
Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease

 , , , and
, , , and
Abstract
:1. Introduction
2. Bart’s Hydrops Fetalis: Clinical Perspectives and Current Diagnostic Procedures
Current Biomarkers for Hemoglobin Bart’s Disease
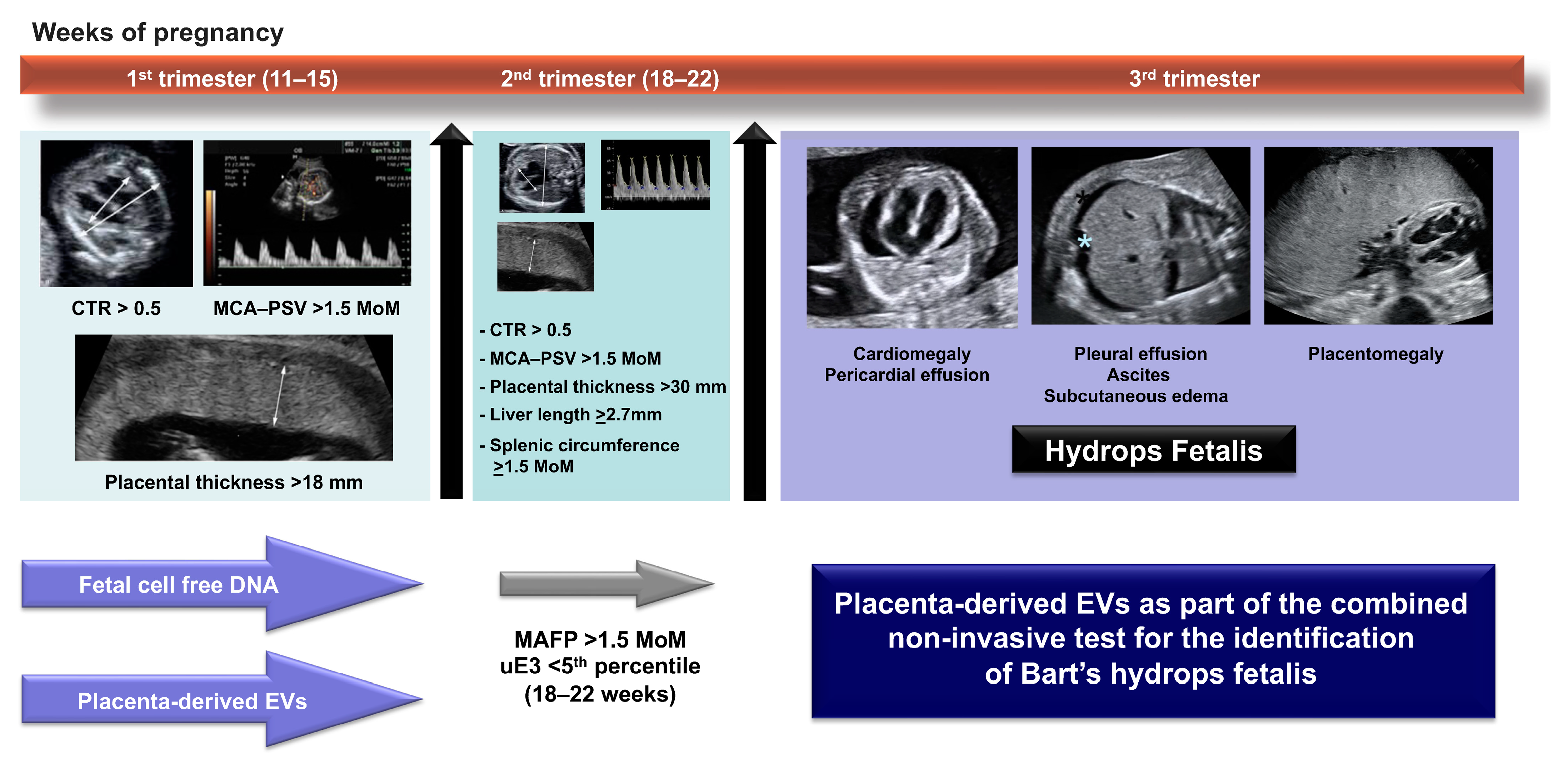
- a cardiac diameter/thoracic diameter ratio (CTR) with a cut-off value of 0.5 at mid-pregnancy;
- a high peak systolic velocity of the middle cerebral artery (MCA-PSV) with a cut-off value of 1.5 multiple of median (MoM), and;
- an increase in the placental thickness with a cut off value of 1.8 cm in the late first trimester and 3 cm at mid-pregnancy.
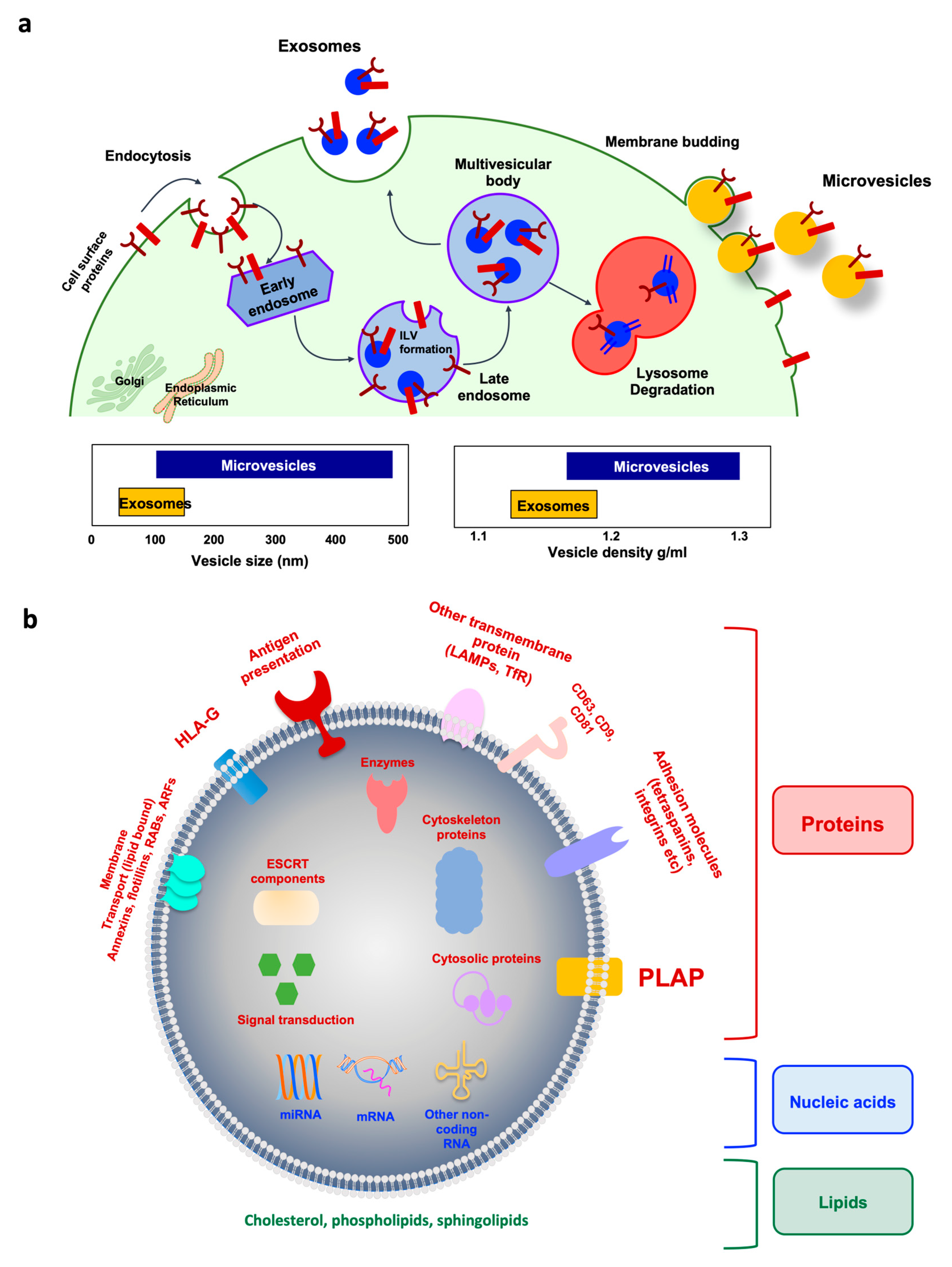
3. Extracellular Vesicles (EVs) as a Source of Non-Invasive Liquid Biopsy
3.1. Extracellular Vesicle Subpopulations
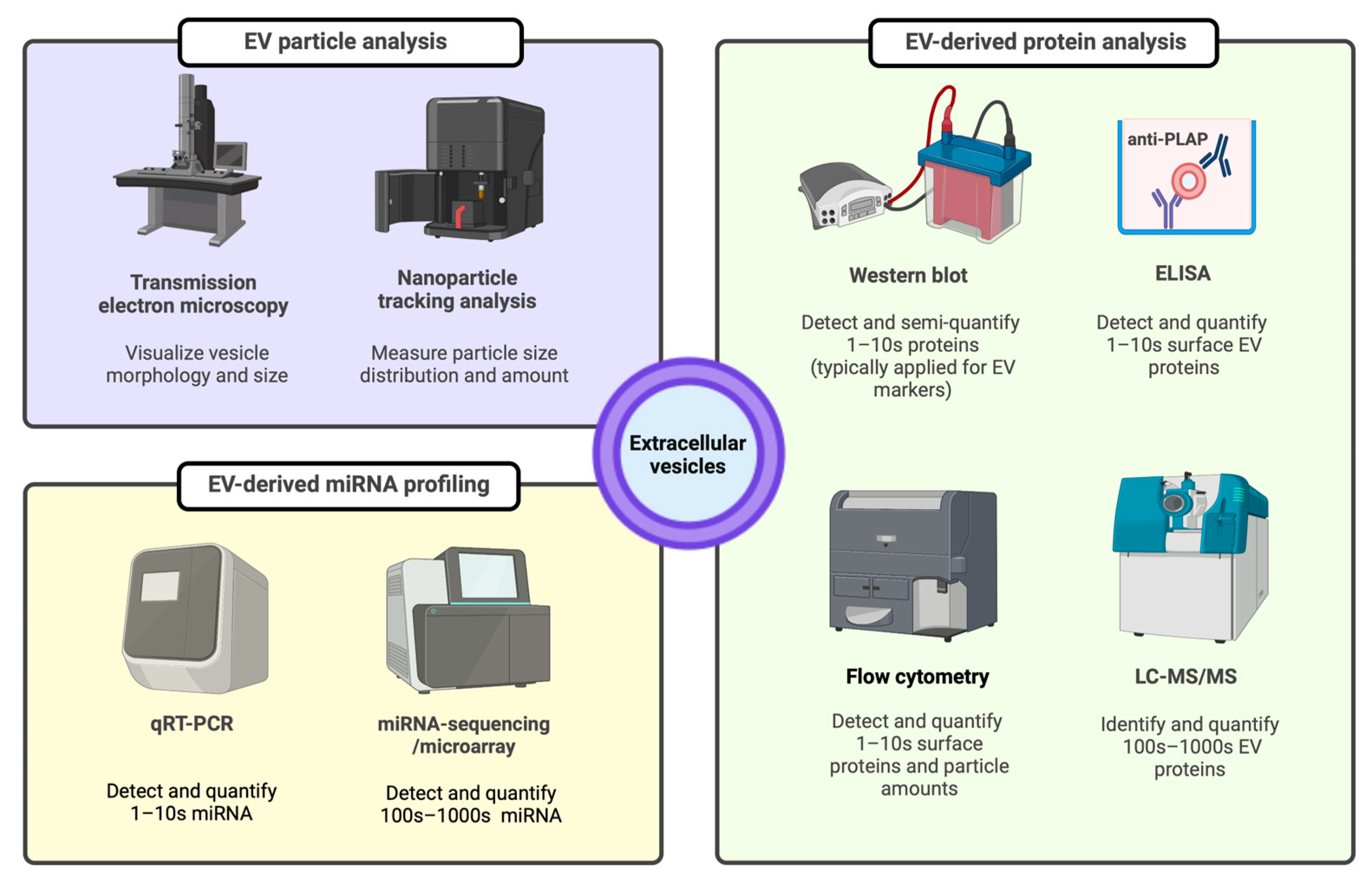
3.2. EV Isolation and Characterization
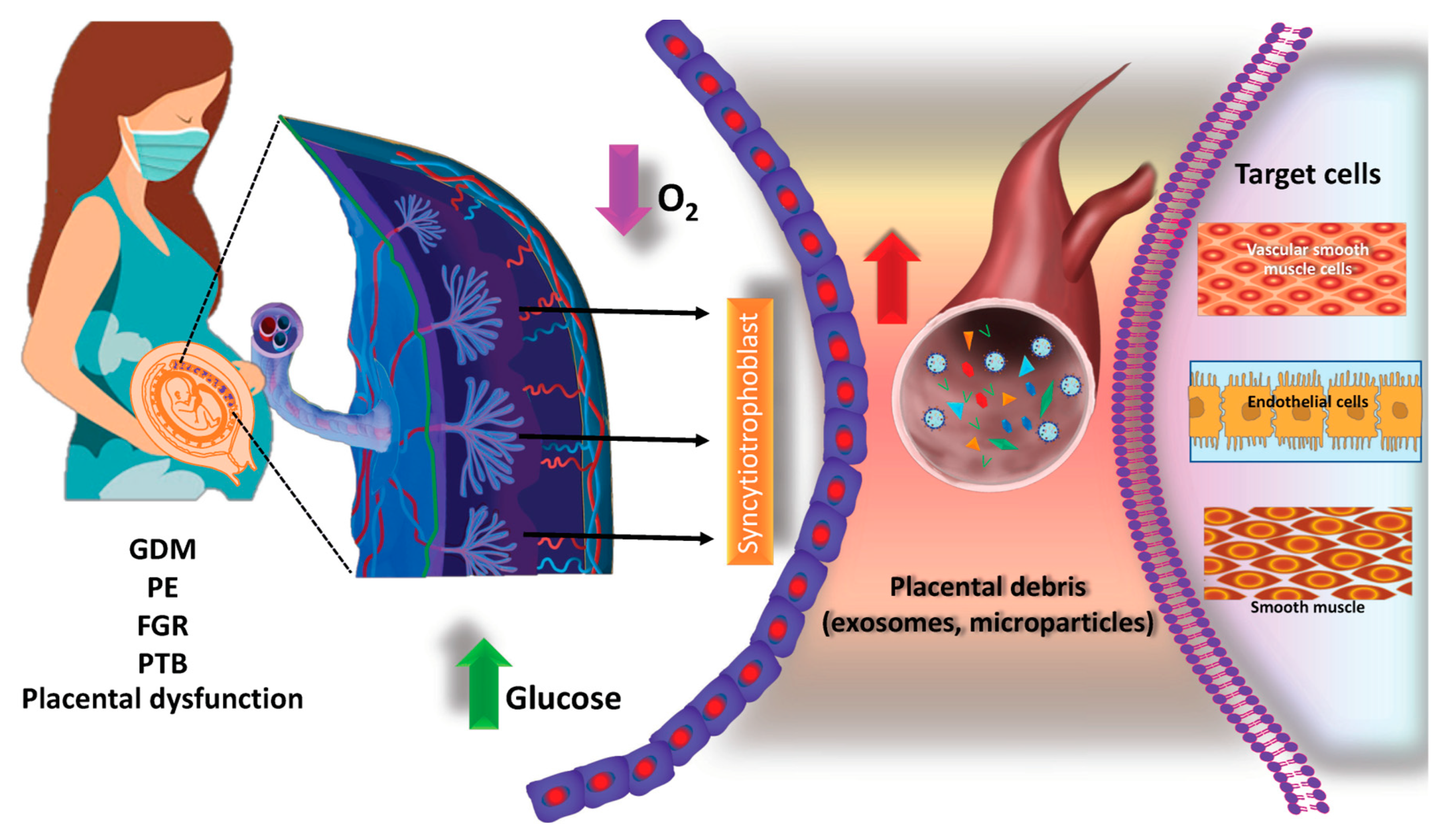
3.3. Placenta-Derived EVs
3.3.1. Regulating Trophoblast Migration and Angiogenesis
3.3.2. Promoting Maternal–Fetal Immune Tolerance
3.3.3. Mediating Maternal–Fetal Intercommunication
3.3.4. Regulating Maternal Metabolic Homeostasis
3.4. Pathophysiological Roles of Placenta-Derived EVs in Pregnancy Complications
4. Placenta-Derived EVs as Hb Bart’s Liquid Biopsy: Challenges and Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flint, J.; Hill, A.V.; Bowden, D.K.; Oppenheimer, S.J.; Sill, P.R.; Serjeantson, S.W.; Bana-Koiri, J.; Bhatia, K.; Alpers, M.P.; Boyce, A.J.; et al. High frequencies of alpha-thalassaemia are the result of natural selection by malaria. Nature 1986, 321, 744–750. [Google Scholar] [CrossRef]
- Wanapirak, C.; Muninthorn, W.; Sanguansermsri, T.; Dhananjayanonda, P.; Tongsong, T. Prevalence of thalassemia in pregnant women at Maharaj Nakorn Chiang Mai Hospital. J. Med. Assoc. Thail. 2004, 87, 1415–1418. [Google Scholar]
- Chui, D.H. Alpha-thalassemia: Hb H disease and Hb Barts hydrops fetalis. Ann. N. Y. Acad. Sci. 2005, 1054, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Modell, B.; Darlison, M. Global epidemiology of haemoglobin disorders and derived service indicators. Bull. World Health Organ. 2008, 86, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Jatavan, P.; Chattipakorn, N.; Tongsong, T. Fetal hemoglobin Bart’s hydrops fetalis: Pathophysiology, prenatal diagnosis and possibility of intrauterine treatment. J. Matern. Fetal. Neonatal Med. 2018, 31, 946–957. [Google Scholar] [CrossRef]
- Yang, J.; Peng, C.F.; Qi, Y.; Rao, X.Q.; Guo, F.; Hou, Y.; He, W.; Wu, J.; Chen, Y.Y.; Zhao, X.; et al. Noninvasive prenatal detection of hemoglobin Bart hydrops fetalis via maternal plasma dispensed with parental haplotyping using the semiconductor sequencing platform. Am. J. Obstet. Gynecol. 2020, 222, 185.e1–185.e17. [Google Scholar] [CrossRef]
- Xiong, F.; Sun, M.; Zhang, X.; Cai, R.; Zhou, Y.; Lou, J.; Zeng, L.; Sun, Q.; Xiao, Q.; Shang, X.; et al. Molecular epidemiological survey of haemoglobinopathies in the Guangxi Zhuang Autonomous Region of southern China. Clin. Genet. 2010, 78, 139–148. [Google Scholar] [CrossRef]
- Lie-Injo, L.E. Alpha-chain thalassemia and hydrops fetalis in Malaya: Report of five cases. Blood 1962, 20, 581–590. [Google Scholar]
- King, A.J.; Higgs, D.R. Potential new approaches to the management of the Hb Bart’s hydrops fetalis syndrome: The most severe form of α-thalassemia. Hematol. Am. Soc. Hematol. Educ. Program 2018, 2018, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Chik, K.W.; Shing, M.M.; Li, C.K.; Leung, T.F.; Tsang, K.S.; Yuen, H.L.; Cheng, S.B.; Yuen, P.M. Treatment of hemoglobin Bart’s hydrops with bone marrow transplantation. J. Pediatr. 1998, 132, 1039–1042. [Google Scholar] [CrossRef]
- Jiang, F.; Li, D.Z. Outcome of survivors with hemoglobin Bart’s hydrops fetalis syndrome: The most severe form of α-thalassemia. Pediatr. Transplant. 2021, 25, e14090. [Google Scholar] [CrossRef]
- Chan, W.Y.K.; Lee, P.P.W.; Lee, V.; Chan, G.C.F.; Leung, W.; Ha, S.Y.; Cheuk, D.K.L. Outcomes of allogeneic transplantation for hemoglobin Bart’s hydrops fetalis syndrome in Hong Kong. Pediatr. Transplant. 2021, 25, e14037. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.T.; Wong, V.C.; So, W.W.; Ma, H.K.; Chan, V.; Todd, D. Homozygous alpha-thalassaemia: Clinical presentation, diagnosis and management. A review of 46 cases. Br. J. Obstet. Gynaecol. 1985, 92, 680–684. [Google Scholar] [CrossRef]
- Thammavong, K.; Luewan, S.; Jatavan, P.; Tongsong, T. Foetal haemodynamic response to anaemia. ESC Heart Fail. 2020, 7, 3473–3482. [Google Scholar] [CrossRef]
- Tongsong, T.; Wanapirak, C.; Sirivatanapa, P.; Sanguansermsri, T.; Sirichotiyakul, S.; Piyamongkol, W.; Chanprapaph, P. Prenatal control of severe thalassaemia: Chiang Mai strategy. Prenat. Diagn. 2000, 20, 229–234. [Google Scholar] [CrossRef]
- Tongsong, T.; Wanapirak, C.; Sirivatanapa, P.; Sa-nguansermsri, T.; Sirichotiyakul, S.; Piyamongkol, W.; Chanprapaph, P.; Steger, H.F.; Sekararithi, R.; Tuggapichitti, A. Prenatal eradication of Hb Bart’s hydrops fetalis. J. Reprod. Med. 2001, 46, 18–22. [Google Scholar]
- Tongsong, T.; Boonyanurak, P. Placental thickness in the first half of pregnancy. J. Clin. Ultrasound JCU 2004, 32, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Tongsong, T.; Tatiyapornkul, T. Cardiothoracic ratio in the first half of pregnancy. J. Clin. Ultrasound 2004, 32, 186–189. [Google Scholar] [CrossRef]
- Tongsong, T.; Wanapirak, C.; Sirichotiyakul, S.; Chanprapaph, P. Sonographic markers of hemoglobin Bart disease at midpregnancy. J. Ultrasound Med. 2004, 23, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Thammavong, K.; Luewan, S.; Tongsong, T. Performance of Fetal Cardiac Volume Derived from VOCAL (Virtual Organ Computer-Aided AnaLysis) in Predicting Hemoglobin (Hb) Bart’s Disease. J. Clin. Med. 2021, 10, 4651. [Google Scholar] [CrossRef]
- Thammavong, K.; Luewan, S.; Wanapirak, C.; Tongsong, T. Ultrasound Features of Fetal Anemia Lessons From Hemoglobin Bart Disease. J. Ultrasound Med. 2021, 40, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Tongprasert, F.; Wanapirak, C.; Tongsong, T. Maternal serum hCG, PAPP-A and AFP as predictors of hemoglobin Bart disease at mid-pregnancy. Prenat. Diagn. 2011, 31, 430–433. [Google Scholar] [CrossRef]
- Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Second trimester maternal serum markers and a predictive model for predicting fetal hemoglobin Bart’s disease. J. Matern. Fetal. Neonatal Med. 2013, 26, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Comparison of maternal serum PlGF and sFlt-1 between pregnancies with and without fetal hemoglobin Bart’s disease. Prenat. Diagn. 2013, 33, 1272–1275. [Google Scholar] [CrossRef]
- Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Second trimester maternal serum inhibin-A in fetal anemia secondary to hemoglobin Bart’s disease. J. Matern. Fetal. Neonatal Med. 2014, 27, 1005–1009. [Google Scholar] [CrossRef]
- Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Comparison of cardiac troponin T and N-terminal pro-B-type natriuretic peptide between fetuses with hemoglobin Bart’s disease and nonanemic fetuses. Prenat. Diagn. 2014, 34, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Wanapirak, C.; Piyamomgkol, W.; Sirichotiyakul, S.; Tongprasert, F.; Srisupundit, K.; Luewan, S.; Traisrisilp, K.; Jatavan, P.; Tongsong, T. Second-trimester maternal serum screening for fetal Down syndrome: As a screening test for hemoglobin Bart’s disease: A prospective population-based study. Prenat. Diagn. 2018, 38, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Mitchell, M.D.; Peiris, H.N.; Kobayashi, M.; Koh, Y.Q.; Duncombe, G.; Illanes, S.E.; Rice, G.E.; Salomon, C. Placental exosomes in normal and complicated pregnancy. Am. J. Obstet. Gynecol. 2015, 213, S173–S181. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, M.Z.; Ratajczak, J. Extracellular microvesicles/exosomes: Discovery, disbelief, acceptance, and the future? Leukemia 2020, 34, 3126–3135. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal. CCS 2021, 19, 47. [Google Scholar] [CrossRef]
- Sarker, S.; Scholz-Romero, K.; Perez, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E.; Salomon, C. Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy. J. Transl. Med. 2014, 12, 204. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Guanzon, D.; Scholz-Romero, K.; Longo, S.; Correa, P.; Illanes, S.E.; Rice, G.E. Placental Exosomes as Early Biomarker of Preeclampsia: Potential Role of Exosomal MicroRNAs Across Gestation. J. Clin. Endocrinol. Metab. 2017, 102, 3182–3194. [Google Scholar] [CrossRef]
- Nair, S.; Salomon, C. Extracellular vesicles as critical mediators of maternal-fetal communication during pregnancy and their potential role in maternal metabolism. Placenta 2020, 98, 60–68. [Google Scholar] [CrossRef]
- Sadovsky, Y.; Ouyang, Y.; Powell, J.S.; Li, H.; Mouillet, J.F.; Morelli, A.E.; Sorkin, A.; Margolis, L. Placental small extracellular vesicles: Current questions and investigative opportunities. Placenta 2020, 102, 34–38. [Google Scholar] [CrossRef]
- Palma, C.; Jellins, J.; Lai, A.; Salas, A.; Campos, A.; Sharma, S.; Duncombe, G.; Hyett, J.; Salomon, C. Extracellular Vesicles and Preeclampsia: Current Knowledge and Future Research Directions. Subcell Biochem. 2021, 97, 455–482. [Google Scholar] [CrossRef] [PubMed]
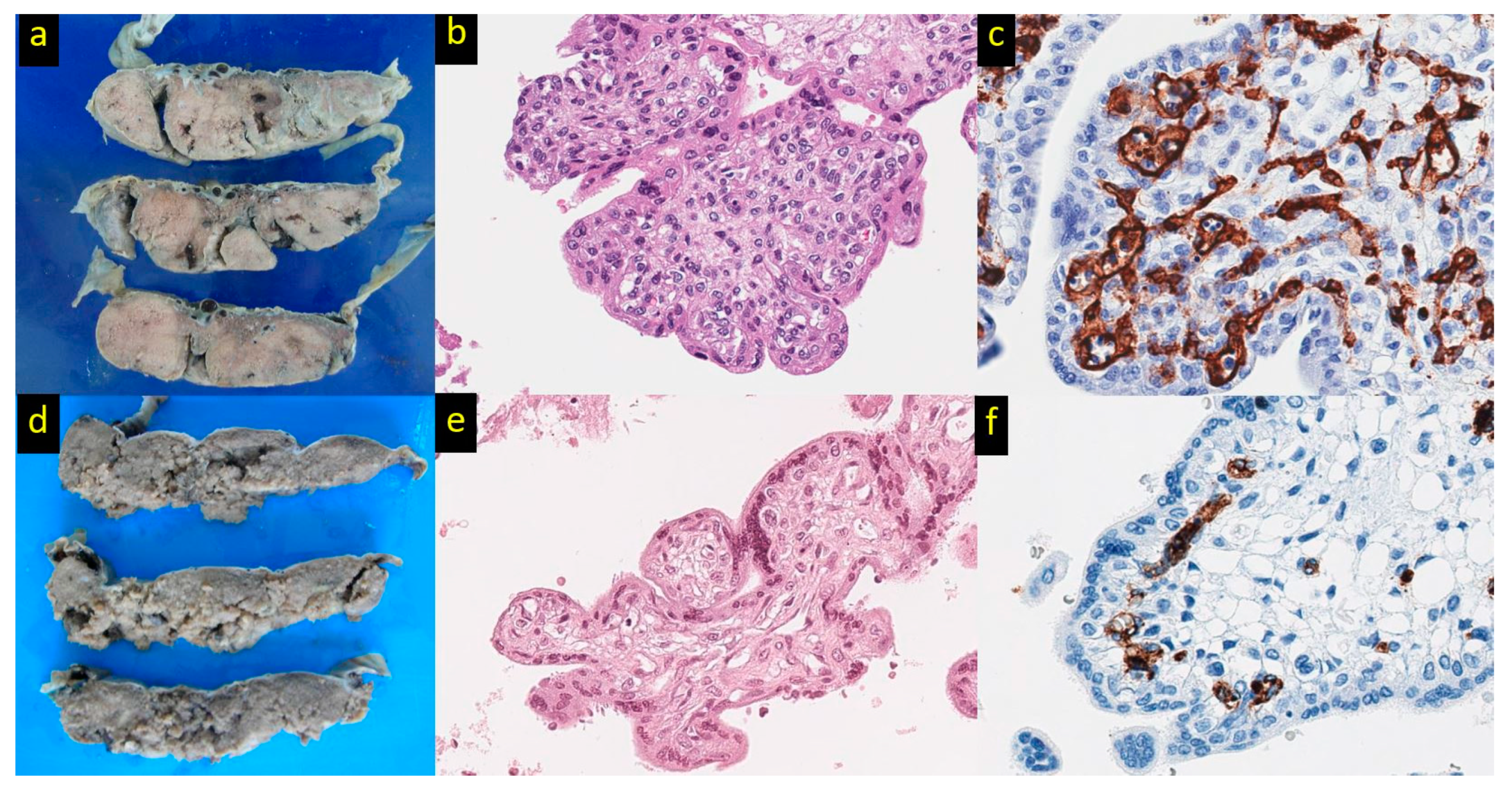
- Taweevisit, M.; Thorner, P.S. Hemoglobin Bart hydrops fetalis: A model for studying vascular changes in placental hypoxia. Placenta 2016, 44, 98–103. [Google Scholar] [CrossRef]
- Taweevisit, M.; Thorner, P.S. Hydrops fetalis in the stillborn: A series from the central region of Thailand. Pediatr. Dev. Pathol. 2010, 13, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Taweevisit, M.; Thorner, P.S. Peripheral villous stromal hyperplasia: A distinctive placental lesion in hemoglobin bart hydrops fetalis. Pediatr. Dev. Pathol. 2012, 15, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Stanek, J. Utility of diagnosing various histological patterns of diffuse chronic hypoxic placental injury. Pediatr. Dev. Pathol. 2012, 15, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Stanek, J. Hypoxic patterns of placental injury: A review. Arch. Pathol. Lab. Med. 2013, 137, 706–720. [Google Scholar] [CrossRef] [PubMed]
- Stanek, J. Placental hypoxic overlap lesions: A clinicoplacental correlation. J. Obstet. Gynaecol. Res. 2015, 41, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Turowski, G.; Vogel, M. Re-view and view on maturation disorders in the placenta. Apmis 2018, 126, 602–612. [Google Scholar] [CrossRef]
- Songdej, D.; Babbs, C.; Higgs, D.R. An international registry of survivors with Hb Bart’s hydrops fetalis syndrome. Blood 2017, 129, 1251–1259. [Google Scholar] [CrossRef] [Green Version]
- Lam, Y.H.; Ghosh, A.; Tang, M.H.; Lee, C.P.; Sin, S.Y. Early ultrasound prediction of pregnancies affected by homozygous alpha-thalassaemia-1. Prenat. Diagn 1997, 17, 327–332. [Google Scholar] [CrossRef]
- Lam, Y.H.; Tang, M.H.; Lee, C.P.; Tse, H.Y. Prenatal ultrasonographic prediction of homozygous type 1 alpha-thalassemia at 12 to 13 weeks of gestation. Am. J. Obstet. Gynecol 1999, 180, 148–150. [Google Scholar] [CrossRef]
- Leung, K.Y.; Cheong, K.B.; Lee, C.P.; Chan, V.; Lam, Y.H.; Tang, M. Ultrasonographic prediction of homozygous alpha0-thalassemia using placental thickness, fetal cardiothoracic ratio and middle cerebral artery Doppler: Alone or in combination? Ultrasound Obstet. Gynecol. 2010, 35, 149–154. [Google Scholar] [CrossRef]
- Sirichotiyakul, S.; Luewan, S.; Srisupundit, K.; Tongprasert, F.; Tongsong, T. Prenatal ultrasound evaluation of fetal Hb Bart’s disease among pregnancies at risk at 11 to 14 weeks of gestation. Prenat. Diagn. 2014, 34, 230–234. [Google Scholar] [CrossRef]
- Leung, K.Y.; Liao, C.; Li, Q.M.; Ma, S.Y.; Tang, M.H.; Lee, C.P.; Chan, V.; Lam, Y.H. A new strategy for prenatal diagnosis of homozygous alpha(0)-thalassemia. Ultrasound Obstet. Gynecol. 2006, 28, 173–177. [Google Scholar] [CrossRef]
- Leung, W.C.; Oepkes, D.; Seaward, G.; Ryan, G. Serial sonographic findings of four fetuses with homozygous alpha-thalassemia-1 from 21 weeks onwards. Ultrasound Obstet. Gynecol. 2002, 19, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Raungrongmorakot, K.; Chaemsaithong, P. Middle cerebral artery peak systolic velocity in fetuses with homozygous alpha-thalassemia-1: Case series. J. Med. Assoc. Thai. 2010, 93, S114–S117. [Google Scholar] [PubMed]
- Srisupundit, K.; Piyamongkol, W.; Tongsong, T. Identification of fetuses with hemoglobin Bart’s disease using middle cerebral artery peak systolic velocity. Ultrasound Obstet. Gynecol. 2009, 33, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Luewan, S.; Tongprasert, F.; Piyamongkol, W.; Wanapirak, C.; Tongsong, T. Fetal liver length measurement at mid-pregnancy among fetuses at risk as a predictor of hemoglobin Bart’s disease. J. Perinatol. 2010, 31, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Tongsong, T.; Piyamongkol, W.; Tongprasert, F.; Srisupundit, K.; Luewan, S. Fetal Splenic Artery Peak Velocity (SPA-PSV) at Mid-Pregnancy as a Predictor of Hb Bart’s Disease. Ultraschall Med. 2011, 32, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Srisupundit, K.; Tongprasert, F.; Luewan, S.; Sirichotiyakul, S.; Tongsong, T. Splenic circumference at midpregnancy as a predictor of hemoglobin Bart’s disease among fetuses at risk. Gynecol. Obstet. Investig. 2011, 72, 63–67. [Google Scholar] [CrossRef]
- Siwawong, W.; Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Fetal cardiac circumference derived by spatiotemporal image correlation as a predictor of fetal hemoglobin Bart disease at midpregnancy. J. Ultrasound Med. 2013, 32, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Harn-a-morn, P.; Wanapirak, C.; Sirichotiyakul, S.; Srisupundit, K.; Tongprasert, F.; Luewan, S.; Tongsong, T. Effectiveness of ultrasound algorithm in prenatal diagnosis of Hemoglobin Bart’s disease among pregnancies at risk. Int. J. Gynecol. Obstet. 2022; in press. [Google Scholar] [CrossRef]
- Li, X.; Qiu, X.; Huang, H.; Zhao, Y.; Li, X.; Li, M.; Tian, X. Fetal heart size measurements as new predictors of homozygous alpha-thalassemia-1 in mid-pregnancy. Congenit. Heart Dis. 2018, 13, 282–287. [Google Scholar] [CrossRef]
- Traisrisilp, K.; Sirilert, S.; Tongsong, T. The performance of cardio-biparietal ratio measured by 2D ultrasound in predicting fetal hemoglobin Bart disease during midpregnancy: A pilot study. Prenat. Diagn. 2019, 39, 647–651. [Google Scholar] [CrossRef]
- Thathan, N.; Traisrisilp, K.; Luewan, S.; Srisupundit, K.; Tongprasert, F.; Tongsong, T. Screening for hemoglobin Bart’s disease among fetuses at risk at mid-pregnancy using the fetal cardiac diameter to biparietal diameter ratio. BMC Pregnancy Childbirth 2014, 14, 230. [Google Scholar] [CrossRef] [PubMed]
- Tongsong, T.; Tongprasert, F.; Srisupundit, K.; Luewan, S.; Traisrisilp, K.; Jatavan, P. Fetal Cardiac Remodeling in Response to Anemia: Using Hemoglobin Bart’s Disease as a Study Model. Ultraschall Med. 2018, 41, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Srisupundit, K.; Piyamongkol, W.; Tongprasert, F.; Luewan, S.; Tongsong, T. Reference range of fetal splenic circumference from 14 to 40 weeks of gestation. Arch. Gynecol. Obstet. 2010, 283, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Tongprasert, F.; Srisupundit, K.; Luewan, S.; Tongsong, T. Second trimester maternal serum alpha-fetoprotein (MSAFP) as predictor of fetal hemoglobin Bart’s disease. Prenat. Diagn. 2014, 34, 1277–1282. [Google Scholar] [CrossRef]
- Tungwiwat, W.; Fucharoen, S.; Fucharoen, G.; Ratanasiri, T.; Sanchaisuriya, K. Development and application of a real-time quantitative PCR for prenatal detection of fetal alpha(0)-thalassemia from maternal plasma. Ann. N. Y. Acad. Sci. 2006, 1075, 103–107. [Google Scholar] [CrossRef]
- Long, X.J.; Long, G.F.; Lin, W.X. [Noninvasive prenatal diagnosis of Hb Bart’s hydrops fetus using cell-free fetal DNA in maternal plasma]. Zhonghua Xue Ye Xue Za Zhi 2009, 30, 175–178. [Google Scholar]
- Pornprasert, S.; Sukunthamala, K.; Kunyanone, N.; Sittiprasert, S.; Thungkham, K.; Junorse, S.; Pongsawatkul, K.; Pattanaporn, W.; Jitwong, C.; Sanguansermsri, T. Analysis of real-time PCR cycle threshold of alpha-thalassemia-1 Southeast Asian type deletion using fetal cell-free DNA in maternal plasma for noninvasive prenatal diagnosis of Bart’s hydrops fetalis. J. Med. Assoc. Thail. 2010, 93, 1243–1248. [Google Scholar]
- Ho, S.S.; Chong, S.S.; Koay, E.S.; Ponnusamy, S.; Chiu, L.; Chan, Y.H.; Rauff, M.; Baig, S.; Chan, J.; Su, L.L.; et al. Noninvasive prenatal exclusion of haemoglobin Bart’s using foetal DNA from maternal plasma. Prenat. Diagn. 2010, 30, 65–73. [Google Scholar] [CrossRef]
- Sirichotiyakul, S.; Charoenkwan, P.; Sanguansermsri, T. Prenatal diagnosis of homozygous alpha-thalassemia-1 by cell-free fetal DNA in maternal plasma. Prenat. Diagn. 2012, 32, 45–49. [Google Scholar] [CrossRef]
- Li, X.; Yang, T.; Li, C.S.; Jin, L.; Lou, H.; Song, Y. Prenatal detection of thalassemia by cell-free fetal DNA (cffDNA) in maternal plasma using surface enhanced Raman spectroscopy combined with PCR. Biomed. Opt. Express 2018, 9, 3167–3176. [Google Scholar] [CrossRef]
- Sawakwongpra, K.; Tangmansakulchai, K.; Ngonsawan, W.; Promwan, S.; Chanchamroen, S.; Quangkananurug, W.; Sriswasdi, S.; Jantarasaengaram, S.; Ponnikorn, S. Droplet-based digital PCR for non-invasive prenatal genetic diagnosis of α and β-thalassemia. Biomed. Rep. 2021, 15, 82. [Google Scholar] [CrossRef]
- Salomon, C.; Yee, S.; Scholz-Romero, K.; Kobayashi, M.; Vaswani, K.; Kvaskoff, D.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. Extravillous trophoblast cells-derived exosomes promote vascular smooth muscle cell migration. Front. Pharmacol. 2014, 5, 175. [Google Scholar] [CrossRef]
- Dragovic, R.A.; Southcombe, J.H.; Tannetta, D.S.; Redman, C.W.; Sargent, I.L. Multicolor flow cytometry and nanoparticle tracking analysis of extracellular vesicles in the plasma of normal pregnant and pre-eclamptic women. Biol. Reprod. 2013, 89, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Niida, S.; Azuma, E.; Yanagibashi, T.; Muramatsu, M.; Huang, T.T.; Sagara, H.; Higaki, S.; Ikutani, M.; Nagai, Y.; et al. Inflammation-induced endothelial cell-derived extracellular vesicles modulate the cellular status of pericytes. Sci. Rep. 2015, 5, 8505. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Y.; Mouillet, J.F.; Coyne, C.B.; Sadovsky, Y. Review: Placenta-specific microRNAs in exosomes—Good things come in nano-packages. Placenta 2014, 35, S69–S73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemmens, H.; Lambert, D.W. Extracellular vesicles: Translational challenges and opportunities. Biochem. Soc. Trans. 2018, 46, 1073–1082. [Google Scholar] [CrossRef]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating Extracellular Vesicles in Human Disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- De Wever, O.; Hendrix, A. A supporting ecosystem to mature extracellular vesicles into clinical application. Embo J. 2019, 38, e101412. [Google Scholar] [CrossRef]
- He, J.; Ren, W.; Wang, W.; Han, W.; Jiang, L.; Zhang, D.; Guo, M. Exosomal targeting and its potential clinical application. Drug Deliv. Transl. Res. 2021, 12, 2385–2402. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Mouillet, J.F.; Sorkin, A.; Sadovsky, Y. Trophoblastic extracellular vesicles and viruses: Friends or foes? Am. J. Reprod. Immunol. 2021, 85, e13345. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C.R. The Variation of Animals and Plants under Domestication; J. Murray: London, UK, 1868; Volume 1. [Google Scholar]
- Liu, Y.; Chen, Q. 150 years of Darwin’s theory of intercellular flow of hereditary information. Nat. Rev. Mol. Cell Biol. 2018, 19, 749–750. [Google Scholar] [CrossRef] [PubMed]
- Schmorl, G. Pathologisch-Anatomische Unteruchungen Uber Puerperal-Eklampsie; Verlag FCW: Leipzig, Germany, 1893. [Google Scholar]
- Lapaire, O.; Holzgreve, W.; Oosterwijk, J.C.; Brinkhaus, R.; Bianchi, D.W. Georg Schmorl on trophoblasts in the maternal circulation. Placenta 2007, 28, 1–5. [Google Scholar] [CrossRef]
- Li, H.; Pinilla-Macua, I.; Ouyang, Y.; Sadovsky, E.; Kajiwara, K.; Sorkin, A.; Sadovsky, Y. Internalization of trophoblastic small extracellular vesicles and detection of their miRNA cargo in P-bodies. J. Extracell. Vesicles 2020, 9, 1812261. [Google Scholar] [CrossRef] [PubMed]
- Sheller-Miller, S.; Choi, K.; Choi, C.; Menon, R. Cyclic-recombinase-reporter mouse model to determine exosome communication and function during pregnancy. Am. J. Obstet. Gynecol. 2019, 221, 502.e1–502.e12. [Google Scholar] [CrossRef] [PubMed]
- Sheller-Miller, S.; Lei, J.; Saade, G.; Salomon, C.; Burd, I.; Menon, R. Feto-Maternal Trafficking of Exosomes in Murine Pregnancy Models. Front. Pharmacol. 2016, 7, 432. [Google Scholar] [CrossRef] [Green Version]
- Borges, F.T.; Reis, L.A.; Schor, N. Extracellular vesicles: Structure, function, and potential clinical uses in renal diseases. Braz. J. Med. Biol. Res. 2013, 46, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef]
- Quesenberry, P.J.; Aliotta, J.M. Cellular phenotype.e switching and microvesicles. Adv. Drug Deliv. Rev. 2010, 62, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, C.D.; Lodha, S.; Sahoo, S. EV Cargo Sorting in Therapeutic Development for Cardiovascular Disease. Cells 2021, 10, 1500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Kobayashi, M.; Ashman, K.; Sobrevia, L.; Mitchell, M.D.; Rice, G.E. Hypoxia-induced changes in the bioactivity of cytotrophoblast-derived exosomes. PLoS ONE 2013, 8, e79636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomon, C.; Ryan, J.; Sobrevia, L.; Kobayashi, M.; Ashman, K.; Mitchell, M.; Rice, G.E. Exosomal signaling during hypoxia mediates microvascular endothelial cell migration and vasculogenesis. PLoS ONE 2013, 8, e68451. [Google Scholar] [CrossRef] [PubMed]
- Rice, G.E.; Scholz-Romero, K.; Sweeney, E.; Peiris, H.; Kobayashi, M.; Duncombe, G.; Mitchell, M.D.; Salomon, C. The Effect of Glucose on the Release and Bioactivity of Exosomes From First Trimester Trophoblast Cells. J. Clin. Endocrinol. Metab. 2015, 100, E1280–E1288. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef]
- Welton, J.L.; Webber, J.P.; Botos, L.A.; Jones, M.; Clayton, A. Ready-made chromatography columns for extracellular vesicle isolation from plasma. J. Extracell. Vesicles 2015, 4, 27269. [Google Scholar] [CrossRef] [PubMed]
- Stranska, R.; Gysbrechts, L.; Wouters, J.; Vermeersch, P.; Bloch, K.; Dierickx, D.; Andrei, G.; Snoeck, R. Comparison of membrane affinity-based method with size-exclusion chromatography for isolation of exosome-like vesicles from human plasma. J. Transl. Med. 2018, 16, 1. [Google Scholar] [CrossRef]
- Koh, Y.Q.; Almughlliq, F.B.; Vaswani, K.; Peiris, H.N.; Mitchell, M.D. Exosome enrichment by ultracentrifugation and size exclusion chromatography. Front. Biosci. (Landmark Ed.) 2018, 23, 865–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, A.-K.; De Miroschedji, K.; Doeppner, T.R.; Börger, V.; Ruesing, J.; Rebmann, V.; Durst, S.; Jansen, S.; Bremer, M.; Behrmann, E.; et al. Precipitation with polyethylene glycol followed by washing and pelleting by ultracentrifugation enriches extracellular vesicles from tissue culture supernatants in small and large scales. J. Extracell. Vesicles 2018, 7, 1528109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.K.; Whiteside, T.L. Immunoaffinity-Based Isolation of Melanoma Cell-Derived and T Cell-Derived Exosomes from Plasma of Melanoma Patients. Methods Mol. Biol. 2021, 2265, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Yousif, G.; Qadri, S.; Parray, A.; Akhthar, N.; Shuaib, A.; Haik, Y. Exosomes Derived Neuronal Markers: Immunoaffinity Isolation and Characterization. NeuroMol. Med. 2021, 24, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Chutipongtanate, S.; Kongsomros, S.; Pongsakul, N.; Panachan, J.; Khowawisetsut, L.; Pattanapanyasat, K.; Hongeng, S.; Thitithanyanont, A. Anti-SARS-CoV-2 effect of extracellular vesicles released from mesenchymal stem cells. J. Extracell. Vesicles 2022, 11, e12201. [Google Scholar] [CrossRef] [PubMed]
- Panachan, J.; Rojsirikulchai, N.; Pongsakul, N.; Khowawisetsut, L.; Pongphitcha, P.; Siriboonpiputtana, T.; Chareonsirisuthigul, T.; Phornsarayuth, P.; Klinkulab, N.; Jinawath, N.; et al. Extracellular Vesicle-Based Method for Detecting MYCN Amplification Status of Pediatric Neuroblastoma. Cancers 2022, 14, 2627. [Google Scholar] [CrossRef]
- Sharma, S.; Rasool, H.I.; Palanisamy, V.; Mathisen, C.; Schmidt, M.; Wong, D.T.; Gimzewski, J.K. Structural-mechanical characterization of nanoparticle exosomes in human saliva, using correlative AFM, FESEM, and force spectroscopy. ACS Nano 2010, 4, 1921–1926. [Google Scholar] [CrossRef] [Green Version]
- Dragovic, R.A.; Gardiner, C.; Brooks, A.S.; Tannetta, D.S.; Ferguson, D.J.; Hole, P.; Carr, B.; Redman, C.W.; Harris, A.L.; Dobson, P.J.; et al. Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomedicine 2011, 7, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Groot Kormelink, T.; Arkesteijn, G.J.; Nauwelaers, F.A.; van den Engh, G.; Nolte-’t Hoen, E.N.; Wauben, M.H. Prerequisites for the analysis and sorting of extracellular vesicle subpopulations by high-resolution flow cytometry. Cytometry A 2016, 89, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Chutipongtanate, S.; Greis, K.D. Multiplex Biomarker Screening Assay for Urinary Extracellular Vesicles Study: A Targeted Label-Free Proteomic Approach. Sci. Rep. 2018, 8, 15039. [Google Scholar] [CrossRef] [Green Version]
- Menon, R.; Dixon, C.L.; Sheller-Miller, S.; Fortunato, S.J.; Saade, G.R.; Palma, C.; Lai, A.; Guanzon, D.; Salomon, C. Quantitative Proteomics by SWATH-MS of Maternal Plasma Exosomes Determine Pathways Associated With Term and Preterm Birth. Endocrinology 2019, 160, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, Y.M.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]
- Bianchi, D.W.; Lo, Y.M. Fetomaternal cellular and plasma DNA trafficking: The Yin and the Yang. Ann. N. Y. Acad. Sci. 2001, 945, 119–131. [Google Scholar] [CrossRef]
- Chiu, R.W.; Lo, Y.M. Application of fetal DNA in maternal plasma for noninvasive prenatal diagnosis. Expert Rev. Mol. Diagn. 2002, 2, 32–40. [Google Scholar] [CrossRef]
- Chiu, R.W.; Lo, Y.M. Non-invasive prenatal diagnosis: On the horizon? Pharmacogenomics 2003, 4, 191–200. [Google Scholar] [CrossRef]
- Bianchi, D.W. Circulating fetal DNA: Its origin and diagnostic potential-a review. Placenta 2004, 25 (Suppl. A), S93–S101. [Google Scholar] [CrossRef]
- Wataganara, T.; Bianchi, D.W. Fetal cell-free nucleic acids in the maternal circulation: New clinical applications. Ann. N. Y. Acad. Sci. 2004, 1022, 90–99. [Google Scholar] [CrossRef]
- Lo, Y.M.; Chiu, R.W. Prenatal diagnosis: Progress through plasma nucleic acids. Nat. Rev. Genet. 2007, 8, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Bianchi, D.W. Fetal fraction and noninvasive prenatal testing: What clinicians need to know. Prenat. Diagn. 2020, 40, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Chiu, R.W.K.; Lo, Y.M.D. Cell-free fetal DNA coming in all sizes and shapes. Prenat. Diagn. 2021, 41, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.M.D. Noninvasive prenatal testing: Advancing through a virtuous circle of science, technology and clinical applications. Prenat. Diagn. 2021, 41, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Placental debris, oxidative stress and pre-eclampsia. Placenta 2000, 21, 597–602. [Google Scholar] [CrossRef]
- Gercel-Taylor, C.; O’Connor, S.M.; Lam, G.K.; Taylor, D.D. Shed membrane fragment modulation of CD3-zeta during pregnancy: Link with induction of apoptosis. J. Reprod. Immunol. 2002, 56, 29–44. [Google Scholar] [CrossRef]
- Kam, W.; Clauser, E.; Kim, Y.S.; Kan, Y.W.; Rutter, W.J. Cloning, sequencing, and chromosomal localization of human term placental alkaline phosphatase cDNA. Proc. Natl. Acad. Sci. USA 1985, 82, 8715–8719. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, M.; Stenqvist, A.C.; Nagaeva, O.; Kjellberg, L.; Wulff, M.; Baranov, V.; Mincheva-Nilsson, L. Human placenta expresses and secretes NKG2D ligands via exosomes that down-modulate the cognate receptor expression: Evidence for immunosuppressive function. J. Immunol. 2009, 183, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Mincheva-Nilsson, L.; Baranov, V. Placenta-derived exosomes and syncytiotrophoblast microparticles and their role in human reproduction: Immune modulation for pregnancy success. Am. J. Reprod. Immunol. 2014, 72, 440–457. [Google Scholar] [CrossRef]
- Salomon, C.; Torres, M.J.; Kobayashi, M.; Scholz-Romero, K.; Sobrevia, L.; Dobierzewska, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. A gestational profile of placental exosomes in maternal plasma and their effects on endothelial cell migration. PLoS ONE 2014, 9, e98667. [Google Scholar] [CrossRef] [Green Version]
- Henthorn, P.S.; Raducha, M.; Kadesch, T.; Weiss, M.J.; Harris, H. Sequence and characterization of the human intestinal alkaline phosphatase gene. J. Biol. Chem. 1988, 263, 12011–12019. [Google Scholar] [CrossRef]
- Kniss, D.A.; Xie, Y.; Li, Y.; Kumar, S.; Linton, E.A.; Cohen, P.; Fan-Havard, P.; Redman, C.W.; Sargent, I.L. ED(27) trophoblast-like cells isolated from first-trimester chorionic villi are genetically identical to HeLa cells yet exhibit a distinct phenotype. Placenta 2002, 23, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Salomon, C.; Yee, S.W.; Mitchell, M.D.; Rice, G.E. The possible role of extravillous trophoblast-derived exosomes on the uterine spiral arterial remodeling under both normal and pathological conditions. Biomed. Res. Int. 2014, 2014, 693157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.C.; Liu, X.B.; Huang, S.; Bi, X.Y.; Wang, H.X.; Xie, L.X.; Wang, Y.Q.; Cao, X.F.; Lv, J.; Xiao, F.J.; et al. Microvesicles derived from human umbilical cord mesenchymal stem cells stimulated by hypoxia promote angiogenesis both in vitro and in vivo. Stem Cells Dev. 2012, 21, 3289–3297. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Nagaeva, O.; Chen, T.; Stendahl, U.; Antsiferova, J.; Mogren, I.; Hernestål, J.; Baranov, V. Placenta-derived soluble MHC class I chain-related molecules down-regulate NKG2D receptor on peripheral blood mononuclear cells during human pregnancy: A possible novel immune escape mechanism for fetal survival. J. Immunol. 2006, 176, 3585–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Salomon, C. Extracellular vesicles and their immunomodulatory functions in pregnancy. Semin. Immunopathol. 2018, 40, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Sabapatha, A.; Gercel-Taylor, C.; Taylor, D.D. Specific isolation of placenta-derived exosomes from the circulation of pregnant women and their immunoregulatory consequences. Am. J. Reprod. Immunol. 2006, 56, 345–355. [Google Scholar] [CrossRef]
- Chang, G.; Mouillet, J.F.; Mishima, T.; Chu, T.; Sadovsky, E.; Coyne, C.B.; Parks, W.T.; Surti, U.; Sadovsky, Y. Expression and trafficking of placental microRNAs at the feto-maternal interface. FASEB J. 2017, 31, 2760–2770. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human villous trophoblasts express and secrete placenta-specific microRNAs into maternal circulation via exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Donker, R.B.; Mouillet, J.F.; Chu, T.; Hubel, C.A.; Stolz, D.B.; Morelli, A.E.; Sadovsky, Y. The expression profile of C19MC microRNAs in primary human trophoblast cells and exosomes. Mol. Hum. Reprod. 2012, 18, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Atay, S.; Gercel-Taylor, C.; Suttles, J.; Mor, G.; Taylor, D.D. Trophoblast-derived exosomes mediate monocyte recruitment and differentiation. Am. J. Reprod. Immunol. 2011, 65, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Kambe, S.; Yoshitake, H.; Yuge, K.; Ishida, Y.; Ali, M.M.; Takizawa, T.; Kuwata, T.; Ohkuchi, A.; Matsubara, S.; Suzuki, M.; et al. Human exosomal placenta-associated miR-517a-3p modulates the expression of PRKG1 mRNA in Jurkat cells. Biol. Reprod. 2014, 91, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collett, G.P.; Redman, C.W.; Sargent, I.L.; Vatish, M. Endoplasmic reticulum stress stimulates the release of extracellular vesicles carrying danger-associated molecular pattern (DAMP) molecules. Oncotarget 2018, 9, 6707–6717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czernek, L.; Düchler, M. Exosomes as Messengers Between Mother and Fetus in Pregnancy. Int. J. Mol. Sci. 2020, 21, 4264. [Google Scholar] [CrossRef]
- Atay, S.; Gercel-Taylor, C.; Taylor, D.D. Human trophoblast-derived exosomal fibronectin induces pro-inflammatory IL-1β production by macrophages. Am. J. Reprod. Immunol. 2011, 66, 259–269. [Google Scholar] [CrossRef]
- Hamilton, S.T.; Hahn, F.; Sonntag, E.; Marschall, M.; Rawlinson, W.D. A placental specific miRNA miR-517a-3p exerts anti-human cytomegalovirus activity. Placenta 2021, 112, 62–65. [Google Scholar] [CrossRef]
- Patil, R.; Ghosh, K.; Shetty, S. Comment on Salomon et al. Gestational Diabetes Mellitus Is Associated With Changes in the Concentration and Bioactivity of Placenta-Derived Exosomes in Maternal Circulation Across Gestation. Diabetes 2016, 65, e24–e25. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Scholz-Romero, K.; Sarker, S.; Sweeney, E.; Kobayashi, M.; Correa, P.; Longo, S.; Duncombe, G.; Mitchell, M.D.; Rice, G.E.; et al. Gestational Diabetes Mellitus Is Associated With Changes in the Concentration and Bioactivity of Placenta-Derived Exosomes in Maternal Circulation Across Gestation. Diabetes 2016, 65, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Rice, G.E. Reply. Am. J. Obstet. Gynecol. 2016, 214, 766–767. [Google Scholar] [CrossRef] [Green Version]
- Gillet, V.; Ouellet, A.; Stepanov, Y.; Rodosthenous, R.S.; Croft, E.K.; Brennan, K.; Abdelouahab, N.; Baccarelli, A.; Takser, L. miRNA Profiles in Extracellular Vesicles From Serum Early in Pregnancies Complicated by Gestational Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2019, 104, 5157–5169. [Google Scholar] [CrossRef]
- Record, M. Intercellular communication by exosomes in placenta: A possible role in cell fusion? Placenta 2014, 35, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Adam, S.; Elfeky, O.; Kinhal, V.; Dutta, S.; Lai, A.; Jayabalan, N.; Nuzhat, Z.; Palma, C.; Rice, G.E.; Salomon, C. Review: Fetal-maternal communication via extracellular vesicles—Implications for complications of pregnancies. Placenta 2017, 54, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, S.K.; Alam, S.M.; Jasti, S.; Hodes, H.; Nauser, T.; Gilliam, M.; Billstrand, C.; Hunt, J.S.; Petroff, M.G. Immunomodulatory molecules are released from the first trimester and term placenta via exosomes. Placenta 2012, 33, 982–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, E.K.; Tsui, N.B.; Lau, T.K.; Leung, T.N.; Chiu, R.W.; Panesar, N.S.; Lit, L.C.; Chan, K.W.; Lo, Y.M. mRNA of placental origin is readily detectable in maternal plasma. Proc. Natl. Acad. Sci. USA 2003, 100, 4748–4753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauniaux, E.; Jurkovic, D.; Campbell, S. Current topic: In vivo investigation of the placental circulations by Doppler echography. Placenta 1995, 16, 323–331. [Google Scholar] [CrossRef]
- Truong, G.; Guanzon, D.; Kinhal, V.; Elfeky, O.; Lai, A.; Longo, S.; Nuzhat, Z.; Palma, C.; Scholz-Romero, K.; Menon, R.; et al. Oxygen tension regulates the miRNA profile and bioactivity of exosomes released from extravillous trophoblast cells—Liquid biopsies for monitoring complications of pregnancy. PLoS ONE 2017, 12, e0174514. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of exosomes as liquid biopsy in clinical diagnosis. Signal Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Escudero, C.A.; Herlitz, K.; Troncoso, F.; Acurio, J.; Aguayo, C.; Roberts, J.M.; Truong, G.; Duncombe, G.; Rice, G.; Salomon, C. Role of Extracellular Vesicles and microRNAs on Dysfunctional Angiogenesis during Preeclamptic Pregnancies. Front. Physiol. 2016, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Jayabalan, N.; Nair, S.; Nuzhat, Z.; Rice, G.E.; Zuñiga, F.A.; Sobrevia, L.; Leiva, A.; Sanhueza, C.; Gutiérrez, J.A.; Lappas, M.; et al. Cross Talk between Adipose Tissue and Placenta in Obese and Gestational Diabetes Mellitus Pregnancies via Exosomes. Front. Endocrinol. (Lausanne) 2017, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Guanzon, D.; Jayabalan, N.; Lai, A.; Scholz-Romero, K.; Kalita de Croft, P.; Ormazabal, V.; Palma, C.; Diaz, E.; McCarthy, E.A.; et al. Extracellular vesicle-associated miRNAs are an adaptive response to gestational diabetes mellitus. J. Transl. Med. 2021, 19, 360. [Google Scholar] [CrossRef]
- Nair, S.; Ormazabal, V.; Lappas, M.; McIntyre, H.D.; Salomon, C. Extracellular vesicles and their potential role inducing changes in maternal insulin sensitivity during gestational diabetes mellitus. Am. J. Reprod. Immunol. 2021, 85, e13361. [Google Scholar] [CrossRef]
- Miranda, J.; Paules, C.; Nair, S.; Lai, A.; Palma, C.; Scholz-Romero, K.; Rice, G.E.; Gratacos, E.; Crispi, F.; Salomon, C. Placental exosomes profile in maternal and fetal circulation in intrauterine growth restriction—Liquid biopsies to monitoring fetal growth. Placenta 2018, 64, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Menon, R.; Debnath, C.; Lai, A.; Guanzon, D.; Bhatnagar, S.; Kshetrapal, P.; Sheller-Miller, S.; Salomon, C. Protein Profile Changes in Circulating Placental Extracellular Vesicles in Term and Preterm Births: A Longitudinal Study. Endocrinology 2020, 161, bqaa009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, R.; Shahin, H. Extracellular vesicles in spontaneous preterm birth. Am. J. Reprod. Immunol. 2021, 85, e13353. [Google Scholar] [CrossRef] [PubMed]
- Radnaa, E.; Richardson, L.S.; Sheller-Miller, S.; Baljinnyam, T.; de Castro Silva, M.; Kumar Kammala, A.; Urrabaz-Garza, R.; Kechichian, T.; Kim, S.; Han, A.; et al. Extracellular vesicle mediated feto-maternal HMGB1 signaling induces preterm birth. Lab. Chip 2021, 21, 1956–1973. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Dixon, C.L.; Cayne, S.; Radnaa, E.; Salomon, C.; Sheller-Miller, S. Differences in cord blood extracellular vesicle cargo in preterm and term births. Am. J. Reprod. Immunol. 2022, 87, e13521. [Google Scholar] [CrossRef]
- Holder, B.S.; Tower, C.L.; Jones, C.J.; Aplin, J.D.; Abrahams, V.M. Heightened pro-inflammatory effect of preeclamptic placental microvesicles on peripheral blood immune cells in humans. Biol. Reprod. 2012, 86, 103. [Google Scholar] [CrossRef]
- Tobin, B.W.; Marchello, M.J. Islet transplantation reverses carcass protein loss in diabetic rats without inducing disproportionate fat accumulation. Diabetologia 1995, 38, 881–888. [Google Scholar] [CrossRef]
- Taylor, D.D.; Shah, S. Methods of isolating extracellular vesicles impact down-stream analyses of their cargoes. Methods 2015, 87, 3–10. [Google Scholar] [CrossRef]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Chutipongtanate, S.; Chatchen, S.; Svasti, J. Plasma prefractionation methods for proteomic analysis and perspectives in clinical applications. Proteom. Clin. Appl. 2017, 11, 1600135. [Google Scholar] [CrossRef]
- Kheansaard, W.; Phongpao, K.; Paiboonsukwong, K.; Pattanapanyasat, K.; Chaichompoo, P.; Svasti, S. Microparticles from β-thalassaemia/HbE patients induce endothelial cell dysfunction. Sci. Rep. 2018, 8, 13033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaihmon, P.; Lertthammakiat, S.; Anurathapan, U.; Pakakasama, S.; Sirachainan, N.; Hongeng, S.; Pattanapanyasat, K. Activated platelets and leukocyte activations in young patients with β-thalassemia/HbE following bone marrow transplantation. Thromb. Res. 2018, 169, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Siriworadetkun, S.; Thubthed, R.; Thiengtavor, C.; Paiboonsukwong, K.; Khuhapinant, A.; Fucharoen, S.; Pattanapanyasat, K.; Vadolas, J.; Svasti, S.; Chaichompoo, P. Elevated levels of circulating monocytic myeloid derived suppressor cells in splenectomised β-thalassaemia/HbE patients. Br. J. Haematol. 2020, 191, e72–e76. [Google Scholar] [CrossRef]
- Atipimonpat, A.; Siwaponanan, P.; Khuhapinant, A.; Svasti, S.; Sukapirom, K.; Khowawisetsut, L.; Pattanapanyasat, K. Extracellular vesicles from thalassemia patients carry iron-containing ferritin and hemichrome that promote cardiac cell proliferation. Ann. Hematol. 2021, 100, 1929–1946. [Google Scholar] [CrossRef] [PubMed]
- Klaihmon, P.; Khuhapinant, A.; Kheansaard, W.; Pattanapanyasat, K. Internalization of cell-derived microparticles triggers endothelial pro-inflammatory responses. Asian Pac. J. Allergy Immunol. 2021. [Google Scholar] [CrossRef]
- Levin, C.; Koren, A.; Rebibo-Sabbah, A.; Levin, M.; Koifman, N.; Brenner, B.; Aharon, A. Extracellular Vesicle MicroRNA That Are Involved in β-Thalassemia Complications. Int. J. Mol. Sci. 2021, 22, 9760. [Google Scholar] [CrossRef] [PubMed]
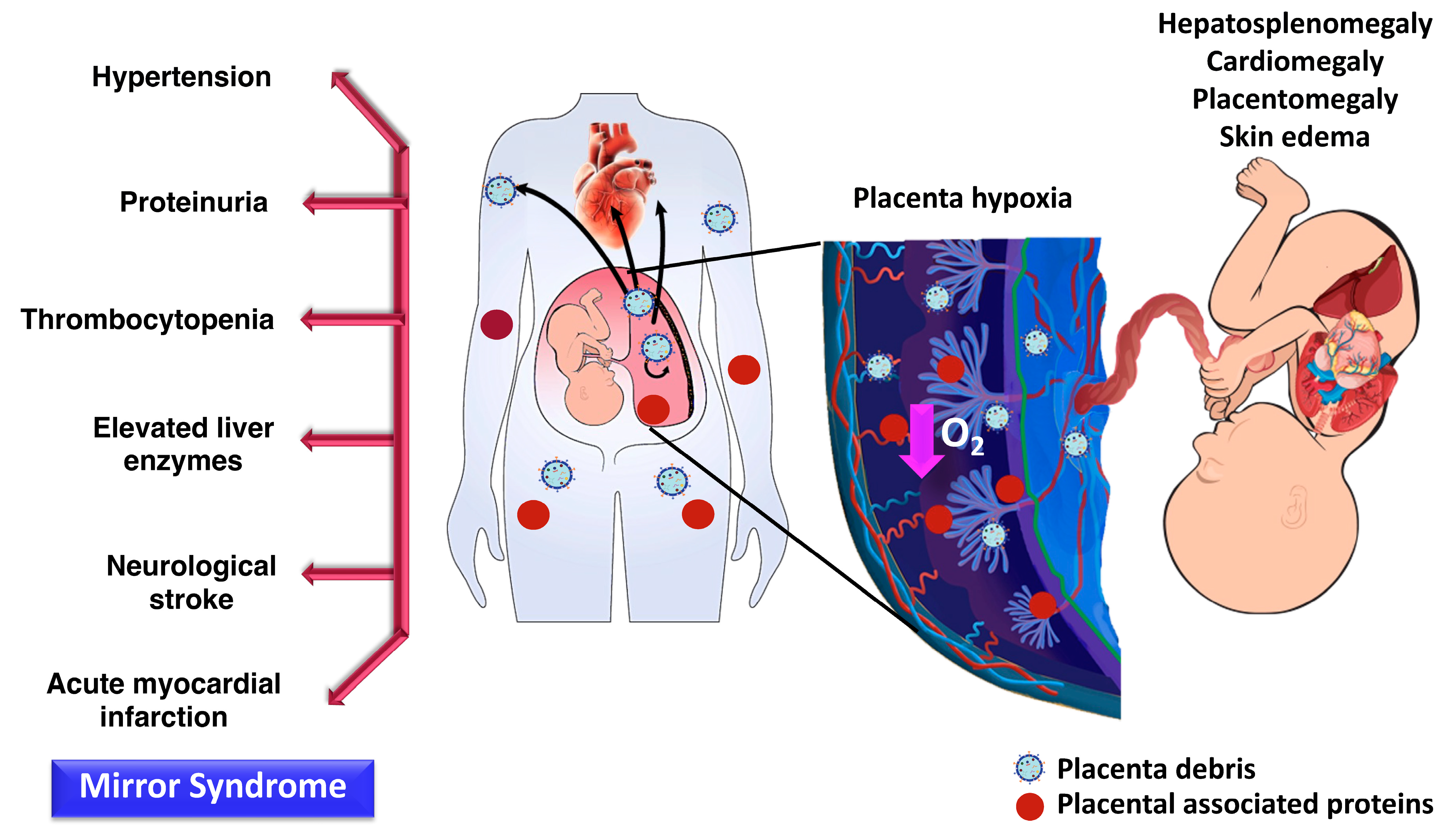
- John, A.H.; Duncan, A.S. The maternal syndrome associated with hydrops foetalis. J. Obstet. Gynaecol. Br. Commonw. 1964, 71, 61–65. [Google Scholar] [CrossRef]
- Kaiser, I.H. Ballantyne and triple edema. Am. J. Obstet. Gynecol. 1971, 110, 115–120. [Google Scholar] [CrossRef]
- Gedikbasi, A.; Oztarhan, K.; Gunenc, Z.; Yildirim, G.; Arslan, O.; Yildirim, D.; Ceylan, Y. Preeclampsia due to fetal non-immune hydrops: Mirror syndrome and review of literature. Hypertens. Pregnancy 2011, 30, 322–330. [Google Scholar] [CrossRef]
- Catalano, M.; O’Driscoll, L. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2019, 9, 1703244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongsomros, S.; Suksatu, A.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Prasongtanakij, S.; Jearawuttanakul, K.; Borwornpinyo, S.; Hongeng, S.; Thitithanyanont, A.; Chutipongtanate, S. Anti-SARS-CoV-2 Activity of Extracellular Vesicle Inhibitors: Screening, Validation, and Combination with Remdesivir. Biomedicines 2021, 9, 1230. [Google Scholar] [CrossRef] [PubMed]
- Chiangjong, W.; Chutipongtanate, S. EV-out or EV-in: Tackling cell-to-cell communication within the tumor microenvironment to enhance anti-tumor efficacy using extracellular vesicle-based therapeutic strategies. OpenNano 2022, 8, 100085. [Google Scholar] [CrossRef]
- Rolnik, D.L.; Nicolaides, K.H.; Poon, L.C. Prevention of preeclampsia with aspirin. Am. J. Obstet. Gynecol. 2022, 226, S1108–S1119. [Google Scholar] [CrossRef]
- Tweet, M.S.; Lewey, J.; Smilowitz, N.R.; Rose, C.H.; Best, P.J.M. Pregnancy-Associated Myocardial Infarction: Prevalence, Causes, and Interventional Management. Circ. Cardiovasc. Interv. 2020, 13, e20008687. [Google Scholar] [CrossRef]
- Verbruggen, M.; Mannaerts, D.; Muys, J.; Jacquemyn, Y. Use of ticagrelor in human pregnancy, the first experience. BMJ Case Rep. 2015, bcr2015212217. [Google Scholar] [CrossRef]
- Nana, M.; Morgan, H.; Moore, S.; Lee, Z.X.; Ang, E.; Nelson-Piercy, C. Antiplatelet therapy in pregnancy: A systematic review. Pharmacol. Res. 2021, 168, 105547. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Goetzl, L.; Karliner, J.S.; Tang, N.; Pulliam, L. Human plasma platelet-derived exosomes: Effects of aspirin. FASEB J. 2016, 30, 2058–2063. [Google Scholar] [CrossRef] [Green Version]
- Casieri, V.; Matteucci, M.; Pasanisi, E.M.; Papa, A.; Barile, L.; Fritsche-Danielson, R.; Lionetti, V. Ticagrelor Enhances Release of Anti-Hypoxic Cardiac Progenitor Cell-Derived Exosomes Through Increasing Cell Proliferation In Vitro. Sci. Rep. 2020, 10, 2494. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Gestational Age (Week) | Cut-Off Value | Sensitivity (%) | Specificity (%) | References | |
|---|---|---|---|---|---|---|
| Potential biomarkers | Cardio-thoracic diameter ratio | 12–15 | ≥0.50 | 75–100 | 90–100 | [18,46,47,48,49,50,59] |
| 18–22 | ≥0.52 | 88–100 | 87–96 | |||
| Middle cerebral artery—peak systolic velocity | 12–15 | ≥1.5 MoM | 17–56 | 79–96 | [48,49,53] | |
| 16–22 | ≥1.5 MoM | 64–85 | 98–100 | |||
| Placental thickness | 12–15 | ≥18 mm | 72.9 | 68.8 | [48,49,52] | |
| 18–22 | ≥30 mm | 74–100 | 96.1 | |||
| Other additional biomarkers | Thickened nuchal translucency | 11–14 | >95th percentile | 16.7 | 98.6 | [49] |
| Cardio-biparietal diameter ratio | 17–22 | ≥0.45 | 84–91 | 77–93 | [60,61] | |
| Cardiac circumference | 15–22 | 1.17 MoM | 86.4 | 78.1 | [57] | |
| Global sphericity index | 18–22 | 1.17 | 74.1 | 88.2 | [62] | |
| Liver length | 18–22 | ≥27.0 mm | 71.3 | 95.5 | [54] | |
| Splenic circumference | 18–22 | ≥1.5 MoM | 70.1 | 83.0 | [63] | |
| Splenic artery—peak systolic velocity | 18–22 | ≥1.51 MoM | 84.4 | 98.1 | [55] |
| Specimens/Procedures | Biomarkers | Cut-Off Value | Test Performance | References |
|---|---|---|---|---|
| Fetal cell-free DNA | Fetal cell-free DNA (GA 11–13 weeks) | - | sens: 98.08% spec: 96.06% | [6] |
| Maternal serum biomarker in second trimester | MAFP | ≥1.5 MoM | sens: 87.2% spec: 74.5% | [23,24,25,27,64] |
| uE3 | <5th percentile | p < 0.001 | ||
| Free β-hCG | Increased | p = 0.543 | ||
| PAPP-A | Increased | p = 0.777 | ||
| Inhibin-A | Increased | p = 0.001 | ||
| PlGF | Increased | p = 0.008 | ||
| sFlt-1 | Increased | p = 0.139 | ||
| sFlt-1/PlGF ratio | Decreased | p = 0.001 | ||
| Combined biochemical and imaging markers | Predictive model (MAFP + uE3) 1/1 + e−[2.876 + 1.333(AFP_MoM) − 6.310(uE3_MoM)] | 0.5 | sens: 61.5% spec: 98.1% | [64] |
| MAFP + MCA-PSV (>1.5 MoM) | - | sens: 97.9% spec: 69.1% | ||
| MAFP + CTR (>0.5) | - | sens: 100.0% spec: 59.3% | ||
| MAFP + Placental thickness (>3.0 cm) | - | sens: 88.9% spec: 69.5% | ||
| MAFP + MCA-PSV + CTR | - | sens: 100.0% spec: 48.1% |
| Advantages | Disadvantages | |
|---|---|---|
| Sonographic markers | Non-invasive technique High predictive performance | Require specialized sonographers Require specific equipment Operator-dependent |
| Biochemical markers | Non-invasive technique Can perform simultaneously with second trimester Down syndrome screening | Poor predictive performance Require validation Other conditions such as infection can interfere with the result |
| Cell-free fetal DNA | Non-invasive technique High predictive performance Can perform simultaneously with Down syndrome screening | Relatively high costs Complex techniques and laboratory device Requires further validation. Thus, the search for other biomarkers is continuing |
| Extracellular vesicles | Non-invasive technique Contain molecular information of dynamic cell and tissue states of disease pathophysiological changes and complications Compatible with multiple etiologies of hydrop fetalis | Relatively high costs Complex techniques and laboratory device Predictive performance requires further studies |
| EV Subpopulation | Diameter | Sedimentation in Centrifugation | Molecular Markers | Cellular Origin |
|---|---|---|---|---|
| Exosomes | 40–150 nm | 100,000–200,000× g for 1–2 h | CD9, CD63, CD81, TSG101, Alix | Multivesicular bodies |
| Microvesicles (or ectosomes) | 100–1000 nm | 10,000–20,000× g for 10–20 min | Integrins, selectins, CD40 ligand | Plasma membrane budding |
| Apoptotic bodies (or apoptotic cell-derived EVs) | 1–5 µm | 1000–2000× g for 10–20 min | Histones, Annexin V | Apoptotic cell membrane blebbing |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaemsaithong, P.; Luewan, S.; Taweevisit, M.; Chiangjong, W.; Pongchaikul, P.; Thorner, P.S.; Tongsong, T.; Chutipongtanate, S. Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease. Int. J. Mol. Sci. 2023, 24, 5658. https://doi.org/10.3390/ijms24065658
Chaemsaithong P, Luewan S, Taweevisit M, Chiangjong W, Pongchaikul P, Thorner PS, Tongsong T, Chutipongtanate S. Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease. International Journal of Molecular Sciences. 2023; 24(6):5658. https://doi.org/10.3390/ijms24065658
Chicago/Turabian StyleChaemsaithong, Piya, Suchaya Luewan, Mana Taweevisit, Wararat Chiangjong, Pisut Pongchaikul, Paul Scott Thorner, Theera Tongsong, and Somchai Chutipongtanate. 2023. "Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease" International Journal of Molecular Sciences 24, no. 6: 5658. https://doi.org/10.3390/ijms24065658
APA StyleChaemsaithong, P., Luewan, S., Taweevisit, M., Chiangjong, W., Pongchaikul, P., Thorner, P. S., Tongsong, T., & Chutipongtanate, S. (2023). Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease. International Journal of Molecular Sciences, 24(6), 5658. https://doi.org/10.3390/ijms24065658