
NANOBODIES®: A Review of Diagnostic and Therapeutic Applications

Abstract
:1. Introduction
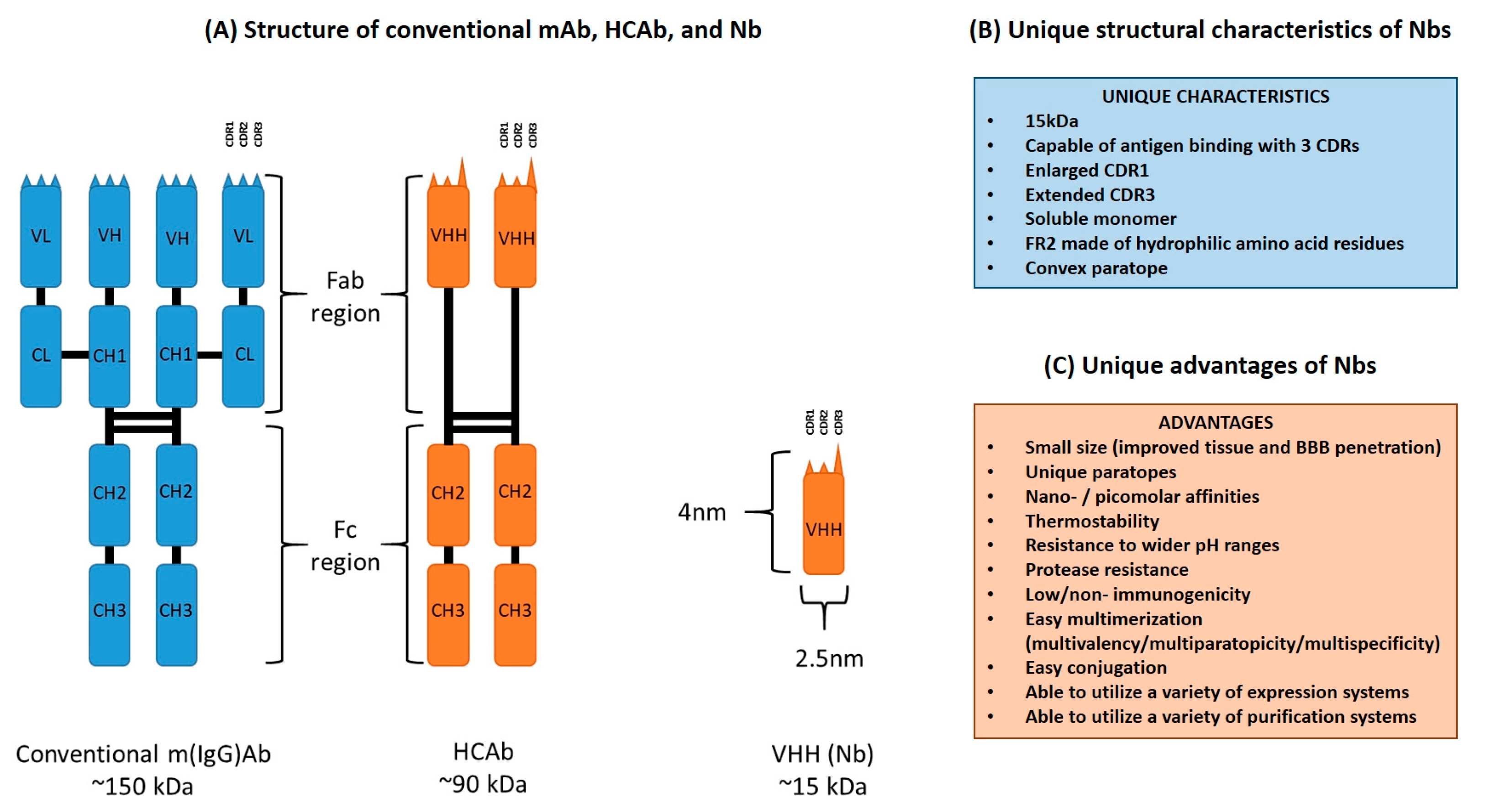
2. Unique Structural Features of Nanobodies
3. Desirable Properties of Nanobodies over Conventional Monoclonal Antibodies
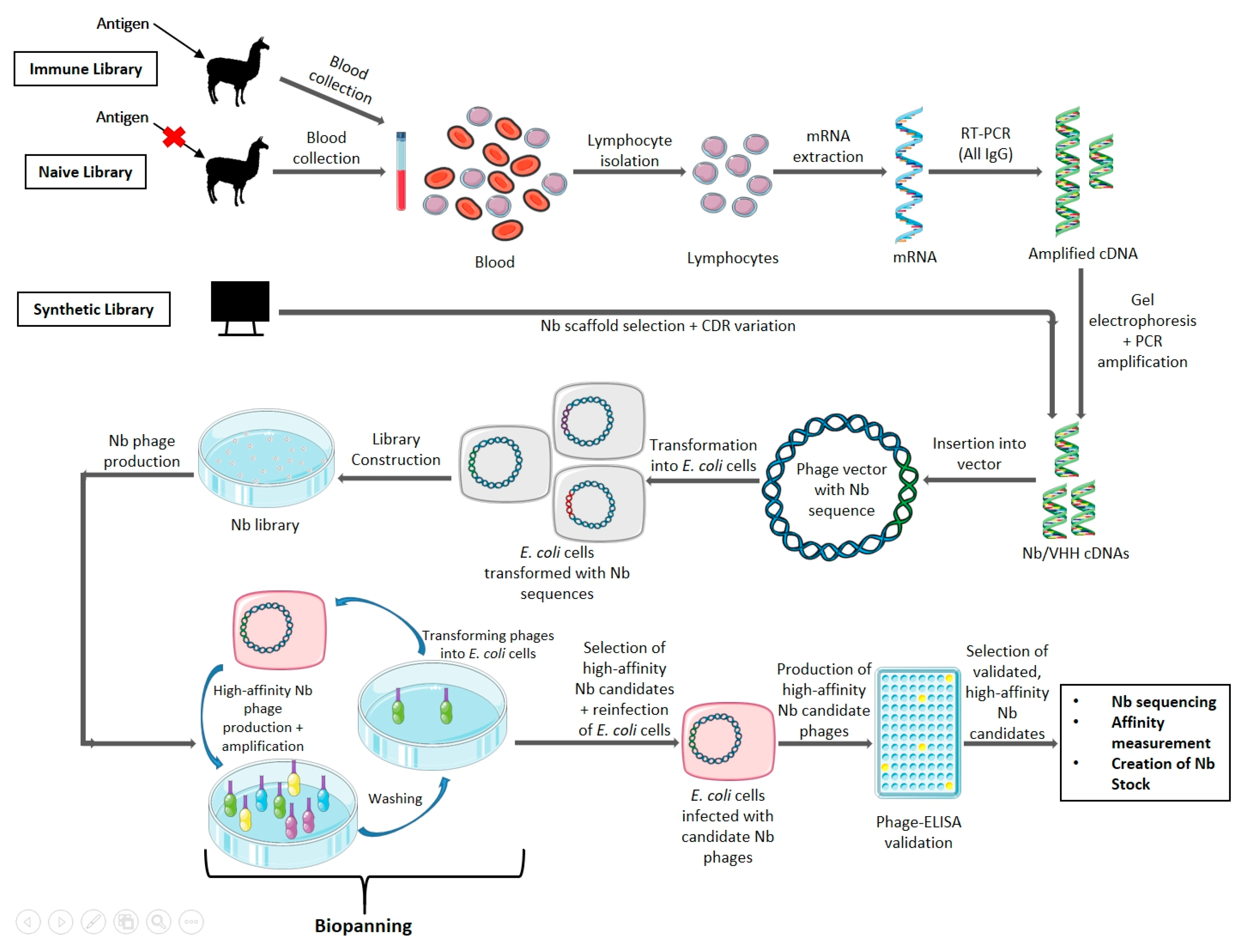
4. Nanobody Generation and Production
4.1. Generation
4.2. Modifications to the Standard Nanobody Generation Protocol
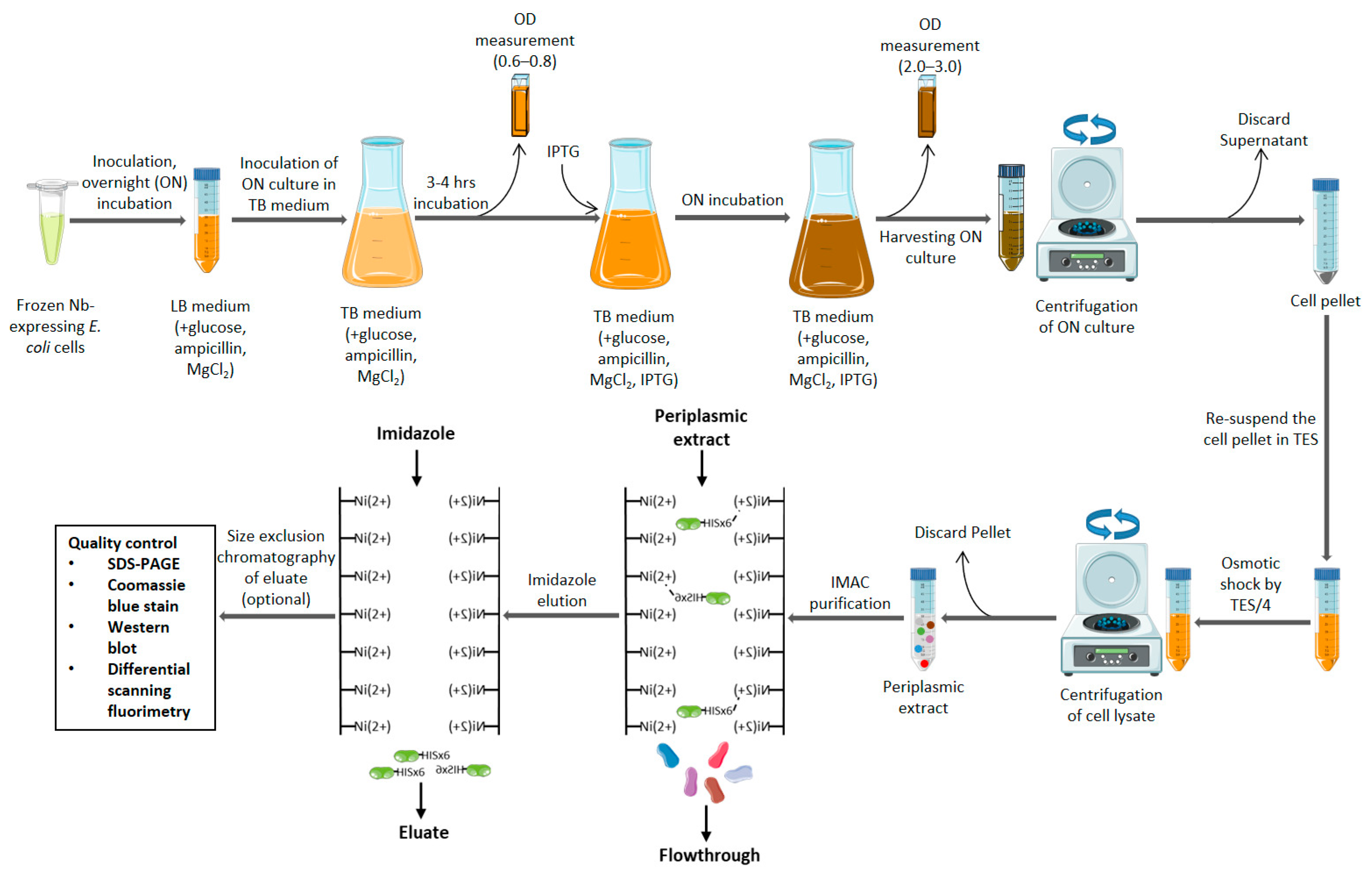
4.3. Production and Purification
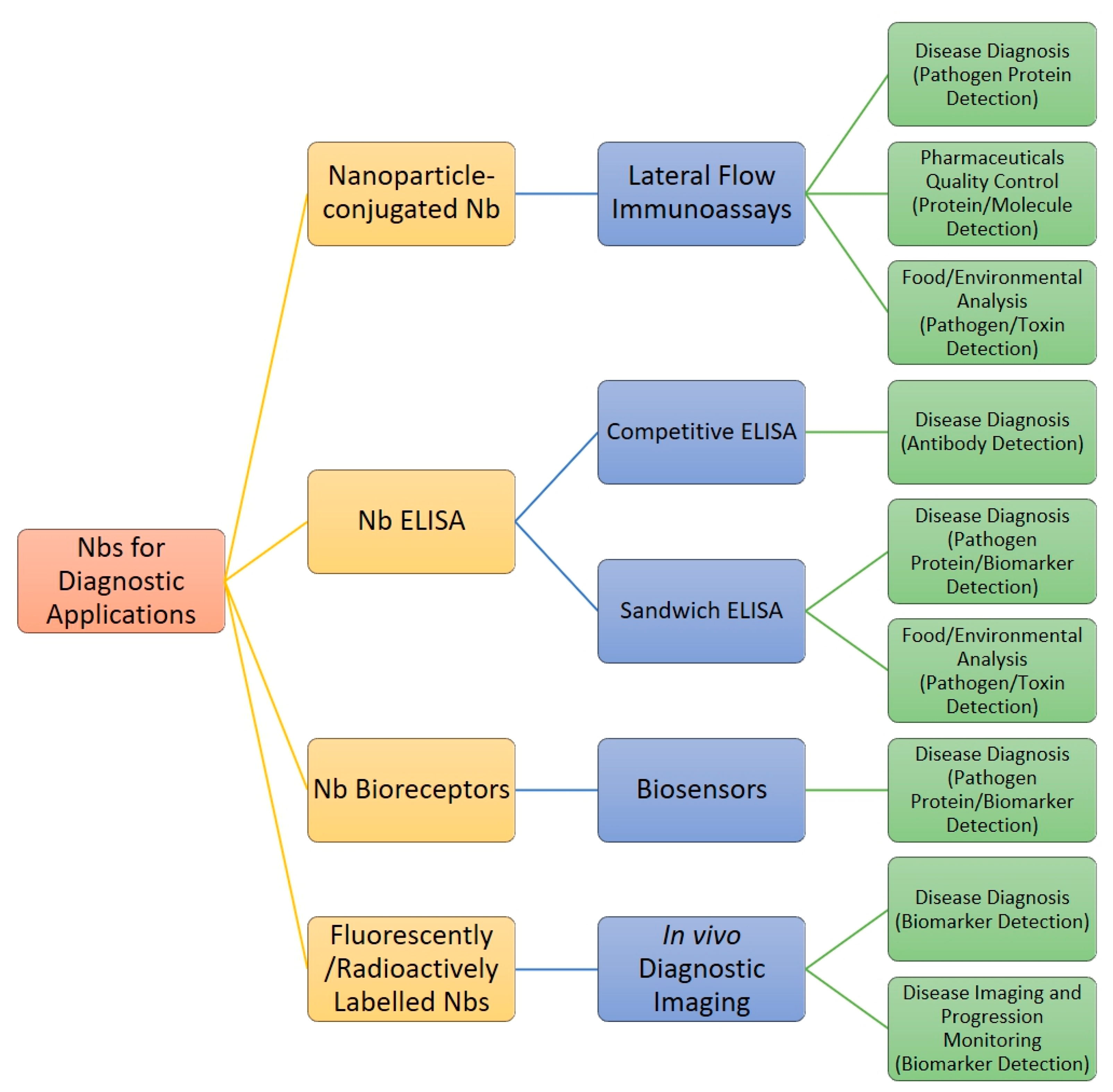
5. Diagnostic Applications of Nanobodies
5.1. Nanobody Immunoassay Format: Lateral Flow Immunoassays and Diagnostic ELISAs
5.2. Biosensors
5.3. In Vivo Diagnostic Imaging
6. Therapeutical Applications of Nanobodies
6.1. Nanobodies Against Cancer
6.2. Nanobodies Against Autoimmune Diseases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product Name | Disease and/or Condition Targeted | Target Antigen | Clinical Trial Status | Manufacturer | References |
|---|---|---|---|---|---|
| 68-GaNOTA-Anti-HER2 VHH1 1 | Breast carcinoma | HER2 | Phase 2 | Universitair Ziekenhuis Brussel, Brussels, Belgium | [113,114] |
| Ciltacabtagene autoleucel, LCAR-B38M | Refractory/relapsed multiple myeloma | B-cell maturation antigen | Approved | Janssen Research & Development, LLC, Raritan, United States | [123,124,125] |
| 131I-GMIB-Anti-HER2-VHH1 | Breast carcinoma | HER2 | Phase 1 | Precirix, Brussels, Belgium | [132] |
| Calplacizumab | Acquired thrombotic thrombocytopenic purpura | Von Willebrand factor | Approved (USA, EU) | Ablynx, Ghent, Belgium | [133,134] |
| Ozoralizumab | Rheumatoid arthritis | Tumor necrosis factor-alpha | Approved (Japan) | Taisho Pharmaceuticals, Tokyo, Japan | [135] |
| Vobarilizumab (ALX-0061) | Rheumatoid arthritis, systemic lupus erythematosus | Interleukin-6 receptor | Phase 2 | Ablynx, Ghent, Belgium | [136,137] |
| Sonelokimab (M1095) | Psoriasis | Interleukin-17A/F | Phase 2 | Bond Avillion 2 Development LP, London, England | [138] |
| Gefurulimab (ALXN1720) | Myasenthia Gravis | Autoantibodies against acetylcholine receptors | Phase 3 | Alexion Pharmaceuticals, Boston, United States | [139] |
| M6495 | Osteoarthritis | A Disintegrin and Metalloproteinase with Thrombospondin Motifs-5 | Phase 1 | Merck KGaA, Darmstadt, Germany | [140] |
| Nb V565 | Crohn’s Disease | Tumor necrosis factor | Phase 2 | VHsquared Ltd., Copenhagen, Denmark | [141] |
| ARP1, VHH batch 203027 | Diarrhea | Rotavirus | Phase 2 | International Centre for Diarrhoeal Disease Research, Dhaka, Bangladesh | [144] |
| ALX-0171 | Lower respiratory tract infection | Respiratory syncytial virus | Phase 2 | Ablynx, Ghent, Belgium | [145] |
| LMN-101 | Campylobacteriosis | Campylobacter jejuni | Phase 2 | Lumen Bioscience, Inc., Seattle, United States | [146] |
6.3. Nanobodies Against Infectious Diseases
6.4. Nanobodies Against Toxins and Venoms
7. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Desmyter, A.; Transue, T.R.; Ghahroudi, M.A.; Thi, M.H.; Poortmans, F.; Hamers, R.; Muyldermans, S.; Wyns, L. Crystal structure of a camel single-domain VH antibody fragment in complex with lysozyme. Nat. Struct. Biol. 1996, 3, 803–811. [Google Scholar] [CrossRef]
- Spinelli, S.; Frenken, L.; Bourgeois, D.; de Ron, L.; Bos, W.; Verrips, T.; Anguille, C.; Cambillau, C.; Tegoni, M. The crystal structure of a llama heavy chain variable domain. Nat. Struct. Biol. 1996, 3, 752–757. [Google Scholar] [CrossRef]
- Arbabi Ghahroudi, M.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar] [CrossRef]
- Nguyen, V.K.; Hamers, R.; Wyns, L.; Muyldermans, S. Loss of splice consensus signal is responsible for the removal of the entire C(H)1 domain of the functional camel IGG2A heavy-chain antibodies. Mol. Immunol. 1999, 36, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Woolven, B.P.; Frenken, L.G.; van der Logt, P.; Nicholls, P.J. The structure of the llama heavy chain constant genes reveals a mechanism for heavy-chain antibody formation. Immunogenetics 1999, 50, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Nanobodies: Natural single-domain antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef]
- Salvador, J.P.; Vilaplana, L.; Marco, M.P. Nanobody: Outstanding features for diagnostic and therapeutic applications. Anal. Bioanal. Chem. 2019, 411, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Lobato, M.N.; Rabbitts, T.H. Single domain intracellular antibodies: A minimal fragment for direct in vivo selection of antigen-specific intrabodies. J. Mol. Biol. 2003, 331, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S.; Lauwereys, M. Unique single-domain antigen binding fragments derived from naturally occurring camel heavy-chain antibodies. J. Mol. Recognit. JMR 1999, 12, 131–140. [Google Scholar] [CrossRef]
- Muyldermans, S.; Baral, T.N.; Retamozzo, V.C.; De Baetselier, P.; De Genst, E.; Kinne, J.; Leonhardt, H.; Magez, S.; Nguyen, V.K.; Revets, H.; et al. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunopathol. 2009, 128, 178–183. [Google Scholar] [CrossRef]
- Sun, S.; Ding, Z.; Yang, X.; Zhao, X.; Zhao, M.; Gao, L.; Chen, Q.; Xie, S.; Liu, A.; Yin, S.; et al. Nanobody: A Small Antibody with Big Implications for Tumor Therapeutic Strategy. Int. J. Nanomed. 2021, 16, 2337–2356. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S.; Cambillau, C.; Wyns, L. Recognition of antigens by single-domain antibody fragments: The superfluous luxury of paired domains. Trends Biochem. Sci. 2001, 26, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.K.; Hamers, R.; Wyns, L.; Muyldermans, S. Camel heavy-chain antibodies: Diverse germline V(H)H and specific mechanisms enlarge the antigen-binding repertoire. EMBO J. 2000, 19, 921–930. [Google Scholar] [CrossRef]
- Decanniere, K.; Muyldermans, S.; Wyns, L. Canonical antigen-binding loop structures in immunoglobulins: More structures, more canonical classes? J. Mol. Biol. 2000, 300, 83–91. [Google Scholar] [CrossRef]
- Muyldermans, S.; Atarhouch, T.; Saldanha, J.; Barbosa, J.A.; Hamers, R. Sequence and structure of VH domain from naturally occurring camel heavy chain immunoglobulins lacking light chains. Protein Eng. 1994, 7, 1129–1135. [Google Scholar] [CrossRef]
- Vu, K.B.; Ghahroudi, M.A.; Wyns, L.; Muyldermans, S. Comparison of llama VH sequences from conventional and heavy chain antibodies. Mol. Immunol. 1997, 34, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- De Genst, E.; Silence, K.; Decanniere, K.; Conrath, K.; Loris, R.; Kinne, J.; Muyldermans, S.; Wyns, L. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. Proc. Natl. Acad. Sci. USA 2006, 103, 4586–4591. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.S.; Colwell, L.J. Analysis of nanobody paratopes reveals greater diversity than classical antibodies. Protein Eng. Des. Sel. PEDS 2018, 31, 267–275. [Google Scholar] [CrossRef]
- Mitchell, L.S.; Colwell, L.J. Comparative analysis of nanobody sequence and structure data. Proteins 2018, 86, 697–706. [Google Scholar] [CrossRef]
- Muyldermans, S. Single domain camel antibodies: Current status. J. Biotechnol. 2001, 74, 277–302. [Google Scholar] [CrossRef]
- Chothia, C.; Novotný, J.; Bruccoleri, R.; Karplus, M. Domain association in immunoglobulin molecules. Pack. Var. Domains. J. Mol. Biol. 1985, 186, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.K.; Desmyter, A.; Muyldermans, S. Functional heavy-chain antibodies in Camelidae. Adv. Immunol. 2001, 79, 261–296. [Google Scholar] [CrossRef] [PubMed]
- Zavrtanik, U.; Lukan, J.; Loris, R.; Lah, J.; Hadži, S. Structural Basis of Epitope Recognition by Heavy-Chain Camelid Antibodies. J. Mol. Biol. 2018, 430, 4369–4386. [Google Scholar] [CrossRef] [PubMed]
- Padlan, E.A. X-ray crystallography of antibodies. Adv. Protein Chem. 1996, 49, 57–133. [Google Scholar] [CrossRef] [PubMed]
- Jovčevska, I.; Muyldermans, S. The Therapeutic Potential of Nanobodies. BioDrugs Clin. Immunother. Biopharm. Gene Ther. 2020, 34, 11–26. [Google Scholar] [CrossRef]
- Khodabakhsh, F.; Behdani, M.; Rami, A.; Kazemi-Lomedasht, F. Single-Domain Antibodies or Nanobodies: A Class of Next-Generation Antibodies. Int. Rev. Immunol. 2018, 37, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Sockolosky, J.T.; Dougan, M.; Ingram, J.R.; Ho, C.C.; Kauke, M.J.; Almo, S.C.; Ploegh, H.L.; Garcia, K.C. Durable antitumor responses to CD47 blockade require adaptive immune stimulation. Proc. Natl. Acad. Sci. USA 2016, 113, E2646–E2654. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.A. Exploring alternative antibody scaffolds: Antibody fragments and antibody mimics for targeted drug delivery. Drug discovery today. Technologies 2018, 30, 35–46. [Google Scholar] [CrossRef]
- Yang, E.Y.; Shah, K. Nanobodies: Next Generation of Cancer Diagnostics and Therapeutics. Front. Oncol. 2020, 10, 1182. [Google Scholar] [CrossRef]
- Ruiz-López, E.; Schuhmacher, A.J. Transportation of Single-Domain Antibodies through the Blood-Brain Barrier. Biomolecules 2021, 11, 1131. [Google Scholar] [CrossRef]
- Caljon, G.; Caveliers, V.; Lahoutte, T.; Stijlemans, B.; Ghassabeh, G.H.; Van Den Abbeele, J.; Smolders, I.; De Baetselier, P.; Michotte, Y.; Muyldermans, S.; et al. Using microdialysis to analyse the passage of monovalent nanobodies through the blood-brain barrier. Br. J. Pharmacol. 2012, 165, 2341–2353. [Google Scholar] [CrossRef] [PubMed]
- Wouters, Y.; Jaspers, T.; De Strooper, B.; Dewilde, M. Identification and in vivo characterization of a brain-penetrating nanobody. Fluids Barriers CNS 2020, 17, 62. [Google Scholar] [CrossRef]
- Steeland, S.; Van Ryckeghem, S.; Van Imschoot, G.; De Rycke, R.; Toussaint, W.; Vanhoutte, L.; Vanhove, C.; De Vos, F.; Vandenbroucke, R.E.; Libert, C. TNFR1 inhibition with a Nanobody protects against EAE development in mice. Sci. Rep. 2017, 7, 13646. [Google Scholar] [CrossRef] [PubMed]
- van de Water, J.A.; Bagci-Onder, T.; Agarwal, A.S.; Wakimoto, H.; Roovers, R.C.; Zhu, Y.; Kasmieh, R.; Bhere, D.; Van Bergen en Henegouwen, P.M.; Shah, K. Therapeutic stem cells expressing variants of EGFR-specific nanobodies have antitumor effects. Proc. Natl. Acad. Sci. USA 2012, 109, 16642–16647. [Google Scholar] [CrossRef] [PubMed]
- Samec, N.; Jovcevska, I.; Stojan, J.; Zottel, A.; Liovic, M.; Myers, M.P.; Muyldermans, S.; Šribar, J.; Križaj, I.; Komel, R. Glioblastoma-specific anti-TUFM nanobody for in-vitro immunoimaging and cancer stem cell targeting. Oncotarget 2018, 9, 17282–17299. [Google Scholar] [CrossRef]
- Zottel, A.; Jovčevska, I.; Šamec, N.; Mlakar, J.; Šribar, J.; Križaj, I.; Skoblar Vidmar, M.; Komel, R. Anti-vimentin, anti-TUFM, anti-NAP1L1 and anti-DPYSL2 nanobodies display cytotoxic effect and reduce glioblastoma cell migration. Ther. Adv. Med. Oncol. 2020, 12, 1758835920915302. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.M.; Renisio, J.G.; Prompers, J.J.; van Platerink, C.J.; Cambillau, C.; Darbon, H.; Frenken, L.G. Thermal unfolding of a llama antibody fragment: A two-state reversible process. Biochemistry 2001, 40, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Dumoulin, M.; Conrath, K.; Van Meirhaeghe, A.; Meersman, F.; Heremans, K.; Frenken, L.G.; Muyldermans, S.; Wyns, L.; Matagne, A. Single-domain antibody fragments with high conformational stability. Protein Sci. Publ. Protein Soc. 2002, 11, 500–515. [Google Scholar] [CrossRef]
- Xu, L.; Song, X.; Jia, L. A camelid nanobody against EGFR was easily obtained through refolding of inclusion body expressed in Escherichia coli. Biotechnol. Appl. Biochem. 2017, 64, 895–901. [Google Scholar] [CrossRef]
- Mohseni, A.; Molakarimi, M.; Taghdir, M.; Sajedi, R.H.; Hasannia, S. Exploring single-domain antibody thermostability by molecular dynamics simulation. J. Biomol. Struct. Dyn. 2019, 37, 3686–3696. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.; Zinner, K.; Mücke, N.; Bartoschik, T.; Muyldermans, S.; Hoheisel, J.D. The structural basis of nanobody unfolding reversibility and thermoresistance. Sci. Rep. 2018, 8, 7934. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.; Flock, T.; Soler, N.; Zaiss, M.; Vincke, C.; Sterckx, Y.; Kastelic, D.; Muyldermans, S.; Hoheisel, J.D. Exploiting sequence and stability information for directing nanobody stability engineering. Biochimica et biophysica acta. Gen. Subj. 2017, 1861, 2196–2205. [Google Scholar] [CrossRef] [PubMed]
- Abbady, A.Q.; Al-Daoude, A.; Al-Mariri, A.; Zarkawi, M.; Muyldermans, S. Chaperonin GroEL a Brucella immunodominant antigen identified using Nanobody and MALDI-TOF-MS technologies. Vet. Immunol. Immunopathol. 2012, 146, 254–263. [Google Scholar] [CrossRef]
- Hussack, G.; Hirama, T.; Ding, W.; Mackenzie, R.; Tanha, J. Engineered single-domain antibodies with high protease resistance and thermal stability. PLoS ONE 2011, 6, e28218. [Google Scholar] [CrossRef]
- Baral, T.N.; Magez, S.; Stijlemans, B.; Conrath, K.; Vanhollebeke, B.; Pays, E.; Muyldermans, S.; De Baetselier, P. Experimental therapy of African trypanosomiasis with a nanobody-conjugated human trypanolytic factor. Nat. Med. 2006, 12, 580–584. [Google Scholar] [CrossRef]
- Coppieters, K.; Dreier, T.; Silence, K.; de Haard, H.; Lauwereys, M.; Casteels, P.; Beirnaert, E.; Jonckheere, H.; Van de Wiele, C.; Staelens, L.; et al. Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis. Arthritis Rheum. 2006, 54, 1856–1866. [Google Scholar] [CrossRef]
- Ackaert, C.; Smiejkowska, N.; Xavier, C.; Sterckx, Y.G.J.; Denies, S.; Stijlemans, B.; Elkrim, Y.; Devoogdt, N.; Caveliers, V.; Lahoutte, T.; et al. Immunogenicity Risk Profile of Nanobodies. Front. Immunol. 2021, 12, 632687. [Google Scholar] [CrossRef]
- Rossotti, M.A.; Bélanger, K.; Henry, K.A.; Tanha, J. Immunogenicity and humanization of single-domain antibodies. FEBS J. 2022, 289, 4304–4327. [Google Scholar] [CrossRef]
- Vincke, C.; Loris, R.; Saerens, D.; Martinez-Rodriguez, S.; Muyldermans, S.; Conrath, K. General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold. J. Biol. Chem. 2009, 284, 3273–3284. [Google Scholar] [CrossRef]
- Moutel, S.; Bery, N.; Bernard, V.; Keller, L.; Lemesre, E.; de Marco, A.; Ligat, L.; Rain, J.C.; Favre, G.; Olichon, A.; et al. NaLi-H1: A universal synthetic library of humanized nanobodies providing highly functional antibodies and intrabodies. eLife 2016, 5, e16228. [Google Scholar] [CrossRef] [PubMed]
- Saerens, D.; Ghassabeh, G.H.; Muyldermans, S. Single-domain antibodies as building blocks for novel therapeutics. Curr. Opin. Pharmacol. 2008, 8, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Le Bas, A.; Ruza, R.R.; Duyvesteyn, H.M.E.; Mikolajek, H.; Malinauskas, T.; Tan, T.K.; Rijal, P.; Dumoux, M.; Ward, P.N.; et al. Neutralizing nanobodies bind SARS-CoV-2 spike RBD and block interaction with ACE2. Nat. Struct. Mol. Biol. 2020, 27, 846–854. [Google Scholar] [CrossRef]
- Hultberg, A.; Temperton, N.J.; Rosseels, V.; Koenders, M.; Gonzalez-Pajuelo, M.; Schepens, B.; Ibañez, L.I.; Vanlandschoot, P.; Schillemans, J.; Saunders, M.; et al. Llama-derived single domain antibodies to build multivalent, superpotent and broadened neutralizing anti-viral molecules. PLoS ONE 2011, 6, e17665. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zeng, W.; Meng, X.; Huang, X.; Yang, Y.; Zhao, D.; Zhou, P.; Wang, X.; Zhao, C.; Sun, Y.; et al. Potent Neutralization of SARS-CoV-2 by Hetero-bivalent Alpaca Nanobodies Targeting the Spike Receptor-Binding Domain. J. Virol. 2021, 95, e02438-20, Advance online publication. [Google Scholar] [CrossRef] [PubMed]
- de Beer, M.A.; Giepmans, B.N.G. Nanobody-Based Probes for Subcellular Protein Identification and Visualization. Front. Cell. Neurosci. 2020, 14, 573278. [Google Scholar] [CrossRef]
- Bobkov, V.; Zarca, A.M.; Van Hout, A.; Arimont, M.; Doijen, J.; Bialkowska, M.; Toffoli, E.; Klarenbeek, A.; van der Woning, B.; van der Vliet, H.J.; et al. Nanobody-Fc constructs targeting chemokine receptor CXCR4 potently inhibit signaling and CXCR4-mediated HIV-entry and induce antibody effector functions. Biochem. Pharmacol. 2018, 158, 413–424. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Chen, X.; Nielsen, J.; Petranovic, D.; Siewers, V. Expression of antibody fragments in Saccharomyces cerevisiae strains evolved for enhanced protein secretion. Microb. Cell Fact. 2021, 20, 134. [Google Scholar] [CrossRef]
- Ta, D.T.; Redeker, E.S.; Billen, B.; Reekmans, G.; Sikulu, J.; Noben, J.P.; Guedens, W.; Adriaensens, P. An efficient protocol towards site-specifically clickable nanobodies in high yield: Cytoplasmic expression in Escherichia coli combined with intein-mediated protein ligation. Protein Eng. Des. Sel. PEDS 2015, 28, 351–363. [Google Scholar] [CrossRef]
- Baghban, R.; Gargari, S.L.; Rajabibazl, M.; Nazarian, S.; Bakherad, H. Camelid-derived heavy-chain nanobody against Clostridium botulinum neurotoxin E in Pichia pastoris. Biotechnol. Appl. Biochem. 2016, 63, 200–205. [Google Scholar] [CrossRef]
- Vanmarsenille, C.; Elseviers, J.; Yvanoff, C.; Hassanzadeh-Ghassabeh, G.; Garcia Rodriguez, G.; Martens, E.; Depicker, A.; Martel, A.; Haesebrouck, F.; Pasmans, F.; et al. In planta expression of nanobody-based designer chicken antibodies targeting Campylobacter. PLoS ONE 2018, 13, e0204222. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Su, W.; Li, S. Construction of Synthetic Nanobody Library in Mammalian Cells by dsDNA-Based Strategies. ChemBioChem Eur. J. Chem. Biol. 2021, 22, 2957–2965. [Google Scholar] [CrossRef]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, M.; Van Vaerenbergh, N.; Kulaya, N.B.; Vincke, C.; D’Huyvetter, M.; Devoogdt, N.; Muyldermans, S.; Xavier, C. Reshaping nanobodies for affinity purification on protein a. New Biotechnol. 2020, 57, 20–28. [Google Scholar] [CrossRef]
- Muyldermans, S. A guide to: Generation and design of nanobodies. FEBS J. 2021, 288, 2084–2102. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, K.; Jung, S.; Conte, A.; Lieberman, J.; Muecksch, F.; Lorenzi, J.C.C.; Park, S.; Schmidt, F.; Wang, Z.; et al. Nanobodies from camelid mice and llamas neutralize SARS-CoV-2 variants. Nature 2021, 595, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Janssens, R.; Dekker, S.; Hendriks, R.W.; Panayotou, G.; van Remoortere, A.; San, J.K.; Grosveld, F.; Drabek, D. Generation of heavy-chain-only antibodies in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 15130–15135. [Google Scholar] [CrossRef]
- Teng, Y.; Young, J.L.; Edwards, B.; Hayes, P.; Thompson, L.; Johnston, C.; Edwards, C.; Sanders, Y.; Writer, M.; Pinto, D.; et al. Diverse human VH antibody fragments with bio-therapeutic properties from the Crescendo Mouse. New Biotechnol. 2020, 55, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Shi, W.; Huang, X.; Wan, Y.; Li, G.; Xing, B.; Xu, Z.P.; Liu, H.; Hammock, B.D.; Yang, X.; et al. Screening, Expression, and Identification of Nanobody against SARS-CoV-2 Spike Protein. Cells 2022, 11, 3355. [Google Scholar] [CrossRef]
- Zimmermann, I.; Egloff, P.; Hutter, C.A.J.; Kuhn, B.T.; Bräuer, P.; Newstead, S.; Dawson, R.J.P.; Geertsma, E.R.; Seeger, M.A. Generation of synthetic nanobodies against delicate proteins. Nat. Protoc. 2020, 15, 1707–1741. [Google Scholar] [CrossRef] [PubMed]
- Pardon, E.; Laeremans, T.; Triest, S.; Rasmussen, S.G.; Wohlkönig, A.; Ruf, A.; Muyldermans, S.; Hol, W.G.; Kobilka, B.K.; Steyaert, J. A general protocol for the generation of Nanobodies for structural biology. Nat. Protoc. 2014, 9, 674–693. [Google Scholar] [CrossRef] [PubMed]
- Uchański, T.; Zögg, T.; Yin, J.; Yuan, D.; Wohlkönig, A.; Fischer, B.; Rosenbaum, D.M.; Kobilka, B.K.; Pardon, E.; Steyaert, J. An improved yeast surface display platform for the screening of nanobody immune libraries. Sci. Rep. 2019, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- McMahon, C.; Baier, A.S.; Pascolutti, R.; Wegrecki, M.; Zheng, S.; Ong, J.X.; Erlandson, S.C.; Hilger, D.; Rasmussen, S.G.F.; Ring, A.M.; et al. Yeast surface display platform for rapid discovery of conformationally selective nanobodies. Nat. Struct. Mol. Biol. 2018, 25, 289–296. [Google Scholar] [CrossRef]
- Li, R.; Kang, G.; Hu, M.; Huang, H. Ribosome Display: A Potent Display Technology used for Selecting and Evolving Specific Binders with Desired Properties. Mol. Biotechnol. 2019, 61, 60–71. [Google Scholar] [CrossRef]
- Salema, V.; Fernández, L.Á. Escherichia coli surface display for the selection of nanobodies. Microb. Biotechnol. 2017, 10, 1468–1484. [Google Scholar] [CrossRef]
- Salema, V.; Mañas, C.; Cerdán, L.; Piñero-Lambea, C.; Marín, E.; Roovers, R.C.; Van Bergen En Henegouwen, P.M.; Fernández, L.Á. High affinity nanobodies against human epidermal growth factor receptor selected on cells by E. coli display. mAbs 2016, 8, 1286–1301. [Google Scholar] [CrossRef]
- Cherf, G.M.; Cochran, J.R. Applications of Yeast Surface Display for Protein Engineering. Methods Mol. Biol. (Clifton N.J.) 2015, 1319, 155–175. [Google Scholar] [CrossRef]
- Kariuki, C.K.; Magez, S. Improving the yield of recalcitrant Nanobodies® by simple modifications to the standard protocol. Protein Expr. Purif. 2021, 185, 105906. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, T.; Hara, K.; Umemura, K. Nanobody production can be simplified by direct secretion from Escherichia coli. Protein Expr. Purif. 2020, 170, 105607. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, Y.; Yu, J.; Liu, W.; Li, F.; Xian, M.; Nian, R.; Song, H.; Feng, D. An efficient constitutive expression system for Anti-CEACAM5 nanobody production in the yeast Pichia pastoris. Protein Expr. Purif. 2019, 155, 43–47. [Google Scholar] [CrossRef]
- Ruano-Gallego, D.; Fraile, S.; Gutierrez, C.; Fernández, L.Á. Screening and purification of nanobodies from E. coli culture supernatants using the hemolysin secretion system. Microb. Cell Fact. 2019, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, W.; Li, H.; Wang, X.; Zhao, Y. Capture and purification of an untagged nanobody by mixed weak cation chromatography and cation exchange chromatography. Protein Expr. Purif. 2022, 192, 106030. [Google Scholar] [CrossRef] [PubMed]
- Mabey, D.; Peeling, R.W.; Ustianowski, A.; Perkins, M.D. Diagnostics for the developing world. Nature reviews. Microbiology 2004, 2, 231–240. [Google Scholar] [CrossRef]
- Koczula, K.M.; Gallotta, A. Lateral flow assays. Essays Biochem. 2016, 60, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Pinto Torres, J.E.; Goossens, J.; Ding, J.; Li, Z.; Lu, S.; Vertommen, D.; Naniima, P.; Chen, R.; Muyldermans, S.; Sterckx, Y.G.; et al. Development of a Nanobody-based lateral flow assay to detect active Trypanosoma congolense infections. Sci. Rep. 2018, 8, 9019. [Google Scholar] [CrossRef] [PubMed]
- Vincke, C.; Gutiérrez, C.; Wernery, U.; Devoogdt, N.; Hassanzadeh-Ghassabeh, G.; Muyldermans, S. Generation of single domain antibody fragments derived from camelids and generation of manifold constructs. Methods Mol. Biol. (Clifton N.J.) 2012, 907, 145–176. [Google Scholar] [CrossRef]
- Goossens, J.; Sein, H.; Lu, S.; Radwanska, M.; Muyldermans, S.; Sterckx, Y.G.-J.; Magez, S. Functionalization of gold nanoparticles with nanobodies through physical adsorption. Anal. Methods 2017, 9, 3430–3440. [Google Scholar] [CrossRef]
- Li, Z.; Pinto Torres, J.E.; Goossens, J.; Vertommen, D.; Caljon, G.; Sterckx, Y.G.; Magez, S. An Unbiased Immunization Strategy Results in the Identification of Enolase as a Potential Marker for Nanobody-Based Detection of Trypanosoma evansi. Vaccines 2020, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Odongo, S.; Sterckx, Y.G.; Stijlemans, B.; Pillay, D.; Baltz, T.; Muyldermans, S.; Magez, S. An Anti-proteome Nanobody Library Approach Yields a Specific Immunoassay for Trypanosoma congolense Diagnosis Targeting Glycosomal Aldolase. PLoS Negl. Trop. Dis. 2016, 10, e0004420. [Google Scholar] [CrossRef]
- Pinto, J.; Odongo, S.; Lee, F.; Gaspariunaite, V.; Muyldermans, S.; Magez, S.; Sterckx, Y.G. Structural basis for the high specificity of a Trypanosoma congolense immunoassay targeting glycosomal aldolase. PLoS Negl. Trop. Dis. 2017, 11, e0005932. [Google Scholar] [CrossRef]
- Doerflinger, S.Y.; Tabatabai, J.; Schnitzler, P.; Farah, C.; Rameil, S.; Sander, P.; Koromyslova, A.; Hansman, G.S. Development of a Nanobody-Based Lateral Flow Immunoassay for Detection of Human Norovirus. mSphere 2016, 1, e00219-16. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Duan, M.; Pei, D.; Lin, J.; Wang, L.; Zhou, P.; Yao, W.; Guo, Y.; Li, X.; Tao, L.; et al. Development of novel-nanobody-based lateral-flow immunochromatographic strip test for rapid detection of recombinant human interferon α2b. J. Pharm. Anal. 2022, 12, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Salvador, J.-P.; Vasylieva, N.; Gonzalez-Garcia, I.; Jin, M.; Caster, R.; Siegel, J.B.; Hammock, B.D. Nanobody-based lateral flow immunoassay for the rapid detection of aflatoxin B1 in almond milk. ACS Food Sci. Technol. 2022, 2, 1276–1282. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, X.; Li, D.; Hu, J.; Wan, D.; Zhang, Z.; Hammock, B.D. Development of nanobody-based flow-through dot ELISA and lateral-flow immunoassay for rapid detection of 3-phenoxybenzoic acid. Anal. Methods Adv. Methods Appl. 2021, 13, 1757–1765. [Google Scholar] [CrossRef]
- Yu, S.; Li, Z.; Li, J.; Zhao, S.; Wu, S.; Liu, H.; Bi, X.; Li, D.; Dong, J.; Duan, S.; et al. Generation of Dual functional Nanobody-Nanoluciferase Fusion and its potential in Bioluminescence Enzyme Immunoassay for trace Glypican-3 in Serum. Sens. Actuators. B Chem. 2021, 336, 129717. [Google Scholar] [CrossRef]
- Zhang, W.T.; Liu, T.T.; Wu, M.; Chen, X.C.; Han, L.; Shi, Z.Z.; Li, Y.Y.; Li, X.Y.; Xu, H.X.; Gong, L.K.; et al. Development of a nanobody-based immunoassay for the sensitive detection of fibrinogen-like protein 1. Acta Pharmacol. Sin. 2021, 42, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Vasylieva, N.; Wan, D.; Yin, Z.; Dong, J.; Hammock, B.D. An Ultrasensitive Bioluminescent Enzyme Immunoassay Based on Nanobody/Nanoluciferase Heptamer Fusion for the Detection of Tetrabromobisphenol A in Sediment. Anal. Chem. 2020, 92, 10083–10090. [Google Scholar] [CrossRef]
- Liu, X.; Tang, Z.; Duan, Z.; He, Z.; Shu, M.; Wang, X.; Gee, S.J.; Hammock, B.D.; Xu, Y. Nanobody-based enzyme immunoassay for ochratoxin A in cereal with high resistance to matrix interference. Talanta 2017, 164, 154–158. [Google Scholar] [CrossRef]
- Tu, Z.; Chen, Q.; Li, Y.; Xiong, Y.; Xu, Y.; Hu, N.; Tao, Y. Identification and characterization of species-specific nanobodies for the detection of Listeria monocytogenes in milk. Anal. Biochem. 2016, 493, 1–7. [Google Scholar] [CrossRef]
- Gu, K.; Song, Z.; Zhou, C.; Ma, P.; Li, C.; Lu, Q.; Liao, Z.; Huang, Z.; Tang, Y.; Li, H.; et al. Development of nanobody-horseradish peroxidase-based sandwich ELISA to detect Salmonella Enteritidis in milk and in vivo colonization in chicken. J. Nanobiotechnol. 2022, 20, 167. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, Y.; Gu, J.; Yang, F.; Wu, S.; Zhang, C.; Ji, X.; Lv, H.; Muyldermans, S.; Wang, S. Selection of specific nanobodies to develop an immuno-assay detecting Staphylococcus aureus in milk. Food Chem. 2021, 353, 129481. [Google Scholar] [CrossRef]
- Gelkop, S.; Sobarzo, A.; Brangel, P.; Vincke, C.; Romão, E.; Fedida-Metula, S.; Strom, N.; Ataliba, I.; Mwiine, F.N.; Ochwo, S.; et al. The Development and Validation of a Novel Nanobody-Based Competitive ELISA for the Detection of Foot and Mouth Disease 3ABC Antibodies in Cattle. Front. Vet. Sci. 2018, 5, 250. [Google Scholar] [CrossRef] [PubMed]
- Katz, H.E. Antigen sensing via nanobody-coated transistors. Nat. Biomed. Eng. 2021, 5, 639–640. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Du, J.; Park, K.W.; Chang, L.H.; Strieter, E.R.; Andrew, T.L. Immobilization of Nanobodies with Vapor-Deposited Polymer Encapsulation for Robust Biosensors. ACS Appl. Polym. Mater. 2021, 3, 2561–2567. [Google Scholar] [CrossRef] [PubMed]
- Campuzano, S.; Salema, V.; Moreno-Guzmán, M.; Gamella, M.; Yáñez-Sedeño, P.; Fernández, L.A.; Pingarrón, J.M. Disposable amperometric magnetoimmunosensors using nanobodies as biorecognition element. Determination of fibrinogen in plasma. Biosens. Bioelectron. 2014, 52, 255–260. [Google Scholar] [CrossRef]
- Guo, K.; Wustoni, S.; Koklu, A.; Díaz-Galicia, E.; Moser, M.; Hama, A.; Alqahtani, A.A.; Ahmad, A.N.; Alhamlan, F.S.; Shuaib, M.; et al. Rapid single-molecule detection of COVID-19 and MERS antigens via nanobody-functionalized organic electrochemical transistors. Nat. Biomed. Eng. 2021, 5, 666–677. [Google Scholar] [CrossRef]
- Koklu, A.; Wustoni, S.; Guo, K.; Silva, R.; Salvigni, L.; Hama, A.; Diaz-Galicia, E.; Moser, M.; Marks, A.; McCulloch, I.; et al. Convection Driven Ultrarapid Protein Detection via Nanobody-Functionalized Organic Electrochemical Transistors. Adv. Mater. 2022, 34, e2202972. [Google Scholar] [CrossRef]
- Noltes, M.E.; van Dam, G.M.; Nagengast, W.B.; van der Zaag, P.J.; Slart, R.H.J.A.; Szymanski, W.; Kruijff, S.; Dierckx, R.A.J.O. Let’s embrace optical imaging: A growing branch on the clinical molecular imaging tree. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 4120–4128. [Google Scholar] [CrossRef]
- Oliveira, S.; van Dongen, G.A.M.S.; van Walsum, M.S.; Roovers, R.C.; Stam, J.C.; Mali, W.; van Diest, P.J.; van Bergen en Henegouwen, P.M.P. Rapid visualization of human tumor xenografts through optical imaging with a near-infrared fluorescent anti–epidermal growth factor receptor nanobody. Mol. Imaging 2012, 11. [Google Scholar] [CrossRef]
- Kijanka, M.M.; van Brussel, A.S.; van der Wall, E.; Mali, W.P.; van Diest, P.J.; van Bergen En Henegouwen, P.M.; Oliveira, S. Optical imaging of pre-invasive breast cancer with a combination of VHHs targeting CAIX and HER2 increases contrast and facilitates tumour characterization. EJNMMI Res. 2016, 6, 14. [Google Scholar] [CrossRef]
- Zheng, F.; Luo, S.; Ouyang, Z.; Zhou, J.; Mo, H.; Schoonooghe, S.; Muyldermans, S.; De Baetselier, P.; Raes, G.; Wen, Y. NIRF-Molecular Imaging with Synovial Macrophages-Targeting Vsig4 Nanobody for Disease Monitoring in a Mouse Model of Arthritis. Int. J. Mol. Sci. 2019, 20, 3347. [Google Scholar] [CrossRef]
- Xavier, C.; Vaneycken, I.; D’huyvetter, M.; Heemskerk, J.; Keyaerts, M.; Vincke, C.; Devoogdt, N.; Muyldermans, S.; Lahoutte, T.; Caveliers, V. Synthesis, preclinical validation, dosimetry, and toxicity of 68Ga-NOTA-anti-HER2 Nanobodies for iPET imaging of HER2 receptor expression in cancer. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2013, 54, 776–784. [Google Scholar] [CrossRef]
- Keyaerts, M.; Xavier, C.; Heemskerk, J.; Devoogdt, N.; Everaert, H.; Ackaert, C.; Vanhoeij, M.; Duhoux, F.P.; Gevaert, T.; Simon, P.; et al. Phase I Study of 68Ga-HER2-Nanobody for PET/CT Assessment of HER2 Expression in Breast Carcinoma. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2016, 57, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Keyaerts, M.; Xavier, C.; Everaert, H.; Vaneycken, I.; Fontaine, C.; Decoster, L.; Vanhoeij, M.; Caveliers, V.; Lahoutte, T. Phase II trial of HER2-PET/CT using 68ga-anti-HER2 VHH1 for characterization of HER2 presence in brain metastases of breast cancer patients. Ann. Oncol. 2019, 30, iii25–iii26. [Google Scholar] [CrossRef]
- Xavier, C.; Blykers, A.; Vaneycken, I.; D’Huyvetter, M.; Heemskerk, J.; Lahoutte, T.; Devoogdt, N.; Caveliers, V. (18)F-nanobody for PET imaging of HER2 overexpressing tumors. Nucl. Med. Biol. 2016, 43, 247–252. [Google Scholar] [CrossRef]
- D’Huyvetter, M.; Vincke, C.; Xavier, C.; Aerts, A.; Impens, N.; Baatout, S.; De Raeve, H.; Muyldermans, S.; Caveliers, V.; Devoogdt, N.; et al. Targeted radionuclide therapy with A 177Lu-labeled anti-HER2 nanobody. Theranostics 2014, 4, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Jailkhani, N.; Ingram, J.R.; Rashidian, M.; Rickelt, S.; Tian, C.; Mak, H.; Jiang, Z.; Ploegh, H.L.; Hynes, R.O. Noninvasive imaging of tumor progression, metastasis, and fibrosis using a nanobody targeting the extracellular matrix. Proc. Natl. Acad. Sci. USA 2019, 116, 14181–14190. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Gainkam, L.O.; Caveliers, V.; Vanhove, C.; Keyaerts, M.; De Baetselier, P.; Bossuyt, A.; Revets, H.; Lahoutte, T. SPECT imaging with 99mTc-labeled EGFR-specific nanobody for in vivo monitoring of EGFR expression. Mol. Imaging Biol. 2008, 10, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, Z.; Mohanta, S.; Li, Y.; López Armbruster, N.; Braeuer, M.; Nekolla, S.G.; Habenicht, A.; Sager, H.B.; Raes, G.; Weber, W.; et al. Targeting mannose receptor expression on macrophages in atherosclerotic plaques of apolipoprotein E-knockout mice using 68Ga-NOTA-anti-MMR nanobody: Non-invasive imaging of atherosclerotic plaques. EJNMMI Res. 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Senders, M.L.; Hernot, S.; Carlucci, G.; van de Voort, J.C.; Fay, F.; Calcagno, C.; Tang, J.; Alaarg, A.; Zhao, Y.; Ishino, S.; et al. Nanobody-Facilitated Multiparametric PET/MRI Phenotyping of Atherosclerosis. JACC. Cardiovasc. Imaging 2019, 12, 2015–2026. [Google Scholar] [CrossRef]
- Benmebarek, M.R.; Karches, C.H.; Cadilha, B.L.; Lesch, S.; Endres, S.; Kobold, S. Killing Mechanisms of Chimeric Antigen Receptor (CAR) T Cells. Int. J. Mol. Sci. 2019, 20, 1283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.H.; Liu, J.; Wang, B.Y.; Chen, Y.X.; Cao, X.M.; Yang, Y.; Zhang, Y.L.; Wang, F.X.; Zhang, P.Y.; Lei, B.; et al. A phase 1, open-label study of LCAR-B38M, a chimeric antigen receptor T cell therapy directed against B cell maturation antigen, in patients with relapsed or refractory multiple myeloma. J. Hematol. Oncol. 2018, 11, 141. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.H.; Wang, B.Y.; Chen, L.J.; Fu, W.J.; Xu, J.; Liu, J.; Jin, S.W.; Chen, Y.X.; Cao, X.M.; Yang, Y.; et al. Four-year follow-up of LCAR-B38M in relapsed or refractory multiple myeloma: A phase 1, single-arm, open-label, multicenter study in China (LEGEND-2). J. Hematol. Oncol. 2022, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- Berdeja, J.G.; Madduri, D.; Usmani, S.Z.; Jakubowiak, A.; Agha, M.; Cohen, A.D.; Stewart, A.K.; Hari, P.; Htut, M.; Lesokhin, A.; et al. Ciltacabtagene autoleucel, a B-cell maturation antigen-directed chimeric antigen receptor T-cell therapy in patients with relapsed or refractory multiple myeloma (CARTITUDE-1): A phase 1b/2 open-label study. Lancet 2021, 398, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. FDA approves second BCMA-targeted CAR-T cell therapy. Nat. Reviews. Drug Discov. 2022, 21, 249. [Google Scholar] [CrossRef] [PubMed]
- De Munter, S.; Van Parys, A.; Bral, L.; Ingels, J.; Goetgeluk, G.; Bonte, S.; Pille, M.; Billiet, L.; Weening, K.; Verhee, A.; et al. Rapid and Effective Generation of Nanobody Based CARs using PCR and Gibson Assembly. Int. J. Mol. Sci. 2020, 21, 883. [Google Scholar] [CrossRef] [PubMed]
- Albert, S.; Arndt, C.; Feldmann, A.; Bergmann, R.; Bachmann, D.; Koristka, S.; Ludwig, F.; Ziller-Walter, P.; Kegler, A.; Gärtner, S.; et al. A novel nanobody-based target module for retargeting of T lymphocytes to EGFR-expressing cancer cells via the modular UniCAR platform. Oncoimmunology 2017, 6, e1287246. [Google Scholar] [CrossRef]
- Jamnani, F.R.; Rahbarizadeh, F.; Shokrgozar, M.A.; Mahboudi, F.; Ahmadvand, D.; Sharifzadeh, Z.; Parhamifar, L.; Moghimi, S.M. T cells expressing VHH-directed oligoclonal chimeric HER2 antigen receptors: Towards tumor-directed oligoclonal T cell therapy. Biochim. Biophys. Acta 2014, 1840, 378–386. [Google Scholar] [CrossRef]
- Huet, H.A.; Growney, J.D.; Johnson, J.A.; Li, J.; Bilic, S.; Ostrom, L.; Zafari, M.; Kowal, C.; Yang, G.; Royo, A.; et al. Multivalent nanobodies targeting death receptor 5 elicit superior tumor cell killing through efficient caspase induction. mAbs 2014, 6, 1560–1570. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, H.; Wang, X.; Bai, Y.; Wang, P.; Wu, J.; Jiang, X.; Wang, Y.; Cai, H.; Xu, T.; et al. Structural basis of a novel PD-L1 nanobody for immune checkpoint blockade. Cell Discov. 2017, 3, 17004. [Google Scholar] [CrossRef]
- Behdani, M.; Zeinali, S.; Karimipour, M.; Khanahmad, H.; Schoonooghe, S.; Aslemarz, A.; Seyed, N.; Moazami-Godarzi, R.; Baniahmad, F.; Habibi-Anbouhi, M.; et al. Development of VEGFR2-specific Nanobody Pseudomonas exotoxin A conjugated to provide efficient inhibition of tumor cell growth. New Biotechnol. 2013, 30, 205–209. [Google Scholar] [CrossRef]
- D’Huyvetter, M.; Vos, J.; Caveliers, V.; Vaneycken, I.; Heemskerk, J.; Duhoux, F.P.; Fontaine, C.; Vanhoeij, M.; Windhorst, A.D.; Aa, F.V.; et al. Phase I Trial of 131I-GMIB-Anti-HER2-VHH1, a New Promising Candidate for HER2-Targeted Radionuclide Therapy in Breast Cancer Patients. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2021, 62, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Duggan, S. Caplacizumab: First Global Approval. Drugs 2018, 78, 1639–1642. [Google Scholar] [CrossRef]
- Hollifield, A.L.; Arnall, J.R.; Moore, D.C. Caplacizumab: An anti-von Willebrand factor antibody for the treatment of thrombotic thrombocytopenic purpura. Am. J. Health-Syst. Pharm. AJHP Off. J. Am. Soc. Health-Syst. Pharm. 2020, 77, 1201–1207. [Google Scholar] [CrossRef]
- Keam, S.J. Ozoralizumab: First Approval. Drugs 2023, 83, 87–92. [Google Scholar] [CrossRef]
- Hannon, C.W.; McCourt, C.; Lima, H.C.; Chen, S.; Bennett, C. Interventions for cutaneous disease in systemic lupus erythematosus. Cochrane Database Syst. Rev. 2021, 3, CD007478. [Google Scholar] [CrossRef]
- Ablynx. A Phase IIb Study for ALX-0061 Monotherapy in Subjects with Rheumatoid Arthritis. Identifier NCT02518620. Available online: https://clinicaltrials.gov/ct2/show/NCT02518620 (accessed on 9 February 2023).
- Papp, K.A.; Weinberg, M.A.; Morris, A.; Reich, K. IL17A/F nanobody sonelokimab in patients with plaque psoriasis: A multicentre, randomised, placebo-controlled, phase 2b study. Lancet 2021, 397, 1564–1575. [Google Scholar] [CrossRef] [PubMed]
- Alexion Pharmaceuticals. Safety and Efficacy of ALXN1720 in Adults with Generalized Myasthenia Gravis. Identifier NCT05556096. Available online: https://clinicaltrials.gov/ct2/show/NCT05556096 (accessed on 9 February 2023).
- Merck KGaA. Multiple Ascending Doses (MAD) of Anti-A Disintegrin and Metalloproteinase with Thrombospondin Motifs-5 (Anti-ADAMTS-5) Nanobody in Participants with Knee Osteoarthritis (OA). Identifier NCT03583346. Available online: https://clinicaltrials.gov/ct2/show/NCT03583346 (accessed on 9 February 2023).
- VHsquared Ltd. A Six Week Efficacy, Safety and Tolerability Study of V565 in Crohn’s Disease (HarbOR). Identifier NCT02976129. Available online: https://clinicaltrials.gov/ct2/show/NCT02976129 (accessed on 9 February 2023).
- Ma, L.; Zhu, M.; Li, G.; Gai, J.; Li, Y.; Gu, H.; Qiao, P.; Li, X.; Ji, W.; Zhao, R.; et al. Preclinical development of a long-acting trivalent bispecific nanobody targeting IL-5 for the treatment of eosinophilic asthma. Respir. Res. 2022, 23, 316. [Google Scholar] [CrossRef] [PubMed]
- Sadeghian-Rizi, T.; Behdani, M.; Khanahmad, H.; Sadeghi, H.M.; Jahanian-Najafabadi, A. Generation and Characterization of a Functional Nanobody Against Inflammatory Chemokine CXCL10, as a Novel Strategy for the Treatment of Multiple Sclerosis. CNS Neurol. Disord. Drug Targets 2019, 18, 141–148. [Google Scholar] [CrossRef]
- Sarker, S.A.; Jäkel, M.; Sultana, S.; Alam, N.H.; Bardhan, P.K.; Chisti, M.J.; Salam, M.A.; Theis, W.; Hammarström, L.; Frenken, L.G. Anti-rotavirus protein reduces stool output in infants with diarrhea: A randomized placebo-controlled trial. Gastroenterology 2013, 145, 740–748.e8. [Google Scholar] [CrossRef]
- Cunningham, S.; Piedra, P.A.; Martinon-Torres, F.; Szymanski, H.; Brackeva, B.; Dombrecht, E.; Detalle, L.; Fleurinck, C. RESPIRE study group Nebulised ALX-0171 for respiratory syncytial virus lower respiratory tract infection in hospitalised children: A double-blind, randomised, placebo-controlled, phase 2b trial. Lancet Respir. Med. 2021, 9, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Lumen Bioscience, Inc. LMN-101 in a Campylobacter Human Challenge Model. Identifier NCT04182490. Available online: https://clinicaltrials.gov/ct2/show/NCT04182490 (accessed on 9 February 2023).
- Maeda, R.; Fujita, J.; Konishi, Y.; Kazuma, Y.; Yamazaki, H.; Anzai, I.; Watanabe, T.; Yamaguchi, K.; Kasai, K.; Nagata, K.; et al. A panel of nanobodies recognizing conserved hidden clefts of all SARS-CoV-2 spike variants including Omicron. Commun. Biol. 2022, 5, 669. [Google Scholar] [CrossRef]
- Cunha-Santos, C.; Perdigao, P.R.L.; Martin, F.; Oliveira, J.G.; Cardoso, M.; Manuel, A.; Taveira, N.; Goncalves, J. Inhibition of HIV replication through siRNA carried by CXCR4-targeted chimeric nanobody. Cell. Mol. Life Sci. CMLS 2020, 77, 2859–2870. [Google Scholar] [CrossRef] [PubMed]
- Gaiotto, T.; Ramage, W.; Ball, C.; Risley, P.; Carnell, G.W.; Temperton, N.; Engelhardt, O.G.; Hufton, S.E. Nanobodies mapped to cross-reactive and divergent epitopes on A(H7N9) influenza hemagglutinin using yeast display. Sci. Rep. 2021, 11, 3126. [Google Scholar] [CrossRef]
- King, M.T.; Huh, I.; Shenai, A.; Brooks, T.M.; Brooks, C.L. Structural basis of VHH-mediated neutralization of the food-borne pathogen Listeria monocytogenes. J. Biol. Chem. 2018, 293, 13626–13635. [Google Scholar] [CrossRef]
- Bernedo-Navarro, R.A.; Romão, E.; Yano, T.; Pinto, J.; De Greve, H.; Sterckx, Y.G.; Muyldermans, S. Structural Basis for the Specific Neutralization of Stx2a with a Camelid Single Domain Antibody Fragment. Toxins 2018, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Liu, Z.; Li, X.; Qin, X.; Hong, H.; Zhou, Z.; Pieters, R.J.; Shi, J.; Wu, Z. Nanobody-Based Bispecific Neutralizer for Shiga Toxin-Producing E. coli. ACS Infect. Dis. 2022, 8, 321–329. [Google Scholar] [CrossRef]
- Okello, E.; Moonens, K.; Erume, J.; De Greve, H. Orally Fed Recombinant Lactococcus lactis Displaying Surface Anti-Fimbrial Nanobodies Protects Piglets against Escherichia coli Causing Post-Weaning Diarrhea. Agriculture 2021, 11, 186. [Google Scholar] [CrossRef]
- Caljon, G.; Stijlemans, B.; Saerens, D.; Van Den Abbeele, J.; Muyldermans, S.; Magez, S.; De Baetselier, P. Affinity is an important determinant of the anti-trypanosome activity of nanobodies. PLoS Negl. Trop. Dis. 2012, 6, e1902. [Google Scholar] [CrossRef]
- Hempelmann, A.; Hartleb, L.; van Straaten, M.; Hashemi, H.; Zeelen, J.P.; Bongers, K.; Papavasiliou, F.N.; Engstler, M.; Stebbins, C.E.; Jones, N.G. Nanobody-mediated macromolecular crowding induces membrane fission and remodeling in the African trypanosome. Cell Rep. 2021, 37, 109923. [Google Scholar] [CrossRef]
- Arias, J.L.; Unciti-Broceta, J.D.; Maceira, J.; Del Castillo, T.; Hernández-Quero, J.; Magez, S.; Soriano, M.; García-Salcedo, J.A. Nanobody conjugated PLGA nanoparticles for active targeting of African Trypanosomiasis. J. Control. Release Off. J. Control. Release Soc. 2015, 197, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Unciti-Broceta, J.D.; Arias, J.L.; Maceira, J.; Soriano, M.; Ortiz-González, M.; Hernández-Quero, J.; Muñóz-Torres, M.; de Koning, H.P.; Magez, S.; Garcia-Salcedo, J.A. Specific Cell Targeting Therapy Bypasses Drug Resistance Mechanisms in African Trypanosomiasis. PLoS Pathog. 2015, 11, e1004942. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.H.; Gabriela, M.; Reaksudsan, K.; Dixon, M.W.A.; Chan, L.J.; Adair, A.; Trickey, S.; O’Neill, M.T.; Tan, L.L.; Lopaticki, S.; et al. Nanobodies against Pfs230 block Plasmodium falciparum transmission. Biochem. J. 2022, 479, 2529–2546. [Google Scholar] [CrossRef]
- Hmila, I.; Saerens, D.; Ben Abderrazek, R.; Vincke, C.; Abidi, N.; Benlasfar, Z.; Govaert, J.; El Ayeb, M.; Bouhaouala-Zahar, B.; Muyldermans, S. A bispecific nanobody to provide full protection against lethal scorpion envenoming. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 3479–3489. [Google Scholar] [CrossRef] [PubMed]
- Ezzine, A.; M’hirsi El Adab, S.; Bouhaouala-Zahar, B.; Hmila, I.; Baciou, L.; Marzouki, M.N. Efficient expression of the anti-AahI’ scorpion toxin nanobody under a new functional form in a Pichia pastoris system. Biotechnol. Appl. Biochem. 2012, 59, 15–21. [Google Scholar] [CrossRef]
- Ben Abderrazek, R.; Vincke, C.; Hmila, I.; Saerens, D.; Abidi, N.; El Ayeb, M.; Muyldermans, S.; Bouhaouala-Zahar, B. Development of Cys38 knock-out and humanized version of NbAahII10 nanobody with improved neutralization of AahII scorpion toxin. Protein Eng. Des. Sel. PEDS 2011, 24, 727–735. [Google Scholar] [CrossRef]
- Richard, G.; Meyers, A.J.; McLean, M.D.; Arbabi-Ghahroudi, M.; MacKenzie, R.; Hall, J.C. In vivo neutralization of α-cobratoxin with high-affinity llama single-domain antibodies (VHHs) and a VHH-Fc antibody. PLoS ONE 2013, 8, e69495. [Google Scholar] [CrossRef]
- Bailon Calderon, H.; Yaniro Coronel, V.O.; Cáceres Rey, O.A.; Colque Alave, E.G.; Leiva Duran, W.J.; Padilla Rojas, C.; Montejo Arevalo, H.; García Neyra, D.; Galarza Pérez, M.; Bonilla, C.; et al. Development of Nanobodies Against Hemorrhagic and Myotoxic Components of Bothrops atrox Snake Venom. Front. Immunol. 2020, 11, 655. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, B.-k.; Odongo, S.; Radwanska, M.; Magez, S. NANOBODIES®: A Review of Diagnostic and Therapeutic Applications. Int. J. Mol. Sci. 2023, 24, 5994. https://doi.org/10.3390/ijms24065994
Jin B-k, Odongo S, Radwanska M, Magez S. NANOBODIES®: A Review of Diagnostic and Therapeutic Applications. International Journal of Molecular Sciences. 2023; 24(6):5994. https://doi.org/10.3390/ijms24065994
Chicago/Turabian StyleJin, Bo-kyung, Steven Odongo, Magdalena Radwanska, and Stefan Magez. 2023. "NANOBODIES®: A Review of Diagnostic and Therapeutic Applications" International Journal of Molecular Sciences 24, no. 6: 5994. https://doi.org/10.3390/ijms24065994
APA StyleJin, B. -k., Odongo, S., Radwanska, M., & Magez, S. (2023). NANOBODIES®: A Review of Diagnostic and Therapeutic Applications. International Journal of Molecular Sciences, 24(6), 5994. https://doi.org/10.3390/ijms24065994