
Changes in the Extracellular Matrix in Endometrial and Cervical Cancer: A Systematic Review




Abstract
:1. Introduction
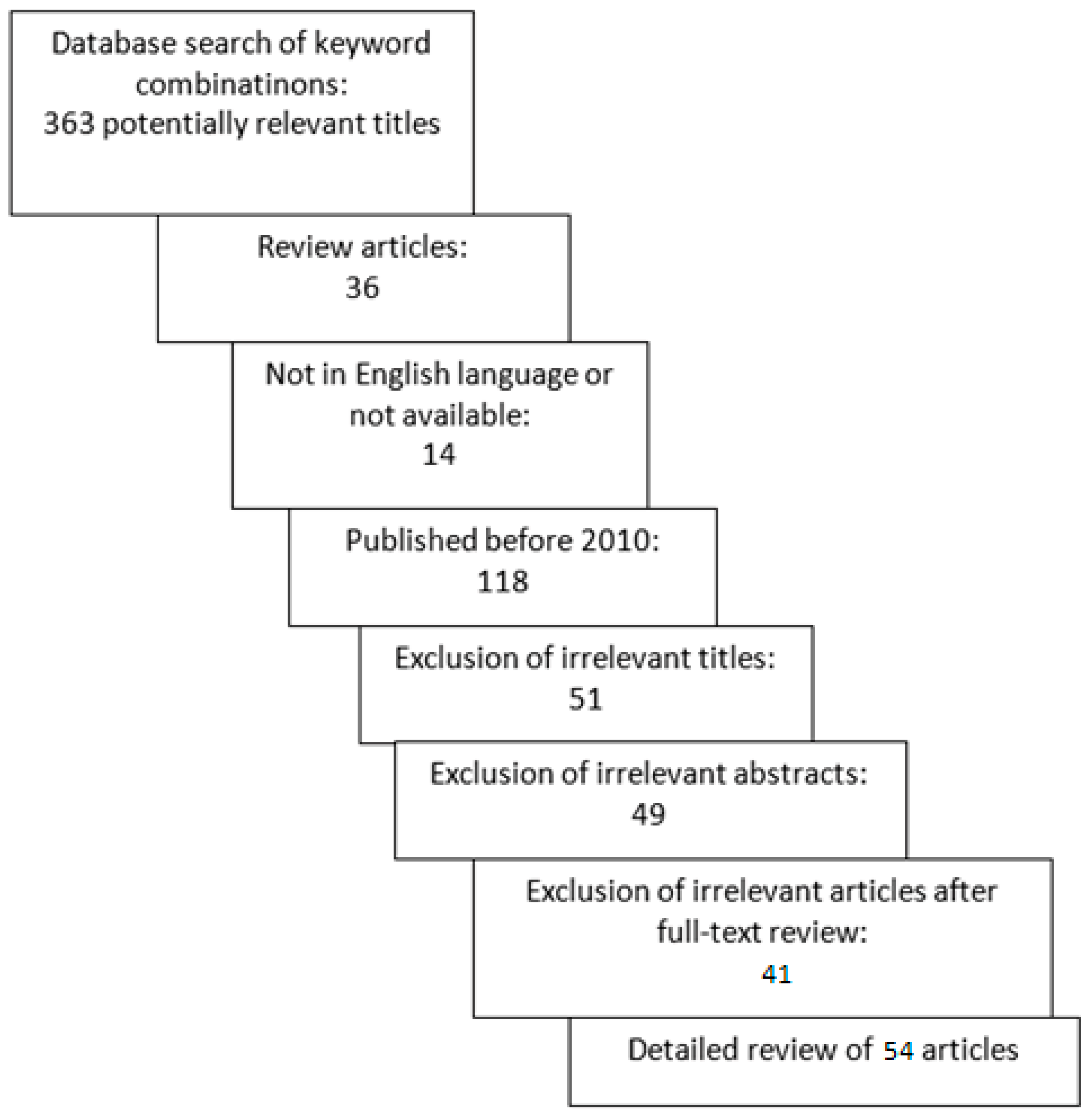
2. Materials and Methods
3. Results
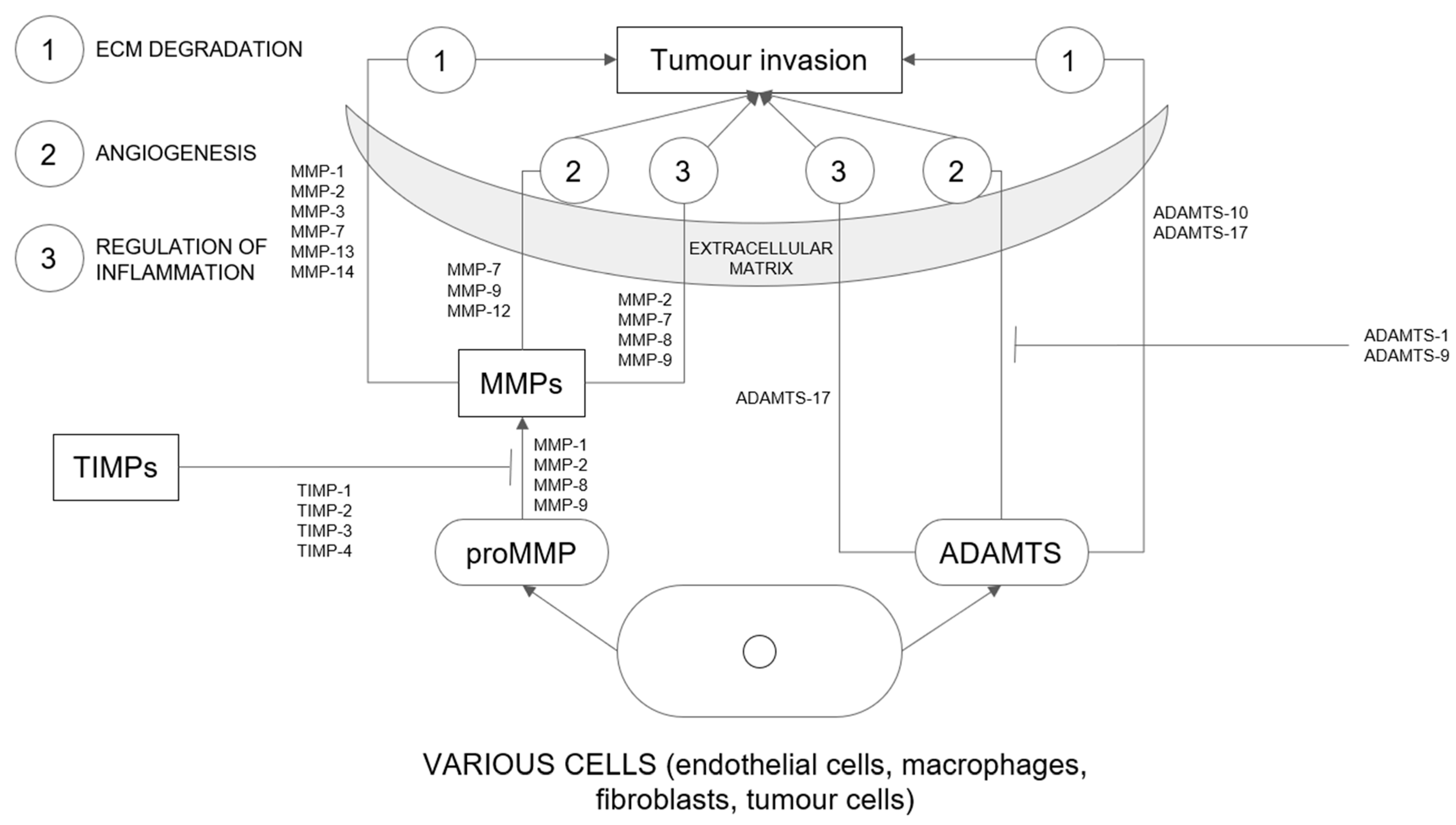
3.1. Matrix Metalloproteinases
3.2. A Disintegrin and Metalloproteinase with Thrombospondin Motifs (ADAMTS)

{kind=link}
{kind=link}
| Type of Cancer | Component of ECM | Action in Tumor Microenvironment |
|---|---|---|
| Endometrial cancer | MMP 2 [18,28,33,34] | Degradation of collagen type 4 in basal membrane |
| MMP 9 [15,18,31,34] | Cancer progression, dependent on hormonal status | |
| MMP 11 [18,35] | Activation of other MMPs | |
| ADAMTS 1 [27] | Antiangiogenic effect | |
| ADAMTS 5 [32] | Involved in tumorigenesis | |
| ADAMTS 8 [32] | Tumour suppressor function | |
| ADAMTS 9 [27,32] | Antiangiogenic effect, degradation of aggrecan and versican | |
| ADAMTS 20 [27] | No statistically significant differences have been found between cancerous and healthy tissues | |
| Cervical cancer | MMP 2 [28] | Degradation of ECM, regulation of other MMPs |
| MMP 9 [15,34] | Associated with better survival rate | |
| MMP1, -3, -11, -13, -14, -15, -17, -24, -28 [18] | Found in Caski, HeLa and SiHa cell lines, their role in tumour development has not been fully understood yet | |
| ADAMTS17 [31] | Correlated with poor prognosis |
3.3. MMPs and CD147
3.4. Interplay of MMPs and Other Molecules
3.5. Tissue Inhibitors of Metalloproteinases (TIMPs)
3.6. Major ECM Adhesion Molecules
3.7. Stromal ECM Components
3.8. Fibulins
3.9. Cell Invasion
4. Discussion
Roots of Further Exploration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Maghrabi, H.; Gomaa, W.; Al-Maghrabi, J. Increased Osteopontin Expression in Endometrial Carcinoma Is Associated with Better Survival Outcome. Ginekol. Pol. 2020, 91, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Zhang, H.; Qiu, H.; Li, B.; Wang, J.; Du, G.; Ren, C.; Wan, X. EFEMP1 Is Repressed by Estrogen and Inhibits the Epithelial-Mesenchymal Transition via Wnt/β-Catenin Signaling in Endometrial Carcinoma. Oncotarget 2016, 7, 25712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres, G.F.; Spadoto-Dias, D.; Bueloni-Dias, F.N.; Leite, N.J.; Elias, L.V.; Custódio Domingues, M.A.; Padovani, C.R.; Dias, R. Immunohistochemical Expression of Hormone Receptors, Ki-67, Endoglin (CD105), Claudins 3 and 4, MMP-2 and-9 in Endometrial Polyps and Endometrial Cancer Type I. OncoTargets Ther. 2018, 11, 3949–3958. [Google Scholar] [CrossRef] [Green Version]
- Lay, V.; Yap, J.; Sonderegger, S.; Dimitriadis, E. Interleukin 11 Regulates Endometrial Cancer Cell Adhesion and Migration via STAT3. Int. J. Oncol. 2012, 41, 759–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashima, H.; Wu, R.C.; Wang, Y.; Sinno, A.K.; Miyamoto, T.; Shiozawa, T.; Wang, T.L.; Fader, A.N.; Shih, I.M. Laminin C1 Expression by Uterine Carcinoma Cells Is Associated with Tumor Progression. Gynecol. Oncol. 2015, 139, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.K.; Lee, T.Y.; Hsu, J.B.K.; Huang, H.-D.; Yang, W.C.V.; Chang, T.H. Computational Analysis for Identification of the Extracellular Matrix Molecules Involved in Endometrial Cancer Progression. PLoS ONE 2020, 15, e0231594. [Google Scholar] [CrossRef] [Green Version]
- Divine, L.M.; Nguyen, M.R.; Meller, E.; Desai, R.A.; Arif, B.; Rankin, E.B.; Bligard, K.H.; Meyerson, C.; Hagemann, I.S.; Massad, M.; et al. AXL Modulates Extracellular Matrix Protein Expression and Is Essential for Invasion and Metastasis in Endometrial Cancer. Oncotarget 2016, 7, 77291. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Watanabe, M.; Hashimoto, K.; Ota, T.; Akimoto, N.; Imada, K.; Nomizu, M.; Ito, A. A Novel Functional Site of Extracellular Matrix Metalloproteinase Inducer (EMMPRIN) That Limits the Migration of Human Uterine Cervical Carcinoma Cells. Int. J. Oncol. 2012, 40, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Wang, J.; Cheng, J.; Zang, C.; Chen, F.; Wang, W.; Zhao, H.; Wang, Y.; Wang, D. Comprehensive Identification of the Human Secretome as Potential Indicators in Treatment Outcome of HPV-Positive and -Negative Cervical Cancer Patients. Gynecol. Obstet. Investig. 2020, 85, 405–415. [Google Scholar] [CrossRef]
- Zhang, R.; Lu, H.; Lyu, Y.Y.; Yang, X.M.; Zhu, L.Y.; Yang, G.D.; Jiang, P.C.; Re, Y.; Song, W.W.; Wang, J.H.; et al. E6/E7-P53-POU2F1-CTHRC1 Axis Promotes Cervical Cancer Metastasis and Activates Wnt/PCP Pathway. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kaewprag, J.; Umnajvijit, W.; Ngamkham, J.; Ponglikitmongkol, M. HPV16 Oncoproteins Promote Cervical Cancer Invasiveness by Upregulating Specific Matrix Metalloproteinases. PLoS ONE 2013, 8, e71611. [Google Scholar] [CrossRef]
- Balasubramaniam, S.D.; Wong, K.K.; Oon, C.E.; Balakrishnan, V.; Kaur, G. Comparative Transcriptomic Profiling in HPV-Associated Cervical Carcinogenesis: Implication of MHC Class II and Immunoglobulin Heavy Chain Genes. Life Sci. 2020, 256. [Google Scholar] [CrossRef]
- Liu, M.; Liu, J.; Yang, B.; Gao, X.; Gao, L.L.; Kong, Q.Y.; Zhang, P.; Li, H. Inversed Expression Patterns of S100A4 and E-Cadherin in Cervical Cancers: Implication in Epithelial–Mesenchymal Transition. Anatomical. Record 2017, 300, 2184–2191. [Google Scholar] [CrossRef] [Green Version]
- Sidorkiewicz, I.; Piskór, B.; Dąbrowska, E.; Guzińska-Ustymowicz, K.; Pryczynicz, A.; Zbucka-Krętowska, M.; Ławicki, S. Plasma Levels and Tissue Expression of Selected Cytokines, Metalloproteinases and Tissue Inhibitors in Patients with Cervical Cancer. Anticancer Res. 2019, 39, 6403–6412. [Google Scholar] [CrossRef]
- Azevedo Martins, J.M.; Rabelo-Santos, S.H.; do Amaral Westin, M.C.; Zeferino, L.C. Tumoral and Stromal Expression of MMP-2, MMP-9, MMP-14, TIMP-1, TIMP-2, and VEGF-A in Cervical Cancer Patient Survival: A Competing Risk Analysis. BMC Cancer 2020, 20, 660. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qi, C.; Liu, X.; Li, C.; Chen, J.; Shi, M. Fibulin-3 Knockdown Inhibits Cervical Cancer Cell Growth and Metastasis in Vitro and in Vivo. Sci. Rep. 2018, 8, 10594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, S.S.; Yuan Quah, M.; Nielsen, S.; Atkins, J.; Au, G.G.; Cairns, M.J.; Nahar, P.; Lombard, J.M.; Tanwar, P.S. Inhibition of Extracellular Matrix Mediated TGF-β Signalling Suppresses Endometrial Cancer Metastasis. Oncotarget 2017, 8, 71400. [Google Scholar] [CrossRef] [Green Version]
- Schröpfer, A.; Kammerer, U.; Kapp, M.; Dietl, J.; Feix, S.; Anacker, J. Expression Pattern of Matrix Metalloproteinases in Human Gynecological Cancer Cell Lines. BMC Cancer 2010, 10, 553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oplawski, M.; Dziobek, K.; Zmarzły, N.; Grabarek, B.; Tomala, B.; Leśniak, E.; Adwent, I.; Januszyk, P.; Dąbruś, D.; Boroń, D. Evaluation of Changes in the Expression Pattern of EDIL3 in Different Grades of Endometrial Cancer. Curr. Pharm. Biotechnol. 2019, 20, 483–488. [Google Scholar] [CrossRef]
- Futyma, K.; Miotła, P.; Rózyńska, K.; Zdunek, M.; Semczuk, A.; Rechberger, T.; Wojcierowski, J. Expression of Genes Encoding Extracellular Matrix Proteins: A Macroarray Study. Oncol. Rep. 2014, 32, 2349–2353. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.J.R.; Crook, M.L. CD147 (EMMPRIN) and Matrix Metalloproteinase-2 Expression in Uterine Endometrioid Adenocarcinoma. Pathol. Res. Pract. 2011, 207, 30–36. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Y.; Cui, D.; Ma, Y.; Sui, L.; Zhu, C.; Kong, H.; Kong, Y. Osteopontin Promotes Invasion, Migration and Epithelial-Mesenchymal Transition of Human Endometrial Carcinoma Cell HEC-1A through AKT and ERK1/2 Signaling. Cell. Physiol. Biochem. 2015, 37, 1503–1512. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.M.; Waheed, S.; Syed, V. RNA Interference Screening Identifies Clathrin-B and Cofilin-1 as Mediators of MT1-MMP in Endometrial Cancer. Exp. Cell Res. 2018, 370, 663–670. [Google Scholar] [CrossRef]
- Gómez-Macías, G.S.; Garza-Rodríguez, M.L.; Garza-Guajardo, R.; Monsiváis-Ovalle, D.; Ancer-Rodríguez, J.; Barrera-Saldaña, H.A.; Barboza-Quintana, O. Overexpression of the Matrix Metalloproteinase 11 Gene Is a Potential Biomarker for Type 1 Endometrial Cancer. Oncol. Lett. 2018, 16, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Shen, N.; Yang, S.Y.; Zhao, L.; Guan, Y.M. Extracellular Matrix Metalloproteinase Inducer and Matrix Metalloproteinase-2 Overexpression Is Associated with Loss of Hormone Receptor Expression and Poor Prognosis in Endometrial Cancer. Oncol. Lett. 2015, 10, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Cho-Clark, M.; Larco, D.O.; Zahn, B.R.; Mani, S.K.; Wu, T.J. GnRH-(1-5) Activates Matrix Metallopeptidase-9 to Release Epidermal Growth Factor and Promote Cellular Invasion. Mol. Cell Endocrinol. 2015, 415, 114–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokmak, A.; Ozaksit, G.; Sarikaya, E.; Kuru-Pekcan, M.; Kosem, A. Decreased ADAMTS-1, -9 and -20 Levels in Women with Endometrial Polyps: A Possible Link between Extracellular Matrix Proteases and Endometrial Pathologies*. J. Obstet. Gynaecol. 2019, 39, 845–850. [Google Scholar] [CrossRef]
- Shukla, S.; Qureshi, S.; Singh, U.; Khattri, S. A Study of Matrix Metalloproteinase-2 and Interleukin-18 in Preinvasive and Invasive Lesions of Cancer Cervix. J. Midlife Health 2020, 11, 236–239. [Google Scholar] [CrossRef]
- Maity, G.; Fahreen, S.; Banerji, A.; Roy Choudhury, P.; Sen, T.; Dutta, A.; Chatterjee, A. Fibronectin-Integrin Mediated Signaling in Human Cervical Cancer Cells (SiHa). Mol. Cell. Biochem. 2010, 336, 65–74. [Google Scholar]
- Skrzypczak, M.; Springwald, A.; Lattrich, C.; Häring, J.; Schüler, S.; Ortmann, O.; Treeck, O. Expression of Cysteine Protease Cathepsin L Is Increased in Endometrial Cancer and Correlates with Expression of Growth Regulatory Genes. Cancer Investig. 2012, 30, 398–403. [Google Scholar] [CrossRef]
- Xu, Q.; Ying, M.; Chen, G.; Lin, A.; Xie, Y.; Ohara, N.; Zhou, D. ADAM17 Is Associated with EMMPRIN and Predicts Poor Prognosis in Patients with Uterine Cervical Carcinoma. Tumor Biol. 2014, 35, 7575–7586. [Google Scholar] [CrossRef]
- Yilmaz, E.; Melekoglu, R.; Taskapan, C.; Olmez Budak, F.; Toprak, S. The Investigation of Serum Levels of ADAMTS 5 and 8 (the A Disintegrin and Metalloproteinase with Thrombospondin Motifs) in the Etiology of Endometrial Cancer. J. Obstet. Gynaecol. 2020, 40, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yi, W.; Wang, C.; Wang, J. The Clinicopathological Significance and Prognostic Value of EMMPRIN Overexpression in Cancers: Evidence from 39 Cohort Studies. Oncotarget 2017, 8, 82643. [Google Scholar] [CrossRef] [Green Version]
- Maity, G.; Sen, T.; Chatterjee, A. Laminin Induces Matrix Metalloproteinase-9 Expression and Activation in Human Cervical Cancer Cell Line (SiHa). J. Cancer Res. Clin. Oncol. 2011, 137, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kawana, K.; Adachi, K.; Fujimoto, A.; Yoshida, M.; Nakamura, H.; Nishida, H.; Inoue, T.; Taguchi, A.; Takahashi, J.; et al. Decreased Expression of the Plasminogen Activator Inhibitor Type 1 Is Involved in Degradation of Extracellular Matrix Surrounding Cervical Cancer Stem Cells. Int. J. Oncol. 2016, 48, 829–835. [Google Scholar] [CrossRef] [Green Version]
- Fullár, A.; Dudás, J.; Oláh, L.; Hollósi, P.; Papp, Z.; Sobel, G.; Karászi, K.; Paku, S.; Baghy, K.; Kovalszky, I. Remodeling of Extracellular Matrix by Normal and Tumor-Associated Fibroblasts Promotes Cervical Cancer Progression. BMC Cancer 2015, 15, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizarraga, F.; Ceballos-Cancino, G.; Espinosa, M.; Vazquez-Santillan, K.; Maldonado, V.; Melendez-Zajgla, J. Tissue Inhibitor of Metalloproteinase-4 Triggers Apoptosis in Cervical Cancer Cells. PLoS ONE 2015, 10, e0135929. [Google Scholar] [CrossRef] [PubMed]
- Lizarraga, F.; Espinosa, M.; Ceballos-Cancino, G.; Vazquez-Santillan, K.; Bahena-Ocampo, I.; Schwarz-Cruz y Celis, A.; Vega-Gordillo, M.; Garcia Lopez, P.; Maldonado, V.; Melendez-Zajgla, J. Tissue Inhibitor of Metalloproteinases-4 (TIMP-4) Regulates Stemness in Cervical Cancer Cells. Mol. Carcinog. 2016, 55, 1952–1961. [Google Scholar] [CrossRef]
- Dogan Gun, B.; Bahadir, B.; Bektas, S.; Barut, F.; Yurdakan, G.; Kandemir, N.O.; Oguz Ozdamar, S. Clinicopathological Significance of Fascin and CD44v6 Expression in Endometrioid Carcinoma; Springer: Cham, Switzerland, 2012. [Google Scholar]
- Li, M.; Xin, X.-Y.; Jin, Z.-S.; Hua, T.; Wang, H.-B.; Wang, H.-B. Transcriptomic Analysis of Stromal Cells from Patients with Endometrial Carcinoma. Int. J. Clin. Exp. Pathol. 2017, 10, 9853. [Google Scholar]
- Gorter, A.; Zijlmans, H.J.; van Gent, H.; Trimbos, J.B.; Fleuren, G.J.; Jordanova, E.S. Versican Expression Is Associated with Tumor-Infiltrating CD8-Positive T Cells and Infiltration Depth in Cervical Cancer. Mod. Pathol. 2010, 23, 1605–1615. [Google Scholar] [CrossRef] [Green Version]
- Turan, T.; Torun, M.; Atalay, F.; Gönenç, A. Assessment of Vitronectin, Soluble Epithelial-Cadherin and TGF-Β1 as a Serum Biomarker with Predictive Value for Endometrial and Ovarian Cancers. Turk. J. Pharm. Sci. 2017, 14, 141–147. [Google Scholar] [CrossRef]
- Yoshida, S.; Asanoma, K.; Yagi, H.; Onoyama, I.; Hori, E.; Matsumura, Y.; Okugawa, K.; Yahata, H.; Kato, K. Fibronectin Mediates Activation of Stromal Fibroblasts by SPARC in Endometrial Cancer Cells. BMC Cancer 2021, 21, 156. [Google Scholar] [CrossRef]
- Yang, T.; Qiu, H.; Bao, W.; Li, B.; Lu, C.; Du, G.; Luo, X.; Wang, L.; Wan, X. Epigenetic Inactivation of EFEMP1 Is Associated with Tumor Suppressive Function in Endometrial Carcinoma. PLoS ONE 2013, 8, e67458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, M.; Fang, S.; Wang, Q.; Fang, R.; Chen, J. Fibulin-4 Is Associated with Prognosis of Endometrial Cancer Patients and Inhibits Cancer Cell Invasion and Metastasis via Wnt/β-Catenin Signaling Pathway. Oncotarget 2017, 8, 18991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, J.; Liu, X.; Fang, R.; Zhao, Y.; Ma, D. Overexpression of Fibulin-4 Is Associated with Tumor Progression and Poor Prognosis in Patients with Cervical Carcinoma. Oncol. Rep. 2014, 31, 2601–2610. [Google Scholar] [CrossRef] [Green Version]
- Winship, A.L.; Rainczuk, K.; Ton, A.; Dimitriadis, E. Fibulin-5 Localisation in Human Endometrial Cancer Shifts from Epithelial to Stromal with Increasing Tumour Grade, and Silencing Promotes Endometrial Epithelial Cancer Cell Proliferation. Oncol. Lett. 2016, 12, 651–657. [Google Scholar] [CrossRef] [Green Version]
- En-lin, S.; Sheng-guo, C.; Hua-qiao, W. The Expression of EFEMP1 in Cervical Carcinoma and Its Relationship with Prognosis. Gynecol. Oncol. 2010, 117, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Song, E.L.; Hou, Y.P.; Yu, S.P.; Chen, S.G.; Huang, J.T.; Luo, T.; Kong, L.P.; Xu, J.; Wang, H.Q. EFEMP1 Expression Promotes Angiogenesis and Accelerates the Growth of Cervical Cancer in Vivo. Gynecol. Oncol. 2011, 121, 174–180. [Google Scholar] [CrossRef]
- Lesjak, M.S.; Marchan, R.; Stewart, J.D.; Rempel, E.; Rahnenführer, J.; Hengstler, J.G. EDI3 Links Choline Metabolism to Integrin Expression, Cell Adhesion and Spreading. Cell Adh. Migr. 2014, 8, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.Y.; Yin, K.M.; Bai, Y.H.; Zhang, Z.G.; Di, W.; Zhang, S. CTHRC1 Promotes M2-like Macrophage Recruitment and Myometrial Invasion in Endometrial Carcinoma by Integrin-Akt Signaling Pathway. Clin. Exp. Metastasis 2019, 36, 351–363. [Google Scholar] [CrossRef]
- Cao, C.; Lin, S.; Zhi, W.; Lazare, C.; Meng, Y.; Wu, P.; Gao, P.; Wei, J.; Wu, P. LOXL2 Expression Status Is Correlated With Molecular Characterizations of Cervical Carcinoma and Associated With Poor Cancer Survival via Epithelial-Mesenchymal Transition (EMT) Phenotype. Front. Oncol. 2020, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Sang, M.M.; Chen, C.; Zhu, W.T.; Gong, Y.-S.; Pei, D.S. CSN6 Promotes the Migration and Invasion of Cervical Cancer Cells by Inhibiting Autophagic Degradation of Cathepsin L. Int. J. Biol. Sci. 2019, 15, 1310–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Min, W.; Liu, X.; Xie, C.; Tang, J.; Yi, T.; Li, Z.; Zhao, X. Identification of FRAS1 as a Human Endometrial Carcinoma-Derived Protein in Serum of Xenograft Model. Gynecol. Oncol. 2012, 127, 406–411. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padežnik, T.; Oleksy, A.; Cokan, A.; Takač, I.; Sobočan, M. Changes in the Extracellular Matrix in Endometrial and Cervical Cancer: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 5463. https://doi.org/10.3390/ijms24065463
Padežnik T, Oleksy A, Cokan A, Takač I, Sobočan M. Changes in the Extracellular Matrix in Endometrial and Cervical Cancer: A Systematic Review. International Journal of Molecular Sciences. 2023; 24(6):5463. https://doi.org/10.3390/ijms24065463
Chicago/Turabian StylePadežnik, Tjaša, Anja Oleksy, Andrej Cokan, Iztok Takač, and Monika Sobočan. 2023. "Changes in the Extracellular Matrix in Endometrial and Cervical Cancer: A Systematic Review" International Journal of Molecular Sciences 24, no. 6: 5463. https://doi.org/10.3390/ijms24065463
APA StylePadežnik, T., Oleksy, A., Cokan, A., Takač, I., & Sobočan, M. (2023). Changes in the Extracellular Matrix in Endometrial and Cervical Cancer: A Systematic Review. International Journal of Molecular Sciences, 24(6), 5463. https://doi.org/10.3390/ijms24065463