
Adipose-Derived Stem Cells in Reinforced Collagen Gel: A Comparison between Two Approaches to Differentiation towards Smooth Muscle Cells

 , , , , , ,
, , , , , ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
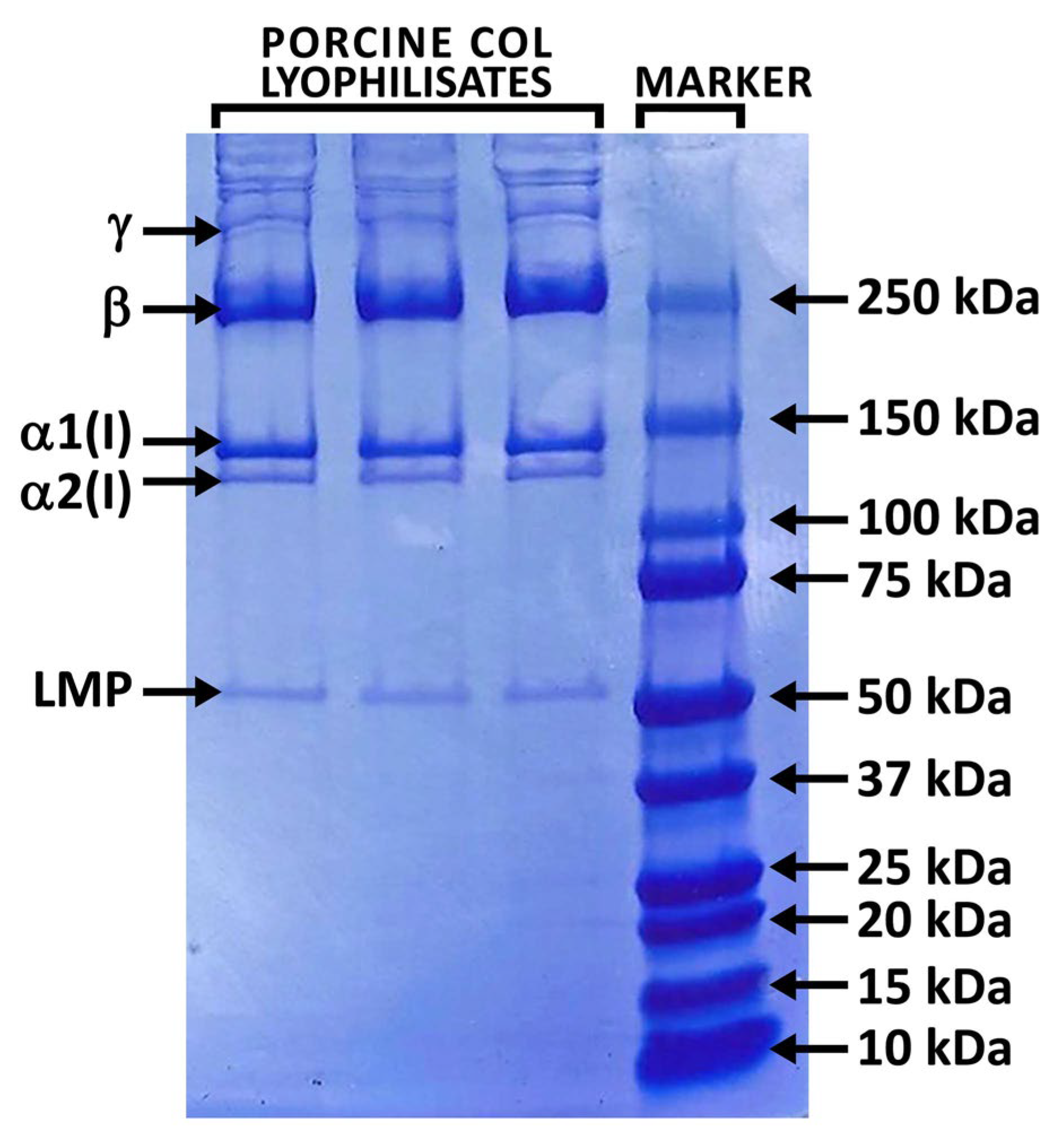
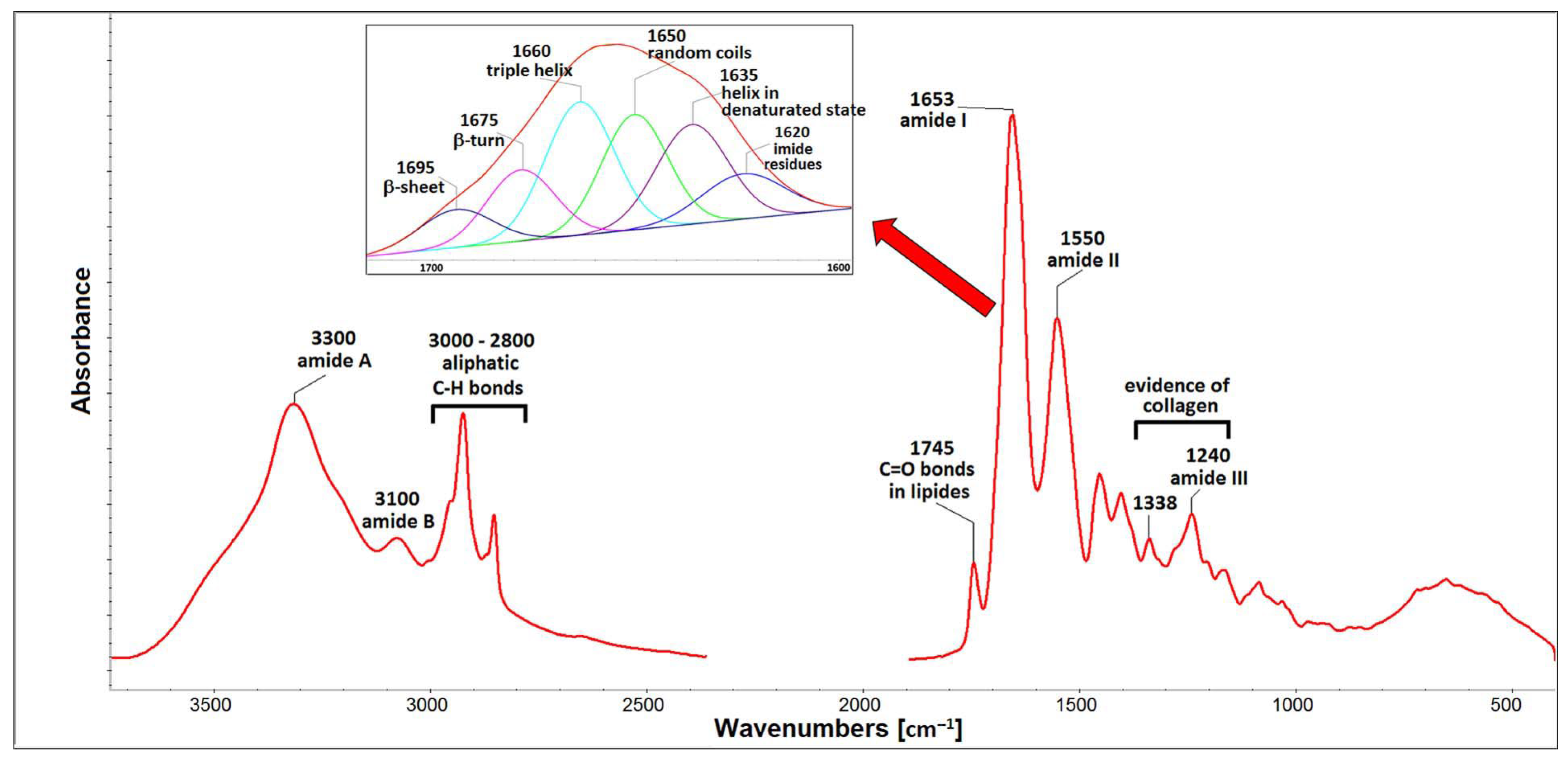
2.1. Properties of Isolated Porcine Collagen
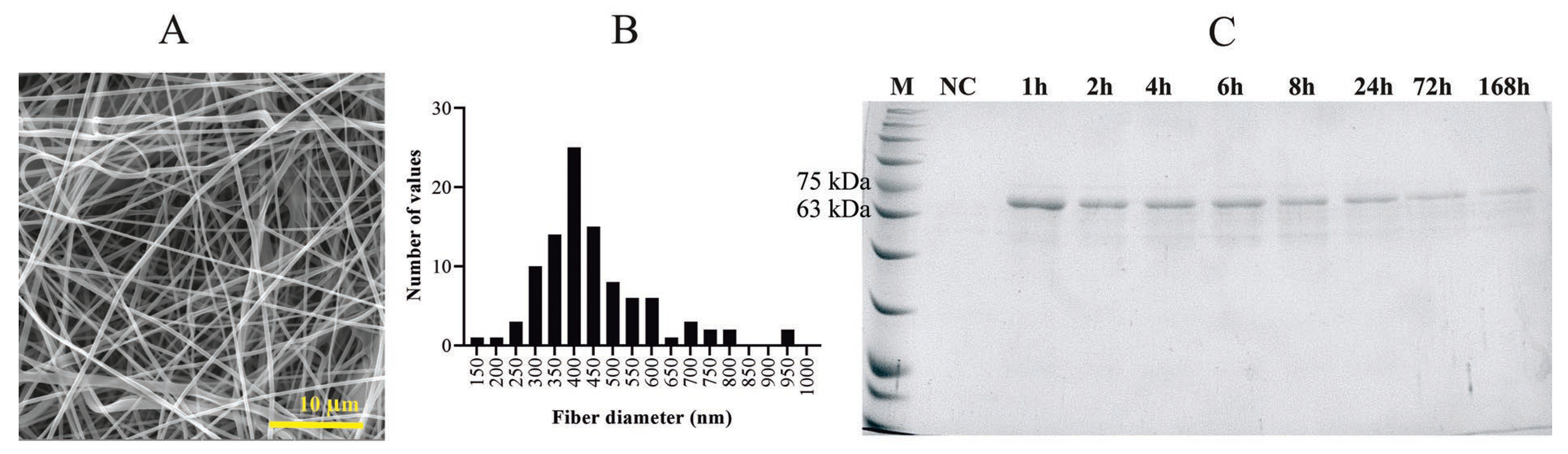
2.2. Morphology of Collagen Fibers and Particles
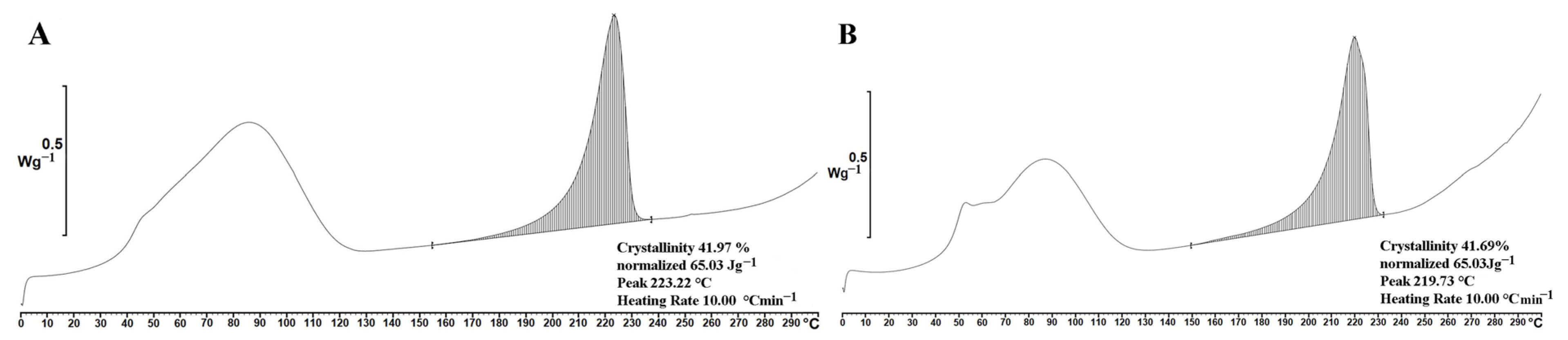
2.3. Properties of PVA and PVA_PL Meshes
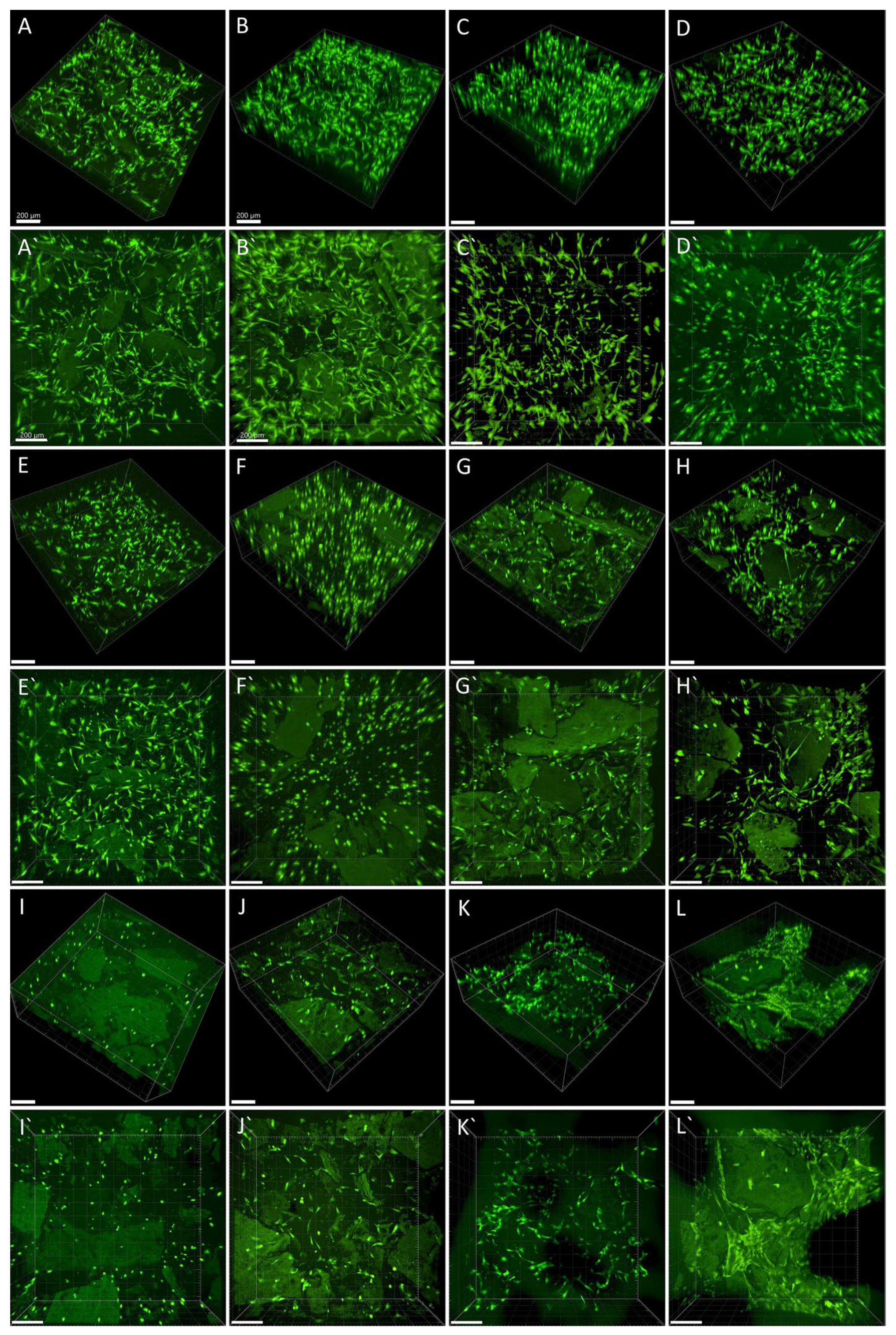
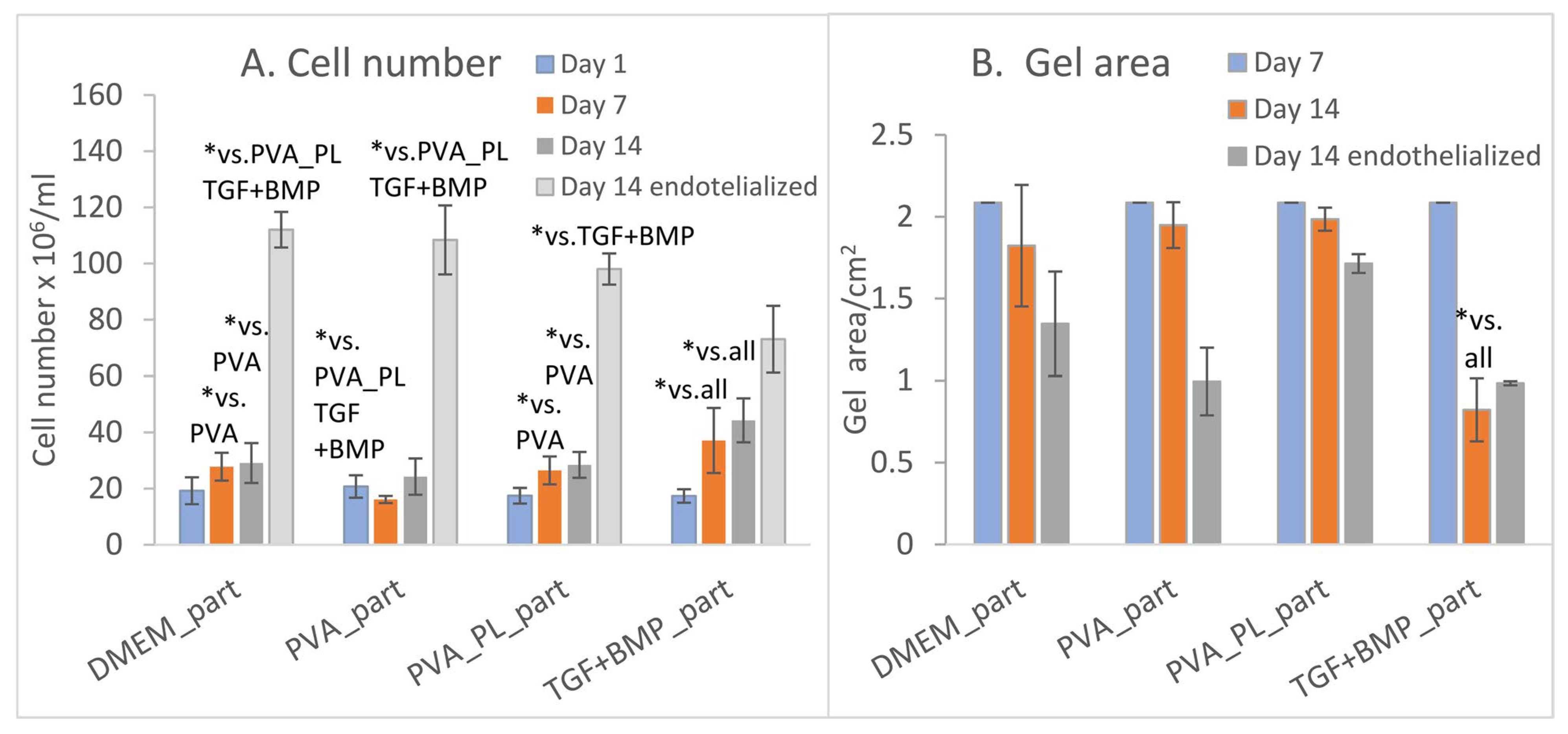
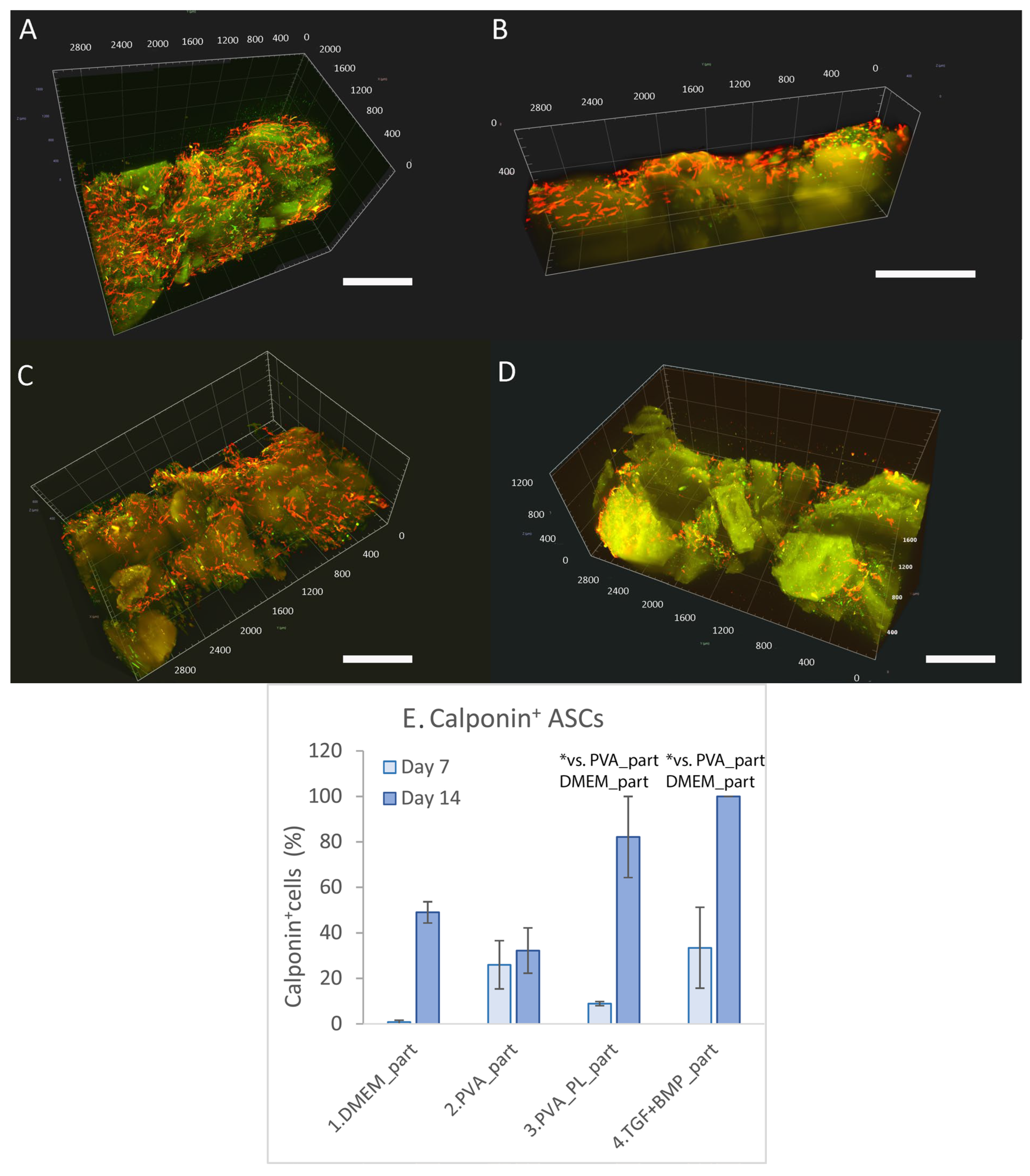
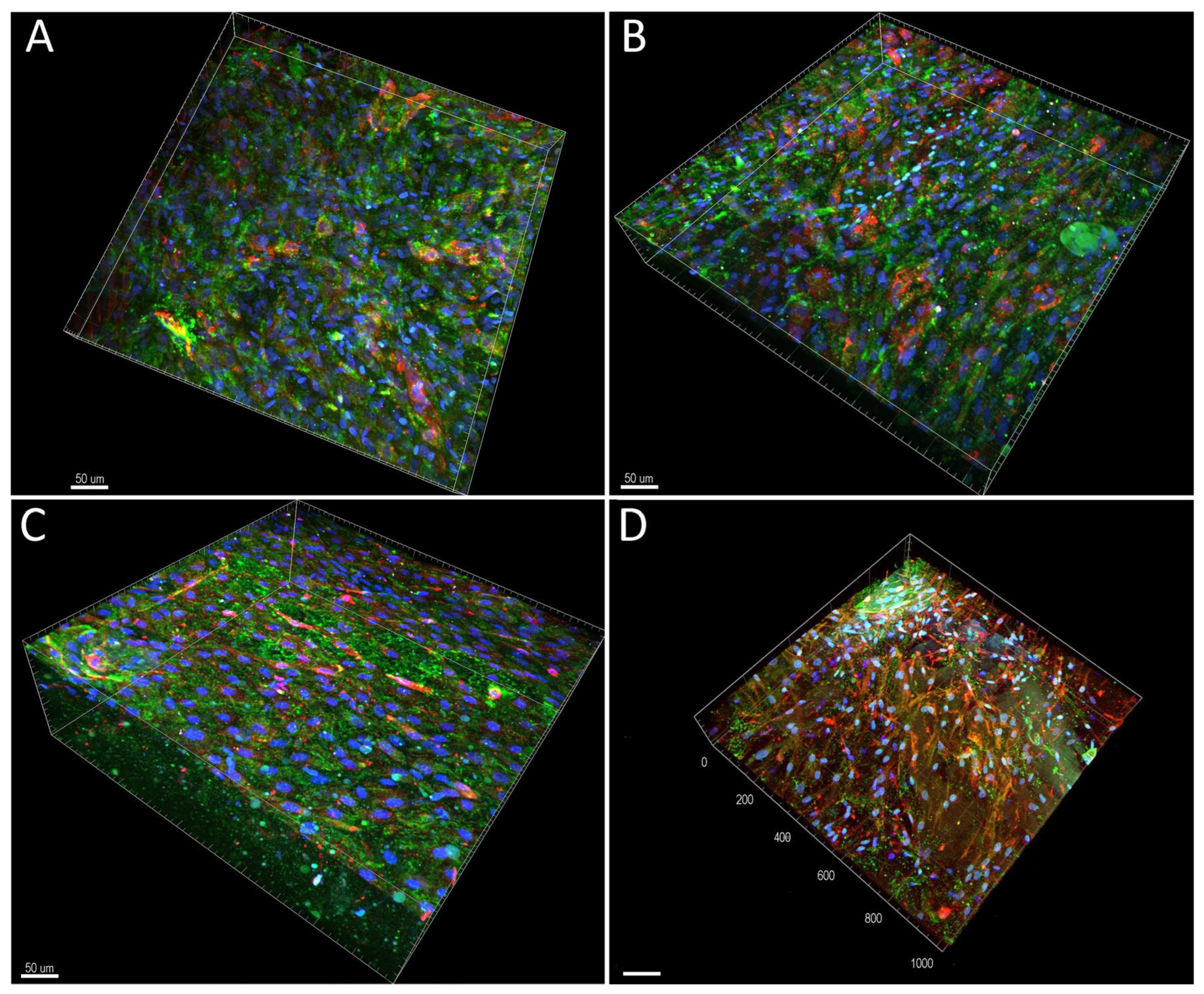
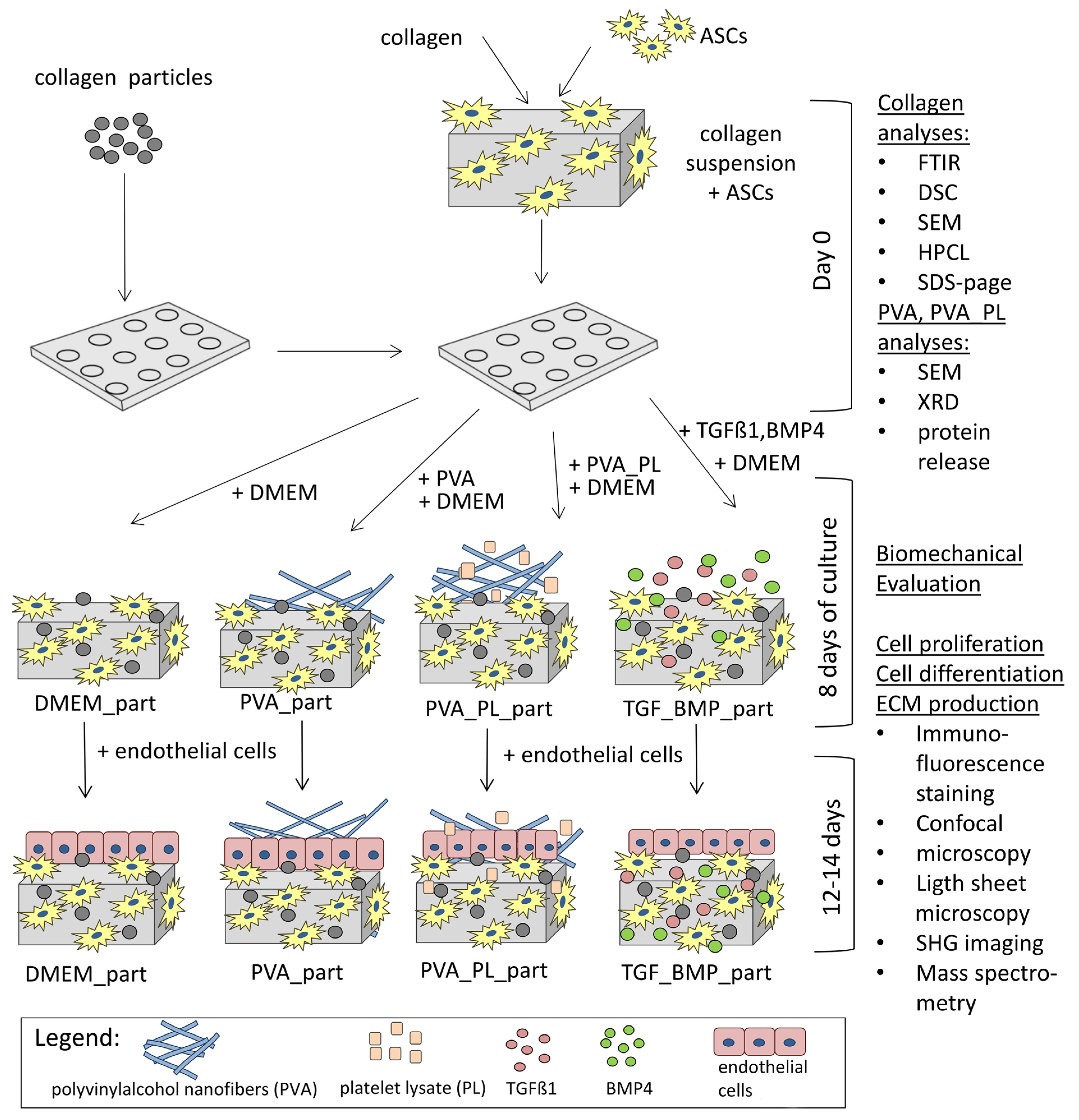
2.4. Cell Colonisation of Collagen Gels Reinforced with Collagen Particles
2.5. Proteomics of the Cell-Material Constructs
2.5.1. Proteins Involved in Cell Differentiation
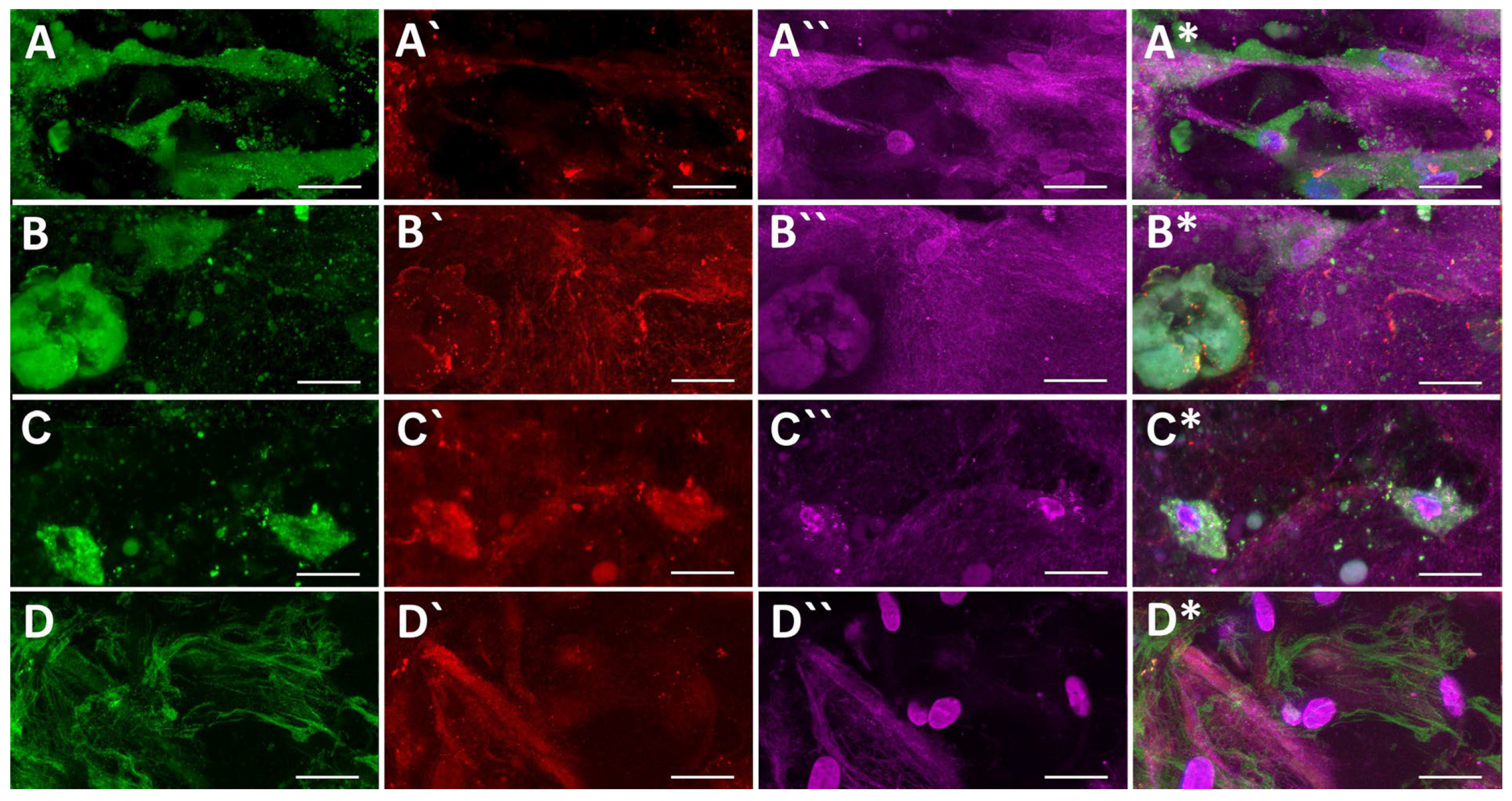
2.5.2. ECM Proteins
2.5.3. Remodelling Proteins
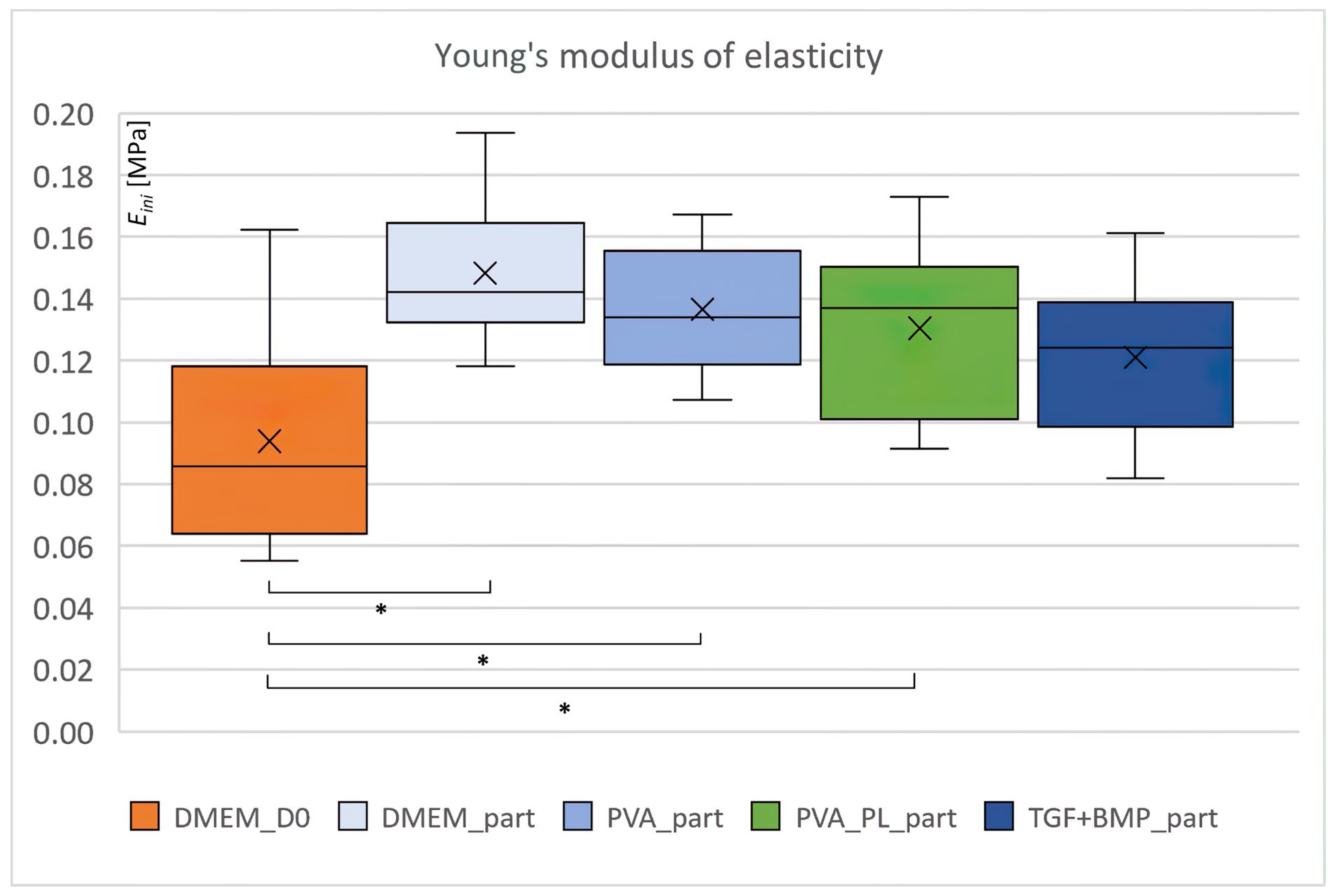
2.6. Biomechanical Properties
3. Discussion
3.1. Collagen Gel Preparation, Gel Reinforcement
3.2. Cellular Component of the Gel, Cell Differentiation
3.3. Mechanical Properties of the Constructs
4. Materials and Methods
4.1. Isolation of Collagen
4.2. Preparation of Collagen Submicrometer Fibers and Particles
4.3. Analyses of Porcine Collagen Composition
4.4. Morphology of Collagen Submicrometer Fibers and Particles
4.5. Platelet Lysate Preparation
4.6. PVA_PL Nanofibrous Mat Preparation
4.7. Protein Content/Release Analysis of PVA_PL
4.8. PVA Crystallinity Analysis
4.9. Cell Isolation, Cultivation and Characterisation
4.10. Preparation of Collagen Hydrogels with Cells, Cell Cultivation
4.11. Visualisation of Living Cells
4.12. Immunofluorescence Staining of Cell Differentiation Markers and ECM Components
4.13. Confocal Microscopy
4.14. Lightsheet Microscopy
4.15. Nonlinear Imaging—Second Harmonic Generation
4.16. Bruker Microscopy
4.17. Evaluation of Biomechanical Properties
4.18. Mass Spectrometry—Label Free Quantification (MS LFQ)
4.19. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organisation. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 8 March 2021).
- Moore, M.J.; Tan, R.P.; Yang, N.; Rnjak-Kovacina, J.; Wise, S.G. Bioengineering artificial blood vessels from natural materials. Trends Biotechnol. 2022, 40, 693–707. [Google Scholar] [CrossRef]
- Ebert, N.; McGinnis, M.; Johnson, W.; Kuhn, E.M.; Mitchell, M.E.; Tweddell, J.S.; Woods, R.K. Comparison of Patch Materials for Pulmonary Artery Reconstruction. Semin. Thorac. Cardiovasc. Surg. 2021, 33, 459–465. [Google Scholar] [CrossRef]
- Zhang, F.; Xie, Y.; Celik, H.; Akkus, O.; Bernacki, S.H.; King, M.W. Engineering small-caliber vascular grafts from collagen filaments and nanofibers with comparable mechanical properties to native vessels. Biofabrication 2019, 11, 035020. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, J.; Cui, Y.; Xu, R.; Wang, Z.; Zhang, J.; Wang, K.; Li, Y.; Zhao, Q.; Kong, D. Effect of sustained heparin release from PCL/chitosan hybrid small-diameter vascular grafts on anti-thrombogenic property and endothelialisation. Acta Biomater. 2014, 10, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, N.; Amorim, I.; Caseiro, A.R.; Pereira, T.; Alvites, R.; Rema, A.; Goncalves, A.; Valadares, G.; Costa, E.; Santos-Silva, A.; et al. Long term performance evaluation of small-diameter vascular grafts based on polyvinyl alcohol hydrogel and dextran and MSCs-based therapies using the ovine pre-clinical animal model. Int. J. Pharm. 2017, 523, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Babrnakova, J.; Pavlinakova, V.; Brtnikova, J.; Sedlacek, P.; Prosecka, E.; Rampichova, M.; Filova, E.; Hearnden, V.; Vojtova, L. Synergistic effect of bovine platelet lysate and various polysaccharides on the biological properties of collagen-based scaffolds for tissue engineering: Scaffold preparation, chemo-physical characterisation, in vitro and ex ovo evaluation. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 236–246. [Google Scholar] [CrossRef]
- Cho, S.W.; Park, H.J.; Ryu, J.H.; Kim, S.H.; Kim, Y.H.; Choi, C.Y.; Lee, M.J.; Kim, J.S.; Jang, I.S.; Kim, D.I.; et al. Vascular patches tissue-engineered with autologous bone marrow-derived cells and decellularised tissue matrices. Biomaterials 2005, 26, 1915–1924. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Zhou, J.; Wang, J.; Zhen, M.; Liu, Y.; Chen, J.; Qi, Z. The development of a tissue-engineered artery using decellularised scaffold and autologous ovine mesenchymal stem cells. Biomaterials 2010, 31, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Malladi, S.; Miranda-Nieves, D.; Leng, L.; Grainger, S.J.; Tarabanis, C.; Nesmith, A.P.; Kosaraju, R.; Haller, C.A.; Parker, K.K.; Chaikof, E.L.; et al. Continuous Formation of Ultrathin, Strong Collagen Sheets with Tunable Anisotropy and Compaction. ACS Biomater. Sci. Eng. 2020, 6, 4236–4246. [Google Scholar] [CrossRef]
- Patil, V.A.; Masters, K.S. Engineered Collagen Matrices. Bioengineering 2020, 7, 163. [Google Scholar] [CrossRef]
- Aras, O.; Kazanci, M. Production of collagen micro- and nanofibers for potential drug-carrier systems. J. Enzym. Inhib. Med. Chem. 2015, 30, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.M.; Ahn, H.; Arenas-Herrera, J.; Kim, C.; Abolbashari, M.; Atala, A.; Yoo, J.J.; Lee, S.J. Electrospun vascular scaffold for cellularised small diameter blood vessels: A preclinical large animal study. Acta Biomater. 2017, 59, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Meghezi, S.; Seifu, D.G.; Bono, N.; Unsworth, L.; Mequanint, K.; Mantovani, D. Engineering 3D Cellularised Collagen Gels for Vascular Tissue Regeneration. J. Vis. Exp. 2015, 100, e52812. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.A.; Caves, J.M.; Haller, C.A.; Dai, E.; Liu, L.; Grainger, S.; Chaikof, E.L. Acellular vascular grafts generated from collagen and elastin analogs. Acta Biomater. 2013, 9, 8067–8074. [Google Scholar] [CrossRef] [Green Version]
- Matsuhashi, A.; Nam, K.; Kimura, T.; Kishida, A. Fabrication of fibrillised collagen microspheres with the microstructure resembling an extracellular matrix. Soft Matter 2015, 11, 2844–2851. [Google Scholar] [CrossRef]
- Hu, Y.; Dan, W.; Xiong, S.; Kang, Y.; Dhinakar, A.; Wu, J.; Gu, Z. Development of collagen/polydopamine complexed matrix as mechanically enhanced and highly biocompatible semi-natural tissue engineering scaffold. Acta Biomater. 2017, 47, 135–148. [Google Scholar] [CrossRef]
- Dewle, A.; Rakshasmare, P.; Srivastava, A. A Polycaprolactone (PCL)-Supported Electrocompacted Aligned Collagen Type-I Patch for Annulus Fibrosus Repair and Regeneration. ACS Appl. Bio Mater. 2021, 4, 1238–1251. [Google Scholar] [CrossRef]
- Leite, F.G.; Marana, J.F.; de Sá, L.F.T.; Alves de Almeida, T.F.R.; do Carmo, H.R.P.; Chaud, M.V.; Grotto, D.; Silveira-Filho, L.D.M. Effects of a collagen hyaluronic acid silk-fibroin patch with the electroconductive element polyaniline on left ventricular remodelling in an infarct heart model. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 1651–1666. [Google Scholar] [CrossRef]
- Goel, H.; Gupta, N.; Santhiya, D.; Dey, N.; Bohidar, H.B.; Bhattacharya, A. Bioactivity reinforced surface patch bound collagen-pectin hydrogel. Int. J. Biol. Macromol. 2021, 174, 240–253. [Google Scholar] [CrossRef]
- Wertheimer, S.; Sharabi, M.; Shelah, O.; Lesman, A.; Haj-Ali, R. Bio-composites reinforced with unique coral collagen fibers: Towards biomimetic-based small diameter vascular grafts. J. Mech. Behav. Biomed. 2021, 119, 104526. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, Y.; Dan, N.; Li, Z.; Dan, W. Anti-calcification potential of collagen based biological patch crosslinked by epoxidised polysaccharide. Int. J. Biol. Macromol. 2022, 209, 1695–1702. [Google Scholar] [CrossRef]
- Hong, H.; Kim, J.; Cho, H.; Park, S.M.; Jeon, M.; Kim, H.K.; Kim, D.S. Ultra-stiff compressed collagen for corneal perforation patch graft realised by in situ photochemical crosslinking. Biofabrication 2020, 12, 045030. [Google Scholar] [CrossRef]
- Yan, M.; An, X.; Duan, S.; Jiang, Z.; Liu, X.; Zhao, X.; Li, Y. A comparative study on cross-linking of fibrillar gel prepared by tilapia collagen and hyaluronic acid with EDC/NHS and genipin. Int. J. Biol. Macromol. 2022, 213, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Orban, J.M.; Wilson, L.B.; Kofroth, J.A.; El-Kurdi, M.S.; Maul, T.M.; Vorp, D.A. Crosslinking of collagen gels by transglutaminase. J. Biomed. Mater. Res. A 2004, 68, 756–762. [Google Scholar] [CrossRef]
- Adamiak, K.; Sionkowska, A. Current methods of collagen cross-linking: Review. Int. J. Biol. Macromol. 2020, 161, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Maarof, M.; Mh Busra, M.F.; Lokanathan, Y.; Bt Hj Idrus, R.; Rajab, N.F.; Chowdhury, S.R. Safety and efficacy of dermal fibroblast conditioned medium (DFCM) fortified collagen hydrogel as acellular 3D skin patch. Drug Deliv. Transl. Res. 2019, 9, 144–161. [Google Scholar] [CrossRef] [PubMed]
- Filova, E.; Steinerova, M.; Travnickova, M.; Knitlova, J.; Musilkova, J.; Eckhardt, A.; Hadraba, D.; Matejka, R.; Prazak, S.; Stepanovska, J.; et al. Accelerated in vitro recellularisation of decellularised porcine pericardium for cardiovascular grafts. Biomed. Mater. 2021, 16, 025024. [Google Scholar] [CrossRef]
- Sun, B.; Chen, B.; Zhao, Y.; Sun, W.; Chen, K.; Zhang, J.; Wei, Z.; Xiao, Z.; Dai, J. Crosslinking heparin to collagen scaffolds for the delivery of human platelet-derived growth factor. J. Biomed. Mater. Res. B Appl. Biomater. 2009, 91, 366–372. [Google Scholar] [CrossRef]
- Miyagi, Y.; Chiu, L.L.; Cimini, M.; Weisel, R.D.; Radisic, M.; Li, R.K. Biodegradable collagen patch with covalently immobilised VEGF for myocardial repair. Biomaterials 2011, 32, 1280–1290. [Google Scholar] [CrossRef]
- Chiu, L.L.; Radisic, M. Scaffolds with covalently immobilised VEGF and Angiopoietin-1 for vascularisation of engineered tissues. Biomaterials 2010, 31, 226–241. [Google Scholar] [CrossRef]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem cells: Their source, potency and use in regenerative therapies with focus on adipose-derived stem cells—A review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Geng, Z.; Nickel, T.; Johnson, C.; Gao, L.; Dutton, J.; Hou, C.; Zhang, J. Differentiation of Human Induced-Pluripotent Stem Cells into Smooth-Muscle Cells: Two Novel Protocols. PLoS ONE 2016, 11, e0147155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogi, A.; Rukhlova, M.; Charlebois, C.; Tian, G.; Stanimirovic, D.B.; Moreno, M.J. Differentiation of Adipose-Derived Stem Cells into Vascular Smooth Muscle Cells for Tissue Engineering Applications. Biomedicines 2021, 9, 797. [Google Scholar] [CrossRef] [PubMed]
- Walters, B.; Turner, P.A.; Rolauffs, B.; Hart, M.L.; Stegemann, J.P. Controlled Growth Factor Delivery and Cyclic Stretch Induces a Smooth Muscle Cell-like Phenotype in Adipose-Derived Stem Cells. Cells 2021, 10, 3123. [Google Scholar] [CrossRef] [PubMed]
- Astori, G.; Amati, E.; Bambi, F.; Bernardi, M.; Chieregato, K.; Schafer, R.; Sella, S.; Rodeghiero, F. Platelet lysate as a substitute for animal serum for the ex-vivo expansion of mesenchymal stem/stromal cells: Present and future. Stem Cell Res. Ther. 2016, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Hwang, S.R.; Yoon, I.S. Advanced Growth Factor Delivery Systems in Wound Management and Skin Regeneration. Molecules 2017, 22, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elcin, A.E.; Parmaksiz, M.; Dogan, A.; Seker, S.; Durkut, S.; Dalva, K.; Elcin, Y.M. Differential gene expression profiling of human adipose stem cells differentiating into smooth muscle-like cells by TGFbeta1/BMP4. Exp. Cell Res. 2017, 352, 207–217. [Google Scholar] [CrossRef]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Osako, K. Type I collagen from the skin of ornate threadfin bream (Nemipterus hexodon): Characteristics and effect of pepsin hydrolysis. Food Chem. 2011, 125, 500–507. [Google Scholar] [CrossRef]
- Veeruraj, A.; Arumugam, M.; Ajithkumar, T.; Balasubramanian, T. Isolation and characterisation of collagen from the outer skin of squid (Doryteuthis singhalensis). Food Hydrocoll. 2015, 43, 708–716. [Google Scholar] [CrossRef]
- Payne, K.J.; Veis, A. Fourier-Transform Ir Spectroscopy of Collagen and Gelatin Solutions—Deconvolution of the Amide I-Band for Conformational Studies. Biopolymers 1988, 27, 1749–1760. [Google Scholar] [CrossRef]
- Jackson, M.; Choo, L.P.; Watson, P.H.; Halliday, W.C.; Mantsch, H.H. Beware of Connective-Tissue Proteins—Assignment and Implications of Collagen Absorptions in Infrared-Spectra of Human Tissues. BBA-Mol. Basis Dis. 1995, 1270, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rabotyagova, E.S.; Cebe, P.; Kaplan, D.L. Collagen structural hierarchy and susceptibility to degradation by ultraviolet radiation. Mater. Sci. Eng. C 2008, 28, 1420–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Koprivova, B.; Lisnenko, M.; Solarska-Sciuk, K.; Prochazkova, R.; Novotny, V.; Mullerova, J.; Mikes, P.; Jencova, V. Large-scale electrospinning of poly (vinylalcohol) nanofibers incorporated with platelet-derived growth factors. Express Polym. Lett. 2020, 14, 987–1000. [Google Scholar] [CrossRef]
- Serpooshan, V.; Zhao, M.M.; Metzler, S.A.; Wei, K.; Shah, P.B.; Wang, A.; Mahmoudi, M.; Malkovskiy, A.V.; Rajadas, J.; Butte, M.J.; et al. The effect of bioengineered acellular collagen patch on cardiac remodelling and ventricular function post myocardial infarction. Biomaterials 2013, 34, 9048–9055. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.K.; Umino, T.; Liu, X.D.; Wang, H.J.; Romberger, D.J.; Spurzem, J.R.; Rennard, S.I. Contraction of fibroblast-containing collagen gels: Initial collagen concentration regulates the degree of contraction and cell survival. Vitr. Cell. Dev.-Anim. 2001, 37, 10–16. [Google Scholar] [CrossRef]
- Nashchekina, Y.A.; Yudintceva, N.M.; Nikonov, P.O.; Ivanova, E.A.; Smagina, L.V.; Voronkina, I.V. Effect of Concentration of Collagen Gel on Functional Activity of Bone Marrow Mesenchymal Stromal Cells. Bull. Exp. Biol. Med. 2017, 163, 123–128. [Google Scholar] [CrossRef]
- Bacakova, M.; Pajorova, J.; Broz, A.; Hadraba, D.; Lopot, F.; Zavadakova, A.; Vistejnova, L.; Beno, M.; Kostic, I.; Jencova, V.; et al. A two-layer skin construct consisting of a collagen hydrogel reinforced by a fibrin-coated polylactide nanofibrous membrane. Int. J. Nanomed. 2019, 14, 5033–5050. [Google Scholar] [CrossRef] [Green Version]
- Velez, D.O.; Tsui, B.; Goshia, T.; Chute, C.L.; Han, A.; Carter, H.; Fraley, S.I. 3D collagen architecture induces a conserved migratory and transcriptional response linked to vasculogenic mimicry. Nat. Commun. 2017, 8, 1651. [Google Scholar] [CrossRef] [Green Version]
- Valero, C.; Amaveda, H.; Mora, M.; Garcia-Aznar, J.M. Combined experimental and computational characterisation of crosslinked collagen-based hydrogels. PLoS ONE 2018, 13, e0195820. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Xie, A.W.; Emani, S.; Yamato, M.; Okano, T.; Emani, S.M.; Wong, J.Y. A Comparison of Human Smooth Muscle and Mesenchymal Stem Cells as Potential Cell Sources for Tissue-Engineered Vascular Patches. Tissue Eng. Part A 2012, 18, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.S.; Moon, H.J.; Lee, M.J.; Song, H.Y.; Kim, Y.M.; Bae, Y.C.; Jung, J.S.; Kim, J.H. Sphingosylphosphorylcholine induces differentiation of human mesenchymal stem cells into smooth-muscle-like cells through a TGF-beta-dependent mechanism. J. Cell Sci. 2006, 119, 4994–5005. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xie, X.Y.; Lian, W.S.; Shi, R.F.; Han, S.L.; Zhang, H.J.; Lu, L.G.; Li, M.Q. Exosomes from adipose-derived stem cells overexpressing Nrf2 accelerate cutaneous wound healing by promoting vascularisation in a diabetic foot ulcer rat model. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-beta and the TGF-beta Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goumans, M.J.; Valdimarsdottir, G.; Itoh, S.; Rosendahl, A.; Sideras, P.; ten Dijke, P. Balancing the activation state of the endothelium via two distinct TGF-beta type I receptors. EMBO J. 2002, 21, 1743–1753. [Google Scholar] [CrossRef]
- Peshavariya, H.M.; Chan, E.C.; Liu, G.S.; Jiang, F.; Dusting, G.J. Transforming growth factor-beta 1 requires NADPH oxidase 4 for angiogenesis in vitro and in vivo. J. Cell. Mol. Med. 2014, 18, 1172–1183. [Google Scholar] [CrossRef]
- Suzuki, Y.; Montagne, K.; Nishihara, A.; Watabe, T.; Miyazono, K. BMPs promote proliferation and migration of endothelial cells via stimulation of VEGF-A/VEGFR2 and Angiopoietin-1/Tie2 signalling. J. Biochem. 2008, 143, 199–206. [Google Scholar] [CrossRef]
- Li, Q.; Kou, X.T.; Qin, X.L.; Li, Z.S.; Li, J.Y.; Chen, C. BMP-4 impedes endothelial cell migration in neointimal hyperplasia via FoXO-3 specific modulation of reactive oxygen species. Atherosclerosis 2022, 351, 9–17. [Google Scholar] [CrossRef]
- Tiaka, E.K.; Papanas, N.; Manolakis, A.C.; Georgiadis, G.S. Epidermal growth factor in the treatment of diabetic foot ulcers: An update. Perspect. Vasc. Surg. Endovasc. Ther. 2012, 24, 37–44. [Google Scholar] [CrossRef]
- Lu, Q.B.; Wan, M.Y.; Wang, P.Y.; Zhang, C.X.; Xu, D.Y.; Liao, X.; Sun, H.J. Chicoric acid prevents PDGF-BB-induced VSMC dedifferentiation, proliferation and migration by suppressing ROS/NFkappaB/mTOR/P70S6K signaling cascade. Redox Biol. 2018, 14, 656–668. [Google Scholar] [CrossRef]
- Gianni-Barrera, R.; Butschkau, A.; Uccelli, A.; Certelli, A.; Valente, P.; Bartolomeo, M.; Groppa, E.; Burger, M.G.; Hlushchuk, R.; Heberer, M.; et al. PDGF-BB regulates splitting angiogenesis in skeletal muscle by limiting VEGF-induced endothelial proliferation. Angiogenesis 2018, 21, 883–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filova, E.; Blanquer, A.; Knitlova, J.; Plencner, M.; Jencova, V.; Koprivova, B.; Lisnenko, M.; Kostakova, E.K.; Prochazkova, R.; Bacakova, L. The Effect of the Controlled Release of Platelet Lysate from PVA Nanomats on Keratinocytes, Endothelial Cells and Fibroblasts. Nanomaterials 2021, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Wu, X.Y.; Tong, J.B.; Yang, X.X.; Zhao, J.L.; Zheng, Q.F.; Zhao, G.B.; Ma, Z.J. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res. Ther. 2015, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Cowper, M.; Frazier, T.; Wu, X.; Curley, J.L.; Ma, M.H.; Mohiuddin, O.A.; Dietrich, M.; McCarthy, M.; Bukowska, J.; Gimble, J.M. Human Platelet Lysate as a Functional Substitute for Fetal Bovine Serum in the Culture of Human Adipose Derived Stromal/Stem Cells. Cells 2019, 8, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camasao, D.B.; Pezzoli, D.; Loy, C.; Kumra, H.; Levesque, L.; Reinhardt, D.P.; Candiani, G.; Mantovani, D. Increasing Cell Seeding Density Improves Elastin Expression and Mechanical Properties in Collagen Gel-Based Scaffolds Cellularised with Smooth Muscle Cells. Biotechnol. J. 2019, 14, e1700768. [Google Scholar] [CrossRef]
- Yamashiro, Y.; Thang, B.Q.; Shin, S.J.; Lino, C.A.; Nakamura, T.; Kim, J.; Sugiyama, K.; Tokunaga, C.; Sakamoto, H.; Osaka, M.; et al. Role of Thrombospondin-1 in Mechanotransduction and Development of Thoracic Aortic Aneurysm in Mouse and Humans. Circ. Res. 2018, 123, 660–672. [Google Scholar] [CrossRef]
- Nikoloudaki, G.; Snider, P.; Simmons, O.; Conway, S.J.; Hamilton, D.W. Periostin and matrix stiffness combine to regulate myofibroblast differentiation and fibronectin synthesis during palatal healing. Matrix Biol. 2020, 94, 31–56. [Google Scholar] [CrossRef] [PubMed]
- Shapland, C.; Hsuan, J.J.; Totty, N.F.; Lawson, D. Purification and Properties of Transgelin—A Transformation and Shape Change Sensitive Actin-Gelling Protein. J. Cell Biol. 1993, 121, 1065–1073. [Google Scholar] [CrossRef]
- Sanz-Fraile, H.; Amoros, S.; Mendizabal, I.; Galvez-Monton, C.; Prat-Vidal, C.; Bayes-Genis, A.; Navajas, D.; Farre, R.; Otero, J. Silk-Reinforced Collagen Hydrogels with Raised Multiscale Stiffness for Mesenchymal Cells 3D Culture. Tissue Eng. Part A 2020, 26, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Filova, E.; Rampichova, M.; Litvinec, A.; Drzik, M.; Mickova, A.; Buzgo, M.; Kost'akova, E.; Martinova, L.; Usvald, D.; Prosecka, E.; et al. A cell-free nanofiber composite scaffold regenerated osteochondral defects in miniature pigs. Int. J. Pharm. 2013, 447, 139–149. [Google Scholar] [CrossRef]
- de Jonge, P.; Simaioforidis, V.; Geutjes, P.; Oosterwijk, E.; Feitz, W. Ureteral reconstruction with reinforced collagen scaffolds in a porcine model. J. Tissue Eng. Regen. Med. 2016, 12, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Syedain, Z.H.; Tranquillo, R.T. TGF-beta 1 diminishes collagen production during long-term cyclic stretching of engineered connective tissue: Implication of decreased ERK signaling. J. Biomech. 2011, 44, 848–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bono, N.; Meghezi, S.; Soncini, M.; Piola, M.; Mantovani, D.; Fiore, G.B. A Dual-Mode Bioreactor System for Tissue Engineered Vascular Models. Ann. Biomed. Eng. 2017, 45, 1496–1510. [Google Scholar] [CrossRef] [PubMed]
- Suchy, T.; Supova, M.; Klapkova, E.; Adamkova, V.; Zavora, J.; Zaloudkova, M.; Ryglova, S.; Ballay, R.; Denk, F.; Pokorny, M.; et al. The release kinetics, antimicrobial activity and cytocompatibility of differently prepared collagen/hydroxyapatite/vancomycin layers: Microstructure vs. nanostructure. Eur. J. Pharm. Sci. 2017, 100, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Ryglova, S.; Suchy, T. Determination of glycosaminoglycans in biological matrices using a simple and sensitive reversed-phase HPLC method with fluorescent detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2021, 1173, 122626. [Google Scholar] [CrossRef] [PubMed]
- Stepanovska, J.; Otahal, M.; Hanzalek, K.; Supova, M.; Matejka, R. pH Modification of High-Concentrated Collagen Bioinks as a Factor Affecting Cell Viability, Mechanical Properties, and Printability. Gels 2021, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Estes, B.T.; Diekman, B.O.; Gimble, J.M.; Guilak, F. Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat. Protoc. 2010, 5, 1294–1311. [Google Scholar] [CrossRef] [Green Version]
- Travnickova, M.; Pajorova, J.; Zarubova, J.; Krocilova, N.; Molitor, M.; Bacakova, L. The Influence of Negative Pressure and of the Harvesting Site on the Characteristics of Human Adipose Tissue-Derived Stromal Cells from Lipoaspirates. Stem Cells Int. 2020, 2020, 1016231. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Number of AA Residues/1000 Units | Amino Acid | Number of AA Residues/1000 Units |
|---|---|---|---|
| Asp + Asn | 48 | Cys | 2 |
| Glu + Gln | 76 | Met | 6 |
| Thr | 17 | Tyr | 3 |
| Ser | 32 | Phe | 15 |
| Gly | 319 | Lys | 31 |
| Ala | 111 | His | 15 |
| Val | 31 | Arg | 52 |
| Ile | 11 | Pro | 121 |
| Leu | 27 | Hyp | 81 |
| Protein (wt%) | 58.45 | ||
| Hyp (wt%) | 5.61 (5.28–5.82) * | ||
| Degree of hydroxylation (%) | 40 | ||
| GAGs (wt%) | 3.92 (3.52–4.81) * | ||
| Lipids (wt%) | 25.49 (22.98–32.82) * | ||
| Water (%) | 7.23 (7.05–7.35) * | ||
| Differentiation | DMEM_Part (a) | PVA_Part (b) | PVA_PL_Part (c) | TGF + BMP_Part (d) |
|---|---|---|---|---|
| Alpha-adducin (ADD1) | 3.37 n = 2 | 2.62 (d) n = 1.25 | 1.89 n.s. D0 n = 1 | 1.66 n.s. D0 n = 1.5 |
| Alpha-parvin (PARVA) | 3.39 n = 3.75 | 3.82 n = 3 | 3.29 n = 3.25 | 2.88 n = 3.5 |
| Caldesmon 1 (CALD1) | 1.77 n = 8.25 | 1.05 n.s. D0 n = 6 | 1.04 n.s. D0 (b), n = 6.75 | 2.62 n.s. D0 (b), n = 5.75 |
| Calponin-2 (CNN2) | 3.11 n = 2 | 3.47 n = 2 | 3.25 n = 1.75 | 3.75 n = 2 |
| Cofilin-2 (CFL2) | 1.99 n = 2.75 | 1.86 n = 2.75 | 1.60 n = 2.75 | 1.62 n = 2.75 |
| Endothelial differentiation-related factor 1 (EDF1) | 2.13 n.s. D0 n = 1.5 | 2.66 n = 1.5 | 2.71 n = 1.75 | 2.97 n = 2 |
| Endothelial monocyte-activating polypeptide 2 (AIMP1) | 3.72 n = 1 | 3.62 n = 0 | 3.55 n = 0.75 | 3.57 n = 1 |
| F-actin-capping protein subunit alpha-1 (CAPZA1) | 3.51 n = 7 | 3.24 n = 6.25 | 3.36 n = 6.5 | 3.14 n = 7 |
| Fascin (FSCN1) | 4.51 n = 3.75 | 4.82 (d) n = 3.75 | 4.40 n = 4 | 3.81 n = 4 |
| Filamin-A (FLNA) | 6.37 n = 39 | 6.16 n = 36.25 | 5.89 n = 36 | 6.43 n = 39.75 |
| Filamin-C (FLNC) | 4.91 n = 25 | 4.91 n = 23 | 5.01 n = 25 | 5.60 n = 27.5 |
| LIM and SH3 domain protein 1 (LASP1) | 3.70 n = 3.5 | 3.18 n = 3.5 | 3.11 n.s. D0 n = 3.75 | 3.13 n = 4 |
| Myosin light polypeptide 6 (MYL6) | 3.50 n = 3.75 | 3.62 n = 4.25 | 3.26 n = 4 | 3.62 n = 4.5 |
| Myosin phosphatase Rho-interacting protein (MPRIP) | 1.43 n.s. D0 n = 0.5 | 1.13 n.s. D0 n = 0 | 1.76 n.s. D0 n = 0.25 | 2.72 (b, c) n = 1.5 |
| Nexilin (NEXN) | 1.98 n.s. D0 n = 1 | 1.47 (d) n = 1 | 1.41 n.s. D0 n = 0.75 | 1.38 n.s. D0 n = 1 |
| Platelet endothelial cell adhesion molecule (PECAM1) | 1.85 n.s. D0 n = 0.75 | 1.79 n.s. D0 n = 1 | 1.29 n.s. D0 n = 0.75 | 1.33 n = 1 |
| Plectin (PLEC) | 4.95 n = 48.5 | 4.92 n = 46.25 | 4.96 n = 45.75 | 4.95 n = 45.25 |
| Transgelin (TAGLN) | 1.59 n = 3.75 | 1.35 n.s. D0 n = 3 | 1.38 n.s. D0 n = 4 | 3.35 (b, c) n = 4 |
| Tropomodulin-3 (TMOD3) | 4.18 n = 4 | 3.89 n = 2.25 | 3.92 n = 2.75 | 4.23 n = 3.25 |
| Tropomyosin alpha-1 chain (TPM1) | 1.11 n.s. D0 n = 4.75 | 1.23 n.s. D0 n = 5 | 1.62 n.s. D0 n = 4.5 | 2.05 (a, b, c) n = 6.75 |
| Tropomyosin alpha-3 chain (TPM3) | 4.01 n = 4.5 | 4.14 n = 5.75 | 3.85 n = 5.5 | 3.61 n = 6.5 |
| Tropomyosin alpha-4 chain (TPM4) | 1.99 n.s. D0 n = 7.5 | 1.48 n.s. D0 n = 7.5 | 1.39 n.s. D0 n = 8 | 1.16 n = 8.5 |
| Utrophin (UTRN) | 2.075 n = 3.75 | 2.70 n = 3 | 1.95 n = 3.25 | 1.62 n = 3 |
| Vinculin (VCL) | 2.39 n = 20.75 | 2.39 n = 20.5 | 2.31 n = 21.25 | 2.27 n = 19.25 |
| von Willebrand factor (VWF) | 2.84 n = 8 | 3.46 n = 9 | 3.56 n = 8 | 3.31 n = 10 |
| Extracellular Matrix | DMEM_Part (a) | PVA_Part (b) | PVA_PL_Part (c) | TGF + BMP_Part (d) |
|---|---|---|---|---|
| Basement membrane-specific heparan sulfate proteoglycan core protein (HSPG2) | 6.17 n = 13 | 6.02 n = 13.5 | 6.08 n = 13 | 5.49 n = 10.75 |
| Collagen alpha-1(V) chain (COL5A1) | 0.65 n.s. D0 n = 2.75 | 0.85 n = 3.5 | 0.8 n = 3.5 | 1.10 n.s. D0 (b, c) n = 4 |
| Collagen alpha-1(XII) chain (COL12A1) | 0.16 n = 0.75 | 0.16 n = 0.5 | 0.15 n = 0.75 | 0.63 n.s. D0 (a, b, c) n = 4.5 |
| Collagen alpha-1(VII) chain (COL7A1) | 0.65 n.s. D0 n = 0 | 0.92 n.s. D0 n = 0 | 0.7 n.s. D0 n = 0 | 3.74 (a, b, c) n = 1.75 |
| Collagen triple helix repeat-containing protein 1 (CTHRC1) | 1.36 n.s. D0 n = 0.75 | 1.34 n = 1 | 1.19 n = 1 | 2.13 (c) n = 1 |
| Decorin (DCN) | 4.47 n = 3.5 | 3.26 n = 3.25 | 3.66 n = 2.75 | 3.66 n = 1 |
| Extracellular matrix protein 1 (ECM1) | 2.81 n = 3.25 | 3.86 (d) n = 4 | 3.05 (d) n = 3.25 | 1.39 n.s. D0 n = 2.5 |
| Fibrillin-1 (FBN1) | 1.99 n.s. D0 n = 0.75 | 1.99 n.s. D0 n = 0.5 | n.s. D0 n = 1 | 1.65 n = 0.75 |
| Fibronectin (FN1) | 6.09 n = 29.25 | 5.80 n = 27 | 5.77 n = 29 | 7.20 n = 36 |
| Fibulin-1 (FBLN1) | 2.68 n = 2.25 | 2.22 n = 2 | 2.32 n = 2 | 2.28 n = 1.75 |
| Galectin-3-binding protein (LGALS3BP) | 2.61 n = 2 | 2.60 (d) n = 2 | 2.99 (d) n = 2 | 1.54 n.s. D0 n = 1.75 |
| Laminin subunit beta-1 (LAMB1) | 5.05 n = 9.25 | 5.26 n = 10 | 5.29 n = 9.75 | 4.89 n = 9 |
| Laminin subunit gamma-1 (LAMC1) | 4.84 n = 11 | 4.85 n = 11 | 4.97 n = 11.25 | 4.43 n = 9.25 |
| Laminin subunit alpha-2 (LAMA2) | 1.47 n = 1.25 | 1.19 n.s. D0 n = 0.75 | 1.11 n.s. D0 n = 0.75 | 1.31 n = 1 |
| Laminin subunit alpha-4 (LAMA4) | 4.68 n = 7 | 4.70 (d) n = 5.25 | 4.77 (d) n = 6.5 | 2.53 n = 4 |
| Lumican (LUM) | 2.23 n = 1.5 | 2.36 n = 2 | 2.41 n = 2 | 2.59 n = 1 |
| Nidogen-1 (NID1) | 2.68 n = 3 | 2.52 (d) n = 3.75 | 2.81 (d) n = 3 | 1.24 n.s. D0 n = 2.5 |
| Tenascin (TNC) | 0.54 n.s. D0 n = 1.75 | 0.77 n.s. D0 n = 1.25 | 0.52 n.s. D0 n = 1.75 | 3.28 (b, c) n = 0 |
| Thrombospondin-4 (THBS4) | 3.45 n = 1.25 | 4.36 (d) n = 1.25 | 4.12 (d) n = 1.25 | 2.65 n = 1 |
| REMODELLING | DMEM (a) | PVA (b) | PVA_PL (c) | TGF + BMP (d) |
|---|---|---|---|---|
| Collagenase, type IV (MMP2) | 6.27 n = 3.5 | 6.27 n = 3.75 | 6.42 n = 3.75 | 6.30 n = 3.25 |
| Disintegrin and metallo-proteinase with thrombo-spondin motifs 1 (ADAMTS1) | 3.46 n = 1 | 3.46 n = 1 | 3.93 n = 1 | 2.86 n = 1 |
| Interstitial collagenase, MMP1 | 3.88 n = 4.25 | 5.04 (d) n = 6.5 | 4.97 (d) n = 5.75 | 3.06 n = 3 |
| Lysyl oxidase homolog 2 (LOXL2) | 1.70 n.s. D0 n = 0.75 | 1.39 n.s. D0 n = 1 | 1.25 n.s. D0 n = 1 | 1.50 n = 1 |
| Matrix metalloproteinase-14 (MMP14) | 1.48 n.s. D0 n = 0.75 | 1.26 n.s. D0 n = 0.75 | 0.6 n = 1 | 0.6 n = 6.25 |
| Matrix-remodelling-associated protein 7 (MXRA7) | 1.56 n.s. D0 n = 1.25 | 1.95 n.s. D0 n = 1 | 1.79 n.s. D0 n = 1 | 3.48 (b) n = 2 |
| Metalloproteinase inhibitor 1 (TIMP1) | 5.81 n = 2 | 6.77 n = 2 | 6.47 n = 2 | 6.14 n = 2 |
| Periostin (POSTN) | 1.17 n.s. D0 n = 4.25 | 1.18 n.s. D0 n = 4.25 | 1.01 n.s. D0 n = 4 | 1.96 (a, b, c) n = 10.5 |
| Procollagen C-endopeptidase enhancer 1 (PCOLCE) | 3.04 n = 1 | 1.86 n.s. D0 n = 1 | 2.47 n.s. D0 n = 1 | 2.96 n = 1 |
| Procollagen-lysine, 2-oxoglutarate 5-dioxygenase 1 (PLOD1) | 2.37 n.s. D0 n = 2 | 2.68 n = 2 | 2.59 n = 2 | 2.45 n = 2 |
| Procollagen-lysine, 2-oxo-glutarate 5-dioxygenase 2 (PLOD2) | 2.91 n = 5.5 | 3.59 n = 5.75 | 3.19 n = 5.25 | 4.80 n = 7.25 |
| TGF-beta-induced protein ig-h3 (TGFBI) | 1.41 n.s. D0 n = 4 | 1.42 n = 6.25 | 1.32 n = 5 | 2.79 n = 7.25 |
| Transforming growth factor beta-1-induced transcript 1 protein (TGFB1I1) | 1.17 n.s. D0 n = 0.25 | 1.24 n.s. D0 n = 0 | 1.82 n.s. D0 n = 0 | 1.97 (c) n = 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filova, E.; Supova, M.; Eckhardt, A.; Vrbacky, M.; Blanquer, A.; Travnickova, M.; Knitlova, J.; Suchy, T.; Ryglova, S.; Braun, M.; et al. Adipose-Derived Stem Cells in Reinforced Collagen Gel: A Comparison between Two Approaches to Differentiation towards Smooth Muscle Cells. Int. J. Mol. Sci. 2023, 24, 5692. https://doi.org/10.3390/ijms24065692
Filova E, Supova M, Eckhardt A, Vrbacky M, Blanquer A, Travnickova M, Knitlova J, Suchy T, Ryglova S, Braun M, et al. Adipose-Derived Stem Cells in Reinforced Collagen Gel: A Comparison between Two Approaches to Differentiation towards Smooth Muscle Cells. International Journal of Molecular Sciences. 2023; 24(6):5692. https://doi.org/10.3390/ijms24065692
Chicago/Turabian StyleFilova, Elena, Monika Supova, Adam Eckhardt, Marek Vrbacky, Andreu Blanquer, Martina Travnickova, Jarmila Knitlova, Tomas Suchy, Sarka Ryglova, Martin Braun, and et al. 2023. "Adipose-Derived Stem Cells in Reinforced Collagen Gel: A Comparison between Two Approaches to Differentiation towards Smooth Muscle Cells" International Journal of Molecular Sciences 24, no. 6: 5692. https://doi.org/10.3390/ijms24065692
APA StyleFilova, E., Supova, M., Eckhardt, A., Vrbacky, M., Blanquer, A., Travnickova, M., Knitlova, J., Suchy, T., Ryglova, S., Braun, M., Burdikova, Z., Schätz, M., Jencova, V., Lisnenko, M., Behalek, L., Prochazkova, R., Sedlacek, R., Kubasova, K., & Bacakova, L. (2023). Adipose-Derived Stem Cells in Reinforced Collagen Gel: A Comparison between Two Approaches to Differentiation towards Smooth Muscle Cells. International Journal of Molecular Sciences, 24(6), 5692. https://doi.org/10.3390/ijms24065692