
Molecular Docking Identifies 1,8-Cineole (Eucalyptol) as A Novel PPARγ Agonist That Alleviates Colon Inflammation

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
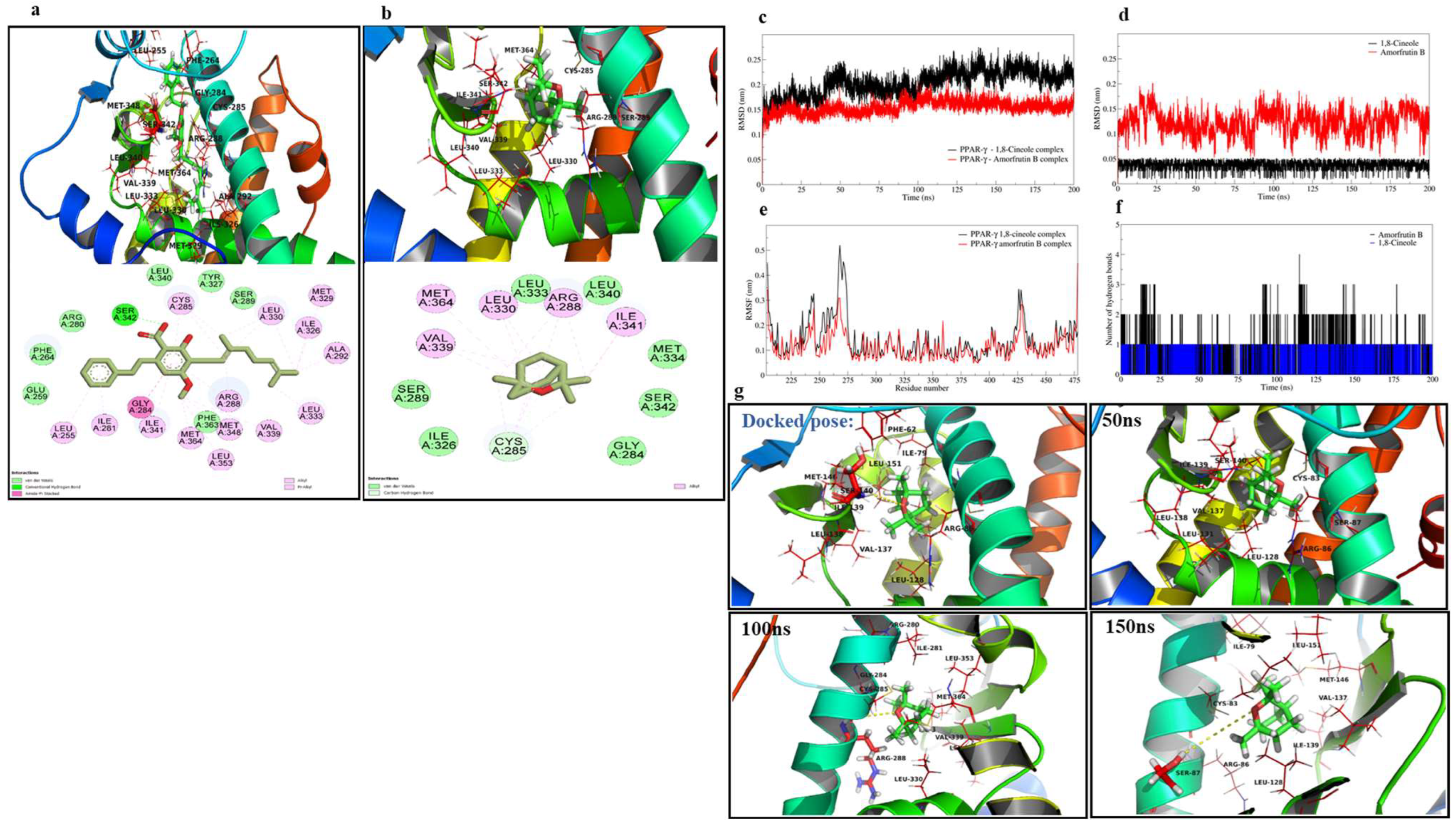
2.1. Molecular Docking Studies
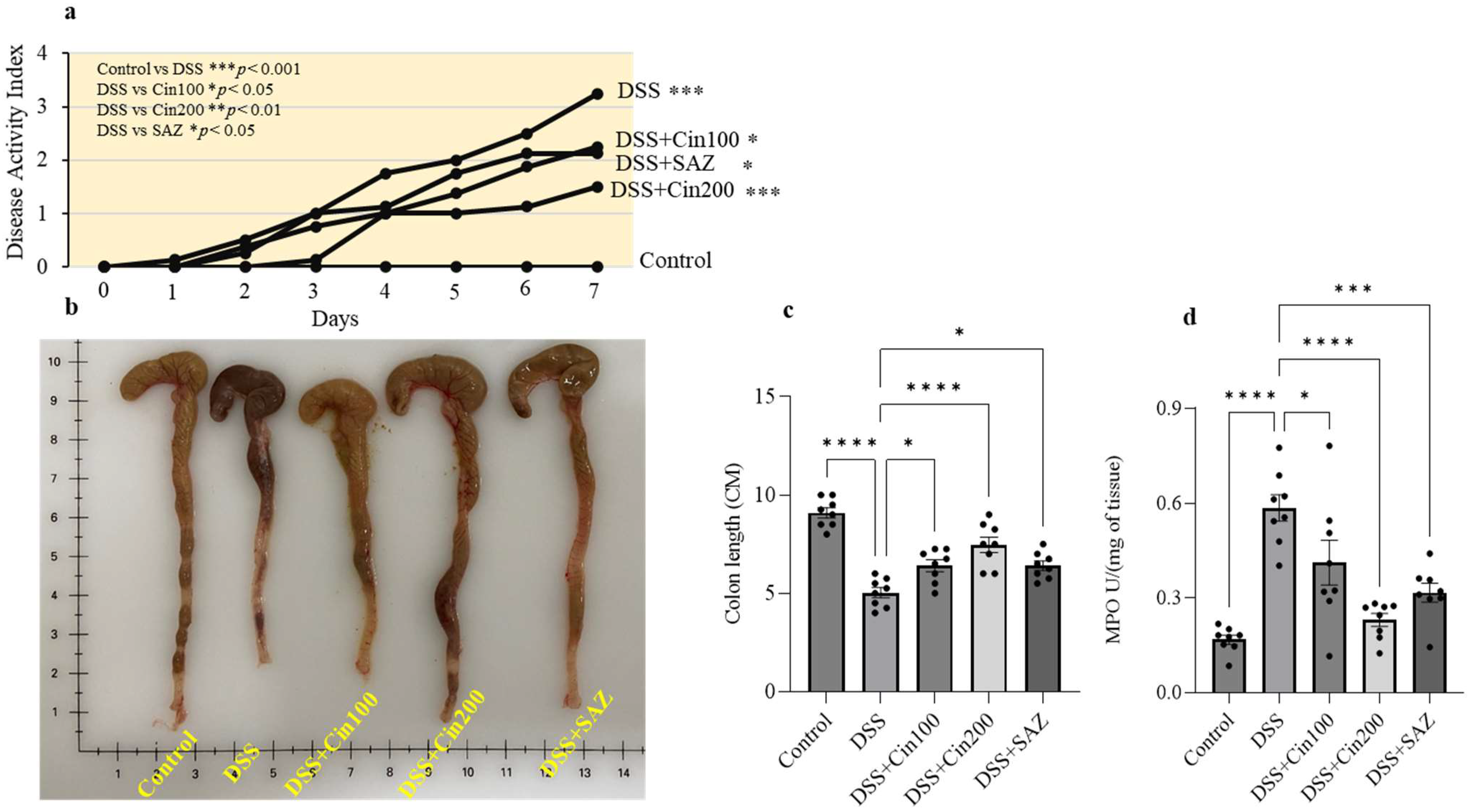
2.2. Effect of 1,8-Cineole on the Disease Activity Index (DAI), Colonic Length, and Myeloperoxidase (MPO) Activity
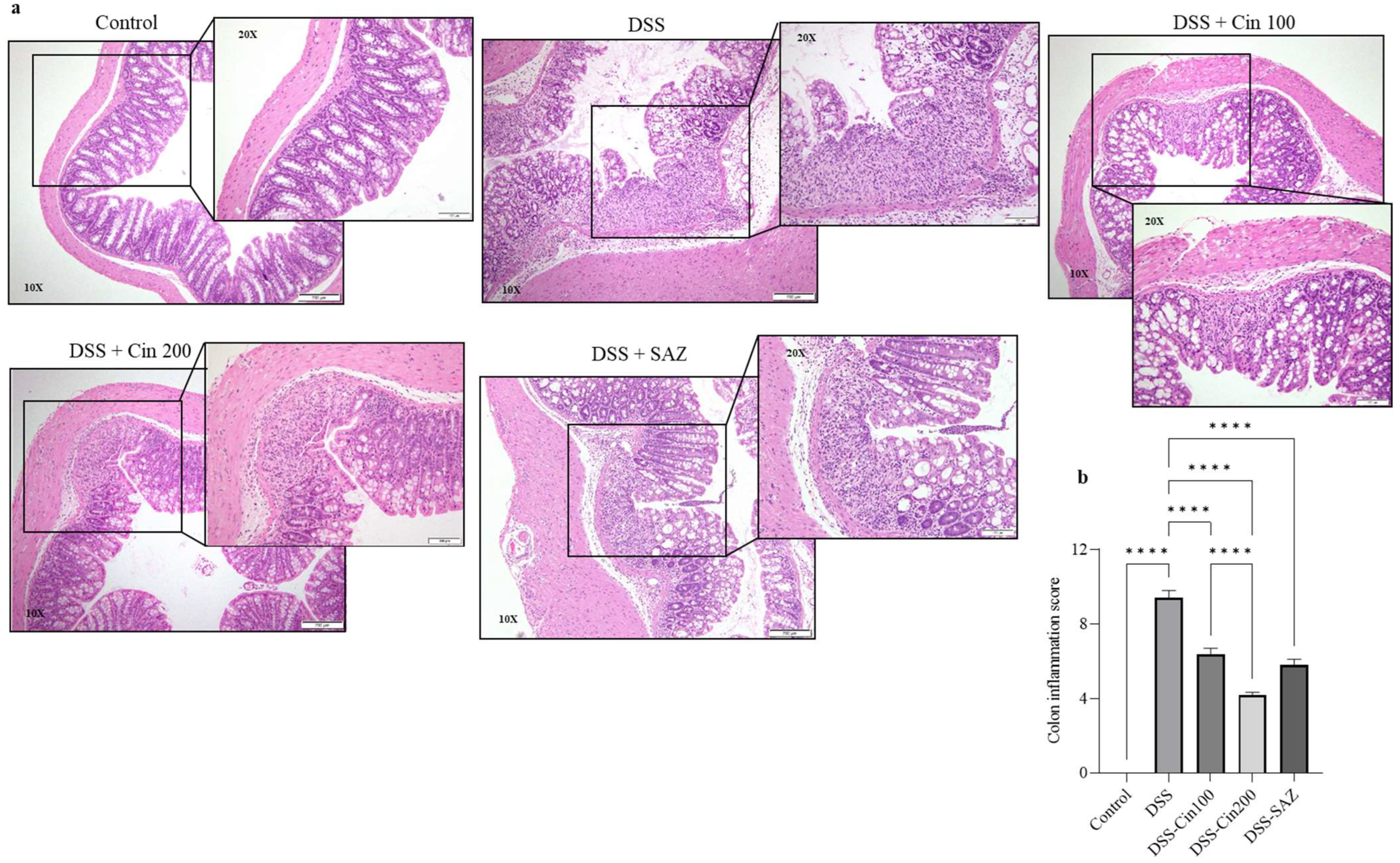
2.3. Effect of 1,8-Cineole on Colonic Histology
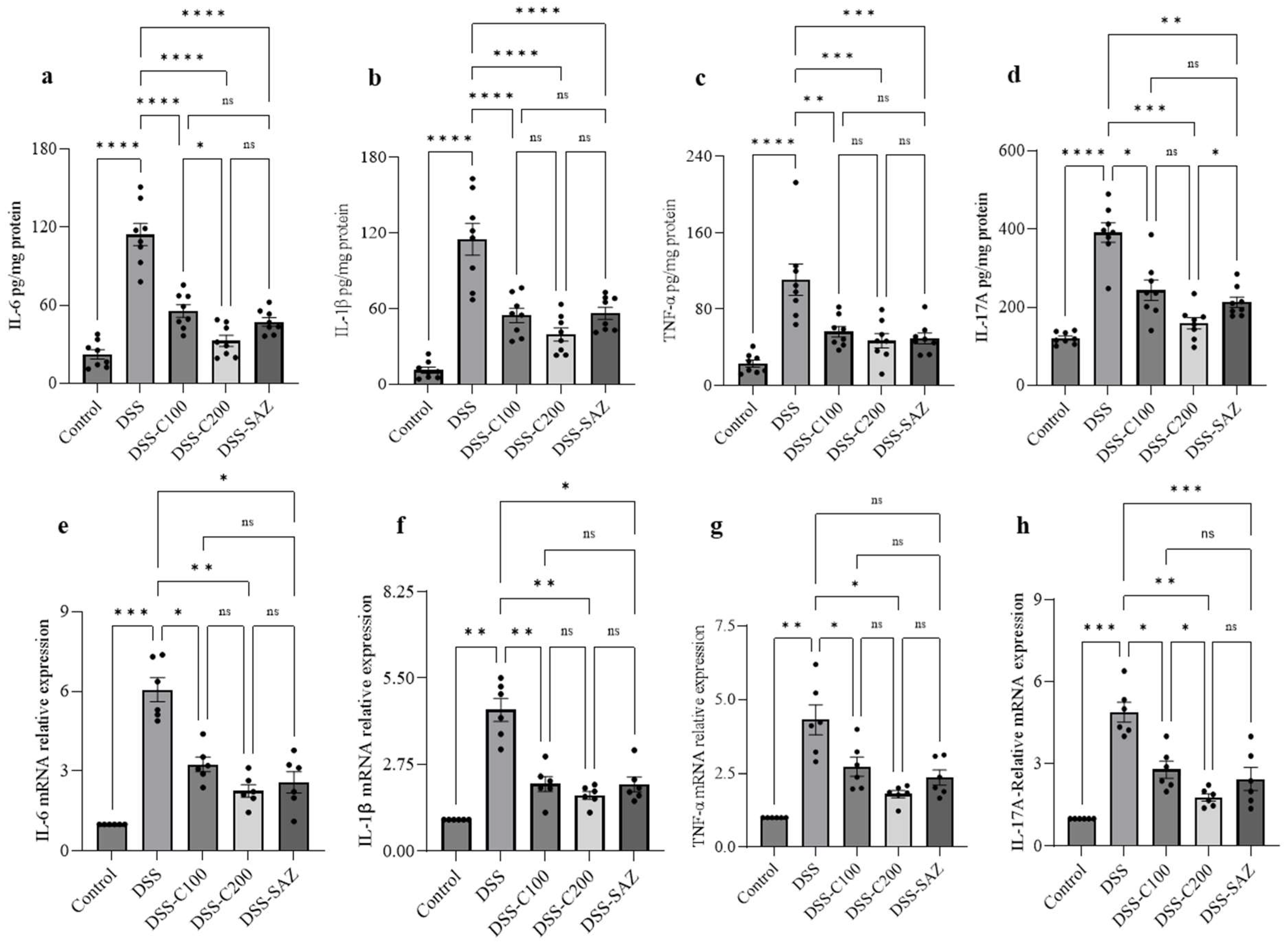
2.4. Effect of 1,8-Cineole on Proinflammatory Cytokine Protein and mRNA
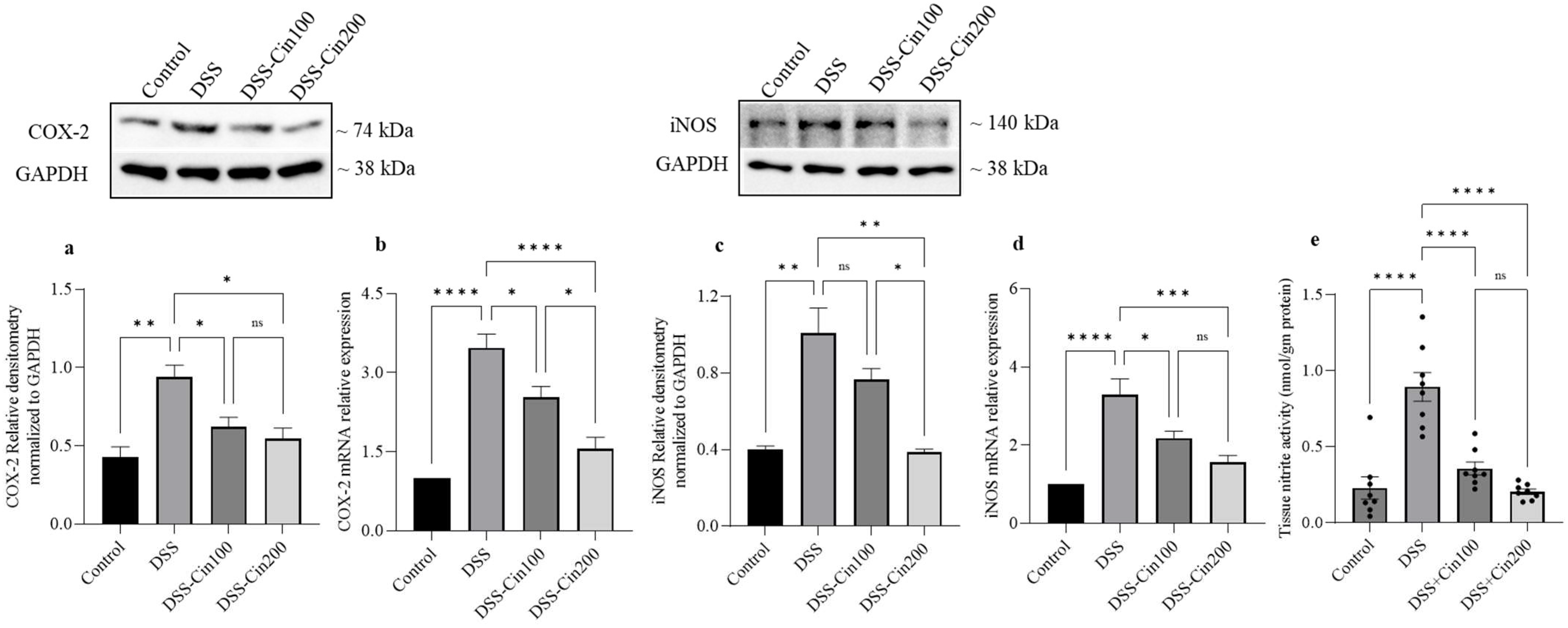
2.5. Effect of 1,8-Cineole on COX2 and iNOS Protein, mRNA, and Tissue Nitrite Levels
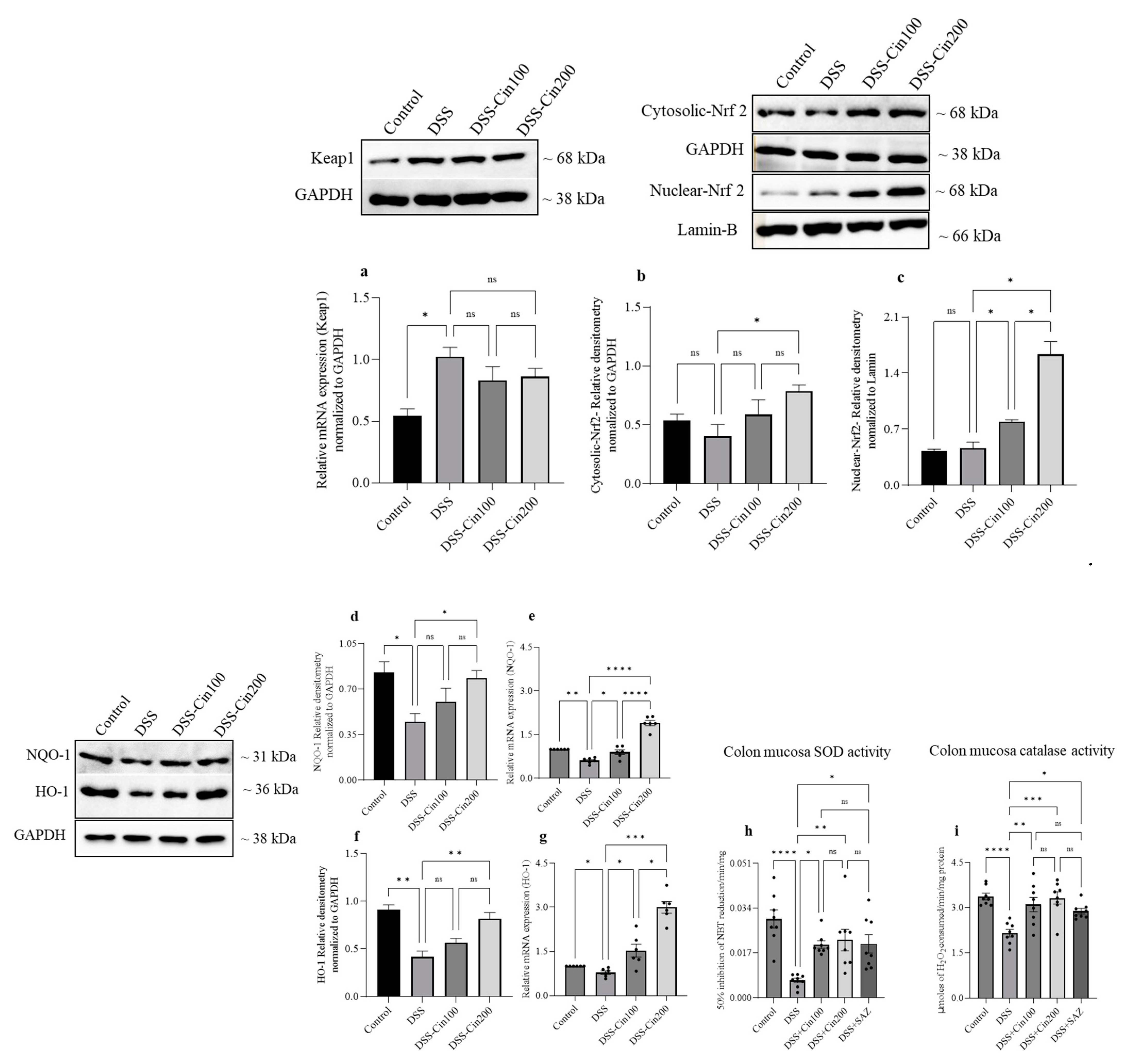
2.6. Effect of 1,8-Cineole on Keap1 and Nrf2 System and Antioxidant Enzymes
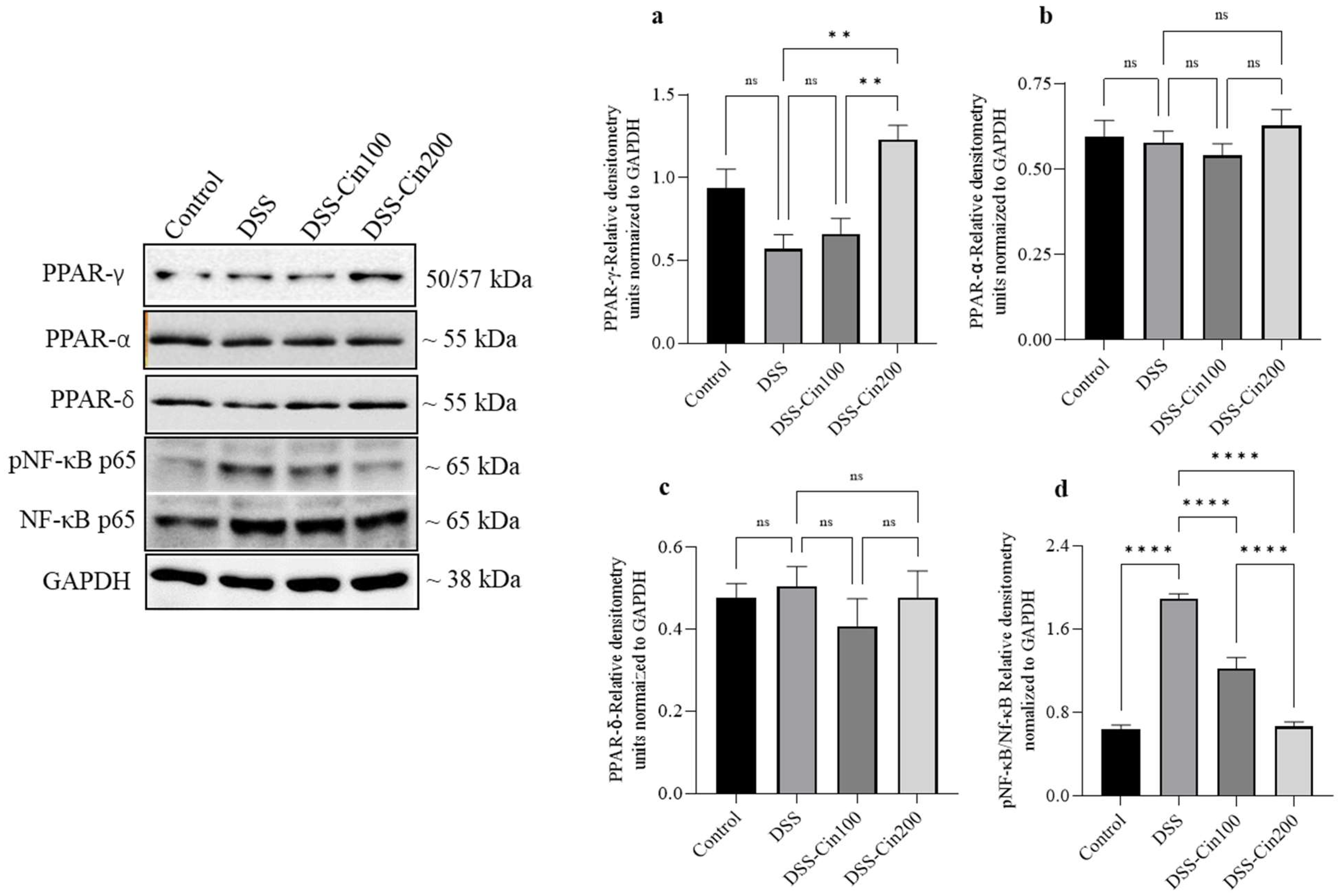
2.7. Effect of 1,8-Cineole on PPARγ, PPARα, PPARβ/δ, and Phospho-Nf-κB (p65) Protein Expression
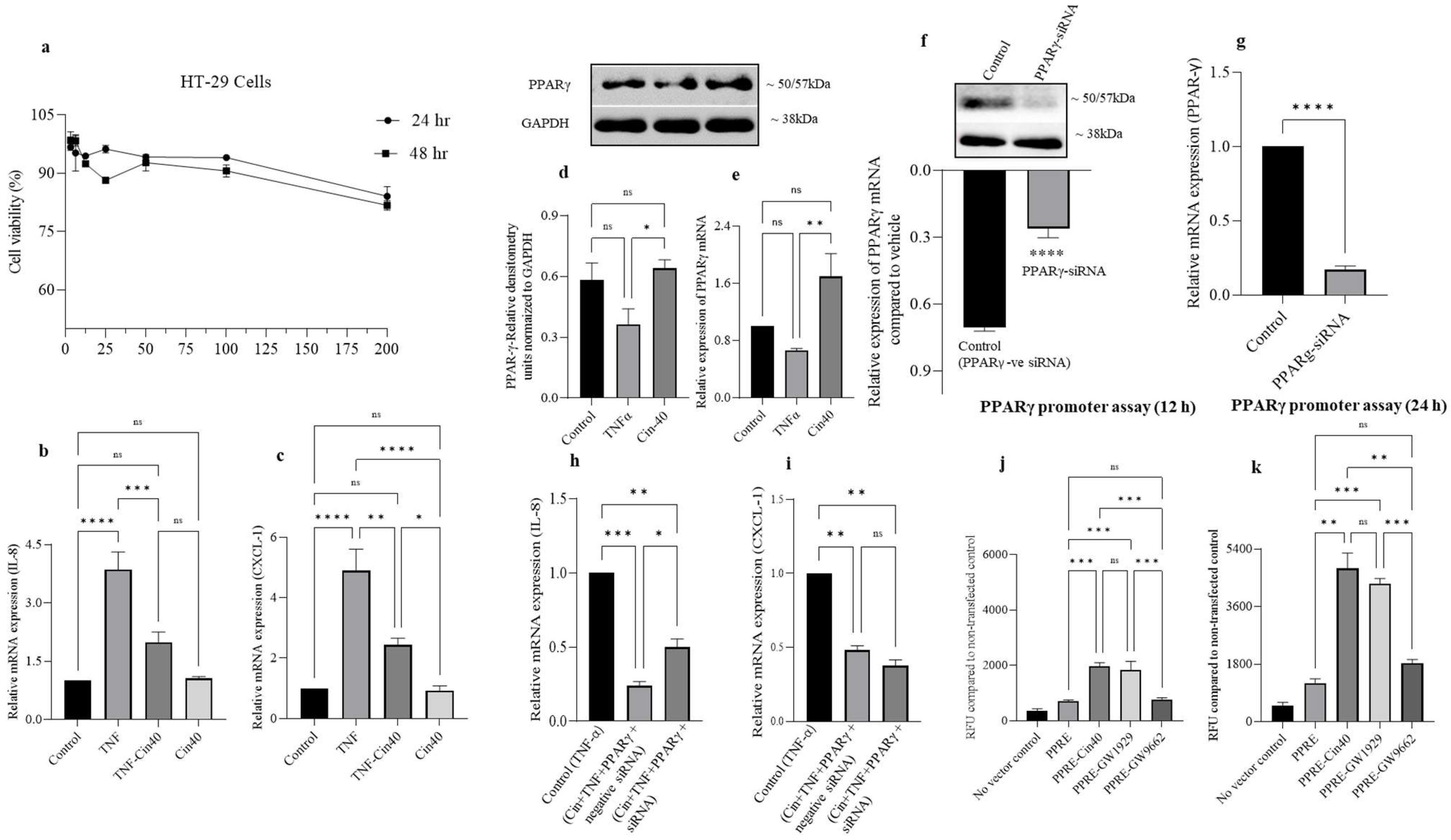
2.8. Effect of 1,8-Ccineole on HT-29 Cell Viability, IL-8, CXCL-1 Chemokine mRNA Expression, and PPARγ Promoter Activation
3. Discussion
4. Materials and Methods
4.1. Molecular Docking and Dynamics Studies
4.2. Chemicals, Reagents, and Cells
4.3. Animals
4.4. Experimental Design
4.5. Evaluation of Disease Activity Index (DAI) Score
4.6. Proinflammatory Cytokine Measurement by Enzyme-linked Immunosorbent Assay (ELISA)
4.7. Myeloperoxidase (MPO) Assay
4.8. Histopathological Evaluation
4.9. RNA Extraction and Real-Time RT-PCR
4.10. Western Blot
4.11. Measurement of Superoxide Dismutase, Catalase Enzyme Activity, and Tissue Nitrite Concentration
4.12. HT-29 Cell Culture
4.13. Cell Viability Assay
4.14. PPARγ Promotor and Nanoluciferase Assay
4.15. PPARγ Knockdown Mediated by siRNA
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDowell, C.; Farooq, U.; Haseeb, M. Inflammatory bowel disease. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470312/ (accessed on 1 January 2023).
- Decara, J.; Rivera, P.; López-Gambero, A.J.; Serrano, A.; Pavón, F.J.; Baixeras, E.; De Fonseca, F.R.; Suárez, J. Peroxisome proliferator-activated receptors: Experimental targeting for the treatment of inflammatory bowel diseases. Front. Pharmacol. 2020, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Fajas, L.; Fruchart, J.C.; Auwerx, J. PPARgamma3 mRNA: A distinct PPARgamma mRNA subtype transcribed from an independent promoter. FEBS Lett. 1998, 438, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Quiles, M.; Broekema, M.F.; Kalkhoven, E. PPARgamma in metabolism, immunity, and cancer: Unified and diverse mechanisms of action. Front. Endocrinol. 2021, 12, 624112. [Google Scholar] [CrossRef]
- Krönke, G.; Kadl, A.; Ikonomu, E.; Blü;ml, S.; Fürnkranz, A.; Sarembock, I.J.; Bochkov, V.N.; Exner, M.; Binder, B.R.; Leitinger, N. Expression of heme oxygenase-1 in human vascular cells is regulated by peroxisome proliferator-activated receptors. Arter. Thromb. Vasc. Biol. 2007, 27, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Dethlefsen, U. Efficacy of cineole in patients suffering from acute bronchitis: A placebo-controlled double-blind trial. Cough 2013, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Worth, H.; Schacher, C.; Dethlefsen, U. Concomitant therapy with Cineole (Eucalyptole) reduces exacerbations in COPD: A placebo-controlled double-blind trial. Respir. Res. 2009, 10, 69. [Google Scholar] [CrossRef] [Green Version]
- Linghu, K.-G.; Wu, G.-P.; Fu, L.-Y.; Yang, H.; Li, H.-Z.; Chen, Y.; Yu, H.; Tao, L.; Shen, X.-C. 1,8-Cineole ameliorates LPS-induced vascular endothelium dysfunction in mice via PPAR-gamma dependent regulation of NF-kappaB. Front. Pharmacol. 2019, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.A.; Santos, F.A.; Silva, R.M.; Campos, A.R.; De Araujo, R.P.; Júnior, R.L.; Rao, V.S.N. 1,8-cineole (eucalyptol), a monoterpene oxide attenuates the colonic damage in rats on acute TNBS-colitis. Food Chem. Toxicol. 2004, 42, 579–584. [Google Scholar] [CrossRef]
- Murad, H.A.; Abdallah, H.M.; Ali, S.S. Mentha longifolia protects against acetic-acid induced colitis in rats. J. Ethnopharmacol. 2016, 190, 354–361. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, J.; Fang, C.; Tang, F. 1,8-cineol attenuates LPS-induced acute pulmonary inflammation in mice. Inflammation 2013, 37, 566–572. [Google Scholar] [CrossRef]
- Juergens, U.R.; Stöber, M.; Vetter, H. Inhibition of cytokine production and arachidonic acid metabolism by eucalyptol (1.8-cineole) in human blood monocytes in vitro. Eur. J. Med. Res. 1998, 3, 508–510. [Google Scholar] [PubMed]
- Yadav, N.; Chandra, H. Suppression of inflammatory and infection responses in lung macrophages by eucalyptus oil and its constituent 1,8-cineole: Role of pattern recognition receptors TREM-1 and NLRP3, the MAP kinase regulator MKP-1, and NFkappaB. PLoS ONE 2017, 12, e0188232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Wu, G.; Li, W.; Yang, J.; Yan, J.; Wang, Y.; Yao, W.; Zhou, X.; He, Z.; Wu, L.; et al. Preparation and protective effects of 1,8-cineole-loaded self-microemulsifying drug delivery system on lipopolysaccharide-induced endothelial injury in mice. Eur. J. Pharm. Sci. 2019, 127, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, Z.; Gulaboglu, M.; Halıcı, Z.; Cınar, I.; Dıyarbakır, B. Biochemical research of the effects of essential oil obtained from the fruit of Myrtus communis L. on cell damage associated with lipopolysaccharide-induced endotoxemia in a human umbilical cord vein endothelial cells. Biochem. Genet. 2021, 59, 315–334. [Google Scholar]
- Juergens, U.; Dethlefsen, U.; Steinkamp, G.; Gillissen, A.; Repges, R.; Vetter, H. Anti-inflammatory activity of 1.8-cineol (eucalyptol) in bronchial asthma: A double-blind placebo-controlled trial. Respir. Med. 2003, 97, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Fujino, S.; Andoh, A.; Bamba, S.; Ogawa, A.; Hata, K.; Araki, Y.; Bamba, T.; Fujiyama, Y. Increased expression of interleukin 17 in inflammatory bowel disease. Gut 2003, 52, 65–70. [Google Scholar] [CrossRef]
- Ito, R.; Kita, M.; Shin-Ya, M.; Kishida, T.; Urano, A.; Takada, R.; Sakagami, J.; Imanishi, J.; Iwakura, Y.; Okanoue, T.; et al. Involvement of IL-17A in the pathogenesis of DSS-induced colitis in mice. Biochem. Biophys. Res. Commun. 2008, 377, 12–16. [Google Scholar] [CrossRef]
- Lee, H.-S.; Park, D.-E.; Song, W.-J.; Park, H.-W.; Kang, H.-R.; Cho, S.-H.; Sohn, S.-W. Effect of 1.8-cineole in dermatophagoides pteronyssinus-stimulated bronchial epithelial cells and mouse model of asthma. Biol. Pharm. Bull. 2016, 39, 946–952. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Wu, P.-C.; Tsao, N.-W.; Tseng, Y.-H.; Chu, F.-H.; Wang, S.-Y. Anti-inflammatory activities of constituents from Cinnamomum insularimontanum hayata leaves and their mechanisms. Plants 2022, 11, 3252. [Google Scholar] [CrossRef]
- Zuzarte, M.; Francisco, V.; Neves, B.; Liberal, J.; Cavaleiro, C.; Canhoto, J.; Salgueiro, L.; Cruz, M.T. Lavandula viridis L Her. essential oil inhibits the inflammatory response in macrophages through blockade of NF-KB signaling cascade. Front. Pharmacol. 2021, 12, 695911. [Google Scholar]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription factor for stress response and beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Song, M.Y.; Kim, E.H. Role of oxidative stress and Nrf2/KEAP1 signaling in colorectal cancer: Mechanisms and Therapeutic perspectives with phytochemicals. Antioxidants 2021, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hernández, A.; Córdova, E.J.; Rosillo-Salazar, O.; García-Ortíz, H.; Contreras-Cubas, C.; Islas-Andrade, S.; Revilla-Monsalve, C.; Salas-Labadía, C.; Orozco, L. Association of HMOX1 and NQO1 polymorphisms with metabolic syndrome components. PLoS ONE 2015, 10, e0123313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryter, S.W. Heme oxygenase-1: An Anti-inflammatory effector in cardiovascular, lung, and related metabolic disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Long, F.; Lin, H.; Wang, S.; Wang, T. Dietary phytochemicals targeting Nrf2 to enhance the radiosensitivity of cancer. Oxidative Med. Cell. Longev. 2022, 2022, 7848811. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tao, D.; Yu, F.; Wang, T.; Qi, M.; Xu, S. Cineole regulates Wnt/beta-catenin pathway through Nrf2/keap1/ROS to inhibit bisphenol A-induced apoptosis, autophagy inhibition and immunosuppression of grass carp hepatocytes. Fish Shellfish Immunol. 2022, 131, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Guo, X.; Zhang, K.; Sekaran, G.; Cao, B.; Zhao, Q.; Zhang, S.; Kirby, G.M.; Zhang, X. The essential oils and eucalyptol from Artemisia vulgaris L. prevent acetaminophen-induced liver injury by activating Nrf2-Keap1 and enhancing APAP clearance through non-toxic metabolic pathway. Front. Pharmacol. 2019, 10, 782. [Google Scholar] [CrossRef] [Green Version]
- Porres-Martínez, M.; González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. In vitro neuroprotective potential of the monoterpenes alpha-pinene and 1,8-cineole against H2O2-induced oxidative stress in PC12 cells. Z. Nat. C J. Biosci. 2016, 71, 191–199. [Google Scholar]
- Annese, V.; Rogai, F.; Settesoldi, A.; Bagnoli, S. PPARgamma in inflammatory bowel disease. PPAR Res. 2012, 2012, 620839. [Google Scholar] [CrossRef] [Green Version]
- Venkataraman, B.; Almarzooqi, S.; Raj, V.; Alhassani, A.T.; Alhassani, A.S.; Ahmed, K.J.; Subramanian, V.S.; Ojha, S.K.; Attoub, S.; Adrian, T.E.; et al. Thymoquinone, a dietary bioactive compound, exerts anti-inflammatory effects in colitis by stimulating expression of the colonic epithelial PPAR-gamma transcription factor. Nutrients 2021, 13, 1343. [Google Scholar] [CrossRef]
- Venkataraman, B.; Almarzooqi, S.; Raj, V.; Dudeja, P.K.; Bhongade, B.A.; Patil, R.B.; Ojha, S.K.; Attoub, S.; Adrian, T.E.; Subramanya, S.B. alpha-bisabolol mitigates colon inflammation by stimulating colon PPAR-gamma transcription factor: In Vivo and in vitro study. PPAR Res. 2022, 2022, 5498115. [Google Scholar] [CrossRef] [PubMed]
- Atreya, I.; Atreya, R.; Neurath, M.F. NF-kappaB in inflammatory bowel disease. J. Intern. Med. 2008, 263, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Dubuquoy, L.; Rousseaux, C.; Thuru, X.; Peyrin-Biroulet, L.; Romano, O.; Chavatte, P.; Chamaillard, M.; Desreumaux, P. PPARgamma as a new therapeutic target in inflammatory bowel diseases. Gut 2006, 55, 1341–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straus, D.S.; Glass, C.K. Anti-inflammatory actions of PPAR ligands: New insights on cellular and molecular mechanisms. Trends Immunol. 2007, 28, 551–558. [Google Scholar] [CrossRef]
- Carothers, A.M.; Davids, J.S.; Damas, B.C.; Bertagnolli, M.M. Persistent cyclooxygenase-2 inhibition downregulates NF-kappaB, resulting in chronic intestinal inflammation in the min/+ mouse model of colon tumorigenesis. Cancer Res. 2010, 70, 4433–4442. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Reynolds, J.M.; Stout, R.D.; Bernlohr, D.A.; Suttles, J. Regulation of Th17 differentiation by epidermal fatty acid-binding protein. J. Immunol. 2009, 182, 7625–7633. [Google Scholar] [CrossRef] [Green Version]
- Ricote, M.; Glass, C.K. PPARs and molecular mechanisms of transrepression. Biochim. Biophys Acta 2007, 1771, 926–935. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.H.; Yang, X.Y.; Zhang, X.; Huang, J.; Hou, J.; Li, J.; Xiong, H.; Mihalic, K.; Zhu, H.; Xiao, W.; et al. Transcriptional inactivation of STAT3 by PPARgamma suppresses IL-6-responsive multiple myeloma cells. Immunity 2004, 20, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Welch, J.S.; Ricote, M.; Akiyama, T.E.; Gonzalez, F.J.; Glass, C.K. PPARgamma and PPARdelta negatively regulate specific subsets of lipopolysaccharide and IFN-gamma target genes in macrophages. Proc. Natl. Acad. Sci. USA 2003, 100, 6712–6717. [Google Scholar] [CrossRef] [Green Version]
- Puleston, J.; Cooper, M.; Murch, S.; Bid, K.; Makh, S.; Ashwood, P.; Bingham, A.H.; Green, H.; Moss, P.; Dhillon, A.; et al. A distinct subset of chemokines dominates the mucosal chemokine response in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2005, 21, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Ponce de Leon-Rodriguez, M.D.C.; Guyot, J.P.; Laurent-Babot, C. Intestinal in vitro cell culture models and their potential to study the effect of food components on intestinal inflammation. Crit. Rev. Food Sci. Nutr. 2019, 59, 3648–3666. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, T.; Haniu, H. Peroxisome proliferator-activated receptor gamma overexpression suppresses proliferation of human colon cancer cells. Biochem. Biophys. Res. Commun. 2012, 424, 524–529. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Van der Spoel, D.; Van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Subramanya, S.B.; Chandran, S.; Almarzooqi, S.; Raj, V.; Al Zahmi, A.S.; Al Katheeri, R.A.; Al Zadjali, S.A.; Collin, P.D.; Adrian, T.E. Frondanol, a nutraceutical extract from cucumaria frondosa, attenuates colonic inflammation in a DSS-induced colitis model in mice. Mar. Drugs 2018, 16, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Raj, V.; Venkataraman, B.; Almarzooqi, S.; Chandran, S.; Ojha, S.K.; Attoub, S.; Adrian, T.E.; Subramanya, S.B. Nerolidol mitigates colonic inflammation: An experimental study using both in vivo and in vitro models. Nutrients 2020, 12, 2032. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar] [PubMed]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.-C.; Wu, H.-W.; Lin, Y.-J.; Chang, S.-F. The essential role of Oct-2 in LPS-induced expression of iNOS in RAW 264.7 macrophages and its regulation by trichostatin A. Am. J. Physiol. Physiol. 2009, 296, C1133–C1139. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Binding Free Energy Estimate (kcal/mol) | Interactions | |
|---|---|---|---|
| H-Bond Interactions | Hydrophobic Interactions | ||
| 1,8-Cineole | −5.7 | - | Ser342, Cys285, Ile341, Leu340, Leu333, Met363, Ser289, Arg288, Val339, and Leu330 |
| Amorfrutin B | −6.9 | Ser342 | Ser342, Ile341, Gly284, Leu255, Ile281, Arg288, Met329, Leu330, Ile326, Ala292, Leu333, Met348, Leu353, Val339, Met364, and Cys285 |
| Gene | Forward | Reverse | PMID |
|---|---|---|---|
| Mouse IL-6 | 5′-TGTGTCGTGCTGTTCAGAACC-3′ | 5′-AGGAATCCCGCAATGATGG-3′ | 22326488 |
| Mouse Il-1β | 5′-TCGCTCAGGGTCACAAGAAA-3′ | 5′-CATCAGAGGCAAGGAGGAAAC-3′ | 21735552 |
| Mouse TNF-α | 5′-AGGCTGCCCCGACTACGT-3′ | 5′-GACTTTCTCCTGGTATGAGATAGCAAA-3′ | 21705622 |
| Mouse IL-17A | 5′-ATCCCTCAAAGCTCAGCGTGTC-3′ | 5′-GGGTCTTCATTGCGGTGGAGAG-3′ | 18606690 |
| Mouse COX2 | 5′-AACCGCATTGCCTCTGAAT-3′ | 5′-CATGTTCCAGGAGGATGGAG-3′ | 22158945 |
| Mouse iNOS | 5′-CGAAACGCTTCACTTCCAA-3′ | 5′-TGAGCCTATATTGCTGTGGCT-3′ | 22158945 |
| Mouse 18S | 5′-CCCCTCGATGACTTTAGCTGAGTGT-3′ | 5′-CGCCGGTCCAAGAATTTCACCTCT-3′ | 22427817 |
| Human CXCL-1 | 5′-GCGGAAAGCTTGCCTCAATC-3′ | 5′-GGTCAGTTGGATTTGTCACTGT-3′ | 25938459 |
| Human IL-8 | 5′-ACTGAGAGTGATTGAGAGTGGAC-3′ | 5′-AACCCTCTGCACCCAGTTTTC-3′ | 31273598 |
| Human PPARγ | 5′-TTCAAGAGTACCAAAGTGCAATCAA-3′ | 5′-AATAAGGTGGAGATGCAGGCTC-3′ | 20421464 |
| Human GAPDH | 5′-TCAAGGCTGAGAACGGGAAG-3′ | 5′-CGCCCCACTTGATTTTGGAG-3′ | 33575432 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venkataraman, B.; Almarzooqi, S.; Raj, V.; Bhongade, B.A.; Patil, R.B.; Subramanian, V.S.; Attoub, S.; Rizvi, T.A.; Adrian, T.E.; Subramanya, S.B. Molecular Docking Identifies 1,8-Cineole (Eucalyptol) as A Novel PPARγ Agonist That Alleviates Colon Inflammation. Int. J. Mol. Sci. 2023, 24, 6160. https://doi.org/10.3390/ijms24076160
Venkataraman B, Almarzooqi S, Raj V, Bhongade BA, Patil RB, Subramanian VS, Attoub S, Rizvi TA, Adrian TE, Subramanya SB. Molecular Docking Identifies 1,8-Cineole (Eucalyptol) as A Novel PPARγ Agonist That Alleviates Colon Inflammation. International Journal of Molecular Sciences. 2023; 24(7):6160. https://doi.org/10.3390/ijms24076160
Chicago/Turabian StyleVenkataraman, Balaji, Saeeda Almarzooqi, Vishnu Raj, Bhoomendra A. Bhongade, Rajesh B. Patil, Veedamali S. Subramanian, Samir Attoub, Tahir A. Rizvi, Thomas E. Adrian, and Sandeep B. Subramanya. 2023. "Molecular Docking Identifies 1,8-Cineole (Eucalyptol) as A Novel PPARγ Agonist That Alleviates Colon Inflammation" International Journal of Molecular Sciences 24, no. 7: 6160. https://doi.org/10.3390/ijms24076160
APA StyleVenkataraman, B., Almarzooqi, S., Raj, V., Bhongade, B. A., Patil, R. B., Subramanian, V. S., Attoub, S., Rizvi, T. A., Adrian, T. E., & Subramanya, S. B. (2023). Molecular Docking Identifies 1,8-Cineole (Eucalyptol) as A Novel PPARγ Agonist That Alleviates Colon Inflammation. International Journal of Molecular Sciences, 24(7), 6160. https://doi.org/10.3390/ijms24076160