
Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides


 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
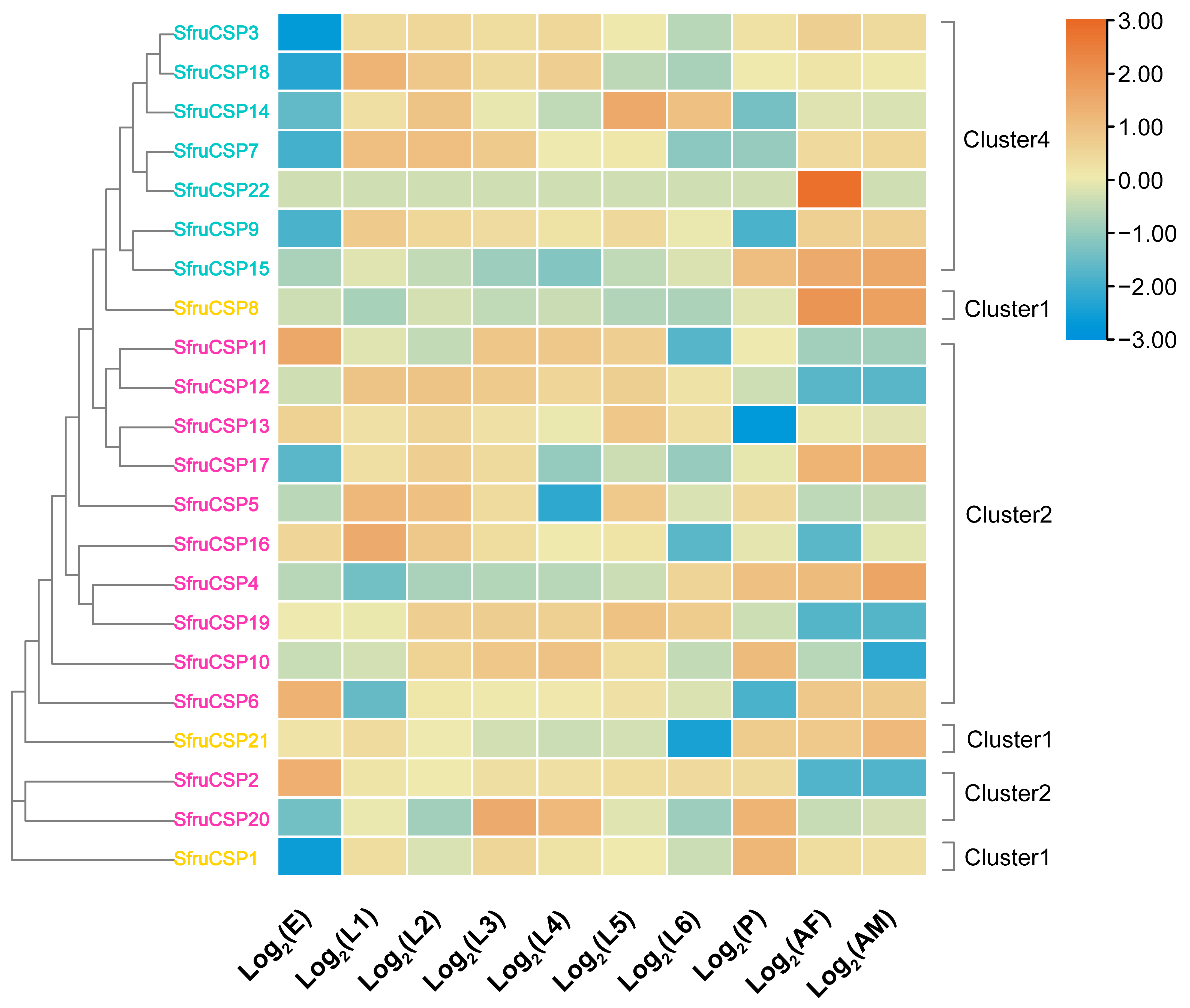
2. Results
2.1. Identification of OBP and CSP Genes in S. frugiperda
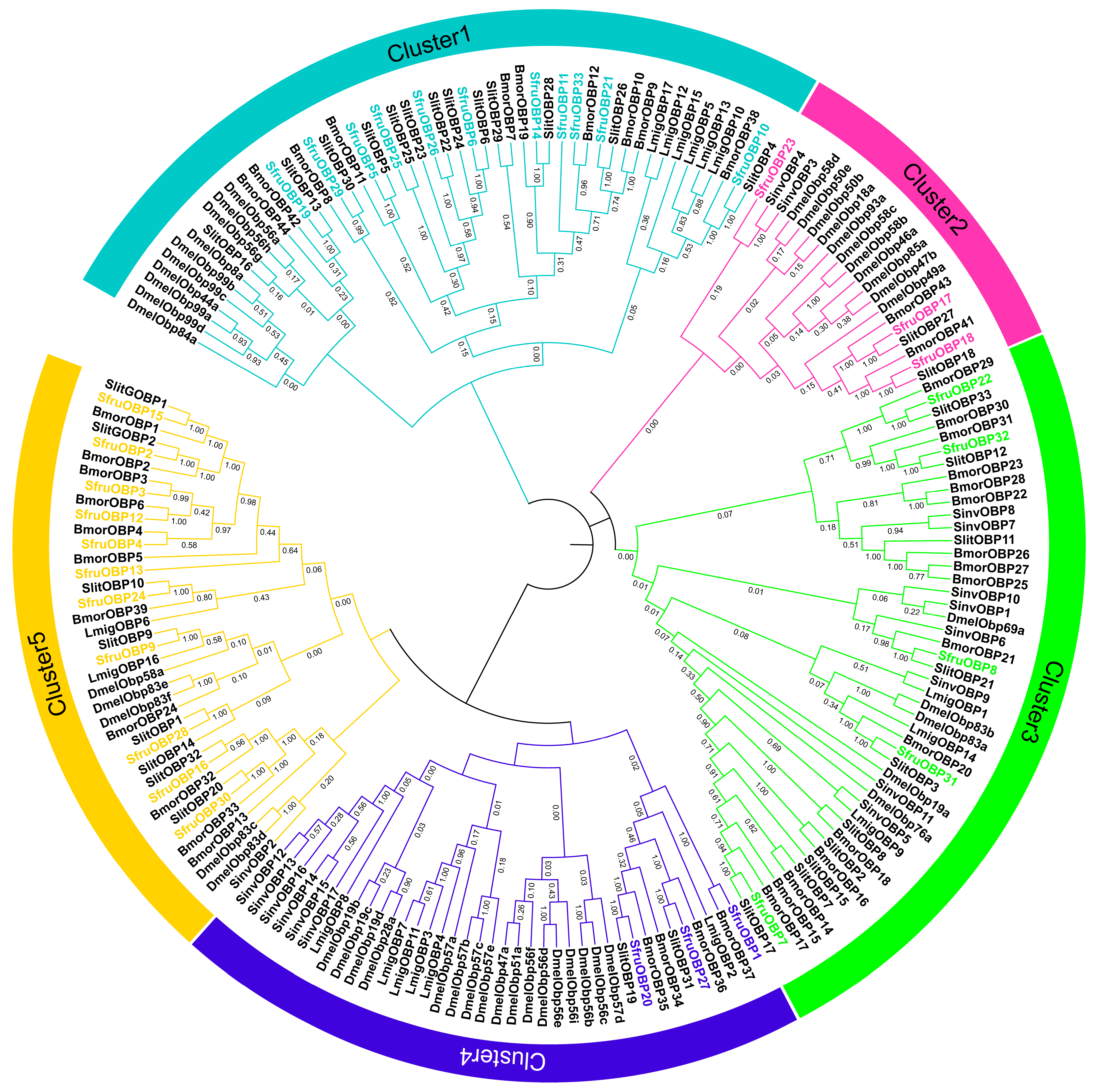
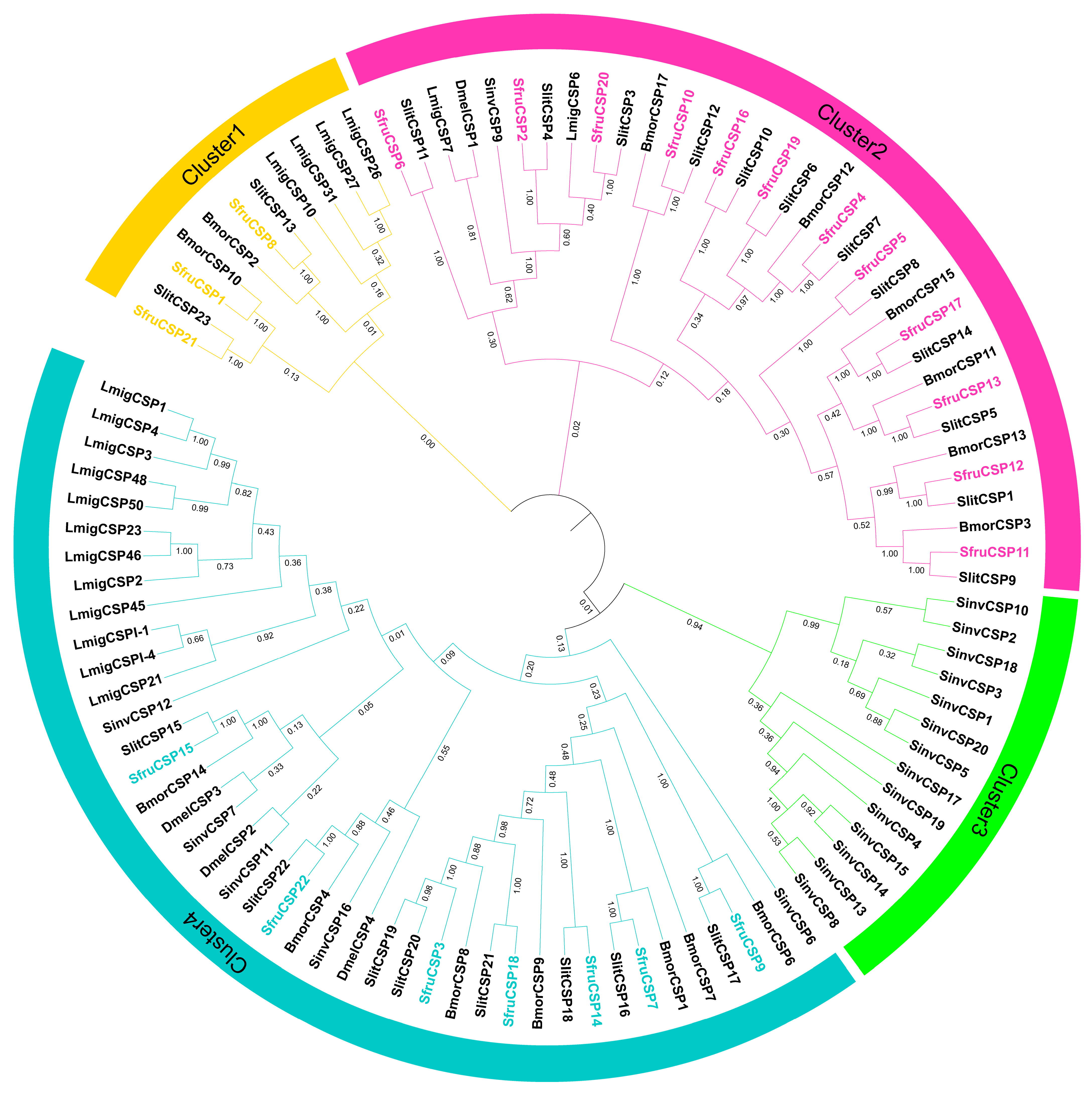
2.2. Phylogenetic Analysis of OBP and CSP in S. frugiperda
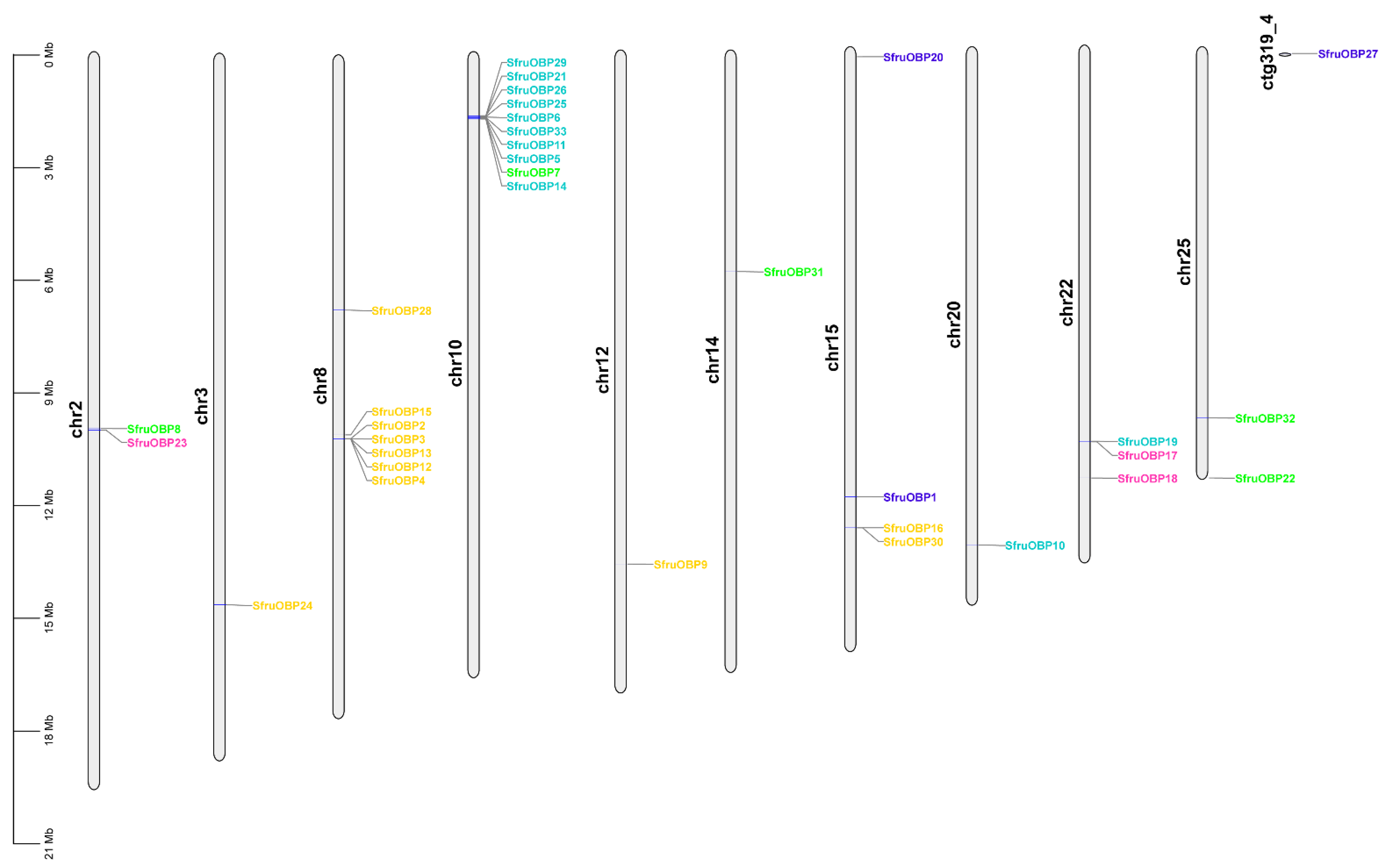
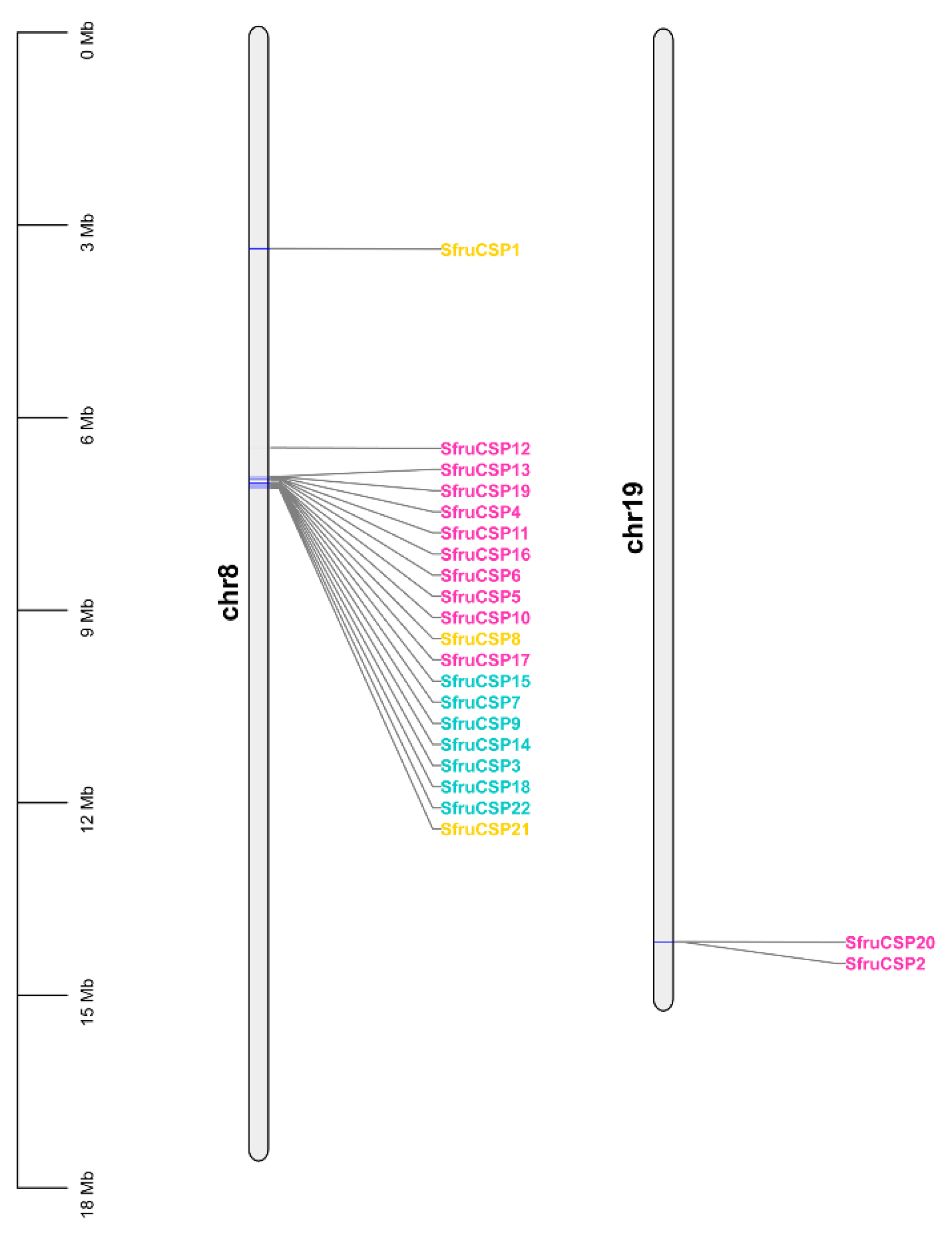
2.3. Chromosomal Distribution of OBP and CSP Genes
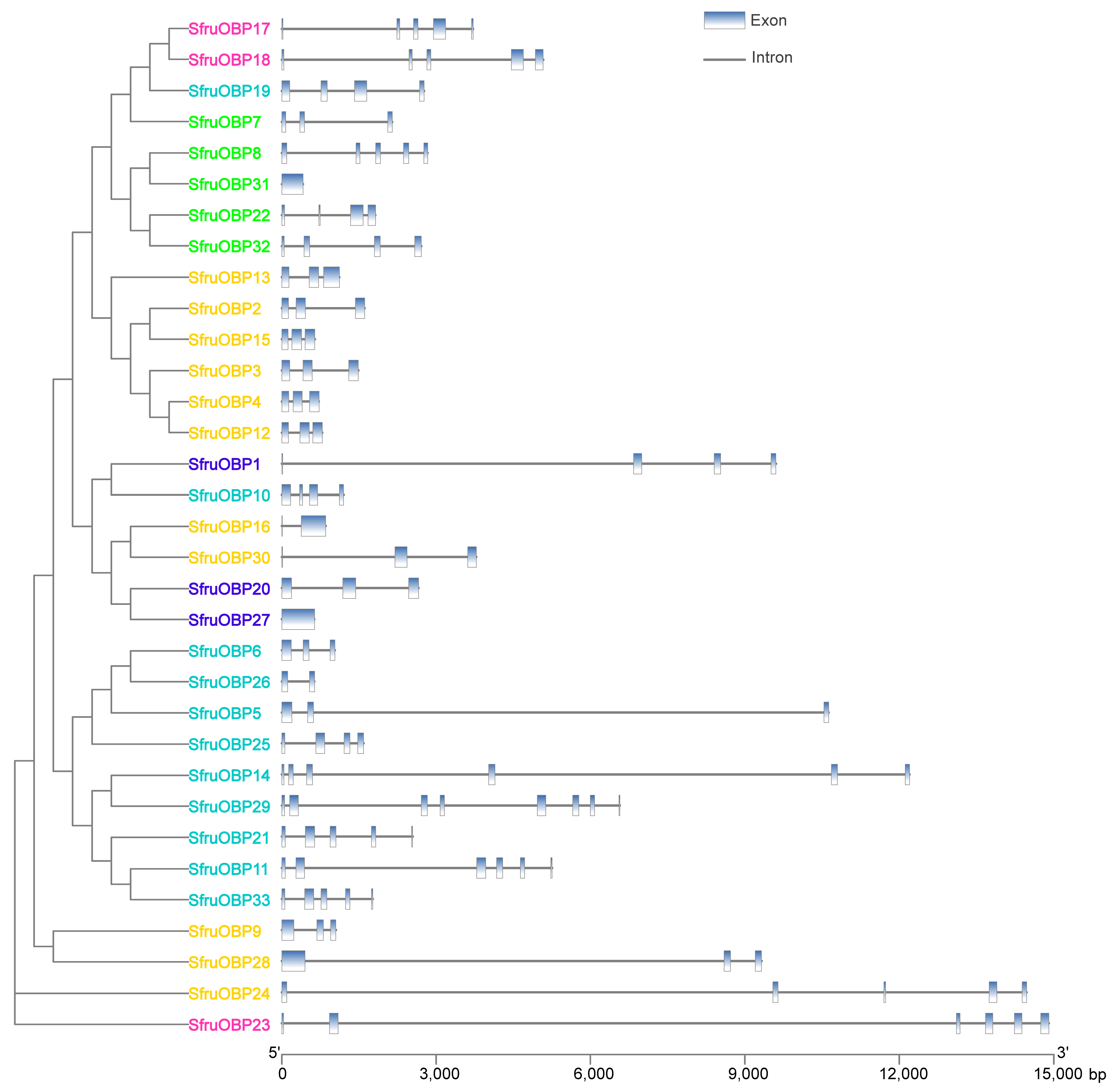
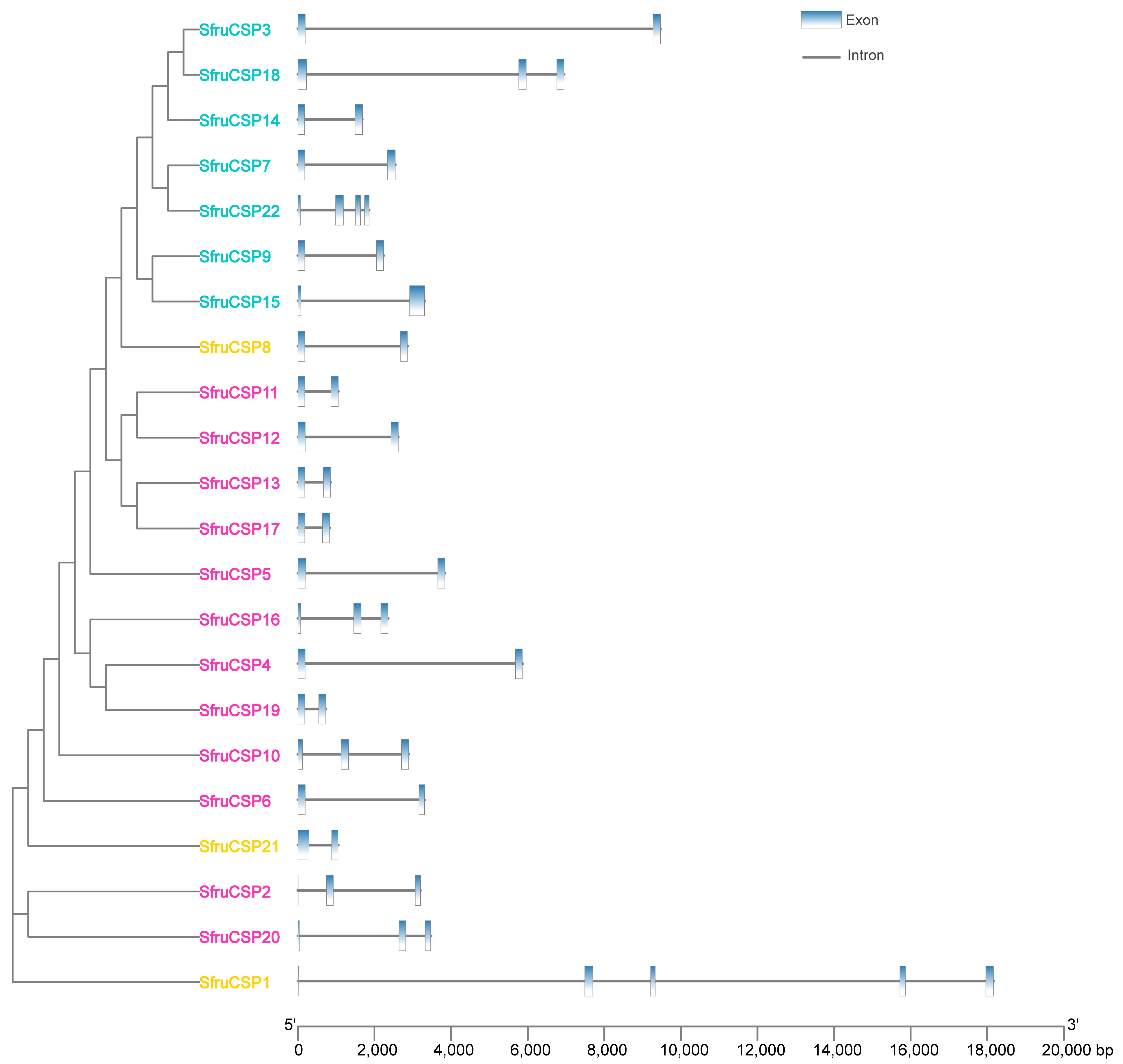
2.4. Intron/Exon Organization of OBP and CSP Genes of S. frugiperda
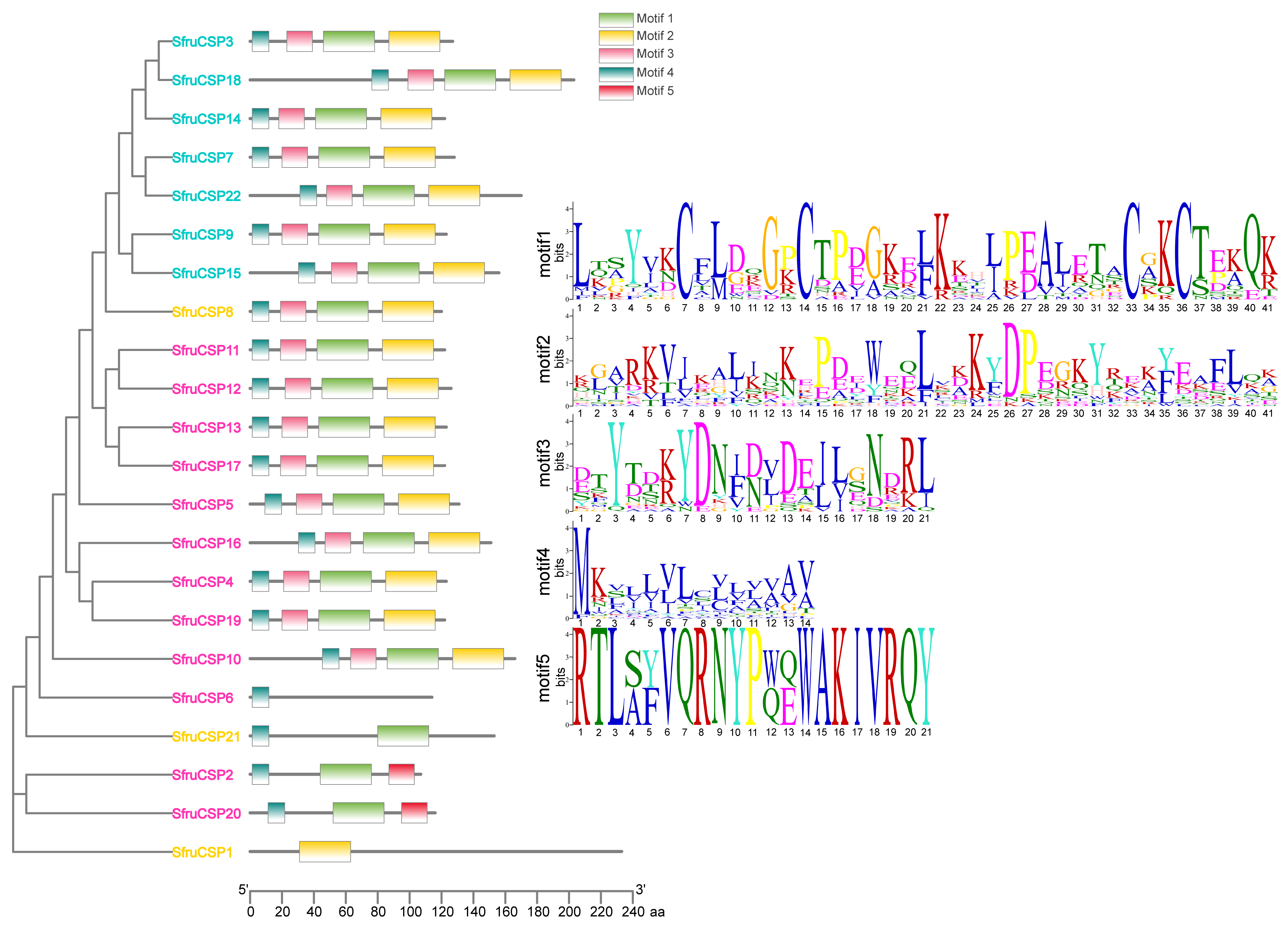
2.5. Motif Analysis of OBPs and CSPs of S. frugiperda
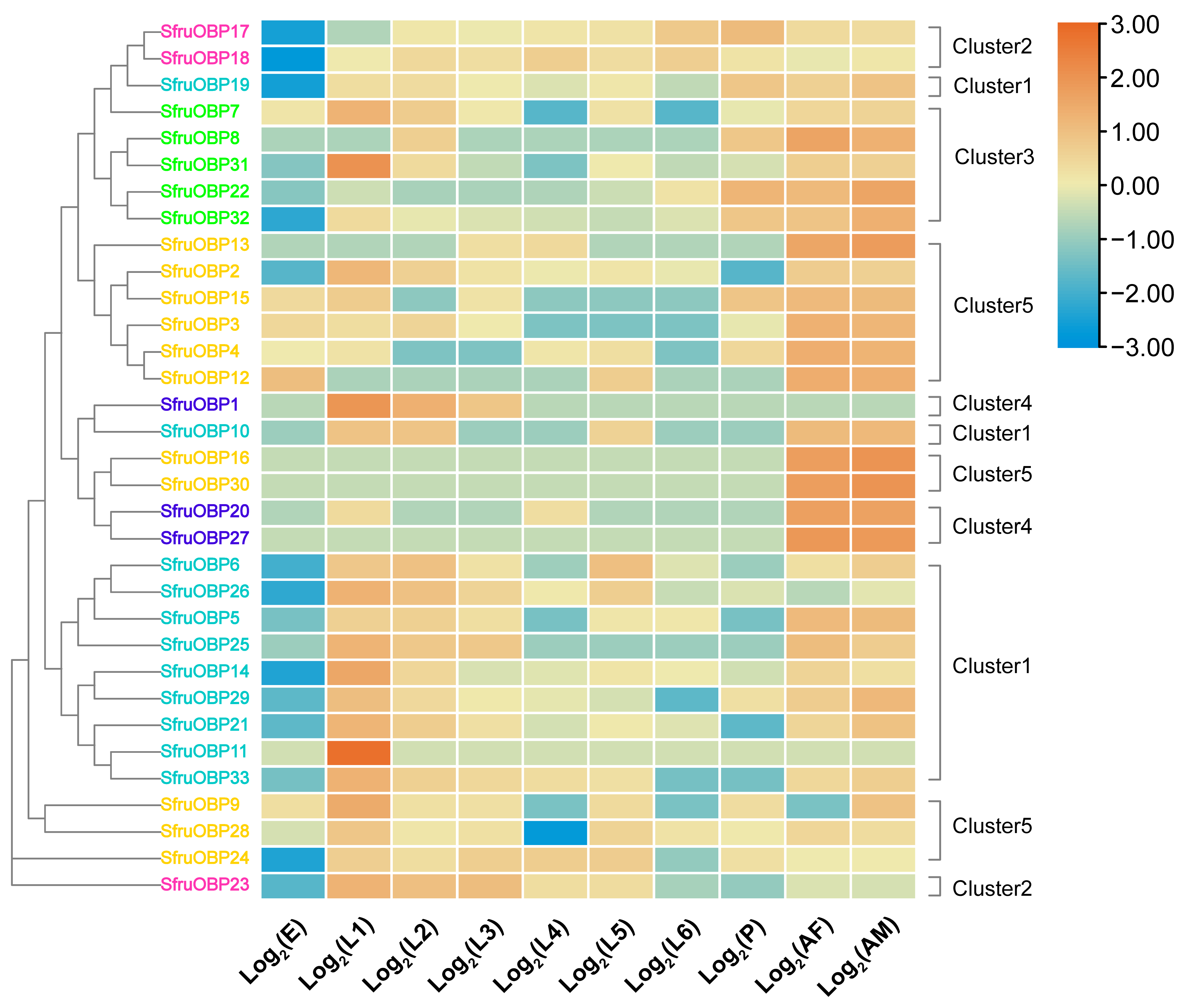
2.6. Gene Expression Analysis of OBPs and CSPs of S. frugiperda
2.7. Binding of SfruOBP31 to Host Volatiles, Pheromones, and Pesticides
3. Discussion
4. Materials and Methods
4.1. Gene Identification
4.2. Sequence Alignment and Phylogenetic Tree Construction
4.3. Chromosomal Distribution, Gene Structure, and Motif Analysis of OBPs and CSPs
4.4. Samples Preparation, RNA Extraction, and RNA-seq
4.5. q-RT-PCR Verification of Selected SfruOBPs and SfruCSPs
4.6. Heterologous Expression and Purification of SfruOBP31 in Bacteria
4.7. Chemical Competitive Binding Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chown, S.L.; Terblanche, J.S. Physiological diversity in insects: Ecological and evolutionary contexts. Adv. Insect Physiol. 2006, 33, 50–152. [Google Scholar] [CrossRef] [Green Version]
- Danks, H.V. The elements of seasonal adaptations in insects. Can. Entomol. 2007, 139, 1–44. [Google Scholar] [CrossRef]
- Van der Goes van Naters, W.; Carlson, J.R. Insects as chemosensors of humans and crops. Nature 2006, 444, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Benton, R. Multigene family evolution: Perspectives from insect chemoreceptors. Trends Ecol. Evol. 2015, 30, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P. Odorant-binding proteins. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 199–228. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Francis, F.; Liu, Y.; Chen, J.L.; Cheng, D.F. An overview of odorant-binding protein functions in insect peripheral olfactory reception. Genet. Mol. Res. 2011, 10, 3056–3069. [Google Scholar] [CrossRef]
- De Bruyne, M.; Warr, C.G. Molecular and cellular organization of insect chemosensory neurons. BioEssays 2006, 28, 23–34. [Google Scholar] [CrossRef]
- Guo, W.; Ren, D.; Zhao, L.; Jiang, F.; Song, J.; Wang, X.; Kang, L. Identification of odorant-binding proteins (OBPs) and functional analysis of phase-related OBPs in the migratory locust. Front. Physiol. 2018, 9, 984. [Google Scholar] [CrossRef] [Green Version]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Vogt, R.G.; Riddiford, L.M.; Prestwich, G.D. Kinetic properties of a sex pheromone-degrading enzyme: The sensillar esterase of Antheraea polyphemus. Proc. Natl. Acad. Sci. USA 1985, 82, 8827–8831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Picimbon, J.-F.; Ji, S.; Kan, Y.; Chuanling, Q.; Zhou, J.-J.; Pelosi, P. Multiple functions of an odorant-binding protein in the mosquito Aedes aegypti. Biochem. Biophys. Res. Commun. 2008, 372, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xuan, N.; Liu, G.; Xie, H.; Lou, Q.; Arnaud, P.; Offmann, B.; Picimbon, J.-F. An expanded survey of the moth PBP/GOBP clade in Bombyx mori: New insight into expression and functional roles. Front. Physiol. 2021, 12, 712593. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, C.; Qiu, C.-L.; Shi, J.-H.; Sun, Z.; Hu, X.-J.; Liu, L.; Wang, M.-Q. A salivary odorant-binding protein mediates Nilaparvata lugens feeding and host plant phytohormone suppression. Int. J. Mol. Sci. 2021, 22, 4988. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.; Mans, B.J.; Ribeiro, J.M.C.; Andersen, J.F. Multifunctionality and mechanism of ligand binding in a mosquito Antiinflammatory Protein. Proc. Natl. Acad. Sci. USA 2009, 106, 3728–3733. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Mao, K.; Ren, Z.; Jin, R.; Zhang, Y.; Cai, T.; He, S.; Li, J.; Wan, H. Odorant binding protein 3 is associated with nitenpyram and sulfoxaflor resistance in Nilaparvata lugens. Int. J. Biol. Macromol. 2022, 209, 1352–1358. [Google Scholar] [CrossRef]
- Zhang, W.; Xie, M.; Eleftherianos, I.; Mohamed, A.; Cao, Y.; Song, B.; Zang, L.-S.; Jia, C.; Bian, J.; Keyhani, N.O.; et al. An odorant binding protein is involved in counteracting detection-avoidance and Toll-pathway innate immunity. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- He, P.; Li, Z.-Q.; Zhang, Y.-F.; Chen, L.; Wang, J.; Xu, L.; Zhang, Y.-N.; He, M. Identification of odorant-binding and chemosensory protein genes and the ligand affinity of two of the encoded proteins suggest a complex olfactory perception system in Periplaneta americana. Insect Mol. Biol. 2017, 26, 687–701. [Google Scholar] [CrossRef]
- Agnihotri, A.; Liu, N.; Xu, W. Chemosensory proteins (CSPs) in the cotton bollworm Helicoverpa armigera. Insects 2022, 13, 29. [Google Scholar] [CrossRef]
- Sabatier, L.; Jouanguy, E.; Dostert, C.; Zachary, D.; Dimarcq, J.-L.; Bulet, P.; Imler, J.-L. Pherokine-2 and -3: Two Drosophila molecules related to pheromone/odor-binding proteins induced by viral and bacterial infections. Eur. J. Biochem. 2003, 270, 3398–3407. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Guo, H.; Huang, L.-Q.; Pelosi, P.; Wang, C.-Z. Unique function of a chemosensory protein in the proboscis of two Helicoverpa species. J. Exp. Biol. 2014, 217, 1821–1826. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Venthur, H.; Wang, S.; Homem, R.A.; Zhou, J.-J. Evidence for the involvement of the chemosensory protein AgosCSP5 in resistance to insecticides in the cotton aphid, Aphis gossypii. Insects 2021, 12, 335. [Google Scholar] [CrossRef] [PubMed]
- Lizana, P.; Mutis, A.; Quiroz, A.; Venthur, H. Insights into chemosensory proteins from non-model insects: Advances and perspectives in the context of pest management. Front. Physiol. 2022, 13, 924750. [Google Scholar] [CrossRef]
- Leal, W.S.; Nikonova, L.; Peng, G. Disulfide structure of the pheromone binding protein from the silkworm moth, Bombyx mori. FEBS Lett. 1999, 464, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Lagarde, A.; Spinelli, S.; Tegoni, M.; He, X.; Field, L.; Zhou, J.-J.; Cambillau, C. The crystal structure of odorant binding protein 7 from Anopheles gambiae exhibits an outstanding adaptability of its binding site. J. Mol. Biol. 2011, 414, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Mao, Y.; Zhang, L. Binding properties of four antennae-expressed chemosensory proteins (CSPs) with insecticides indicates the adaption of Spodoptera litura to environment. Pestic. Biochem. Physiol. 2018, 146, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Falchetto, M.; Ciossani, G.; Scolari, F.; Di Cosimo, A.; Nenci, S.; Field, L.M.; Mattevi, A.; Zhou, J.-J.; Gasperi, G.; Forneris, F. Structural and biochemical evaluation of Ceratitis capitata odorant-binding protein 22 affinity for odorants involved in intersex communication. Insect Mol. Biol. 2019, 28, 431–443. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Du, G.; Ng, C.-S.; Prestwich, G.D. Odorant binding by a pheromone binding protein: Active site mapping by photoaffinity labeling. Biochemistry 1994, 33, 4812–4819. [Google Scholar] [CrossRef]
- Xu, Y.-L.; He, P.; Zhang, L.; Fang, S.-Q.; Dong, S.-L.; Zhang, Y.-J.; Li, F. Large-scale identification of odorant-binding proteins and chemosensory proteins from expressed sequence tags in insects. BMC Genom. 2009, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Li, T.; Liu, J.; Sun, M.; Chen, X.; Liu, Y.; Xu, P. Sex-and stage-dependent expression patterns of odorant-binding and chemosensory protein genes in Spodoptera exempta. PeerJ 2021, 9, e12132. [Google Scholar] [CrossRef] [PubMed]
- Honson, N.S.; Plettner, E. Disulfide connectivity and reduction in pheromone-binding proteins of the gypsy moth, Lymantria dispar. Naturwissenschaften 2006, 93, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.-P.; Zhang, H.; Zhao, P.; Lin, Y.; Xia, Q.-Y.; Xiang, Z.-H. Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2007, 37, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Jacquin-Joly, E. Functional and expression pattern analysis of chemosensory proteins expressed in antennae and pheromonal gland of Mamestra brassicae. Chem. Senses 2001, 26, 833–844. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Field, L.M.; He, X.L. Insect odorant-binding proteins: Do they offer an alternative pest control strategy? Outlooks Pest Manag. 2010, 21, 31–34. [Google Scholar] [CrossRef]
- Wanchoo, A.; Zhang, W.; Ortiz-Urquiza, A.; Boswell, J.; Xia, Y.; Keyhani, N.O. Red imported fire ant (Solenopsis invicta) chemosensory proteins are expressed in tissue, developmental, and caste-specific patterns. Front. Physiol. 2020, 11, 585883. [Google Scholar] [CrossRef]
- Bista, S.; Thapa, M.K.; Khanal, S. Fall armyworm: Menace to nepalese farming and the integrated management approaches. Int. J. Agric. Environ. Biotechnol. 2020, 5, 1001–1018. [Google Scholar] [CrossRef]
- Ríos-Díez, J.D.; Saldamando-Benjumea, C.I. Susceptibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) strains from central Colombia to two insecticides, methomyl and lambda-cyhalothrin: A study of the genetic basis of resistance. J. Econ. Entomol. 2011, 104, 1698–1705. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Luttrell, R.G.; Pitre, H.N. Feeding sites and distribution of Fall armyworm (Lepidoptera: Noctuidae) larvae on cotton. Environ. Entomol. 1990, 19, 1060–1067. [Google Scholar] [CrossRef]
- Yu, S.J.; Nguyen, S.N.; Abo-Elghar, G.E. Biochemical characteristics of insecticide resistance in the Fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Physiol. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Tang, X.; Lyu, B.; Lu, H.; Tang, J.; Zhang, Y. Effects of cannibalism on the growth and development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Int. J. Pest Manag. 2022, 1–11. [Google Scholar] [CrossRef]
- Yang, C.; Belawat, P.; Hafen, E.; Jan, L.Y.; Jan, Y.-N. Drosophila egg-laying site selection as a system to study simple decision-making processes. Science 2008, 319, 1679–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Ye, X.; Xu, H.; Mei, Y.; Yang, Y.; Chen, X.; Yang, Y.; Liu, T.; Yu, Y.; Yang, W.; et al. The genetic adaptations of Fall armyworm Spodoptera frugiperda facilitated its rapid global dispersal and invasion. Mol. Ecol. Resour. 2020, 20, 1050–1068. [Google Scholar] [CrossRef]
- Legeai, F.; Gimenez, S.; Duvic, B.; Escoubas, J.-M.; Gosselin Grenet, A.-S.; Blanc, F.; Cousserans, F.; Séninet, I.; Bretaudeau, A.; Mutuel, D.; et al. Establishment and analysis of a reference transcriptome for Spodoptera frugiperda. BMC Genom. 2014, 15, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; He, L.; Tan, X.; Zhang, Z.; Wang, Y.; Li, X.; He, H.; Ding, W.; Li, Y. Identification and phylogenetics of Spodoptera frugiperda chemosensory proteins based on antennal transcriptome data. Comp. Biochem. Physiol. D Genom. Proteom. 2020, 34, 100680. [Google Scholar] [CrossRef] [PubMed]
- Bentzur, A.; Shmueli, A.; Omesi, L.; Ryvkin, J.; Knapp, J.-M.; Parnas, M.; Davis, F.P.; Shohat-Ophir, G. Odorant binding protein 69a connects social interaction to modulation of social responsiveness in Drosophila. PLoS Genet. 2018, 14, e1007328. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Große-Wilde, E.; Gohl, T.; Bouché, E.; Breer, H.; Krieger, J. Candidate pheromone receptors provide the basis for the response of distinct antennal neurons to pheromonal compounds. Eur. J. Neurosci. 2007, 25, 2364–2373. [Google Scholar] [CrossRef]
- Montagné, N.; Chertemps, T.; Brigaud, I.; François, A.; François, M.-C.; de Fouchier, A.; Lucas, P.; Larsson, M.C.; Jacquin-Joly, E. Functional characterization of a sex pheromone receptor in the pest moth Spodoptera littoralis by heterologous expression in Drosophila. Eur. J. Neurosci. 2012, 36, 2588–2596. [Google Scholar] [CrossRef]
- Cattaneo, A.M.; Gonzalez, F.; Bengtsson, J.M.; Corey, E.A.; Jacquin-Joly, E.; Montagné, N.; Salvagnin, U.; Walker, W.B.; Witzgall, P.; Anfora, G.; et al. Candidate pheromone receptors of codling moth Cydia pomonella respond to pheromones and kairomones. Sci. Rep. 2017, 7, 41105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattaneo, A.M. Current status on the functional characterization of chemosensory receptors of Cydia pomonella (Lepidoptera: Tortricidae). Front. Behav. Neurosci. 2018, 12, 189. [Google Scholar] [CrossRef]
- Vogt, R.G.; Große-Wilde, E.; Zhou, J.J. The Lepidoptera Odorant Binding Protein gene family: Gene gain and loss within the GOBP/PBP complex of moths and butterflies. Insect Biochem. Mol. Biol. 2015, 62, 142–153. [Google Scholar] [CrossRef]
- Crava, C.M.; Bobkov, Y.V.; Sollai, G.; Anfora, G.; Crnjar, R.; Cattaneo, A.M. Chemosensory receptors in the larval maxilla of Papilio hospiton. Front. Ecol. Evol. 2022, 9, 795994. [Google Scholar] [CrossRef]
- Nagnan-Le Meillour, P.; Cain, A.H.; Jacquin-Joly, E.; François, M.C.; Ramachandran, S.; Maida, R.; Steinbrecht, R.A. Chemosensory proteins from the proboscis of Mamestra brassicae. Chem. Senses 2000, 25, 541–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, K.; Utoguchi, A.; Yamada, A.; Yoshikawa, H. Identification and genomic structure of chemosensory proteins (CSP) and odorant binding proteins (OBP) genes expressed in foreleg tarsi of the swallowtail butterfly Papilio xuthus. Insect Biochem. Mol. Biol. 2008, 38, 969–976. [Google Scholar] [CrossRef]
- Gong, D.P.; Zhang, H.J.; Zhao, P.; Xia, Q.Y.; Xiang, Z.H. The odorant binding protein gene family from the genome of silkworm, Bombyx mori. BMC Genom. 2009, 10, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlmeier, P.; Billeter, J. Genetic mechanisms modulating behaviour through plastic chemosensory responses in insects. Mol. Ecol. 2022, 32, 45–60. [Google Scholar] [CrossRef]
- Liu, N.-Y.; Zhang, T.; Ye, Z.-F.; Li, F.; Dong, S.-L. Identification and characterization of candidate chemosensory gene families from Spodoptera exigua developmental transcriptomes. Int. J. Biol. Sci. 2015, 11, 1036–1048. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-L.; Xu, J.-W.; Yao, W.-C.; Yang, H.-H.; Dewer, Y.; Zhang, F.; Zhu, X.-Y.; Zhang, Y.-N. Chemosensory genes in the head of Spodoptera litura larvae. Bull. Entomol. Res. 2021, 111, 454–463. [Google Scholar] [CrossRef]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.-H.; Zhou, J.-J.; Gao, S.; Wang, D.-H.; Li, X.-C.; Guo, Y.-Y.; Zhang, Y.-J. Identification and comparative expression analysis of odorant binding protein genes in the tobacco cutworm Spodoptera litura. Sci. Rep. 2015, 5, 13800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, W.B.; Roy, A.; Anderson, P.; Schlyter, F.; Hansson, B.S.; Larsson, M.C. Transcriptome analysis of gene families involved in chemosensory function in Spodoptera littoralis (Lepidoptera: Noctuidae). BMC Genom. 2019, 20, 428. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.G.; Rozas, J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the Arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondón, J.J.; Moreyra, N.N.; Pisarenco, V.A.; Rozas, J.; Hurtado, J.; Hasson, E. Evolution of the odorant-binding protein gene family in Drosophila. Front. Ecol. Evol. 2022, 10, 957247. [Google Scholar] [CrossRef]
- Garczynski, S.F.; Cattaneo, A.M.; Walker, W.B. Alternative splicing of the CpomOR53 gene produces three transcripts expressed in codling moth (Lepidoptera: Tortricidae) Antennae. J. Econ. Entomol. 2019, 112, 991–996. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, A.M.; Bengtsson, J.M.; Montagné, N.; Jacquin-Joly, E.; Rota-Stabelli, O.; Salvagnin, U.; Bassoli, A.; Witzgall, P.; Anfora, G. TRPA5, an ankyrin subfamily insect TRP channel, is expressed in antennae of Cydia pomonella (Lepidoptera: Tortricidae) in multiple splice variants. J. Insect Sci. 2016, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Rooney, A.P. Concerted and birth-and-death evolution of multigene families. Annu. Rev. Genet. 2005, 39, 121–152. [Google Scholar] [CrossRef] [Green Version]
- Vieira, F.G.; Sanchez-Gracia, A.; Rozas, J. Comparative genomic analysis of the odorant-binding protein family in 12 Drosophila genomes: Purifying selection and birth-and-death evolution. Genome Biol. 2007, 8, R235. [Google Scholar] [CrossRef] [Green Version]
- Pracana, R.; Levantis, I.; Martínez-Ruiz, C.; Stolle, E.; Priyam, A.; Wurm, Y. Fire ant social chromosomes: Differences in number, sequence and expression of odorant binding proteins. Evol. Lett. 2017, 1, 199–210. [Google Scholar] [CrossRef]
- Singh, S.; Tyagi, C.; Rather, I.A.; Sabir, J.S.M.; Hassan, M.I.; Singh, A.; Singh, I.K. Molecular modeling of chemosensory protein 3 from Spodoptera litura and its binding property with plant defensive metabolites. Int. J. Mol. Sci. 2020, 21, 4073. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Guidolin, A.; Syed, Z.; Cornel, A.J.; Leal, W.S. Knockdown of a mosquito odorant-binding protein involved in the sensitive detection of oviposition attractants. J. Chem. Ecol. 2010, 36, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhou, Y.-T.; Tan, Y.; Zhou, X.-R.; Pang, B.-P. Identification of odorant-binding protein genes in Galeruca daurica (Coleoptera: Chrysomelidae) and analysis of their expression profiles. Bull. Entomol. Res. 2017, 107, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Rong, J.; Liu, N.-y.; Liu, Y.; Dong, S.-l. A larval specific OBP able to bind the major female sex pheromone component in Spodoptera exigua (Hübner). J. Integr. Agric. 2015, 14, 1356–1366. [Google Scholar] [CrossRef] [Green Version]
- De Biasio, F.; Riviello, L.; Bruno, D.; Grimaldi, A.; Congiu, T.; Sun, Y.F.; Falabella, P. Expression pattern analysis of odorant-binding proteins in the pea aphid Acyrthosiphon Pisum. Insect Sci. 2014, 22, 220–234. [Google Scholar] [CrossRef]
- Vandermoten, S.; Francis, F.; Haubruge, E.; Leal, W.S. Conserved odorant-binding proteins from aphids and eavesdropping predators. PLoS ONE 2011, 6, e23608. [Google Scholar] [CrossRef] [Green Version]
- Kitabayashi, A.N.; Arai, T.; Kubo, T.; Natori, S. Molecular cloning of cDNA for p10, a novel protein that increases in the regenerating legs of Periplaneta americana (American cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Smadja, C.; Butlin, R.K. On the scent of speciation: The chemosensory system and its role in premating isolation. Heredity 2008, 102, 77–97. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Zhou, X.; Gu, F.; Zhong, G.; Yi, X. Olfactory plasticity: Variation in the expression of chemosensory receptors in Bactrocera dorsalis in different physiological states. Front. Physiol. 2017, 8, 672. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.-H.; Wang, S.-Y.; Zhang, X.-Y.; Ji, P.; Liu, J.-T.; Wang, G.-R.; Wu, K.-M.; Guo, Y.-Y.; Zhou, J.-J.; Zhang, Y.-J. Functional characterizations of chemosensory proteins of the alfalfa plant bug Adelphocoris lineolatus indicate their involvement in host recognition. PLoS ONE 2012, 7, e42871. [Google Scholar] [CrossRef]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory proteins in the honey bee: Insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Maleszka, J.; Forêt, S.; Saint, R.; Maleszka, R. RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev. Genes Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Li, L.; Yao, W.-C.; Yin, M.-Z.; Li, J.-Q.; Xu, J.-W.; Dewer, Y.; Zhu, X.-Y.; Zhang, Y.-N. Two odorant-binding proteins involved in the recognition of sex pheromones in Spodoptera litura larvae. J. Agric. Food Chem. 2022, 70, 12372–12382. [Google Scholar] [CrossRef]
- Yi, X.; Qi, J.; Zhou, X.; Hu, M.Y.; Zhong, G.H. Differential expression of chemosensory-protein genes in midguts in response to diet of Spodoptera litura. Sci. Rep. 2017, 7, 296. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, X.; Liu, J.; Hu, M.; Zhong, G.; Geng, P.; Yi, X. Molecular cloning, expression and molecular modeling of chemosensory protein from Spodoptera litura and its binding properties with Rhodojaponin III. PLoS ONE 2012, 7, e47611. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Jiang, Y.; Zhang, L.; Cai, Y. Effects of insecticides chlorpyrifos, emamectin benzoate and fipronil on Spodoptera litura might be mediated by OBPs and CSPs. Bull. Entomol. Res. 2018, 108, 658–666. [Google Scholar] [CrossRef]
- Sokame, B.M.; Subramanian, S.; Kilalo, D.C.; Juma, G.; Calatayud, P.A. Larval dispersal of the invasive Fall armyworm, Spodoptera frugiperda, the exotic stemborer Chilo partellus, and indigenous maize stemborers in Africa. Entomol. Exp. Appl. 2020, 168, 322–331. [Google Scholar] [CrossRef]
- Van den Berg, J.; du Plessis, H. Chemical control and insecticide resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2022, 115, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Murúa, M.G.; Vera, M.T.; Abraham, S.; Juárez, M.L.; Prieto, S.; Head, G.P.; Willink, E. Fitness and mating compatibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) populations from different host plant species and regions in Argentina. Ann. Entomol. Soc. Am. 2008, 101, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Sun, X.-T.; Ge, S.-S.; Yang, X.-M.; Wu, K.-M. Mating competitiveness of male Spodoptera frugiperda (Smith) irradiated by X-rays. Insects 2023, 14, 137. [Google Scholar] [CrossRef]
- Tumlinson, J.H.; Mitchell, E.R.; Teal, P.E.A.; Heath, R.R.; Mengelkoch, L.J. Sex pheromone of Fall armyworm, Spodoptera frugiperda (J.E. Smith). J. Chem. Ecol. 1986, 12, 1909–1926. [Google Scholar] [CrossRef] [PubMed]
- De Bruyne, M.; Baker, T.C. Odor detection in insects: Volatile codes. J. Chem. Ecol. 2008, 34, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.J.; Schmelz, E.A.; Meagher, R.L.; Teal, P.E.A. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings. J. Chem. Ecol. 2006, 32, 1911–1924. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Wang, R.; Du, Y.; Gao, B.; Gui, F.; Lu, K. Olfactory perception of herbicide butachlor by GOBP2 elicits ecdysone biosynthesis and detoxification enzyme responsible for chlorpyrifos tolerance in Spodoptera litura. Environ. Pollut. 2021, 285, 117409. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.; Ma, Z.; Zhang, Y.; Yan, S.; Shen, J. RNA interference cannot be operated in lepidopteran insect? A nanocarrier breaks bottlenecks at all developmental stages of Spodoptera frugiperda. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Li, J.-J.; Shi, Y.; Wu, J.-N.; Li, H.; Smagghe, G.; Liu, T.-X. CRISPR/Cas9 in in lepidopteran insects: Progress, application and prospects. J. Insect Physiol. 2021, 135, 104325. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; Keyhani, N.O.; Zhang, Z.; Li, S.; Xia, Y. Comparative transcriptomic analysis of immune responses of the migratory locust, Locusta migratoria, to challenge by the fungal insect pathogen, Metarhizium acridum. BMC Genom. 2015, 16, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Onofrio, C.; Zaremska, V.; Zhu, J.; Knoll, W.; Pelosi, P. Ligand-binding assays with OBPs and CSPs. Methods Enzymol. 2020, 642, 229–258. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genome ID | Chromosome No. | Length (bp) | Introns | Exons | Amino Acids (aa) | Molecular Weight (kDa) | Isoelectric Point | Signal Peptide (aa) |
|---|---|---|---|---|---|---|---|---|---|
| OBP1 | Sfru136920 | chr15 | 9606 | 2 | 3 | 131 | 14.90 | 9.01 | ND |
| OBP2 | Sfru112680 | chr8 | 1607 | 2 | 3 | 162 | 18.23 | 5.31 | 1-21 |
| OBP3 | Sfru112690 | chr8 | 1488 | 2 | 3 | 173 | 19.14 | 5.16 | 1-30 |
| OBP4 | Sfru112730 | chr8 | 722 | 2 | 3 | 164 | 18.75 | 5.47 | 1-23 |
| OBP5 | Sfru220490 | chr10 | 10,632 | 2 | 3 | 146 | 16.16 | 4.47 | 1-21 |
| OBP6 | Sfru220520 | chr10 | 1035 | 2 | 3 | 147 | 15.73 | 5.42 | 1-21 |
| OBP7 | Sfru220480 | chr10 | 2148 | 2 | 3 | 121 | 13.97 | 8.15 | ND |
| OBP8 | Sfru131490 | chr2 | 2834 | 4 | 5 | 149 | 17.22 | 4.63 | 1-26 |
| OBP9 | Sfru078880 | chr12 | 1052 | 2 | 3 | 155 | 18.22 | 4.88 | ND |
| OBP10 | Sfru072390 | chr20 | 1203 | 3 | 4 | 118 | 12.69 | 8.72 | 1-20 |
| OBP11 | Sfru220500 | chr10 | 5252 | 5 | 6 | 211 | 23.91 | 5.52 | 1-23 |
| OBP12 | Sfru112710 | chr8 | 783 | 2 | 3 | 164 | 18.74 | 5.36 | 1-19 |
| OBP13 | Sfru112700 | chr8 | 1121 | 2 | 3 | 209 | 23.92 | 8.07 | 1-22 |
| OBP14 | Sfru220470 | chr10 | 12,205 | 5 | 6 | 191 | 21.37 | 5.84 | ND |
| OBP15 | Sfru112570 | chr8 | 644 | 2 | 3 | 164 | 19.27 | 5.51 | 1-19 |
| OBP16 | Sfru136280 | chr15 | 852 | 1 | 2 | 159 | 18.58 | 8.46 | 1-19 |
| OBP17 | Sfru036850 | chr22 | 3719 | 4 | 5 | 141 | 15.70 | 6.86 | ND |
| OBP18 | Sfru153150 | chr22 | 5083 | 4 | 5 | 135 | 14.44 | 8.96 | ND |
| OBP19 | Sfru036860 | chr22 | 2766 | 3 | 4 | 202 | 22.75 | 5.12 | 1-19 |
| OBP20 | Sfru194150 | chr15 | 2659 | 2 | 3 | 160 | 18.14 | 9.97 | ND |
| OBP21 | Sfru220550 | chr10 | 2544 | 4 | 5 | 154 | 16.98 | 4.89 | 1-23 |
| OBP22 | Sfru221430 | chr25 | 1817 | 3 | 4 | 156 | 16.93 | 7.51 | 1-22 |
| OBP23 | Sfru131450 | chr2 | 14,905 | 5 | 6 | 241 | 27.63 | 5.82 | ND |
| OBP24 | Sfru032040 | chr3 | 14,476 | 4 | 5 | 156 | 17.81 | 6.38 | ND |
| OBP25 | Sfru220530 | chr10 | 1592 | 3 | 4 | 152 | 16.32 | 4.69 | 1-21 |
| OBP26 | Sfru220540 | chr10 | 635 | 1 | 2 | 148 | 16.15 | 4.89 | ND |
| OBP27 | Sfru116380 | ctg319_4 | 635 | 0 | 1 | 211 | 24.78 | 6.91 | ND |
| OBP28 | Sfru007560 | chr8 | 9327 | 2 | 3 | 244 | 26.94 | 5.61 | 1-18 |
| OBP29 | Sfru220560 | chr10 | 6574 | 7 | 8 | 272 | 30.59 | 5.16 | 1-20 |
| OBP30 | Sfru136290 | chr15 | 3781 | 2 | 3 | 136 | 15.53 | 7.63 | 1-19 |
| OBP31 | Sfru155250 | chr14 | 413 | 0 | 1 | 137 | 14.71 | 4.36 | 1-20 |
| OBP32 | Sfru139020 | chr25 | 2713 | 3 | 4 | 133 | 15.09 | 8.88 | 1-16 |
| OBP33 | Sfru220510 | chr10 | 1769 | 4 | 5 | 153 | 17.10 | 4.72 | 1-21 |
| Gene Name | Genome ID | Chromosome No. | Length (bp) | Introns | Exons | Amino Acids (aa) | Molecular Weight (kDa) | Isoelectric Point | Signal Peptide (aa) |
|---|---|---|---|---|---|---|---|---|---|
| CSP1 | Sfru089520 | chr8 | 18,161 | 4 | 5 | 233 | 25.59 | 10.44 | ND |
| CSP2 | Sfru158600 | chr19 | 3201 | 2 | 3 | 107 | 11.94 | 9.49 | 1-22 |
| CSP3 | Sfru069670 | chr8 | 9467 | 1 | 2 | 127 | 14.80 | 6.74 | 1-18 |
| CSP4 | Sfru007770 | chr8 | 5864 | 1 | 2 | 123 | 13.57 | 5.23 | 1-18 |
| CSP5 | Sfru069550 | chr8 | 3841 | 1 | 2 | 131 | 15.61 | 9.42 | 1-25 |
| CSP6 | Sfru007800 | chr8 | 3306 | 1 | 2 | 114 | 12.92 | 5.04 | 1-16 |
| CSP7 | Sfru069640 | chr8 | 2542 | 1 | 2 | 128 | 14.60 | 5.43 | 1-16 |
| CSP8 | Sfru069600 | chr8 | 2859 | 1 | 2 | 120 | 13.77 | 5.86 | 1-16 |
| CSP9 | Sfru069650 | chr8 | 2240 | 1 | 2 | 123 | 14.36 | 6.82 | 1-18 |
| CSP10 | Sfru069590 | chr8 | 2891 | 2 | 3 | 166 | 18.59 | 9.57 | ND |
| CSP11 | Sfru007780 | chr8 | 1057 | 1 | 2 | 122 | 13.82 | 8.95 | 1-16 |
| CSP12 | Sfru007430 | chr8 | 2617 | 1 | 2 | 126 | 14.10 | 7.65 | 1-15 |
| CSP13 | Sfru007750 | chr8 | 853 | 1 | 2 | 123 | 13.78 | 8.77 | 1-16 |
| CSP14 | Sfru069660 | chr8 | 1679 | 1 | 2 | 122 | 14.20 | 6.12 | 1-16 |
| CSP15 | Sfru069630 | chr8 | 3308 | 1 | 2 | 156 | 18.30 | 9.04 | ND |
| CSP16 | Sfru007790 | chr8 | 2358 | 2 | 3 | 151 | 17.14 | 5.61 | ND |
| CSP17 | Sfru069620 | chr8 | 828 | 1 | 2 | 122 | 14.03 | 5.84 | 1-17 |
| CSP18 | Sfru069680 | chr8 | 6950 | 2 | 3 | 203 | 23.31 | 6.90 | ND |
| CSP19 | Sfru158610 | chr19 | 3462 | 2 | 3 | 116 | 12.87 | 9.38 | 1-29 |
| CSP20 | Sfru069530 | chr8 | 735 | 1 | 2 | 122 | 13.62 | 5.14 | 1-17 |
| CSP21 | Sfru069700 | chr8 | 1054 | 1 | 2 | 153 | 17.12 | 8.86 | ND |
| CSP22 | Sfru069690 | chr8 | 1857 | 3 | 4 | 170 | 19.22 | 4.93 | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, C.; Mohamed, A.; Cattaneo, A.M.; Huang, X.; Keyhani, N.O.; Gu, M.; Zang, L.; Zhang, W. Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides. Int. J. Mol. Sci. 2023, 24, 5595. https://doi.org/10.3390/ijms24065595
Jia C, Mohamed A, Cattaneo AM, Huang X, Keyhani NO, Gu M, Zang L, Zhang W. Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides. International Journal of Molecular Sciences. 2023; 24(6):5595. https://doi.org/10.3390/ijms24065595
Chicago/Turabian StyleJia, Chen, Amr Mohamed, Alberto Maria Cattaneo, Xiaohua Huang, Nemat O. Keyhani, Maiqun Gu, Liansheng Zang, and Wei Zhang. 2023. "Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides" International Journal of Molecular Sciences 24, no. 6: 5595. https://doi.org/10.3390/ijms24065595
APA StyleJia, C., Mohamed, A., Cattaneo, A. M., Huang, X., Keyhani, N. O., Gu, M., Zang, L., & Zhang, W. (2023). Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides. International Journal of Molecular Sciences, 24(6), 5595. https://doi.org/10.3390/ijms24065595