
Genome-Wide Identification and Characterization of Bovine Fibroblast Growth Factor (FGF) Gene and Its Expression during Adipocyte Differentiation

Abstract
:1. Introduction
2. Results
2.1. Identification of Members of the Bovine FGF Family
2.2. Structural Characteristics of Members of the Bovine FGF Family
2.3. Phylogenetic Analysis of Bovine FGF Protein
2.4. Chromosome Distribution and Collinearity Analysis of FGF Gene
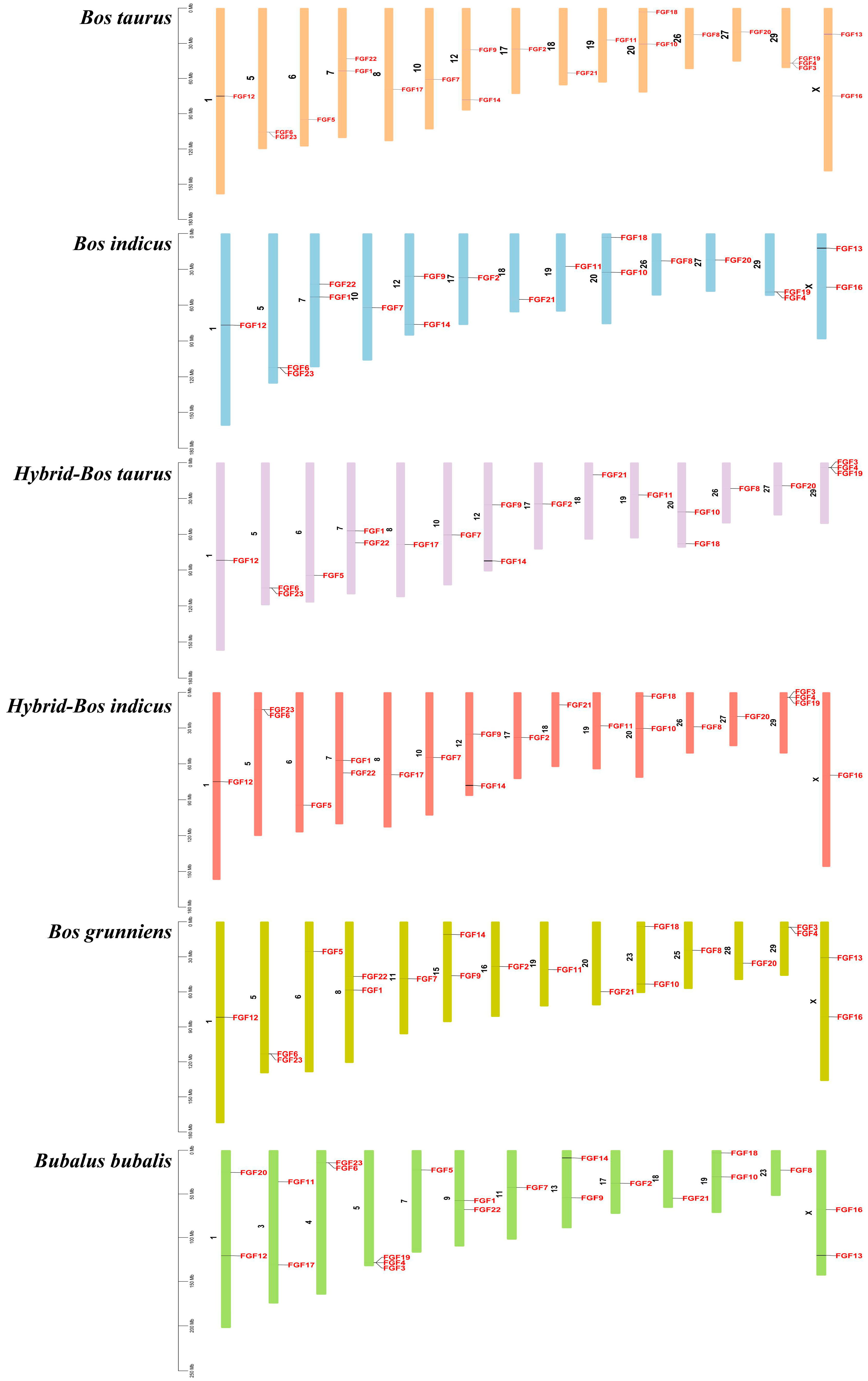
2.5. Collinear Analysis of FGF Gene in Several Bovine Subfamily Species
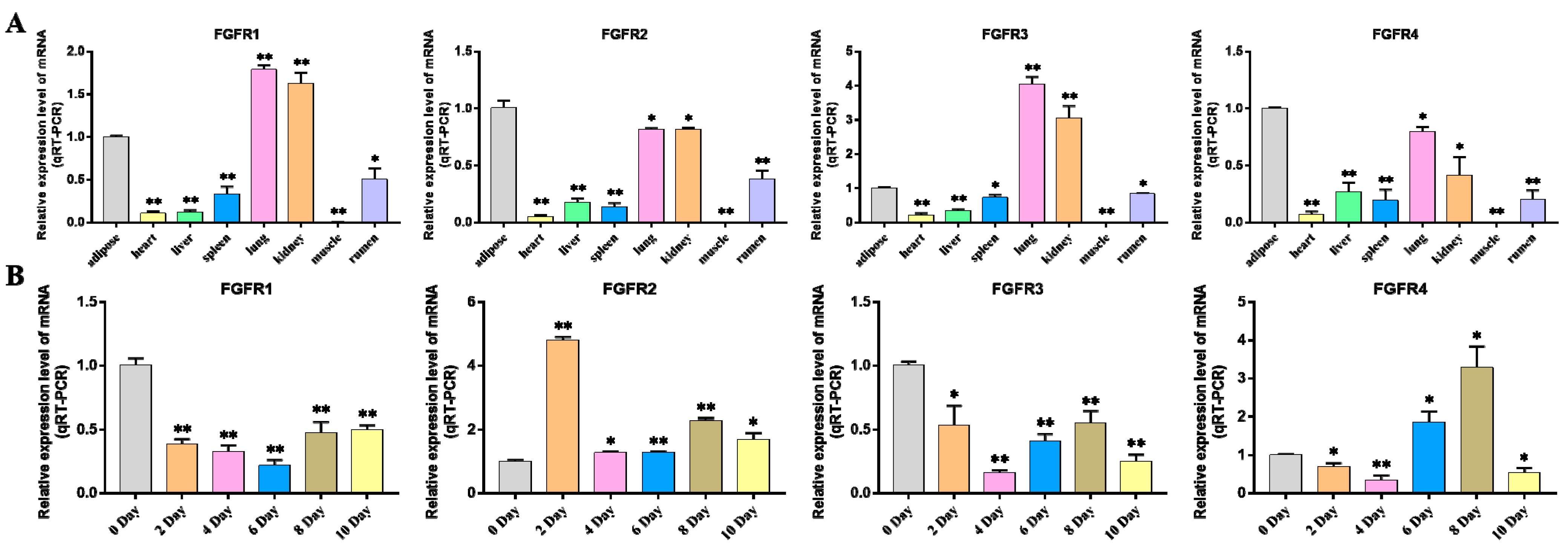
2.6. Expression Analysis of FGF Gene in Different Tissues
2.7. Expression Analysis of FGF Gene during Differentiation of Bovine Adipocytes
3. Discussion
3.1. Identification and Phylogenetic Analysis of Bovine FGF Family Proteins
3.2. Analysis of Physicochemical Properties and Structural Characteristics of the Bovine FGF Protein
3.3. Chromosome Distribution, Replication, and Collinearity Analysis of Bovine FGF Protein
3.4. FGF Gene Affects Adipocyte Differentiation
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of FGF Gene
4.2. Conservative Motif and Gene Structure Analysis
4.3. Chromosome Distribution, Gene Replication, and Collinearity Analysis
4.4. Culture and Induced Differentiation of Bovine Primary Adipocytes
4.5. RNA Extraction and qRT-PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kazala, E.C.; Lozeman, F.J.; Mir, P.S.; Laroche, A.; Bailey, D.R.C.; Weselake, R.J. Relationship of fatty acid composition to intramuscular fat content in beef from crossbred Wagyu cattle. J. Anim. Sci. 1999, 77, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.E.; O’Rahilly, S.; Rochford, J.J. Adipogenesis at a glance. J. Cell Sci. 2011, 124, 2681–2686. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Song, T.-J.; Li, X.; Hu, L.; He, Q.; Liu, M.; Lane, M.D.; Tang, Q.-Q. BMP signaling pathway is required for commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2009, 106, 12670–12675. [Google Scholar] [CrossRef] [Green Version]
- Widberg, C.H.; Newell, F.S.; Bachmann, A.W.; Ramnoruth, S.N.; Spelta, M.C.; Whitehead, J.P.; Hutley, L.J.; Prins, J.B. Fibroblast growth factor receptor 1 is a key regulator of early adipogenic events in human preadipocytes. Am. J. Physiol. Metab. 2009, 296, E121–E131. [Google Scholar] [CrossRef] [Green Version]
- Ohta, H.; Itoh, N. Roles of FGFs as Adipokines in Adipose Tissue Development, Remodeling, and Metabolism. Front. Endocrinol. 2014, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Eswarakumar, V.; Lax, I.; Schlessinger, J. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev. 2005, 16, 139–149. [Google Scholar] [CrossRef]
- Shamsi, F.; Xue, R.; Huang, T.L.; Lundh, M.; Liu, Y.; Leiria, L.O.; Lynes, M.D.; Kempf, E.; Wang, C.-H.; Sugimoto, S.; et al. FGF6 and FGF9 regulate UCP1 expression independent of brown adipogenesis. Nat. Commun. 2020, 11, 1421. [Google Scholar] [CrossRef] [Green Version]
- Mathes, S.; Fahrner, A.; Ghoshdastider, U.; Rüdiger, H.A.; Leunig, M.; Wolfrum, C.; Krützfeldt, J. FGF-2–dependent signaling activated in aged human skeletal muscle promotes intramuscular adipogenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2021013118. [Google Scholar] [CrossRef] [PubMed]
- Kakudo, N.; Morimoto, N.; Ogawa, T.; Taketani, S.; Kusumoto, K. FGF-2 combined with bilayer artificial dermis composed of collagen matrix prompts generation of fat pad in subcutis of mice. Med. Mol. Morphol. 2019, 52, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, H.; Konishi, M.; Itoh, N. FGF10 and FGF21 as regulators in adipocyte development and metabolism. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 302–309. [Google Scholar] [CrossRef]
- Giralt, M.; Gavaldà-Navarro, A.; Villarroya, F. Fibroblast growth factor-21, energy balance and obesity. Mol. Cell. Endocrinol. 2015, 418, 66–73. [Google Scholar] [CrossRef]
- Thisse, B.; Thisse, C. Functions and regulations of fibroblast growth factor signaling during embryonic development. Dev. Biol. 2005, 287, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, R.T.; Niehrs, C. Fibroblast Growth Factor Signaling during Early Vertebrate Development. Endocr. Rev. 2005, 26, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Ornitz, D.M. Evolution of the Fgf and Fgfr gene families. Trends Genet. 2004, 20, 563–569. [Google Scholar] [CrossRef]
- Itoh, N.; Konishi, M.; Simmons, A.E.; Karimi, I.; Talwar, M.; Simmons, T.W.; Su, F.; Juarez, M.A.; Cooke, C.L.; LaPointe, L.; et al. The Zebrafish fgf Family. Zebrafish 2007, 4, 179–186. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, J.; Sun, Z.; Tang, Y.; Wu, Y. Genome-Wide Identification and Analysis of the Polycomb Group Family in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 7537. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, S.; Dong, C.; Chen, B.; Feng, J.; Peng, W.; Mahboob, S.; Al-Ghanim, K.A.; Xu, P. Genome-wide identification, phylogeny, and expression of fibroblast growth genes in common carp. Gene 2016, 578, 225–231. [Google Scholar] [CrossRef]
- Itoh, N.; Ornitz, D. Functional evolutionary history of the mouseFgf gene family. Dev. Dyn. 2008, 237, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Draper, B.W.; Stock, D.W.; Kimmel, C.B. Zebrafish fgf24 functions with fgf8 to promote posterior mesodermal development. Development 2003, 130, 4639–4654. [Google Scholar] [CrossRef] [Green Version]
- Popovici, C.; Roubin, R.; Coulier, F.; Birnbaum, D. An evolutionary history of the FGF superfamily. Bioessays 2005, 27, 849–857. [Google Scholar] [CrossRef]
- Coulier, F.; Pontarotti, P.; Roubin, R.; Hartung, H.; Goldfarb, M.; Birnbaum, D. Of Worms and Men: An Evolutionary Perspective on the Fibroblast Growth Factor (FGF) and FGF Receptor Families. J. Mol. Evol. 1997, 44, 43–56. [Google Scholar] [CrossRef]
- Oulion, S.; Bertrand, S.; Escriva, H. Evolution of the FGF Gene Family. Int. J. Evol. Biol. 2012, 2012, 298147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanta, T.K.; Khan, A.; Hashem, A.; Abd_Allah, E.F.; Al-Harrasi, A. The molecular mass and isoelectric point of plant proteomes. BMC Genom. 2019, 20, 631. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.L. The evolution of functionally novel proteins after gene duplication. Proc. R. Soc. B Boil. Sci. 1994, 256, 119–124. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol. 2001, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Chen, D.L.; Samocha-Bonet, D.; Gillinder, K.; Barclay, J.; Magor, G.W.; Perkins, A.; Greenfield, J.; Yang, G.; Whitehead, J.P. Fibroblast growth factor-1 (FGF-1) promotes adipogenesis by downregulation of carboxypeptidase A4 (CPA4)—A negative regulator of adipogenesis implicated in the modulation of local and systemic insulin sensitivity. Growth Factors 2016, 34, 210–216. [Google Scholar] [CrossRef]
- Lu, G.-M.; Rong, Y.-X.; Liang, Z.-J.; Hunag, D.-L.; Wu, F.-X.; Ma, Y.-F.; Luo, Z.-Z.; Liu, X.-H.; Mo, S.; Li, H.-M. FGF2-induced PI3K/Akt signaling evokes greater proliferation and adipogenic differentiation of human adipose stem cells from breast than from abdomen or thigh. Aging 2020, 12, 14830–14848. [Google Scholar] [CrossRef]
- Xu, Q.; Lin, S.; Wang, Y.; Zhu, J.; Lin, Y. Fibroblast growth factor 10 (FGF10) promotes the adipogenesis of intramuscular preadipocytes in goat. Mol. Biol. Rep. 2018, 45, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Price, C.A. Mechanisms of fibroblast growth factor signaling in the ovarian follicle. J. Endocrinol. 2016, 228, R31–R43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Ealy, A. Disruption of fibroblast growth factor receptor signaling in bovine cumulus-oocyte complexes during in vitro maturation reduces subsequent embryonic development. Domest. Anim. Endocrinol. 2012, 42, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Xu, J.; Colvin, J.S.; McEwen, D.G.; MacArthur, C.A.; Coulier, F.; Gao, G.; Goldfarb, M. Receptor Specificity of the Fibroblast Growth Factor Family. J. Biol. Chem. 1996, 271, 15292–15297. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor Specificity of the Fibroblast Growth Factor Family: The Complete Mammalian FGF Family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Wilhelm, K.; Dubrac, A.; Tung, J.K.; Alves, T.C.; Fang, J.S.; Xie, Y.; Zhu, J.; Chen, Z.; De Smet, F.; et al. FGF-dependent metabolic control of vascular development. Nature 2017, 545, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Dai, J.; Yang, M.; Deng, G.; Xu, S.; Jia, Y.; Boden, G.; Ma, Z.A.; Yang, G.; Li, L. Silencing of FGF-21 expression promotes hepatic gluconeogenesis and glycogenolysis by regulation of the STAT3-SOCS3 signal. FEBS J. 2014, 281, 2136–2147. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zeng, L.; Wang, Y.-J.; An, Z.-M.; Ying, B.-W. Associations of Fibroblast Growth Factor 21 Gene 3′ Untranslated Region Single-Nucleotide Polymorphisms with Metabolic Syndrome, Obesity, and Diabetes in a Han Chinese Population. DNA Cell Biol. 2012, 31, 547–552. [Google Scholar] [CrossRef]
- Wang, S.; Cao, S.; Arhatte, M.; Li, D.; Shi, Y.; Kurz, S.; Hu, J.; Wang, L.; Shao, J.; Atzberger, A.; et al. Adipocyte Piezo1 mediates obesogenic adipogenesis through the FGF1/FGFR1 signaling pathway in mice. Nat. Commun. 2020, 11, 2303. [Google Scholar] [CrossRef]
- Johnson, L.S.; Eddy, S.R.; Portugaly, E. Hidden Markov model speed heuristic and iterative HMM search procedure. BMC Bioinform. 2010, 11, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, K.-F.; Chen, J.; Chen, Y.-F.; Wu, L.-J.; Xie, D.-X. Molecular Phylogenetic and Expression Analysis of the Complete WRKY Transcription Factor Family in Maize. DNA Res. 2012, 19, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Lei, Z.; Wang, S.; Wang, X.; Wei, D.; Cai, X.; Luoreng, Z.; Wang, L.; Ma, Y. Genome-wide identification of cyclin-dependent kinase (CDK) genes affecting adipocyte differentiation in cattle. BMC Genom. 2021, 22, 532. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zheng, Q.; Wang, S.; Wei, X.; Li, F.; Ma, Y. High-Throughput RNA Sequencing Reveals NDUFC2-AS lncRNA Promotes Adipogenic Differentiation in Chinese Buffalo (Bubalus bubalis L.). Genes 2019, 10, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Goulart, R.A.; Pantanowitz, L. Oil red O staining in cytopathology. Diagn. Cytopathol. 2011, 39, 272–273. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Morton, G.; Hadi, S. Analysis of rpoS and bolA gene expression under various stress-induced environments in planktonic and biofilm phase using 2−ΔΔCT method. Mol. Cell. Biochem. 2011, 357, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Arocho, A.; Chen, B.; Ladanyi, M.; Pan, Q. Validation of the 2-−ΔΔCt Calculation as an Alternate Method of Data Analysis for Quantitative PCR of BCR-ABL P210 Transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene ID | Transcript ID | pI | Mw/Da | Amino Acids | Description |
|---|---|---|---|---|---|---|
| FGF1 | ENSBTAG00000005198 | ENSBTAT00000078959 | 6.51 | 17,492.86 | 156 | Fibroblast growth factor 1 |
| FGF2 | ENSBTAG00000005691 | ENSBTAT00000007477 | 9.58 | 17,249.83 | 156 | Fibroblast growth factor 2 |
| FGF3 | ENSBTAG00000008623 | ENSBTAT00000011373 | 11.44 | 26,672.65 | 237 | Fibroblast growth factor 3 |
| FGF4 | ENSBTAG00000012563 | ENSBTAT00000029946 | 9.89 | 22,068.57 | 207 | Fibroblast growth factor 4 |
| FGF5 | ENSBTAG00000017348 | ENSBTAT00000023064 | 10.67 | 29,640.87 | 271 | Fibroblast growth factor 5 |
| FGF6 | ENSBTAG00000006800 | ENSBTAT00000016675 | 10.24 | 22,623.16 | 209 | Fibroblast growth factor 6 |
| FGF7 | ENSBTAG00000051898 | ENSBTAT00000073054 | 9.35 | 22,489.17 | 195 | Fibroblast growth factor 7 |
| FGF8 | ENSBTAG00000001530 | ENSBTAT00000002001 | 10.36 | 27,605.6 | 245 | Fibroblast growth factor 8 |
| FGF9 | ENSBTAG00000048237 | ENSBTAT00000064356 | 7.06 | 23,382.45 | 209 | Fibroblast growth factor 9 |
| FGF10 | ENSBTAG00000051910 | ENSBTAT00000086367 | 9.61 | 23,768.15 | 214 | Fibroblast growth factor 10 |
| FGF11 | ENSBTAG00000019242 | ENSBTAT00000025622 | 10.06 | 25,081.1 | 226 | Fibroblast growth factor 11 |
| FGF12 | ENSBTAG00000012413 | ENSBTAT00000083812 | 9.28 | 23,236.66 | 208 | Fibroblast growth factor 12 |
| FGF13 | ENSBTAG00000051480 | ENSBTAT00000075597 | 9.15 | 28,757.89 | 256 | Fibroblast growth factor 13 |
| FGF14 | ENSBTAG00000052496 | ENSBTAT00000070063 | 9.32 | 20,378.4 | 185 | Fibroblast growth factor 14 |
| FGF16 | ENSBTAG00000006722 | ENSBTAT00000008839 | 9.36 | 23,791.89 | 208 | Fibroblast growth factor 16 |
| FGF17 | ENSBTAG00000046951 | ENSBTAT00000065993 | 10.5 | 24,919.41 | 217 | Fibroblast growth factor 17 |
| FGF18 | ENSBTAG00000000128 | ENSBTAT00000000139 | 9.86 | 23,920.57 | 208 | Fibroblast growth factor 18 |
| FGF19 | ENSBTAG00000017285 | ENSBTAT00000022973 | 7.7 | 24,072.59 | 219 | Fibroblast growth factor 19 |
| FGF20 | ENSBTAG00000044043 | ENSBTAT00000061318 | 8.54 | 23,489.59 | 212 | Fibroblast growth factor 20 |
| FGF21 | ENSBTAG00000011624 | ENSBTAT00000015438 | 6.08 | 22,585.83 | 210 | Fibroblast growth factor 21 |
| FGF22 | ENSBTAG00000027357 | ENSBTAT00000086016 | 11.48 | 23,305.82 | 219 | Fibroblast growth factor 22 |
| FGF23 | ENSBTAG00000030343 | ENSBTAT00000008940 | 9.64 | 26,733.24 | 246 | Fibroblast growth factor 23 |
| Gene | Bos indicus | Hybrid-Bos taurus | Hybrid-Bos indicus | Bos grunnines | Bubalus bubalis |
|---|---|---|---|---|---|
| FGF1 | Y | Y | Y | Y | Y |
| FGF2 | Y | Y | Y | Y | Y |
| FGF3 | - | Y | Y | Y | Y |
| FGF4 | Y | Y | Y | Y | Y |
| FGF5 | - | Y | Y | Y | Y |
| FGF6 | Y | Y | Y | Y | Y |
| FGF7 | Y | Y | Y | Y | Y |
| FGF8 | Y | Y | Y | Y | Y |
| FGF9 | Y | Y | Y | Y | N |
| FGF10 | Y | Y | Y | Y | Y |
| FGF11 | N | N | N | N | N |
| FGF12 | Y | Y | Y | Y | Y |
| FGF13 | Y | - | - | Y | Y |
| FGF14 | Y | Y | Y | Y | - |
| FGF16 | Y | - | Y | Y | Y |
| FGF17 | - | Y | Y | - | Y |
| FGF18 | Y | Y | Y | Y | Y |
| FGF19 | Y | Y | Y | - | Y |
| FGF20 | Y | Y | Y | Y | Y |
| FGF21 | Y | Y | Y | Y | Y |
| FGF22 | Y | Y | Y | Y | Y |
| FGF23 | Y | Y | Y | Y | Y |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, H.; Zhang, J.; Li, F.; Pan, C.; Yang, M.; Liu, Y.; Cai, B.; Zhang, L.; Ma, Y. Genome-Wide Identification and Characterization of Bovine Fibroblast Growth Factor (FGF) Gene and Its Expression during Adipocyte Differentiation. Int. J. Mol. Sci. 2023, 24, 5663. https://doi.org/10.3390/ijms24065663
Sheng H, Zhang J, Li F, Pan C, Yang M, Liu Y, Cai B, Zhang L, Ma Y. Genome-Wide Identification and Characterization of Bovine Fibroblast Growth Factor (FGF) Gene and Its Expression during Adipocyte Differentiation. International Journal of Molecular Sciences. 2023; 24(6):5663. https://doi.org/10.3390/ijms24065663
Chicago/Turabian StyleSheng, Hui, Junxing Zhang, Fen Li, Cuili Pan, Mengli Yang, Yuan Liu, Bei Cai, Lingkai Zhang, and Yun Ma. 2023. "Genome-Wide Identification and Characterization of Bovine Fibroblast Growth Factor (FGF) Gene and Its Expression during Adipocyte Differentiation" International Journal of Molecular Sciences 24, no. 6: 5663. https://doi.org/10.3390/ijms24065663
APA StyleSheng, H., Zhang, J., Li, F., Pan, C., Yang, M., Liu, Y., Cai, B., Zhang, L., & Ma, Y. (2023). Genome-Wide Identification and Characterization of Bovine Fibroblast Growth Factor (FGF) Gene and Its Expression during Adipocyte Differentiation. International Journal of Molecular Sciences, 24(6), 5663. https://doi.org/10.3390/ijms24065663