

Lactate Dehydrogenase Superfamily in Rice and Arabidopsis: Understanding the Molecular Evolution and Structural Diversity

 and
and
Abstract
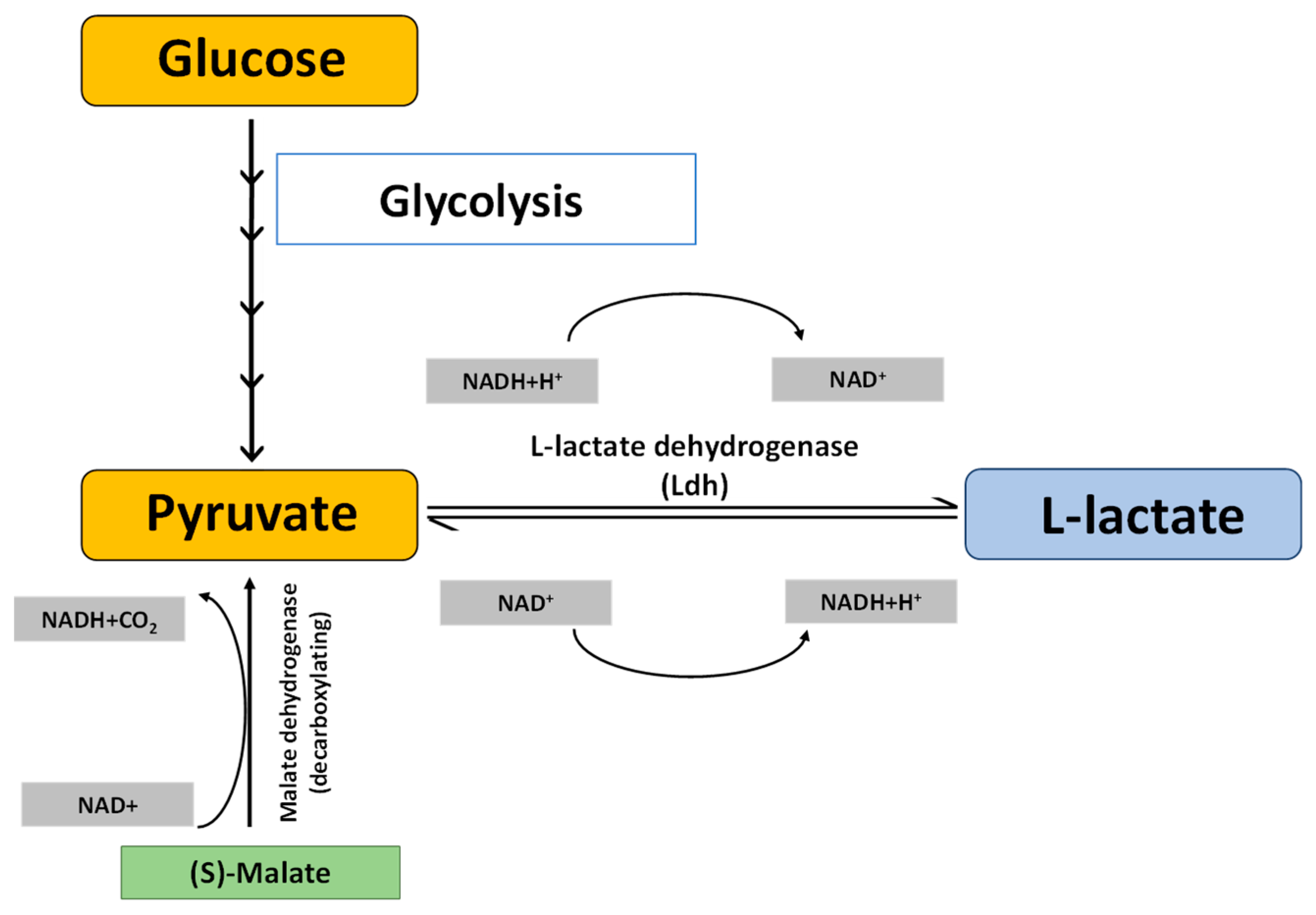
:1. Introduction
2. Results
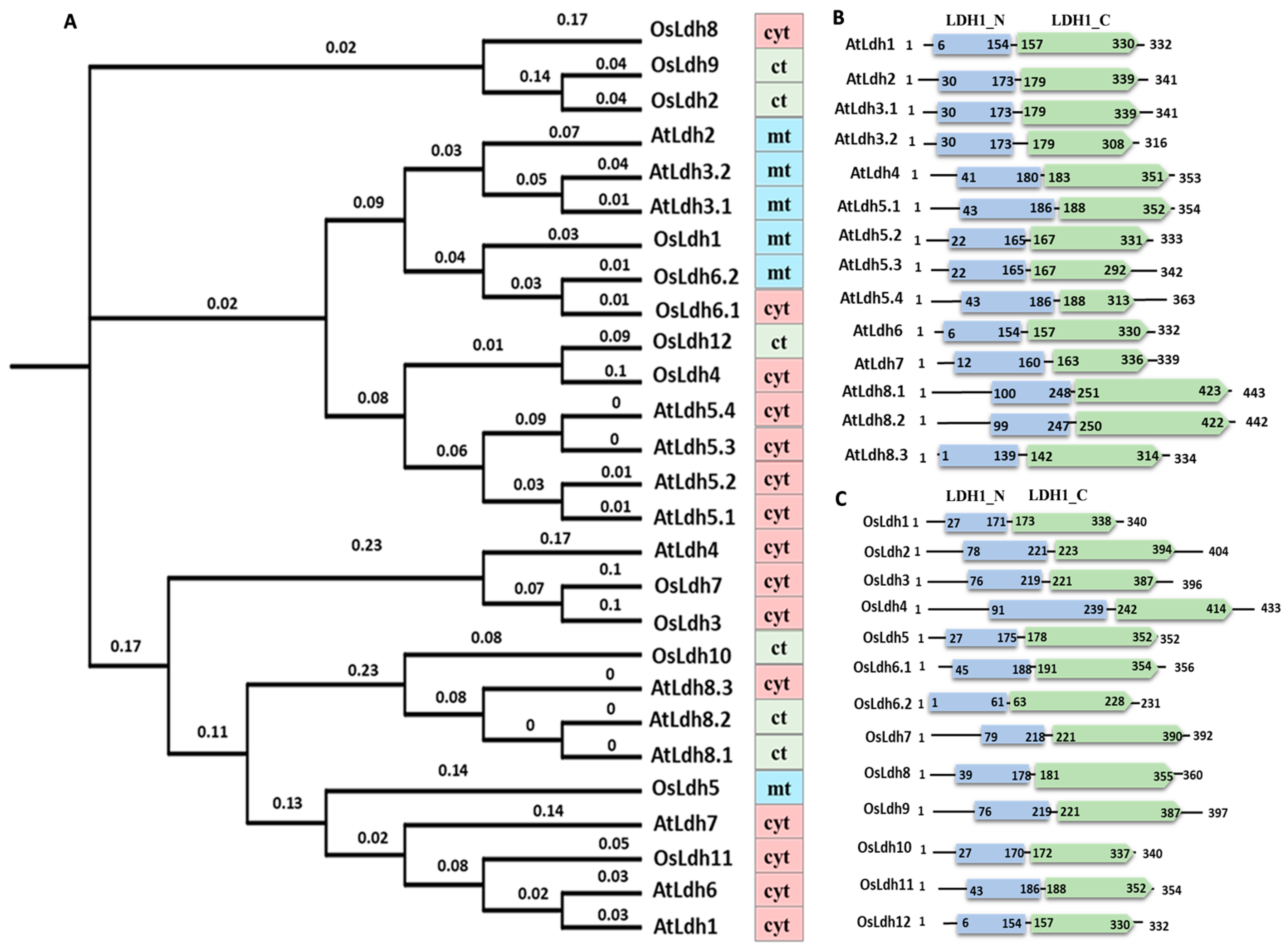
2.1. Identification and Evolutionary Analysis of Ldh Genes across Plant Species
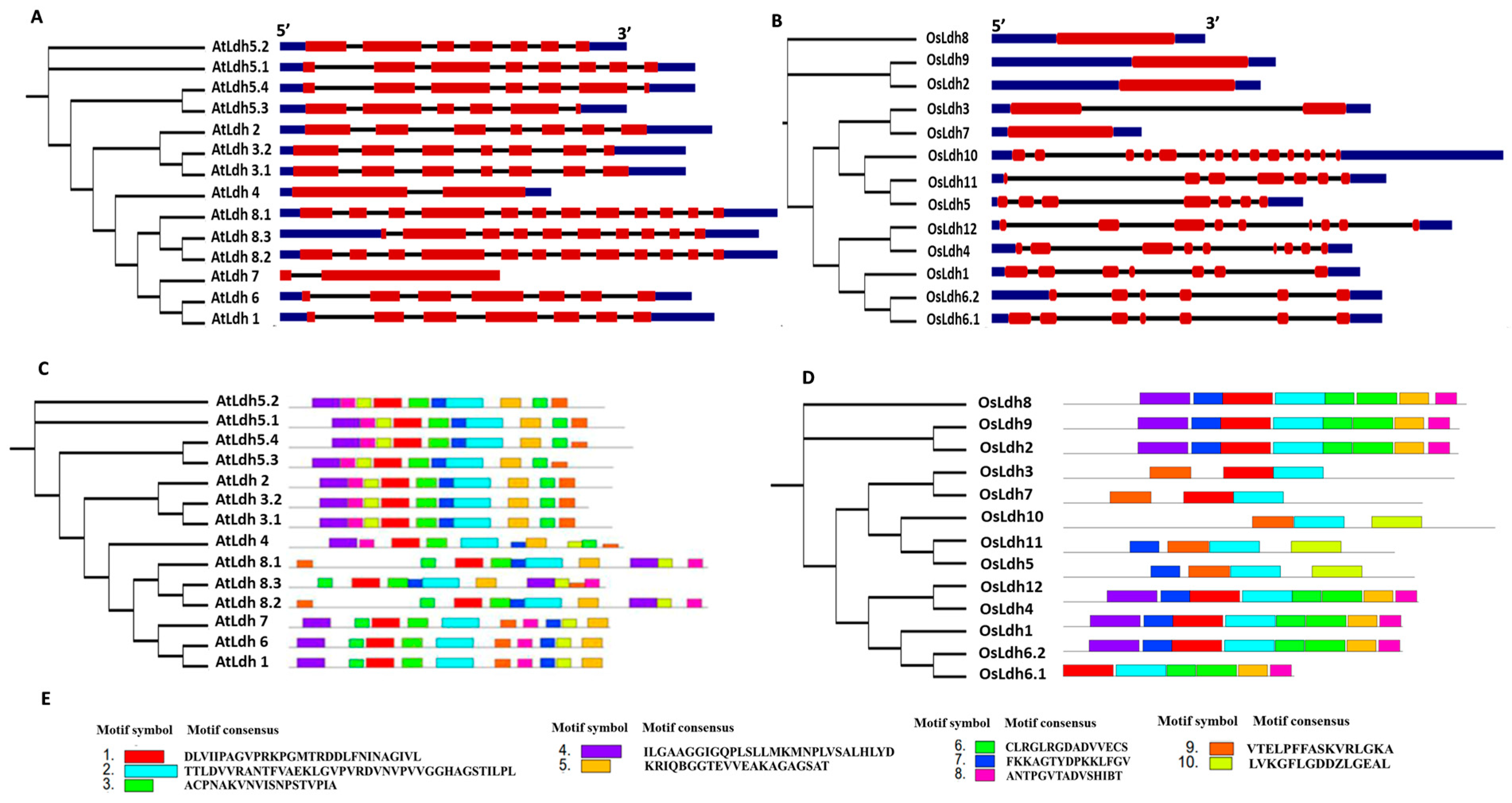
2.2. Exon/Intron Organization and Protein Motif Analysis of the Rice and Arabidopsis Ldh Genes and Proteins
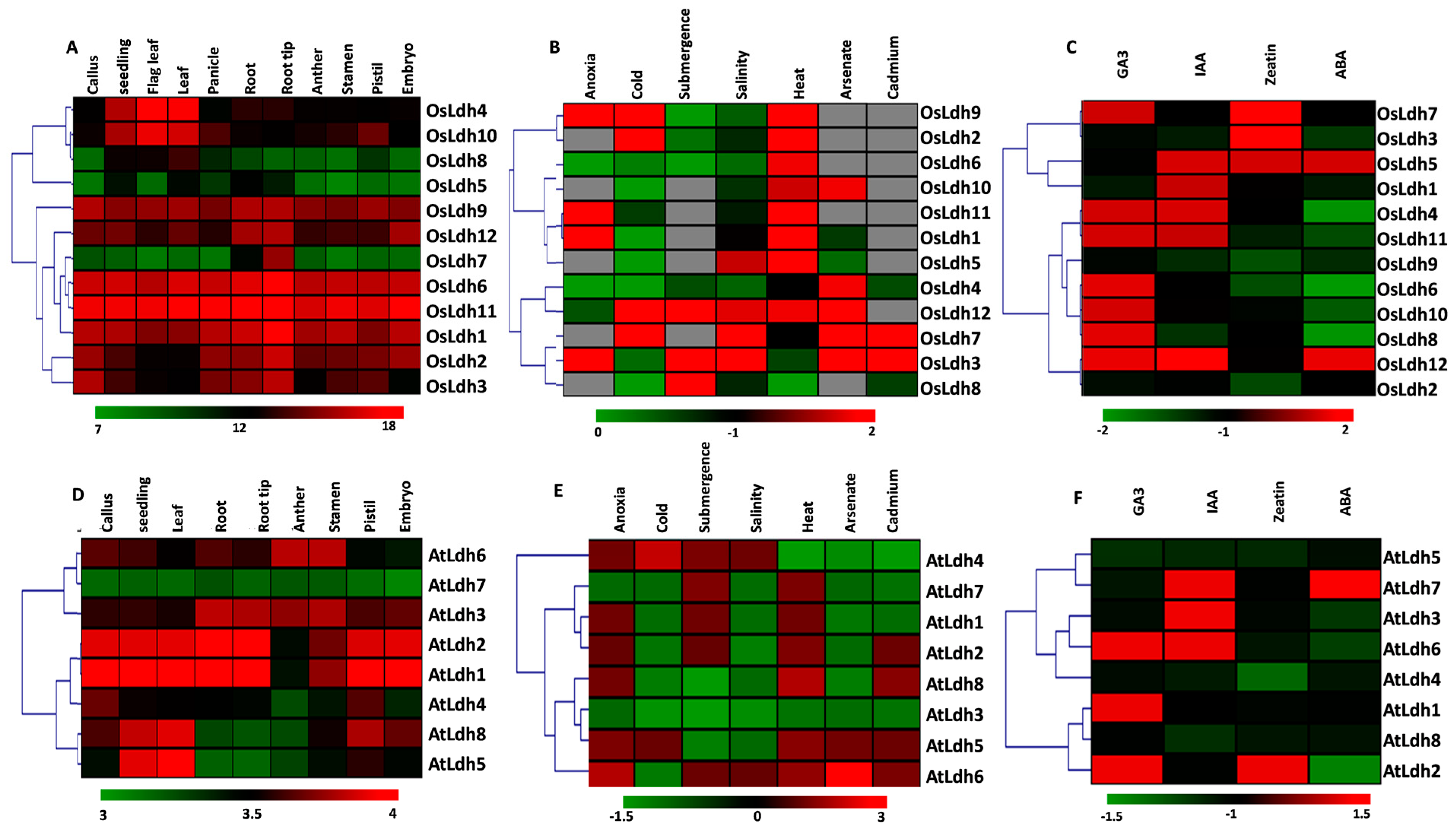
2.3. Tissue-Specific Variations and Stress-Mediated Expression Profiling of Ldh Genes
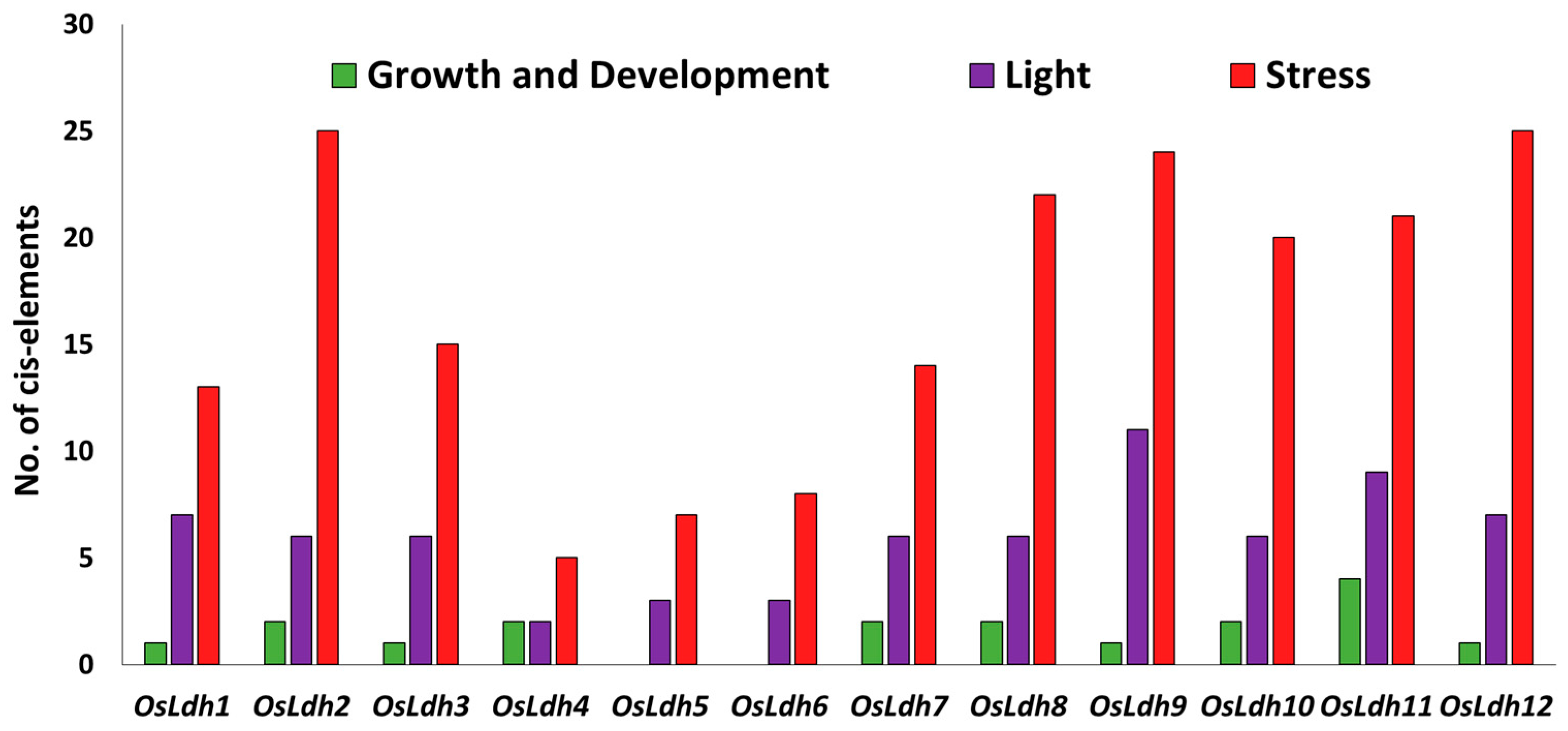
2.4. Identification of cis-Elements in the Promoter Region of Rice Ldh Genes
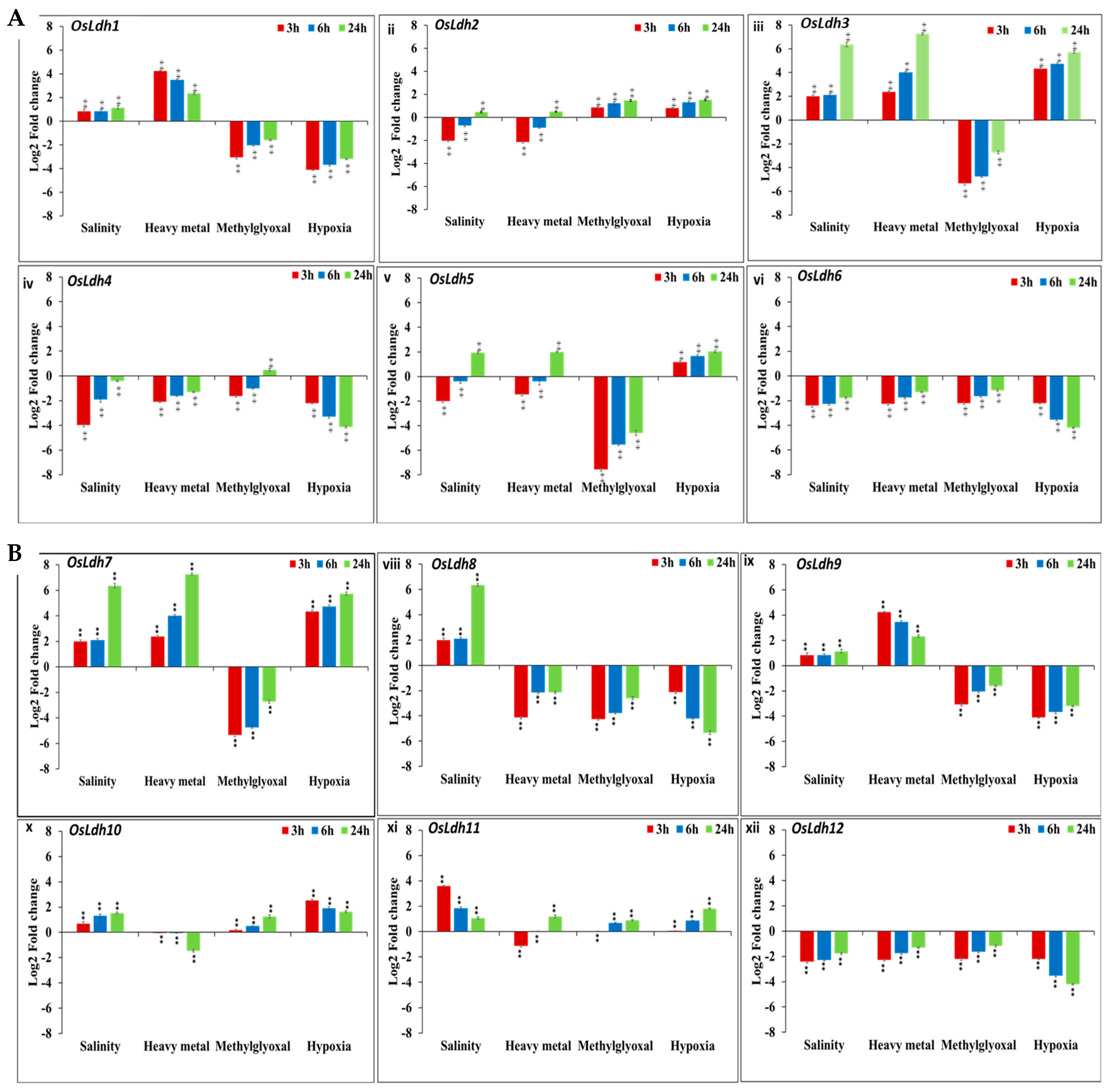
2.5. Temporal Profiling of Rice Ldh Genes under Different Abiotic Stresses Using qRT PCR
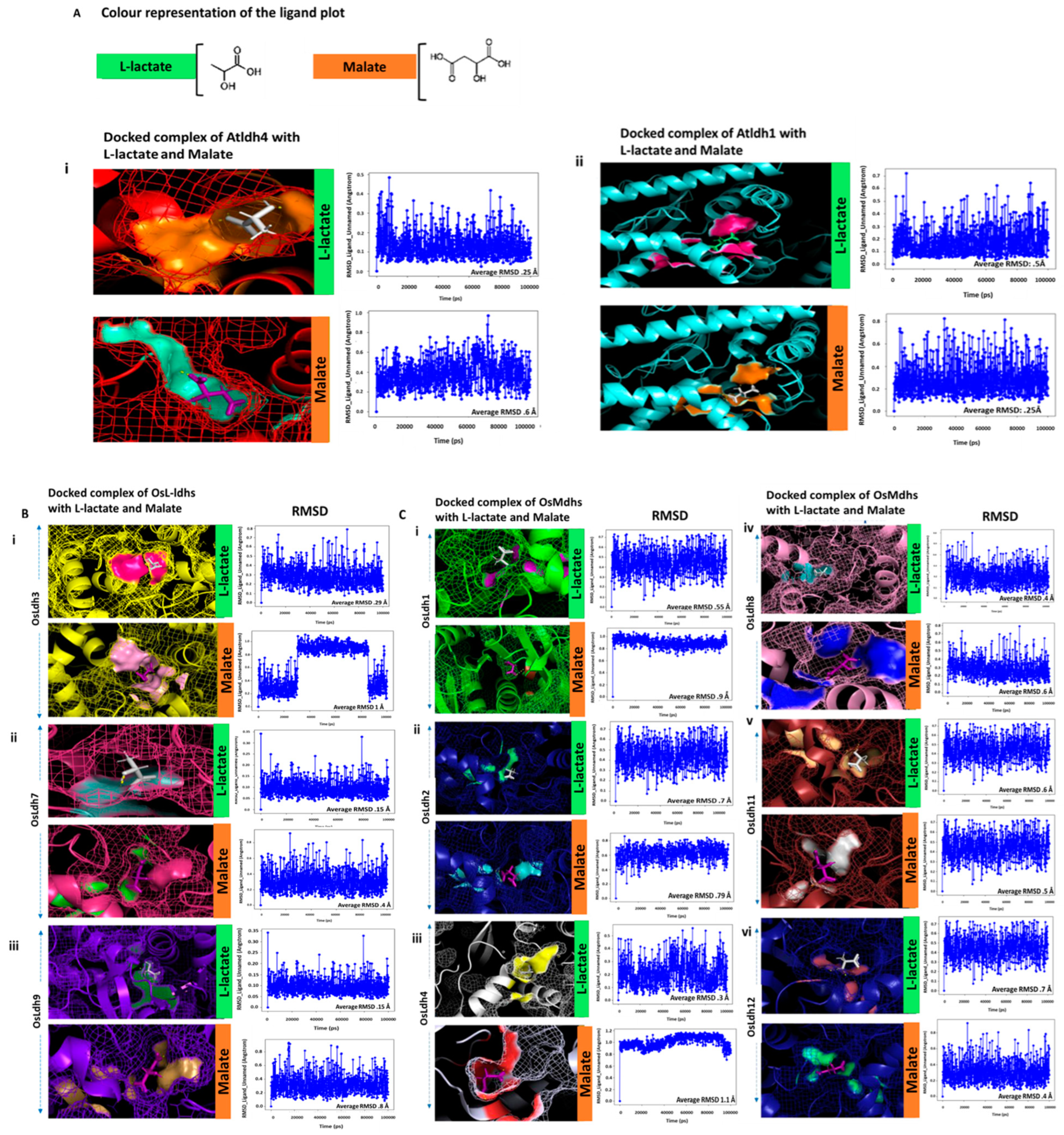
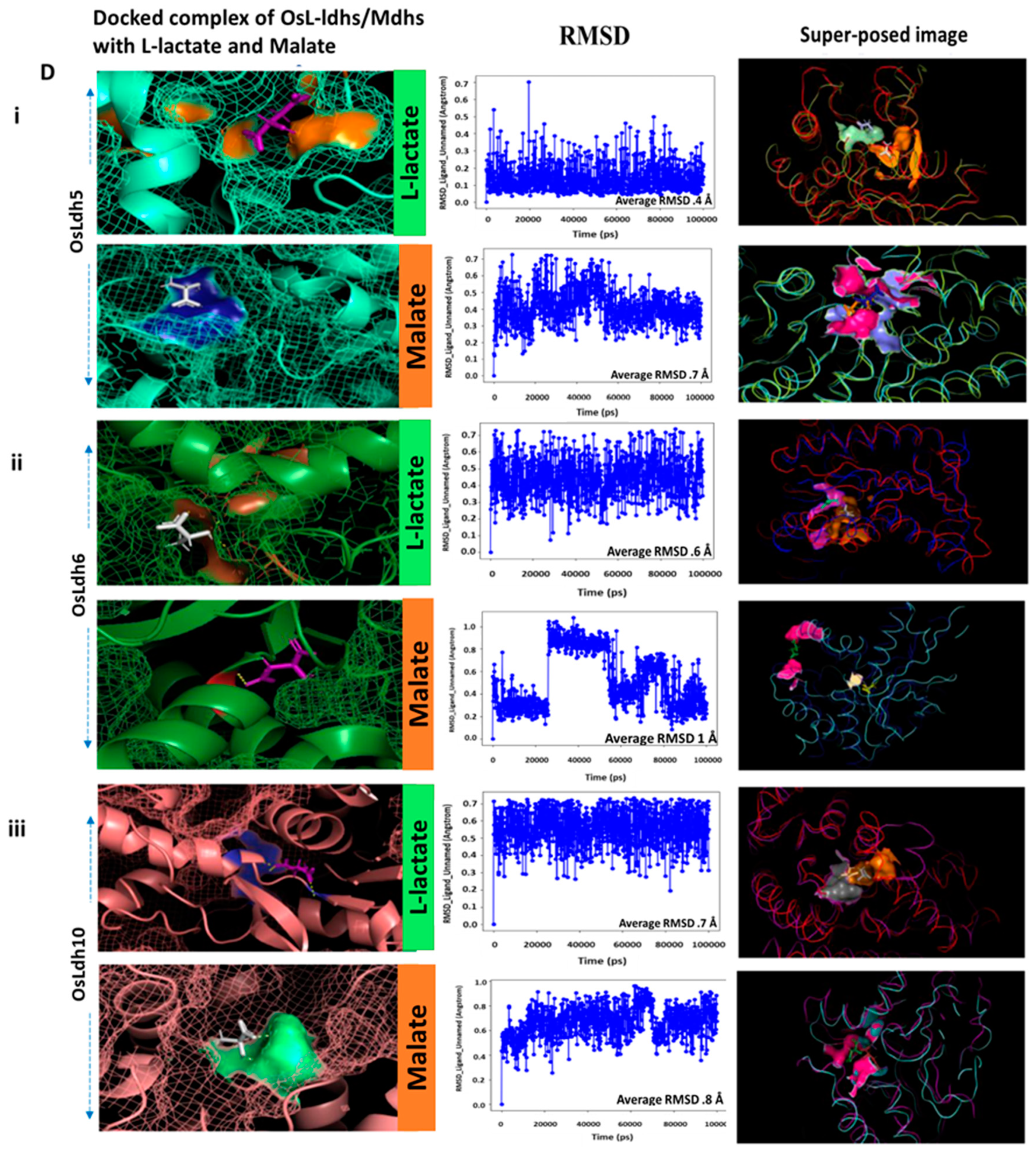
2.6. Identification of Functionally Active L-Lactate Dehydrogenases Using Schrodinger Suite
3. Discussion
4. Materials and Methods
4.1. Identification, Characterization and Domain Assessment of Ldh Genes/Proteins across Plant Species
4.2. Evolutionary Analysis of Genes Encoding Ldh across Plant Species
4.3. Gene Structure Analysis and Motif Identification of the Ldh Genes/Proteins
4.4. Identification of Putative cis-Regulatory Elements in the Promoter Region of Rice Ldh
4.5. Tissue-Specific and Stress Mediated Expression Profiling of Ldh Proteins in Rice and Arabidopsis
4.6. Plant Materials and Stress Treatments
4.7. RNA Isolation, cDNA Synthesis and qRT PCR Analysis of Rice Ldh Genes
4.8. Protein Modelling with Schrodinger Suite
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holbrook, J.J.; Liljas, A.; Steindel, S.J.; Rossmann, M.G. 4 Lactate Dehydrogenase. In The Enzymes; Boyer, P.D., Ed.; Academic Press: Cambridge, MA, USA, 1975; Volume 11, pp. 191–292. [Google Scholar]
- Roche, J.; Girard, E.; Mas, C.; Madern, D. The archaeal LDH-like malate dehydrogenase from Ignicoccus islandicus displays dual substrate recognition, hidden allostery and a non-canonical tetrameric oligomeric organization. J. Struct. Biol. 2019, 208, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Musrati, R.A.; Kollárová, M.; Mernik, N.; Mikulásová, D. Malate dehydrogenase: Distribution, function and properties. Gen. Physiol. Biophys. 1998, 17, 193–210. [Google Scholar]
- Poeydomenge, O.; Marolda, M.; Boudet, A.M.; Grima-Pettenati, J. Nucleotide sequence of a cDNA encoding mitochondrial malate dehydrogenase from Eucalyptus. Plant Physiol. 1995, 107, 1455–1456. [Google Scholar] [CrossRef] [Green Version]
- Schulze, J.; Tesfaye, M.; Litjens, R.H.M.G.; Bucciarelli, B.; Trepp, G.; Miller, S.; Samac, D.; Allan, D.; Vance, C.P. Malate plays a central role in plant nutrition. Plant Soil 2002, 247, 133–139. [Google Scholar] [CrossRef]
- Yudina, R. Malate dehydrogenase in plants: Its genetics, structure, localization and use as a marker. J. Adv. Boil. Biotechnol. 2012, 3, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.A.; Li, Q.; Ge, X.Y.; Yang, C.L.; Luo, X.L.; Zhang, A.H.; Xiao, J.L.; Tian, Y.C.; Xia, G.X.; Chen, X.Y.; et al. The mitochondrial malate dehydrogenase 1 gene GhmMDH1 is involved in plant and root growth under phosphorus deficiency conditions in cotton. Sci. Rep. 2015, 5, 10343–10357. [Google Scholar] [CrossRef] [Green Version]
- Schumann, G.; Bonora, R.; Ceriotti, F.; Clerc-Renauld, P.; Ferrero, C.A.; Férard, G.; Frank, P.F.H.; Gella, F.J.; Hoelzel, W.; Jørgensen, P.J.; et al. IFCC Primary reference procedures for the measurement of catalytic activity concentrations of enzymes at 37 °C. Part 3. Reference procedure for the measurement of catalytic concentration of lactate dehydrogenase. CCLM 2002, 40, 643–648. [Google Scholar] [CrossRef]
- James, W.O.; Cragg, J.M. The ascorbic acid system as an agent in barley respiration. New Phytol. 1943, 42, 28–44. [Google Scholar] [CrossRef]
- King, J. The isolation, properties, and physiological role of lactic dehydrogenase from soybean cotyledons. Can. J. Bot. 1970, 48, 533–540. [Google Scholar] [CrossRef]
- Davies, D.D.; Davies, S. Purification and properties of L (+)-lactate dehydrogenase from potato tubers. Biochem. J. 1972, 129, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Rothe, G. Catalytic properties of three lactate dehydrogenases from potato tubers (Solanum tuberosum). Arch. Biochem. Biophys. 1974, 162, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Oba, K.; Murakami, S.; Uritani, I. Partial purification and characterization of L-lactate dehydrogenase isozymes from sweet potato roots. J. Biochem. 1977, 81, 1193–1201. [Google Scholar]
- Betsche, T. L-Lactate dehydrogenase from leaves of higher plants-Kinetics and regulation of the enzyme from lettuce (Lactuca sativa L.). Biochem. J. 1981, 195, 615–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Carra, P.; Mulcahy, P. Lactate dehydrogenase in plants: Distribution and function. Phytochemistry 1996, 42, 581–587. [Google Scholar] [CrossRef]
- Hondred, D.; Hanson, A.D. Hypoxically inducible barley lactate dehydrogenase: cDNA cloning and molecular analysis. Proc. Natl. Acad. Sci. USA 1990, 87, 7300–7304. [Google Scholar] [CrossRef] [Green Version]
- Christopher, M.E.; Good, A.G. Characterization of hypoxically inducible lactate dehydrogenase in Maize. Plant Physiol. 1996, 112, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Germain, V.; Ricard, B. Two ldh genes from tomato and their expression in different organs, during fruit ripening and in response to stress. Plant Mol. Biol. 1997, 35, 949–954. [Google Scholar] [CrossRef]
- Dolferus, R.; Wolansky, M.; Carroll, R.; Miyashita, Y.; Ismond, K.; Good, A. Functional analysis of lactate dehydrogenase during hypoxic stress in Arabidopsis. Funct. Plant Biol. 2008, 35, 131–140. [Google Scholar] [CrossRef]
- Imran, M.; Tang, K.; Liu, J.Y. Comparative genome-wide analysis of the malate dehydrogenase gene families in cotton. PLoS ONE 2016, 11, e0166341. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Yuan, Y.; Gao, M.; Xing, L.; Li, C.; Li, M.; Ma, F. Genome-wide identification, classification, molecular evolution and expression analysis of malate dehydrogenases in Apple. Int. J. Mol. Sci. 2018, 19, 3312. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, J.; Zhang, C.; Wang, S.; Yang, M. Genome-wide investigation of malate dehydrogenase gene family in poplar (Populus trichocarpa) and their expression analysis under salt stress. Acta Physiol. Plant 2021, 43, 28. [Google Scholar] [CrossRef]
- Tihanyi, K.; Fontanell, A.; Talbot, B.; Thirion, J.P. Soybean L-(+)-lactate dehydrogenases: Purification, characterization, and resolution of subunit structure. Arch. Biochem. Biophys. 1989, 274, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Paventi, G.; Pizzuto, R.; Chieppa, G.; Passarella, S. L-Lactate metabolism in potato tuber mitochondria. FEBS J. 2007, 274, 1459–1469. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.X.; Li, M.; Zhai, H.; You, C.X.; Hao, Y.J. Isolation and characterization of an Apple cytosolic malate dehydrogenase gene reveal its function in malate synthesis. J. Plant Physiol. 2010, 168, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.S.S.; Lam, P.K.S. Glucose-6-phosphate dehydrogenase and lactate dehydrogenase in the green-lipped mussel (Pernaviridis): Possible biomarkers for hypoxia in the marine environment. Water Res. 1997, 31, 2797–2801. [Google Scholar] [CrossRef]
- Bajguz, A. Suppression of Chlorella vulgaris growth by cadmium, lead, and copper stress and its restoration by endogenous brassinolide. Arch. Environ. Contam. Toxicol. 2011, 60, 406–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niczyporuk, P.A.; Bajguz, A.; Zambrzycka, E.; Godlewska-Żyłkiewicz, B. Phytohormones as regulators of heavy metal biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiol. Biochem. 2012, 52, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Histidine kinases in plants: Cross talk between hormone and stress responses. Plant Signal. Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Singla-Pareek, S.L.; Pareek, A. Two-component signaling system in plants: Interaction network and specificity in response to stress and hormones. Plant Cell Rep. 2021, 40, 2037–2046. [Google Scholar] [CrossRef]
- Tseng, Y.C.; Lee, J.R.; Chang, J.C.H.; Kuo, C.H.; Lee, S.J.; Hwang, P.P. Regulation of lactate dehydrogenase in tilapia (Oreochromis mossambicus) gills during acclimation to salinity challenge. Zool. Stud. 2008, 47, 473–480. [Google Scholar]
- Pereira, C.M.; Fonseca, J.S.; Paiva, E.S.; Costa, P.G.; Mies, M.; Silva, A.G.; Calderon, E.N.; Bianchini, A.; Castro, C.B. Larvae of the South Atlantic coral Favia gravida are tolerant to salinity and nutrient concentrations associated with river discharges. Mar. Environ. Res. 2020, 161, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S. Glyoxalase and methylglyoxal as biomarkers for plant stress tolerance. CRC Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Singla-Pareek, S.L.; Kaur, C.; Kumar, B.; Pareek, A.; Sopory, S.K. Reassessing plant glyoxalases: Large family and expanding functions. New Phytol. 2020, 227, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilks, H.M.; Hart, K.W.; Feeney, R.; Dunn, C.R.; Muirhead, H.; Chia, W.N.; Barstow, D.A.; Atkinson, T.; Clarke, A.R.; Holbrook, J.J. A specific, highly active malate dehydrogenase by redesign of a lactate dehydrogenase framework. Science 1988, 242, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.J.; Miller, J.; Scawen, M.D.; Clarke, A.R.; Holbrook, J.J.; Atkinson, T.; Goward, C.R. The importance of arginine 102 for the substrate specificity of Escherichia coli malate dehydrogenase. Biochem. Biophys. Res. Commun. 1992, 189, 1057–1062. [Google Scholar] [CrossRef]
- Cendrin, F.; Chroboczek, J.; Zaccai, G.; Eisenberg, H.; Mevarech, M. Cloning, sequencing, and expression in Escherichia coli of the gene coding for malate dehydrogenase of the extremely halophilic archaebacterium Haloarcula marismortui. Biochem. 1993, 32, 4308–4313. [Google Scholar] [CrossRef]
- Barthová, J.; Plachá, P.; Leblová, S. Isolation of plant lactate dehydrogenase by affinity chromatography and the role of histidine in the molecule of the enzyme. Collect. Czech. Chem. Commun. 1980, 45, 1608–1615. [Google Scholar] [CrossRef]
- Madern, D. The putative l-lactate dehydrogenase from Methanococcus jannaschii is an NADPH dependent l-malate dehydrogenase. Mol. Microbiol. 2000, 37, 1515–1520. [Google Scholar] [CrossRef] [Green Version]
- Madern, D. Molecular evolution within the L-Malate and L-Lactate Dehydrogenase Super-Family. J. Mol. Evol. 2002, 54, 825–840. [Google Scholar] [CrossRef]
- Armenteros, J.J.A.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1995, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Friesner, R.A.; Xiang, Z.; Honig, B. On the role of crystal packing forces in determining protein sidechain conformations. J. Mol. Biol. 2002, 320, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A hierarchical approach to all-atom protein loop prediction. Proteins Struct. Funct. Bioinform. 2004, 55, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Kevin, J.B.; Edmond, C.; Huafeng, X.; Dror, R.; Eastwood, M.; Gregerson, B.A.; Klepeis, J.L.; Kolossváry, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable algorithms for molecular dynamics simulations on commodity clusters. In Proceedings of the ACM/IEEE conference on Supercomputing (SC ‘06). Association for Computing Machinery, New York, NY, USA, 11–17 November 2006; pp. 455–544. [Google Scholar]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | Transcripts | Coordinate (5′-3′) | CDS Length (bp) | Protein | Localisation | ||
|---|---|---|---|---|---|---|---|---|
| Amino Acid Length | pI | Mol. wt (kDa) | ||||||
| AtLdh1 | AT1G04410 | AT1G04410.1 | 1189078-1191412 | 999 | 332 | 6.11 | 35.57 | cytoplasm |
| AtLdh2 | AT1G53240 | AT1G53240.1 | 19854615-19856937 | 1026 | 341 | 8.54 | 35.8 | mitochondria |
| AtLdh3.1 | AT3G15020 | AT3G15020.1 | 5056068-5058248 | 1026 | 341 | 8.3 | 35.87 | mitochondria |
| AtLdh3.2 | AT3G15020.2 | 5056068-5058248 | 951 | 316 | 9.49 | 33.13 | mitochondria | |
| AtLdh4 | AT4G17260 | AT4G17260.1 | 9673991-9675448 | 1062 | 353 | 6.07 | 37.95 | cytoplasm |
| AtLdh5.1 | AT5G09660 | AT5G09660.1 | 2993444-2995675 | 1065 | 354 | 8.14 | 37.36 | cytoplasm |
| AtLdh5.2 | AT5G09660.2 | 2993444-2995307 | 1002 | 333 | 7.56 | 34.95 | cytoplasm | |
| AtLdh5.3 | AT5G09660.3 | 2993444-2995307 | 1029 | 342 | 9.06 | 36.34 | cytoplasm | |
| AtLdh5.4 | AT5G09660 | AT5G09660.4 | 2993444-2995675 | 1092 | 363 | 9.16 | 38.73 | cytoplasm |
| AtLdh6 | AT5G43330 | AT5G43330.1 | 17390433-17392645 | 999 | 332 | 6.33 | 35.67 | cytoplasm |
| AtLdh7 | AT5G56720 | AT5G56720.1 | 22945537-22946718 | 1020 | 339 | 5.75 | 36.87 | cytoplasm |
| AtLdh8.1 | AT5G58330 | AT5G58330.1 | 3579722-23582395 | 1332 | 443 | 5.81 | 48.3 | chloroplast |
| AtLdh8.2 | AT5G58330.2 | 23579722-23582395 | 1329 | 442 | 5.81 | 48.22 | chloroplast | |
| AtLdh8.3 | AT5G58330.3 | 23579722-23582295 | 1005 | 334 | 4.95 | 36.39 | cytoplasm | |
| OsLdh1 | Os01g46070 | Os01g46070.1 | 26190752-26194517 | 1023 | 340 | 8.74 | 35.46 | mitochondria |
| OsLdh2 | Os01g61380 | Os01g61380.1 | 35499017-35501765 | 1191 | 396 | 7.63 | 41.78 | chloroplast |
| OsLdh3 | Os02g01510 | Os02g01510.1 | 295302-299174 | 1179 | 392 | 6.74 | 42.71 | cytoplasm |
| OsLdh4 | Os03g56280 | Os03g56280.1 | 32089685-32086001 | 1065 | 354 | 8.13 | 37.02 | cytoplasm |
| OsLdh5 | Os04g46560 | Os04g46560.1 | 27605166-27608347 | 1059 | 353 | 6.72 | 38.29 | mitochondria |
| OsLdh6 | Os05g49880 | Os05g49880.1 | 28621585-28617595 | 1023 | 340 | 8.22 | 35.43 | mitochondria |
| OsLdh6 | Os05g49880 | Os05g49880.2 | 28621585-28617595 | 696 | 231 | 7.21 | 32.93 | mitochondria |
| OsLdh7 | Os06g01590 | Os06g01590.1 | 348516-346985 | 1083 | 360 | 7.9 | 38.72 | cytoplasm |
| OsLdh8 | Os07g43700 | Os07g43700.1 | 26155933-26153825 | 1215 | 404 | 9 | 42.22 | cytoplasm |
| OsLdh9 | Os08g33720 | Os08g33720.1 | 21057561-21054659 | 1194 | 397 | 7.02 | 41.53 | chloroplast |
| OsLdh10 | Os08g44810 | Os08g44810.1 | 28141042-28146270 | 1302 | 434 | 6.96 | 47 | chloroplast |
| OsLdh11 | Os10g33800 | Os10g33800.1 | 17913818-17917850 | 999 | 333 | 5.75 | 35.56 | cytoplasm |
| OsLdh12 | Os12g43630 | Os12g43630.1 | 27099351-27094647 | 1071 | 357 | 8.09 | 37.38 | chloroplast |
| Proteins | Ligand | |||||
|---|---|---|---|---|---|---|
| Malate | L-Lactate | |||||
| Interacting Amino Acids | Docking Score | RMSD Value (Å) | Interacting Amino Acids | Docking Score | RMSD Value (Å) | |
| AtLdh1 | Asn-193 | −6.993 | 0.25 | Arg-227 | −4.3 | 0.5 |
| Val-195 | −6.993 | His-251 | −4.3 | |||
| Arg-227 | −6.993 | |||||
| AtLdh4 | Ser-195 | −4.304 | 0.6 | Ser-219 | −4.67 | 0.25 |
| Ser-219 | Gly-220 | |||||
| Leu-223 | Leu-223 | |||||
| His-251 | Ser-313 | |||||
| Ser-313 | ||||||
| Osldh1 | Thr-222 | −4.384 | 0.9 | Arg-227 | −3.619 | 0.55 |
| His-251 | ||||||
| OsLdh2 | Asn-193 | −4.249 | 0.79 | Arg-227 | −3.619 | 0.7 |
| Arg-227 | His-251 | |||||
| OsLdh3 | Leu-223 | −3.443 | 0.6-1 | Asn-196 | −3.864 | 0.29 |
| Asp-224 | Ser-219 | |||||
| Gly-220 | ||||||
| OsLdh4 | Ile-158 | −4.234 | 1.1 | Asn-160 | −4.205 | 0.3 |
| Asn-160 | Val-187 | |||||
| Leu-190 | Leu-190 | |||||
| OsLdh5 | Asn-193 | −4.634 | 0.7 | Val-151 | −3.325 | 0.4 |
| Val-195 | Asn-153 | |||||
| Arg-227 | ||||||
| OsLdh6 | Thr-222 | −3.619 | 1 | Arg-227 | −3.619 | 0.6 |
| His-251 | ||||||
| OsLdh7 | Asn-193 | −2.4 | 0.4 | Ser-219 | −6.693 | 0.15 |
| Val-195 | Gly-220 | |||||
| Arg-227 | ||||||
| OsLdh8 | Arg-157 | −3.489 | 0.6 | Val-222 | −3.258 | 0.4 |
| Asn-195 | Leu-225 | |||||
| Arg-229 | Asp-226 | |||||
| OsLdh9 | Asn-193 | −1.865 | 0.8 | Arg-227 | −4.042 | 0.15 |
| Val-195 | His-251 | |||||
| Arg-227 | ||||||
| OsLdh10 | Leu-159 | −3.167 | 0.8 | Val-130 | −3.253 | 0.7 |
| OsLdh11 | Asn-193 | −4.634 | 0.5 | Arg-227 | −3.619 | 0.6 |
| Val-195 | His-251 | |||||
| Arg-227 | ||||||
| OsLdh12 | Asn-193 | −4.634 | 0.4 | Arg-227 | −3.619 | 0.7 |
| Val-195 | His-251 | |||||
| Arg-227 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatterjee, Y.; Bhowal, B.; Gupta, K.J.; Pareek, A.; Singla-Pareek, S.L. Lactate Dehydrogenase Superfamily in Rice and Arabidopsis: Understanding the Molecular Evolution and Structural Diversity. Int. J. Mol. Sci. 2023, 24, 5900. https://doi.org/10.3390/ijms24065900
Chatterjee Y, Bhowal B, Gupta KJ, Pareek A, Singla-Pareek SL. Lactate Dehydrogenase Superfamily in Rice and Arabidopsis: Understanding the Molecular Evolution and Structural Diversity. International Journal of Molecular Sciences. 2023; 24(6):5900. https://doi.org/10.3390/ijms24065900
Chicago/Turabian StyleChatterjee, Yajnaseni, Bidisha Bhowal, Kapuganti Jagadis Gupta, Ashwani Pareek, and Sneh Lata Singla-Pareek. 2023. "Lactate Dehydrogenase Superfamily in Rice and Arabidopsis: Understanding the Molecular Evolution and Structural Diversity" International Journal of Molecular Sciences 24, no. 6: 5900. https://doi.org/10.3390/ijms24065900
APA StyleChatterjee, Y., Bhowal, B., Gupta, K. J., Pareek, A., & Singla-Pareek, S. L. (2023). Lactate Dehydrogenase Superfamily in Rice and Arabidopsis: Understanding the Molecular Evolution and Structural Diversity. International Journal of Molecular Sciences, 24(6), 5900. https://doi.org/10.3390/ijms24065900