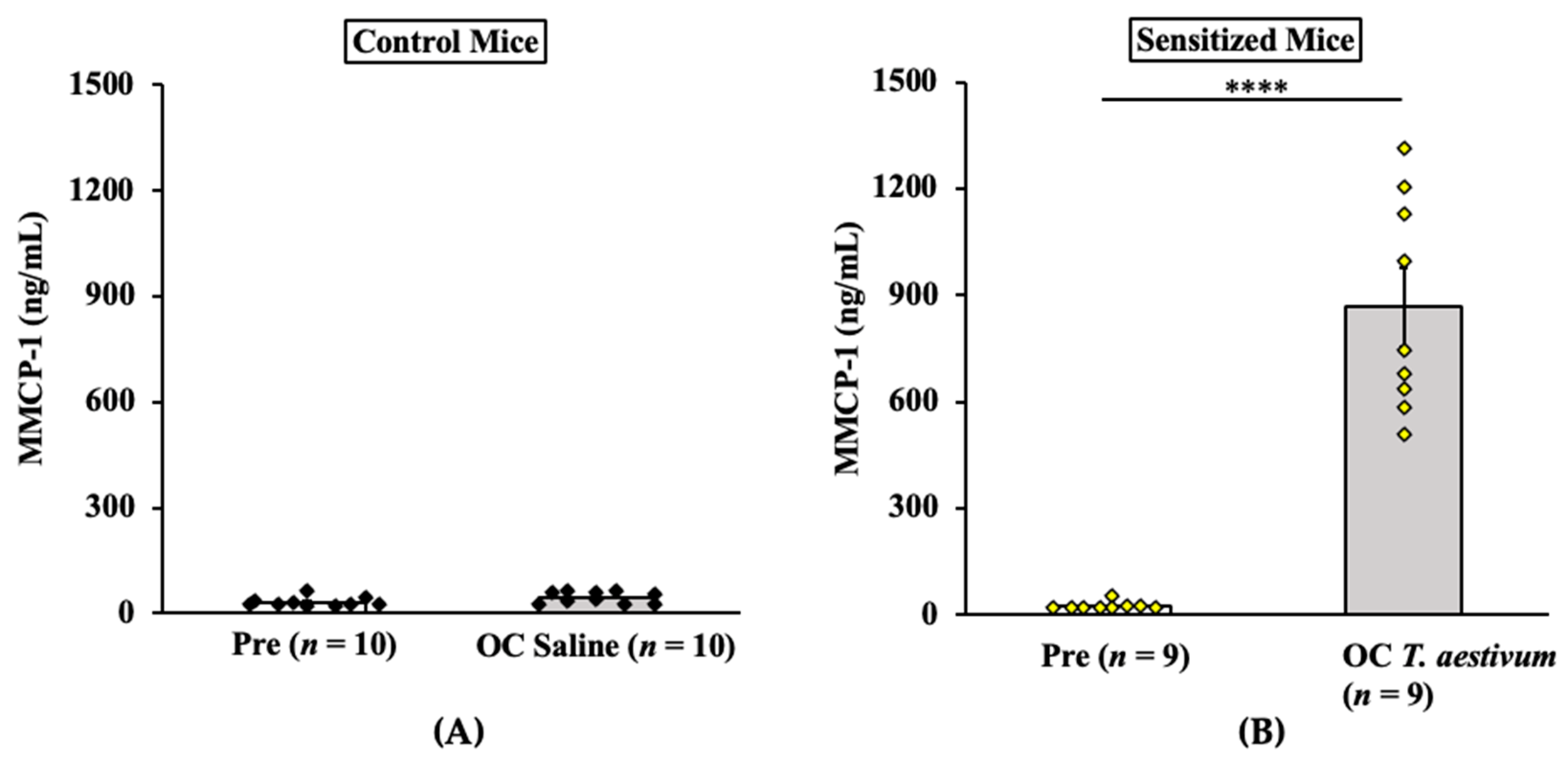
2.2.3. Oral Anaphylaxis Elicited by T. aestivum SSPE Is Associated with Robust Mucosal Mast Cell Response (MMCR) in Balb/c Mice
2.3. Validation of Transdermal Sensitization and Oral Elicitation of Disease Model Using Salt-Soluble Protein Extract from Aegilops tauschii
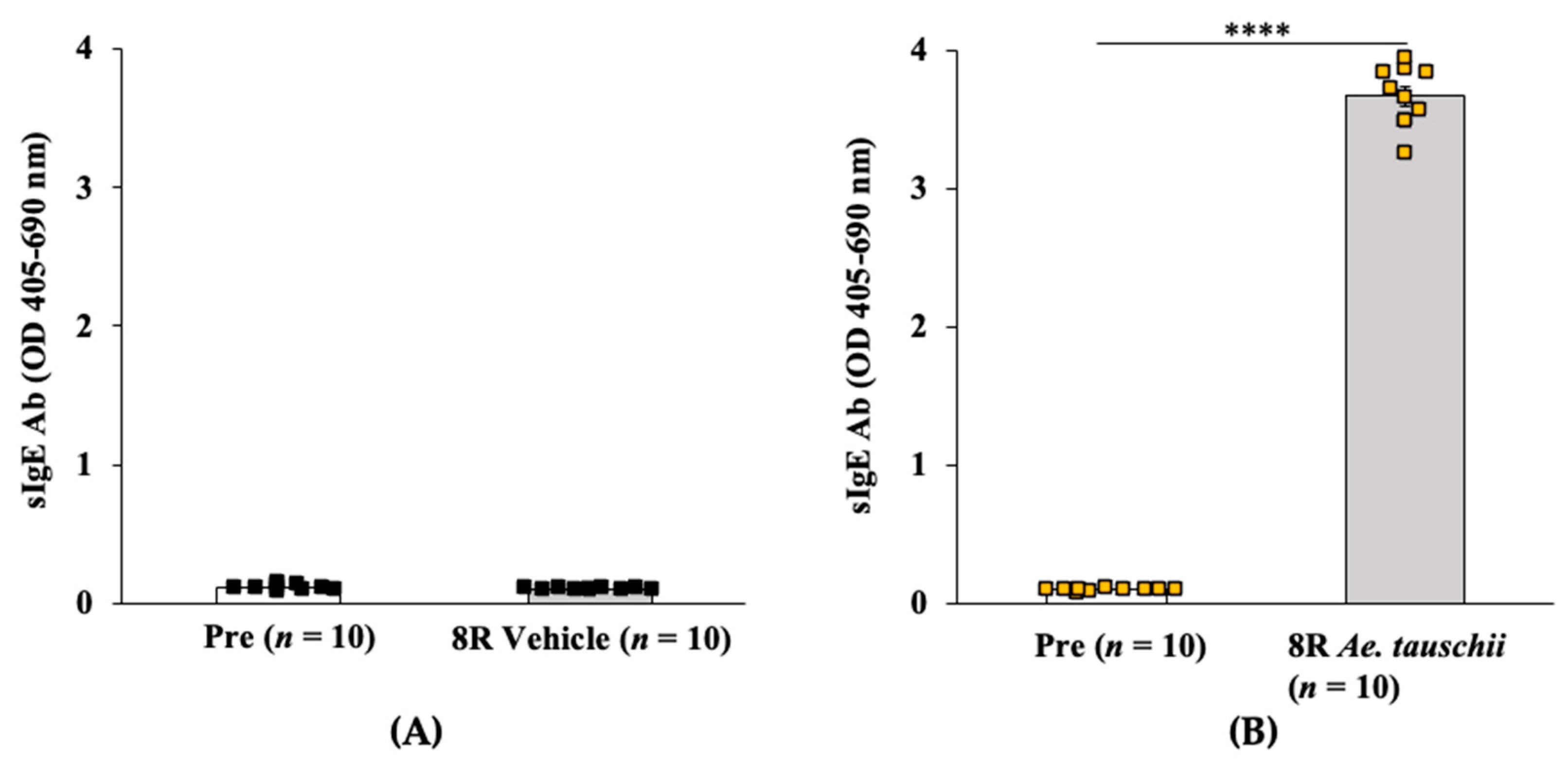
2.3.1. Transdermal Exposure to Salt-Soluble Protein Extract from Aegilops tauschii Elicits Robust Specific-IgE Antibody Response in Balb/c Mice
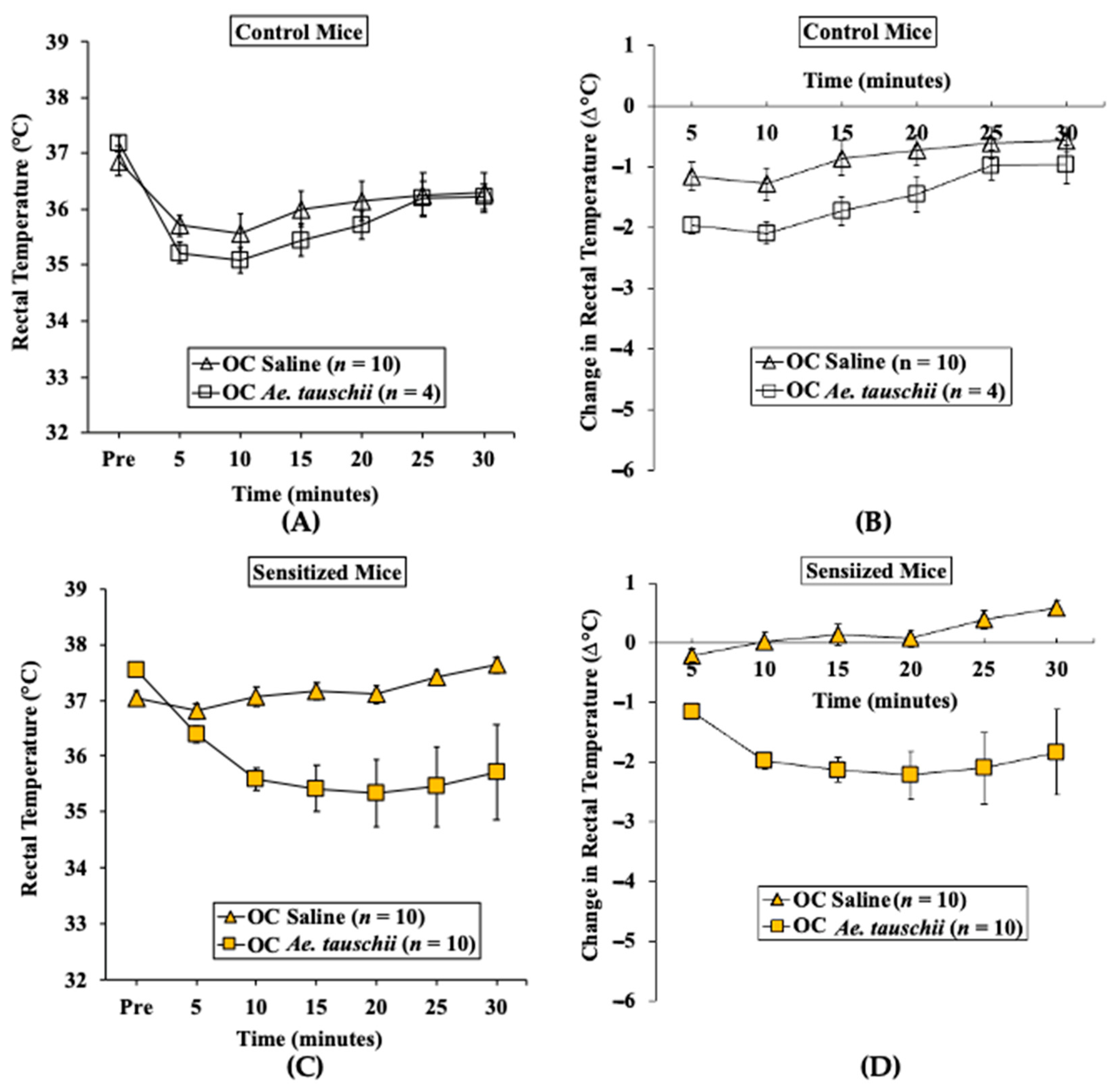
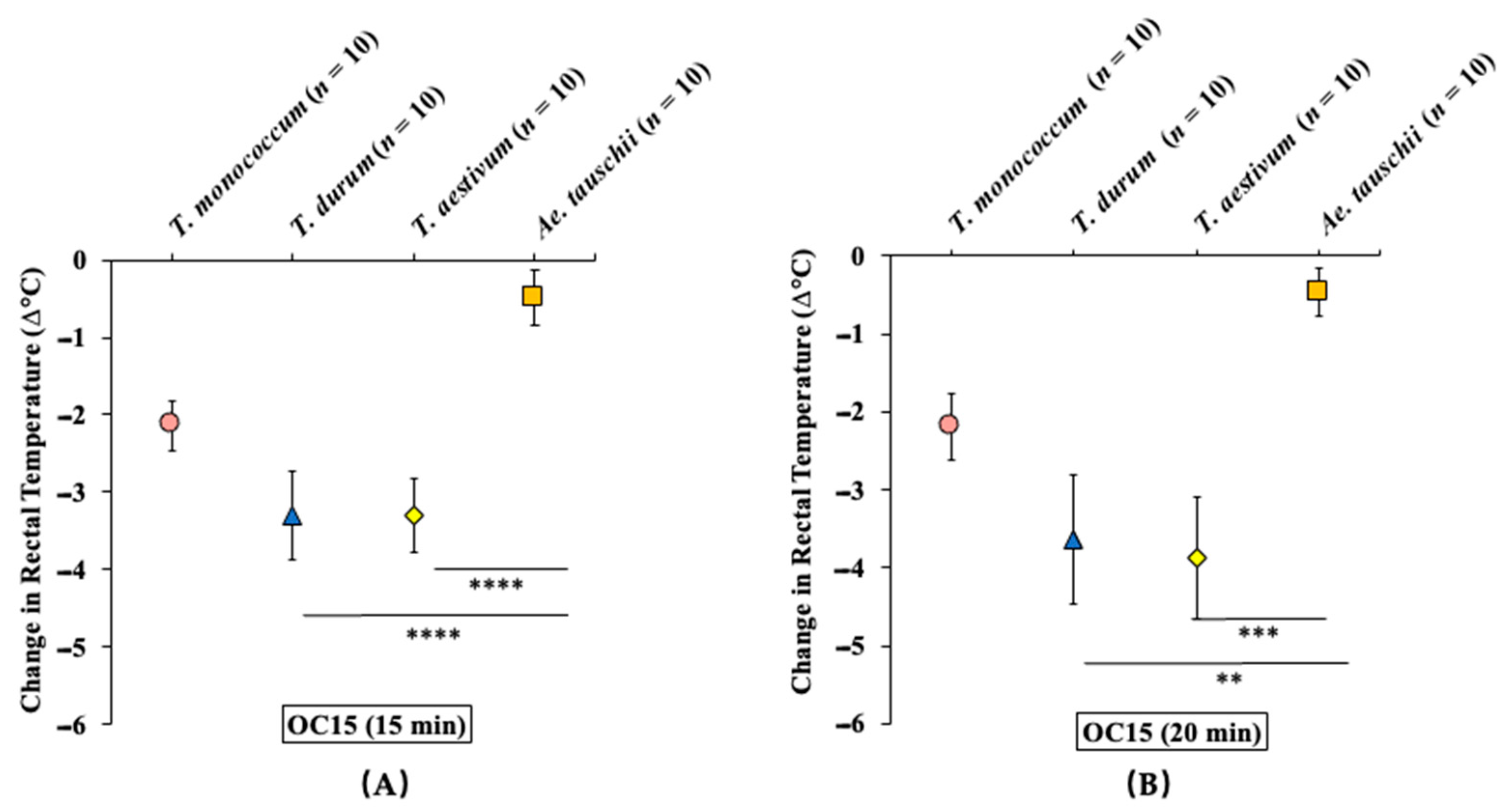
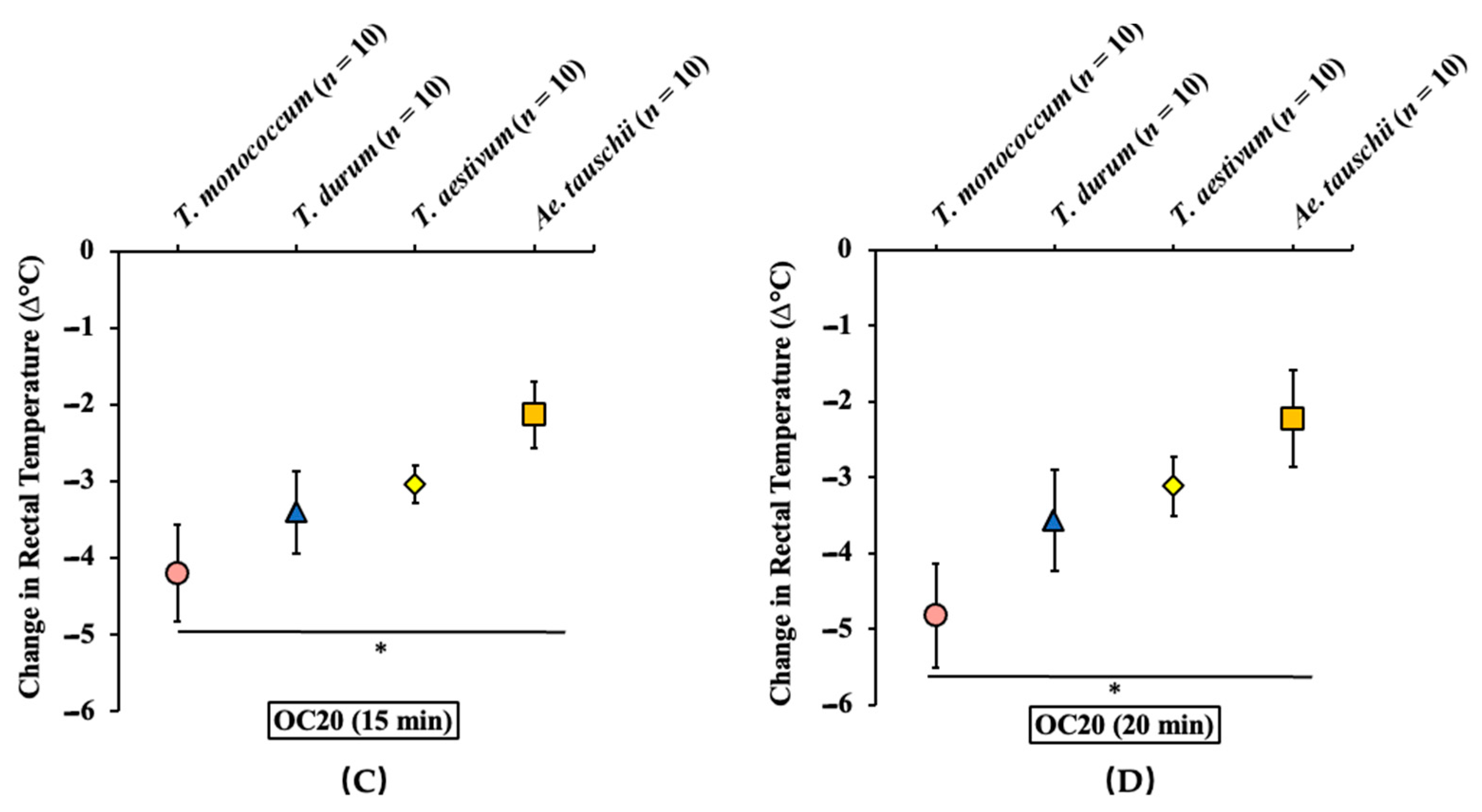
2.3.2. Oral Challenge with Ae. tauschii SSPE Elicits Hypothermia Shock Responses in Skin-Sensitized Mice
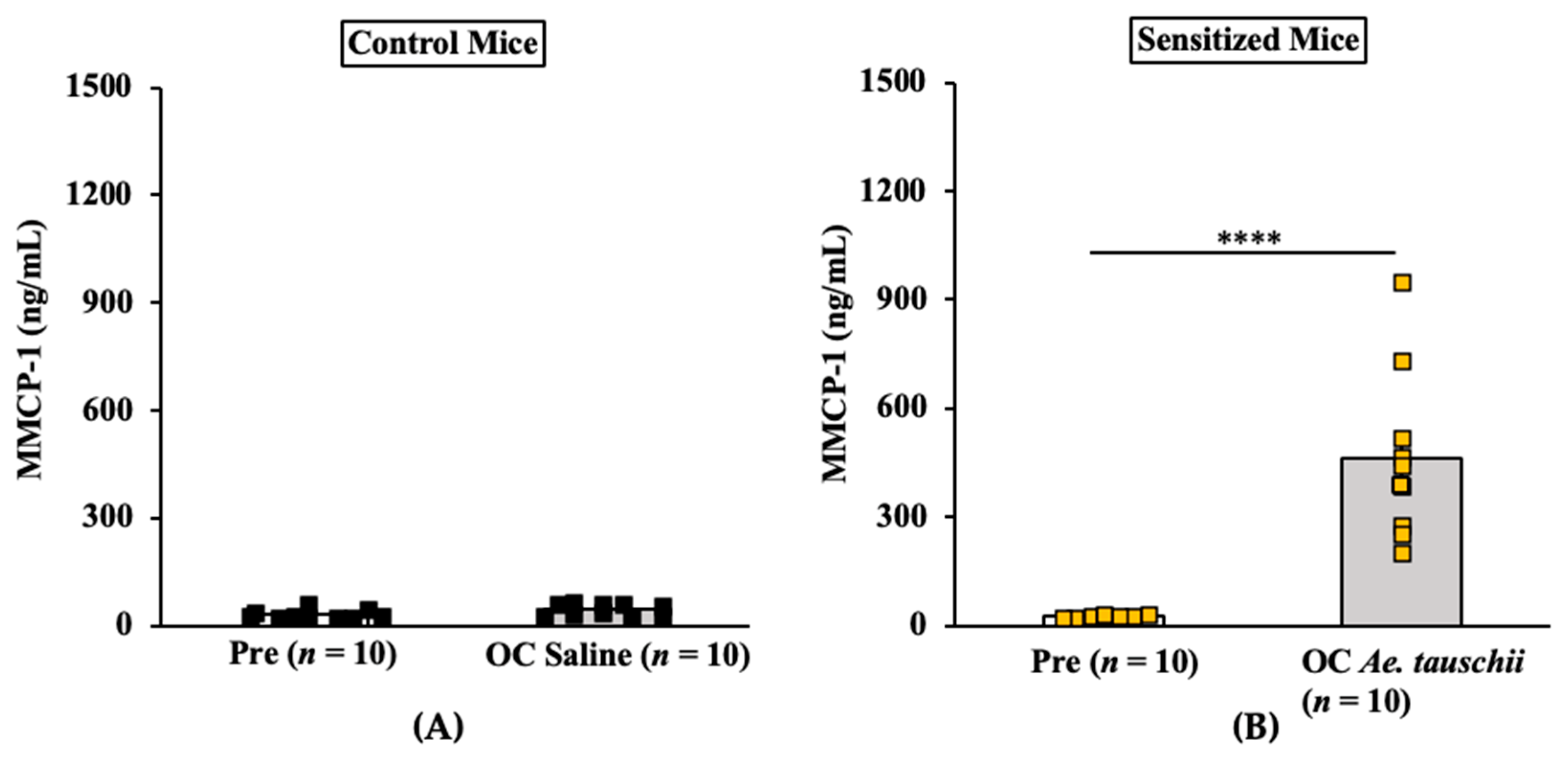
2.3.3. Oral Anaphylaxis Elicited by Ae. tauschii Is Associated with Robust Mucosal Mast Cell Response (MMCR) in Balb/c Mice
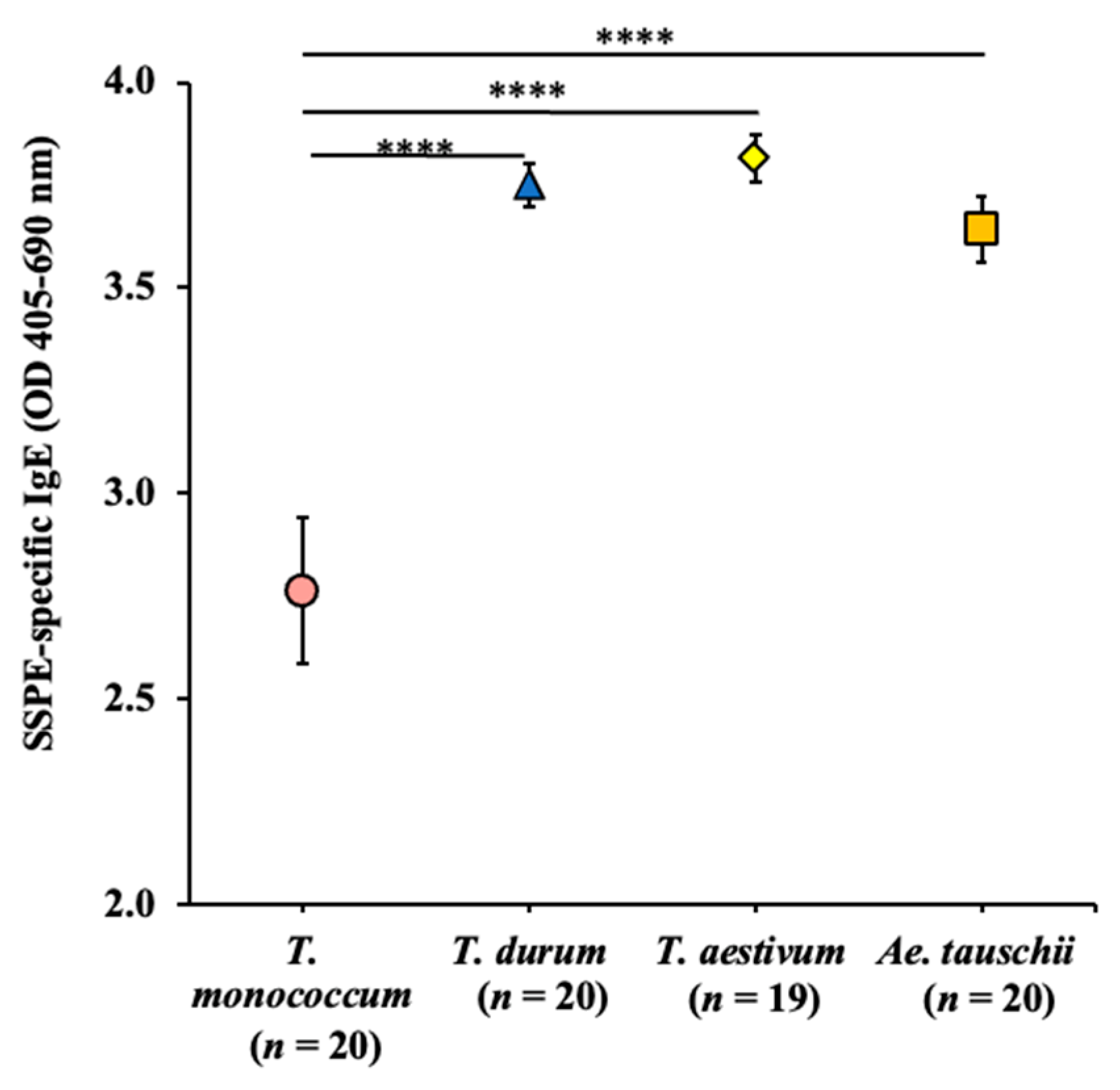
2.4. Comparative Map of the Intrinsic Allergenicity Sensitization Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid, and Hexaploid Wheat
2.5. Comparative Map of the Intrinsic Allergenicity Disease Elicitation Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid, and Hexaploid Wheats
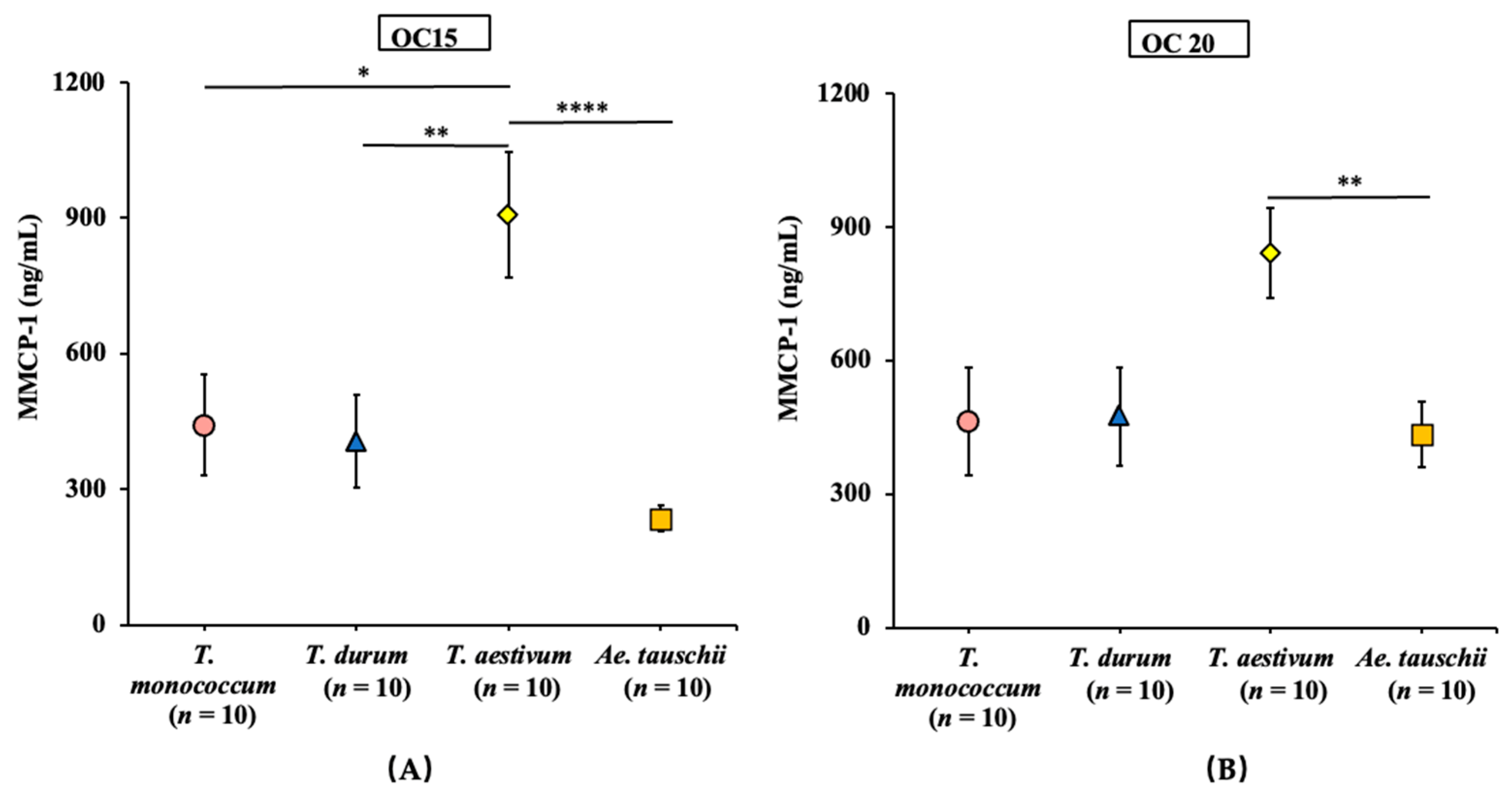
2.6. Comparative Map of the Mucosal Mast Cell Response (MMCR) Elicitation Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid, and Hexaploid Wheats
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Generation of a Plant-Protein-Free Mouse Colony
4.3. Preparation of Salt-Soluble Protein Extract from Wheat Flours
4.4. Skin Sensitization, Bleeding, and Plasma Sample Preparation
4.5. Elicitation of Oral Anaphylaxis and Hypothermic Shock Responses
4.6. Measurement of Wheat SSPE-Specific IgE Antibody Levels
4.7. Quantification of Mucosal Mast Cell Protease-1 (MMCP-1) Level
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sampson, H.A.; O’Mahony, L.; Burks, A.W.; Plaut, M.; Lack, G.; Akdis, C.A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018, 141, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Blumenstock, J.A.; Jiang, J.; Davis, M.M.; Nadeau, K.C. The Public Health Impact of Parent-Reported Childhood Food Allergies in the United States. Pediatrics 2018, 142, e20181235. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Jiang, J.; Blumenstock, J.A.; Davis, M.M.; Schleimer, R.P.; Nadeau, K.C. Prevalence and Severity of Food Allergies Among US Adults. JAMA Netw. Open 2019, 2, e185630. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Scientific Opinion on the evaluation of allergenic foods and food ingredients for labelling purposes. EFSA J. 2014, 12, 3894. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration. Food Allergies. 2022. Available online: https://www.fda.gov/food/food-labeling-nutrition/food-allergies (accessed on 20 September 2022).
- Food Standards Australia New Zealand. Food Allergies and Food Intolerances. 2021. Available online: https://www.foodstandards.gov.au/consumer/foodallergies/allergies/Pages/default.aspx (accessed on 28 September 2022).
- Health Canada. Food Allergen Labeling. 2021. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-labelling/allergen-labelling.html (accessed on 28 September 2022).
- Japan. Food Allergy Labeling. 2019. Available online: https://expatsguide.jp/articles/features/food-allergy-labeling/ (accessed on 28 September 2022).
- UK Food Standards Agency. Food Allergy and Intolerance. 2021. Available online: https://www.food.gov.uk/safety-hygiene/food-allergy-and-intolerance (accessed on 28 September 2022).
- Gupta, R.; Holdford, D.; Bilaver, L.; Dyer, A.; Holl, J.L.; Meltzer, D. The Economic Impact of Childhood Food Allergy in the United States. JAMA Pediatr. 2013, 167, 1026–1031. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.M.; Jiang, J.; Gupta, R.S. Epidemiology and Burden of Food Allergy. Curr. Allergy Asthma Rep. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Sears, E.R. Homoeologous chromosomes in Triticum aestivum. Genet 1952, 37, 624. [Google Scholar]
- McFadden, E.S.; Sears, E.R. The Origin of Triticum Spelta and Its Free-Threshing Hexaploid Relatives. J. Hered. 1946, 37, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Poole, J.A.; Barriga, K.; Leung, D.Y.; Hoffman, M.; Eisenbarth, G.S.; Rewers, M.; Norris, J.M. Timing of Initial Exposure to Cereal Grains and the Risk of Wheat Allergy. Pediatrics 2006, 117, 2175–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, C.; Pereira, B.; Voigt, K.; Grundy, J.; Clayton, C.B.; Higgins, B.; Arshad, S.H.; Dean, T. Original article: Prevalence and cumulative incidence of food hypersensitivity in the first 3 years of life. Allergy 2007, 63, 354–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, C.; Pereira, B.; Grundy, J.; Clayton, C.B.; Arshad, S.H.; Dean, T. Prevalence of sensitization reported and objectively assessed food hypersensitivity amongst six-year-old children: A population-based study. Pediatr. Allergy Immunol. 2006, 17, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Verrill, L.; Bruns, R.; Luccioli, S. Prevalence of self-reported food allergy in U.S. adults: 2001, 2006, and 2010. In Allergy and Asthma Proceedings; Oceanside Publications Inc.: East Providence, RI, USA, 2015; Volume 36, pp. 458–467. [Google Scholar]
- Vierk, K.A.; Koehler, K.M.; Fein, S.B.; Street, D.A. Prevalence of self-reported food allergy in American adults and use of food labels. J. Allergy Clin. Immunol. 2007, 119, 1504–1510. [Google Scholar] [CrossRef]
- Rance, F.; Grandmottet, X.; Grandjean, H. Prevalence and main characteristics of schoolchildren diagnosed withfood allergies in France. Clin. Exp. Allergy 2005, 35, 167–172. [Google Scholar] [CrossRef]
- Woods, R.; Stoney, R.; Raven, J.; Walters, E.; Abramson, M.; Thien, F. Reported adverse food reactions overestimate true food allergy in the community. Eur. J. Clin. Nutr. 2002, 56, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, B.; Venter, C.; Grundy, J.; Clayton, C.B.; Arshad, S.H.; Dean, T. Prevalence of sensitization to food allergens, reported adverse reaction to foods, food avoidance, and food hypersensitivity among teenagers. J. Allergy Clin. Immunol. 2005, 116, 884–892. [Google Scholar] [CrossRef]
- Ebisawa, M.; Ito, K.; Fujisawa, T.; Committee for Japanese Pediatric Guideline for Food Allergy; The Japanese Society of Pediatric Allergy and Clinical Immunology; The Japanese Society of Allergology. Japanese guidelines for food allergy 2020. Allergol. Int. 2020, 69, 370–386. [Google Scholar] [CrossRef]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The Prevalence, Severity, and Distribution of Childhood Food Allergy in the United States. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renz, H.; Allen, K.J.; Sicherer, S.H.; Sampson, H.A.; Lack, G.; Beyer, K.; Oettgen, H.C. Food allergy. Nat. Rev. Dis. Prim. 2018, 4, 17098. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration. Food Allergies: What You Need to Know. 2022. Available online: https://www.fda.gov/food/buy-store-serve-safe-food/food-allergies-what-you-need-know (accessed on 26 January 2022).
- Jin, Y.; Acharya, H.G.; Acharya, D.; Jorgensen, R.; Gao, H.; Secord, J.; Ng, P.K.W.; Gangur, V. Advances in Molecular Mechanisms of Wheat Allergenicity in Animal Models: A Comprehensive Review. Molecules 2019, 24, 1142. [Google Scholar] [CrossRef] [Green Version]
- Cianferoni, A. Wheat allergy: Diagnosis and management. J. Asthma Allergy 2016, 9, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Jorgensen, R.; Raghunath, R.; Nagisetty, S.; Ng, P.K.W.; Gangur, V. Creating hypo-/nonallergenic wheat products using processing methods: Fact or fiction? Compr. Rev. Food Sci. Food Saf. 2021, 20, 6089–6115. [Google Scholar] [CrossRef]
- Nakamura, A.; Tanabe, S.; Watanabe, J.; Makino, T. Primary Screening of Relatively Less Allergenic Wheat Varieties. J. Nutr. Sci. Vitaminol. 2005, 51, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Takahashi, H.; Endo, T.R.; Matsuo, H.; Shiwaku, K.; Morita, E. Characterization of a hypoallergenic wheat line lacking ω-5 gliadin. Allergol. Int. 2016, 65, 400–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larré, C.; Lupi, R.; Gombaud, G.; Brossard, C.; Branlard, G.; Moneret-Vautrin, D.; Rogniaux, H.; Denery-Papini, S. Assessment of allergenicity of diploid and hexaploid wheat genotypes: Identification of allergens in the albumin/globulin fraction. J. Proteom. 2011, 74, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jorgensen, R.; Raghunath, R.; Ng, P.K.W.; Gangur, V. An Adjuvant-Free Mouse Model Using Skin Sensitization Without Tape-Stripping Followed by Oral Elicitation of Anaphylaxis: A Novel Pre-Clinical Tool for Testing Intrinsic Wheat Allergenicity. Front. Allergy 2022, 3, 80. [Google Scholar] [CrossRef]
- Khodoun, M.V.; Strait, R.; Armstrong, L.; Yanase, N.; Finkelman, F.D. Identification of markers that distinguish IgE- from IgG-mediated anaphylaxis. Proc. Natl. Acad. Sci. USA 2011, 108, 12413–12418. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.M.; Siler, L.; Souza, E.; Ng, P.K.W.; Dong, Y.; Brown-Guedira, G.; Jiang, G.-L.; Ward, R.W. Registration of ‘Ambassador’ Wheat. J. Plant Regist. 2010, 4, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Sissons, M. Role of Durum Wheat Composition on the Quality of Pasta and Bread. Food 2008, 2, 75–90. [Google Scholar]
- Nagelkirk, M.; Black, R. Wheat Varieties Used in Michiagn. 2012. Available online: https://www.canr.msu.edu/news/wheat_varieties_used_in_michigan (accessed on 28 September 2022).
- Hidalgo, A.; Brandolini, A. Nutritional properties of einkorn wheat (Triticum monococcum L.). J. Sci. Food Agric. 2014, 94, 601–612. [Google Scholar] [CrossRef]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Gonipeta, B.; Kim, E.; Gangur, V. Mouse Models of Food Allergy: How Well do They Simulate the Human Disorder? Crit. Rev. Food Sci. Nutr. 2015, 55, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Birmingham, N.P.; Parvataneni, S.; Hassan, H.M.A.; Harkema, J.; Samineni, S.; Navuluri, L.; Kelly, C.J.; Gangur, V. An Adjuvant-Free Mouse Model of Tree Nut Allergy Using Hazelnut as a Model Tree Nut. Int. Arch. Allergy Immunol. 2007, 144, 203–210. [Google Scholar] [CrossRef]
- Masthoff, L.J.; Hoff, R.; Verhoeckx, K.C.M.; Van Os-Medendorp, H.; Michelsen-Huisman, A.; Baumert, J.L.; Pasmans, S.G.; Meijer, Y.; Knulst, A.C. A systematic review of the effect of thermal processing on the allergenicity of tree nuts. Allergy 2013, 68, 983–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanga, S.K.; Singh, A.; Raghavan, V. Review of conventional and novel food processing methods on food allergens. Crit. Rev. Food Sci. Nutr. 2017, 57, 2077–2094. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.; Jorgensen, R.; Raghunath, R.; Chandra, S.; Othman, A.; Olson, E.; Ng, P.K.W.; Gangur, V. Intrinsic Allergenicity Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid and Hexaploid Wheats: Validation Using an Adjuvant-Free Mouse Model. Int. J. Mol. Sci. 2023, 24, 5453. https://doi.org/10.3390/ijms24065453
Gao H, Jorgensen R, Raghunath R, Chandra S, Othman A, Olson E, Ng PKW, Gangur V. Intrinsic Allergenicity Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid and Hexaploid Wheats: Validation Using an Adjuvant-Free Mouse Model. International Journal of Molecular Sciences. 2023; 24(6):5453. https://doi.org/10.3390/ijms24065453
Chicago/Turabian StyleGao, Haoran, Rick Jorgensen, Rajsri Raghunath, Shivam Chandra, Aqilah Othman, Eric Olson, Perry K. W. Ng, and Venu Gangur. 2023. "Intrinsic Allergenicity Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid and Hexaploid Wheats: Validation Using an Adjuvant-Free Mouse Model" International Journal of Molecular Sciences 24, no. 6: 5453. https://doi.org/10.3390/ijms24065453
APA StyleGao, H., Jorgensen, R., Raghunath, R., Chandra, S., Othman, A., Olson, E., Ng, P. K. W., & Gangur, V. (2023). Intrinsic Allergenicity Potential of Salt-Soluble Protein Extracts from the Diploid, Tetraploid and Hexaploid Wheats: Validation Using an Adjuvant-Free Mouse Model. International Journal of Molecular Sciences, 24(6), 5453. https://doi.org/10.3390/ijms24065453