
Mesenchymal Stem Cells from COPD Patients Are Capable of Restoring Elastase-Induced Emphysema in a Murine Experimental Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. AD-MSC from COPD Patients Present Different Functional Responses Than Those from Donors in Migration and Cigarette Smoke Exposure
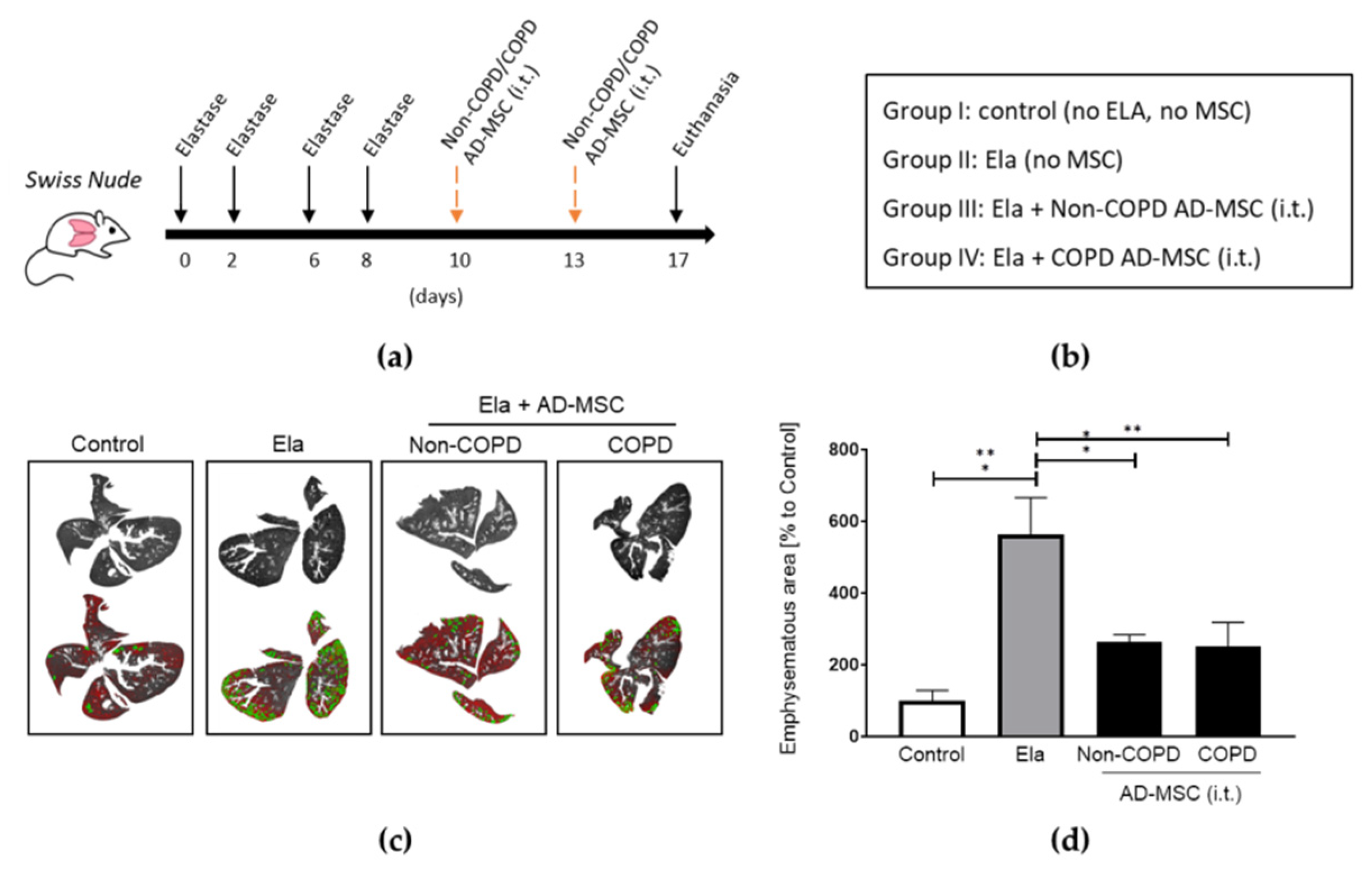
2.2. AD-MSC from COPD Patients Reduced Elastase-Induced Emphysema at the Same Extent as Those from Non-COPD
2.3. Administration of Human UC-MSC Improved Elastase-Induced Emphysema
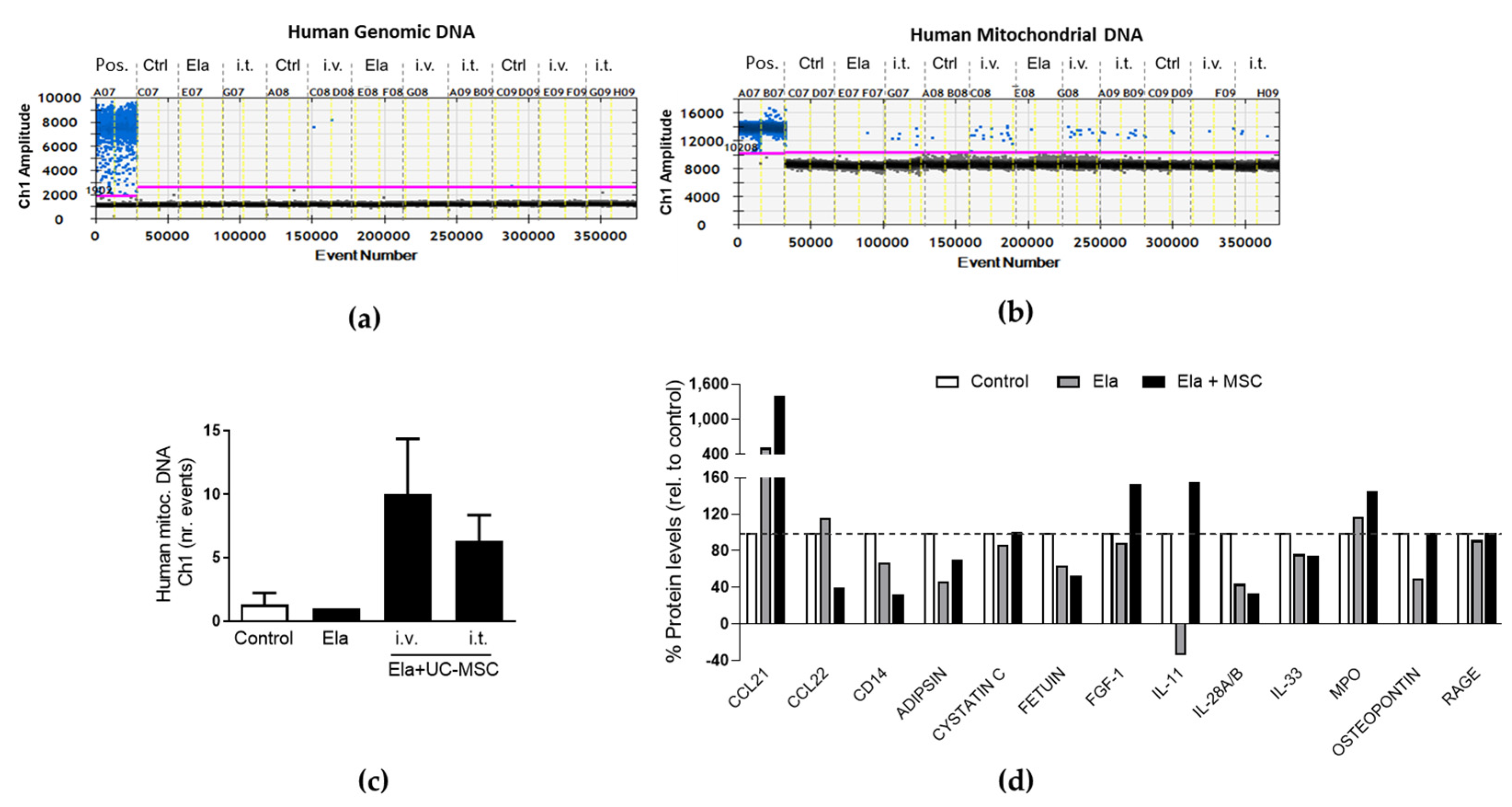
2.4. Human UC-MSC Leave Mitochondrial DNA and Modified the Immunomodulatory Cytokine and Chemokine Profile in the Elastase-Treated Mouse Lungs
3. Discussion
4. Materials and Methods
4.1. Patient Data
4.2. Isolation and Characterization of Human MSC
4.3. MSC Differentiation Assays
4.4. Cellular In Vitro Treatments
4.4.1. Cigarette Smoke Medium (CSM)
4.4.2. VEG-121 Stimulations
4.5. Cellular Migration and Attachment Assays
4.6. Droplet Digital PCR
4.7. MSC Staining and Image Acquisition
4.8. Animal Elastase Emphysema Model
4.9. Protein Array
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agusti, A.; Vogelmeier, C.; Faner, R. COPD 2020: Changes and challenges. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L879–L883. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Timens, W. The pathology of chronic obstructive pulmonary disease. Annu. Rev. Pathol. 2009, 4, 435–459. [Google Scholar] [CrossRef]
- Halpin, D.M.G.; Criner, G.J.; Papi, A.; Singh, D.; Anzueto, A.; Martinez, F.J.; Agusti, A.A.; Vogelmeier, C.F. Global Initiative for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease. The 2020 GOLD Science Committee Report on COVID-19 and Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2021, 203, 24–36. [Google Scholar] [CrossRef]
- Wang, Q.; Sundar, I.K.; Lucas, J.H.; Muthumalage, T.; Rahman, I. Molecular clock REV-ERBalpha regulates cigarette smoke-induced pulmonary inflammation and epithelial-mesenchymal transition. JCI Insight 2021, 6, e145200. [Google Scholar] [CrossRef]
- Behnke, J.; Kremer, S.; Shahzad, T.; Chao, C.M.; Bottcher-Friebertshauser, E.; Morty, R.E.; Bellusci, S.; Ehrhardt, H. MSC Based Therapies-New Perspectives for the Injured Lung. J. Clin. Med. 2020, 9, 682. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, S.; Duan, Y.; Nie, H.; Cui, X.; Du, J.; Yao, Y. Mesenchymal stem cells: Biological characteristics and application in disease therapy. Biochimie 2021, 185, 9–21. [Google Scholar] [CrossRef]
- Armitage, J.D.; Tan, D.B.A.; Sturm, M.; Moodley, Y.P. Transcriptional profiling of circulating mononuclear cells from patients with chronic obstructive pulmonary disease receiving mesenchymal stromal cell infusions. Stem. Cells Transl. Med. 2021, 10, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Casaburi, R.; Flannery, R.; LeRoux-Williams, M.; Tashkin, D.P. A placebo-controlled, randomized trial of mesenchymal stem cells in COPD. Chest 2013, 143, 1590–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Li, F.; Zhou, X.; Chung, K.F.; Wang, W.; Wang, J. Stem cell therapies for chronic obstructive pulmonary disease: Current status of pre-clinical studies and clinical trials. J. Thorac. Dis. 2018, 10, 1084–1098. [Google Scholar] [CrossRef] [Green Version]
- Musial-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transpl. 2019, 28, 801–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Yang, Y.; Yang, D.; Luo, F.; Liang, W.; Guo, S.; Xu, J. The immunomodulatory activity of human umbilical cord blood-derived mesenchymal stem cells in vitro. Immunology 2009, 126, 220–232. [Google Scholar] [CrossRef]
- Chen, Y.T.; Miao, K.; Zhou, L.; Xiong, W.N. Stem cell therapy for chronic obstructive pulmonary disease. Chin. Med. J. 2021, 134, 1535–1545. [Google Scholar] [CrossRef]
- Li, T.; Luo, C.; Zhang, J.; Wei, L.; Sun, W.; Xie, Q.; Liu, Y.; Zhao, Y.; Xu, S.; Wang, L. Efficacy and safety of mesenchymal stem cells co-infusion in allogeneic hematopoietic stem cell transplantation: A systematic review and meta-analysis. Stem. Cell Res. Ther. 2021, 12, 246. [Google Scholar] [CrossRef]
- Cardenes, N.; Alvarez, D.; Sellares, J.; Peng, Y.; Corey, C.; Wecht, S.; Nouraie, S.M.; Shanker, S.; Sembrat, J.; Bueno, M.; et al. Senescence of bone marrow-derived mesenchymal stem cells from patients with idiopathic pulmonary fibrosis. Stem. Cell Res. Ther. 2018, 9, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; von See, M.P.; Yu, E.; Gunel, B.; Lu, K.; Vazin, T.; Schaffer, D.V.; Goodwill, P.W.; Conolly, S.M. Quantitative Magnetic Particle Imaging Monitors the Transplantation, Biodistribution, and Clearance of Stem Cells In Vivo. Theranostics 2016, 6, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [Green Version]
- Janczewski, A.M.; Wojtkiewicz, J.; Malinowska, E.; Doboszynska, A. Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD? Int. J. Mol. Sci. 2017, 18, 2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadalipour, A.; Dumbali, S.P.; Wenzel, P.L. Mitochondrial Transfer and Regulators of Mesenchymal Stromal Cell Function and Therapeutic Efficacy. Front Cell Dev. Biol. 2020, 8, 603292. [Google Scholar] [CrossRef]
- Schmidt, A.; Ladage, D.; Schinkothe, T.; Klausmann, U.; Ulrichs, C.; Klinz, F.J.; Brixius, K.; Arnhold, S.; Desai, B.; Mehlhorn, U.; et al. Basic fibroblast growth factor controls migration in human mesenchymal stem cells. Stem Cells 2006, 24, 1750–1758. [Google Scholar] [CrossRef]
- Kanazawa, H.; Asai, K.; Hirata, K.; Yoshikawa, J. Possible effects of vascular endothelial growth factor in the pathogenesis of chronic obstructive pulmonary disease. Am. J. Med. 2003, 114, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, Y.; Tuder, R.M.; Cool, C.D.; Lynch, D.A.; Flores, S.C.; Voelkel, N.F. Endothelial cell death and decreased expression of vascular endothelial growth factor and vascular endothelial growth factor receptor 2 in emphysema. Am. J. Respir Crit Care Med. 2001, 163 Pt 3, 737–744. [Google Scholar] [CrossRef] [Green Version]
- Westergren-Thorsson, G.; Bagher, M.; Andersson-Sjoland, A.; Thiman, L.; Lofdahl, C.G.; Hallgren, O.; Bjermer, L.; Larsson-Callerfelt, A.K. VEGF synthesis is induced by prostacyclin and TGF-beta in distal lung fibroblasts from COPD patients and control subjects: Implications for pulmonary vascular remodelling. Respirology 2018, 23, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, E.A.; Schenck, T.L.; Machens, H.G.; Egana, J.T. Acute stimulation of mesenchymal stem cells with cigarette smoke extract affects their migration, differentiation, and paracrine potential. Sci. Rep. 2016, 6, 22957. [Google Scholar] [CrossRef] [Green Version]
- Antunes, M.A.; Abreu, S.C.; Cruz, F.F.; Teixeira, A.C.; Lopes-Pacheco, M.; Bandeira, E.; Olsen, P.C.; Diaz, B.L.; Takyia, C.M.; Freitas, I.P.; et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir. Res. 2014, 15, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, N.; Kitabatake, M.; Ouji-Sageshima, N.; Ibaraki, T.; Kumamoto, M.; Fujita, Y.; Hontsu, S.; Yamauchi, M.; Yoshikawa, M.; Muro, S.; et al. Human Adipose-Derived Mesenchymal Stem Cells Ameliorate Elastase-Induced Emphysema in Mice by Mesenchymal-Epithelial Transition. Int. J. Chron. Obs. Pulmon Dis. 2021, 16, 2783–2793. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Segal, K.; Casaburi, R.; Hayes, J.; Tashkin, D. Effect of mesenchymal stromal cell infusions on lung function in COPD patients with high CRP levels. Respir. Res. 2021, 22, 142. [Google Scholar] [CrossRef]
- de Oliveira, H.G.; Cruz, F.F.; Antunes, M.A.; de Macedo Neto, A.V.; Oliveira, G.A.; Svartman, F.M.; Borgonovo, T.; Rebelatto, C.L.; Weiss, D.J.; Brofman, P.R.; et al. Combined Bone Marrow-Derived Mesenchymal Stromal Cell Therapy and One-Way Endobronchial Valve Placement in Patients with Pulmonary Emphysema: A Phase I Clinical Trial. Stem. Cells Transl. Med. 2017, 6, 962–969. [Google Scholar] [CrossRef]
- Armitage, J.; Tan, D.B.A.; Troedson, R.; Young, P.; Lam, K.V.; Shaw, K.; Sturm, M.; Weiss, D.J.; Moodley, Y.P. Mesenchymal stromal cell infusion modulates systemic immunological responses in stable COPD patients: A phase I pilot study. Eur. Respir. J. 2018, 51, 1702369. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef]
- Xunian, Z.; Kalluri, R. Biology and therapeutic potential of mesenchymal stem cell-derived exosomes. Cancer Sci. 2020, 111, 3100–3110. [Google Scholar] [CrossRef]
- Katsha, A.M.; Ohkouchi, S.; Xin, H.; Kanehira, M.; Sun, R.; Nukiwa, T.; Saijo, Y. Paracrine factors of multipotent stromal cells ameliorate lung injury in an elastase-induced emphysema model. Mol. Ther. 2011, 19, 196–203. [Google Scholar] [CrossRef]
- Michaeloudes, C.; Li, X.; Mak, J.C.W.; Bhavsar, P.K. Study of Mesenchymal Stem Cell-Mediated Mitochondrial Transfer in In Vitro Models of Oxidant-Mediated Airway Epithelial and Smooth Muscle Cell Injury. Methods Mol. Biol. 2021, 2269, 93–105. [Google Scholar]
- Ahmad, T.; Mukherjee, S.; Pattnaik, B.; Kumar, M.; Singh, S.; Kumar, M.; Rehman, R.; Tiwari, B.K.; Jha, K.A.; Barhanpurkar, A.P.; et al. Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy. EMBO J. 2014, 33, 994–1010. [Google Scholar] [PubMed]
- Velarde, F.; Ezquerra, S.; Delbruyere, X.; Caicedo, A.; Hidalgo, Y.; Khoury, M. Mesenchymal stem cell-mediated transfer of mitochondria: Mechanisms and functional impact. Cell Mol. Life Sci. 2022, 79, 177. [Google Scholar] [CrossRef] [PubMed]
- Liesveld, J.L.; Sharma, N.; Aljitawi, O.S. Stem cell homing: From physiology to therapeutics. Stem. Cells 2020, 38, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef] [Green Version]
- Ghazavi, H.; Hoseini, S.J.; Ebrahimzadeh-Bideskan, A.; Mashkani, B.; Mehri, S.; Ghorbani, A.; Sadri, K.; Mahdipour, E.; Ghasemi, F.; Forouzanfar, F.; et al. Fibroblast Growth Factor Type 1 (FGF1)-Overexpressed Adipose-Derived Mesenchaymal Stem Cells (AD-MSC(FGF1)) Induce Neuroprotection and Functional Recovery in a Rat Stroke Model. Stem Cell Rev. Rep. 2017, 13, 670–685. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, P.; Hong, C.; Jiang, Q.; Xing, Y.; Tang, X.; Jiang, H.; Luo, S.; Chen, X. Serum cytokine profiles in patients with chronic obstructive pulmonary disease associated pulmonary hypertension identified using protein array. Cytokine 2018, 111, 342–349. [Google Scholar] [CrossRef]
- Wang, X.Y. MSCs transplantation may be a potential therapeutic strategy for COVID-19 treatment. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4537–4538. [Google Scholar]
- Wang, M.Y.; Zhou, T.Y.; Zhang, Z.D.; Liu, H.Y.; Zheng, Z.Y.; Xie, H.Q. Current therapeutic strategies for respiratory diseases using mesenchymal stem cells. MedComm 2021, 2, 351–380. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | COPD | Non-COPD | Sig. (p) |
|---|---|---|---|
| N | 12 | 13 | |
| Age | 64.75 ± 6.01 | 58 ± 12.4 | 0.101 |
| Gender (%) m/f | 10 (83.33)/2 (16.66) | 8 (61.53)/5 (38.46) | 0.292 |
| Smoking history | |||
| smoker % | 6 (50) | 7 (53.84) | |
| exsmoker % | 6 (50) | 4 (12.5) | 0.469 |
| nonsmoker % | - | 2 (6.25) | |
| smoking index (p/y) | 92.75 (40–400) | 32.13 (3–65) | 0.152 |
| Spirometry | |||
| FVC (L) | 2.95 ± 0.68 | 3.95 ± 0.71 | 0.0015 * |
| FVC (% predicted) | 80.7 ± 14.25 | 99.33 ± 2.87 | 0.0001 * |
| FEV1 (L) | 1.48 ± 0.36 | 2.89 ± 0.61 | <0.0001 * |
| FEV1 (% predicted) | 55.55 ± 12.49 | 91.67 ± 8.99 | <0.0001 * |
| FEV1/FVC % | 50.43 ± 10.59 | 69.94 ± 7.91 | <0.0001 * |
| GOLD Stage | |||
| GOLD I | 1 (8.33) | - | |
| GOLD II | 8 (66.66) | - | |
| GOLD III | 3 (25.0) | - | |
| GOLD IV | 0 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Río, C.; Jahn, A.K.; Martin-Medina, A.; Calvo Bota, A.M.; De Francisco Casado, M.T.; Pont Antona, P.J.; Gigirey Castro, O.; Carvajal, Á.F.; Villena Portella, C.; Gómez Bellvert, C.; et al. Mesenchymal Stem Cells from COPD Patients Are Capable of Restoring Elastase-Induced Emphysema in a Murine Experimental Model. Int. J. Mol. Sci. 2023, 24, 5813. https://doi.org/10.3390/ijms24065813
Río C, Jahn AK, Martin-Medina A, Calvo Bota AM, De Francisco Casado MT, Pont Antona PJ, Gigirey Castro O, Carvajal ÁF, Villena Portella C, Gómez Bellvert C, et al. Mesenchymal Stem Cells from COPD Patients Are Capable of Restoring Elastase-Induced Emphysema in a Murine Experimental Model. International Journal of Molecular Sciences. 2023; 24(6):5813. https://doi.org/10.3390/ijms24065813
Chicago/Turabian StyleRío, Carlos, Andreas K. Jahn, Aina Martin-Medina, Alba Marina Calvo Bota, Mª Teresa De Francisco Casado, Pere Joan Pont Antona, Orlando Gigirey Castro, Ángel Francisco Carvajal, Cristina Villena Portella, Cristina Gómez Bellvert, and et al. 2023. "Mesenchymal Stem Cells from COPD Patients Are Capable of Restoring Elastase-Induced Emphysema in a Murine Experimental Model" International Journal of Molecular Sciences 24, no. 6: 5813. https://doi.org/10.3390/ijms24065813
APA StyleRío, C., Jahn, A. K., Martin-Medina, A., Calvo Bota, A. M., De Francisco Casado, M. T., Pont Antona, P. J., Gigirey Castro, O., Carvajal, Á. F., Villena Portella, C., Gómez Bellvert, C., Iglesias, A., Calvo Benito, J., Gayà Puig, A., Ortiz, L. A., & Sala-Llinàs, E. (2023). Mesenchymal Stem Cells from COPD Patients Are Capable of Restoring Elastase-Induced Emphysema in a Murine Experimental Model. International Journal of Molecular Sciences, 24(6), 5813. https://doi.org/10.3390/ijms24065813