
Effect of Electric Field on α-Synuclein Fibrils: Revealed by Molecular Dynamics Simulations

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
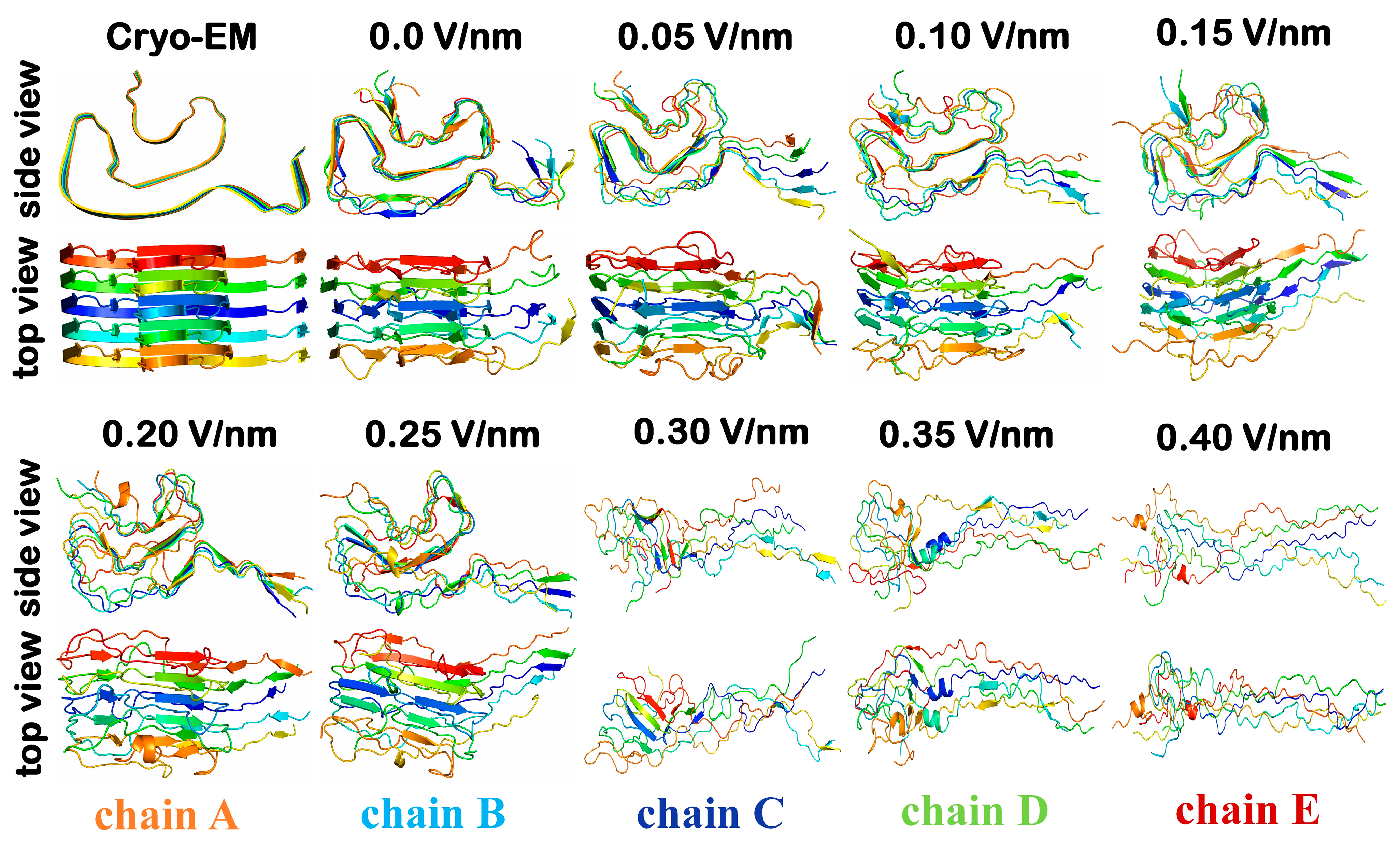
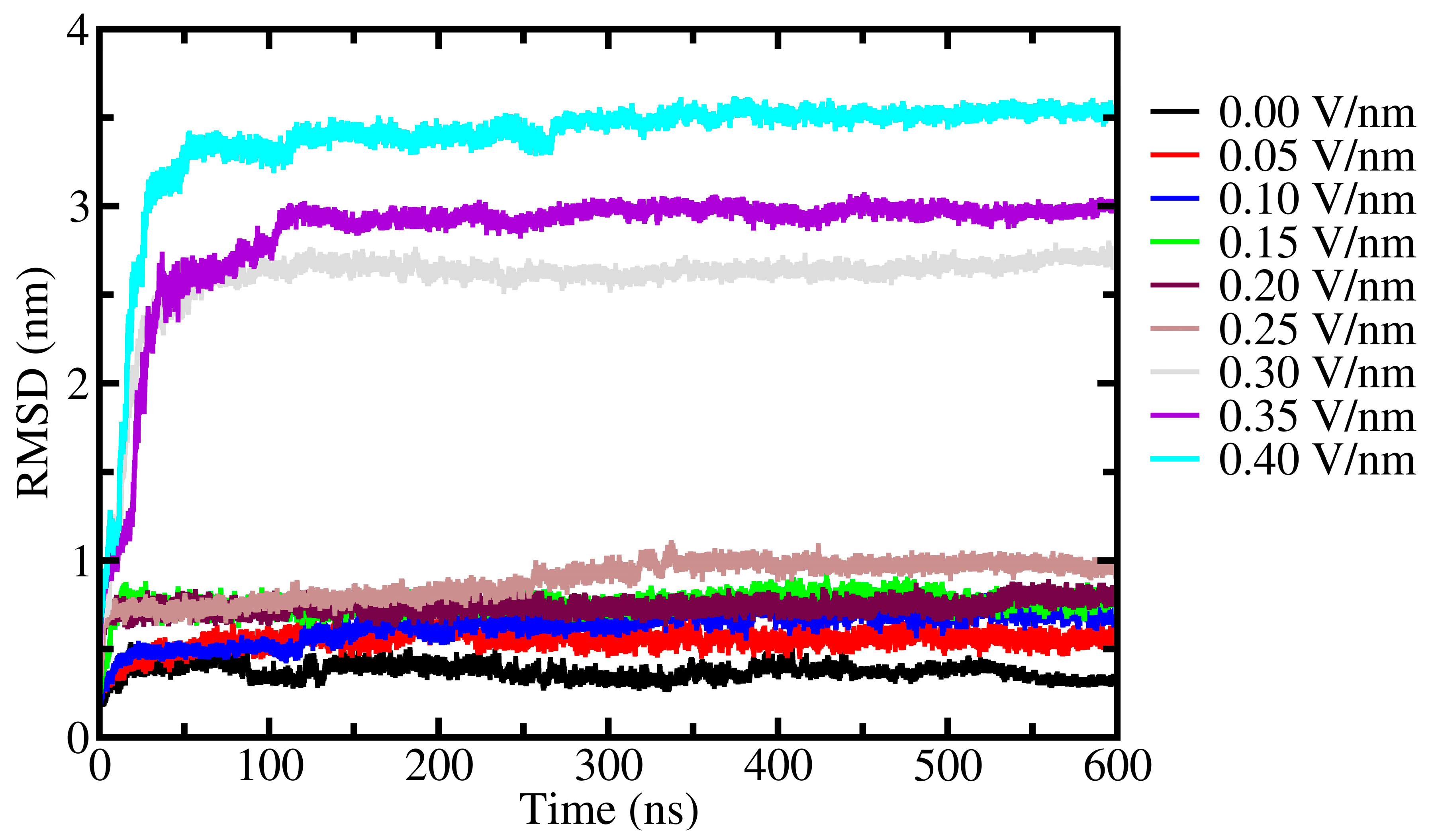
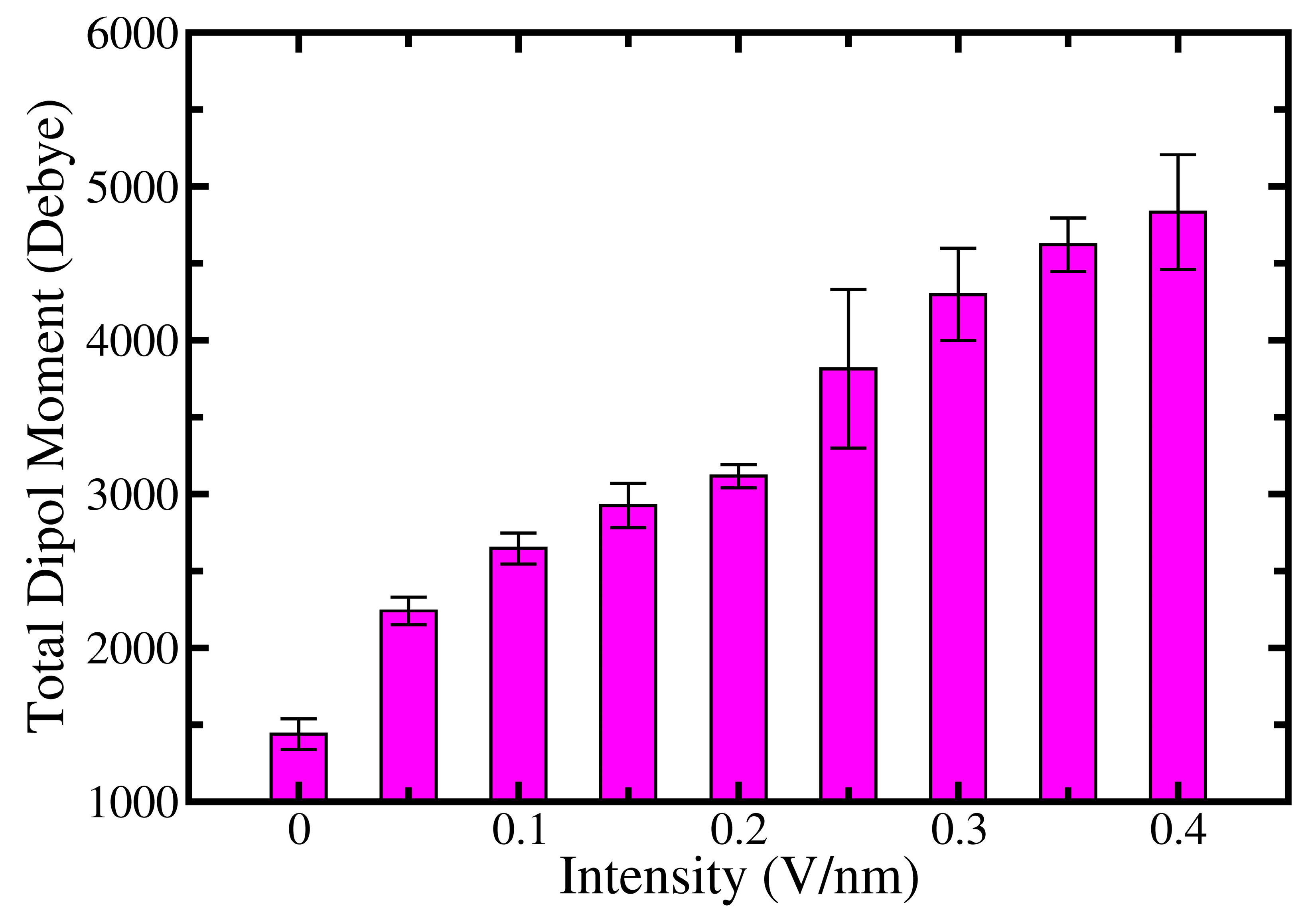
2. Results and Discussion
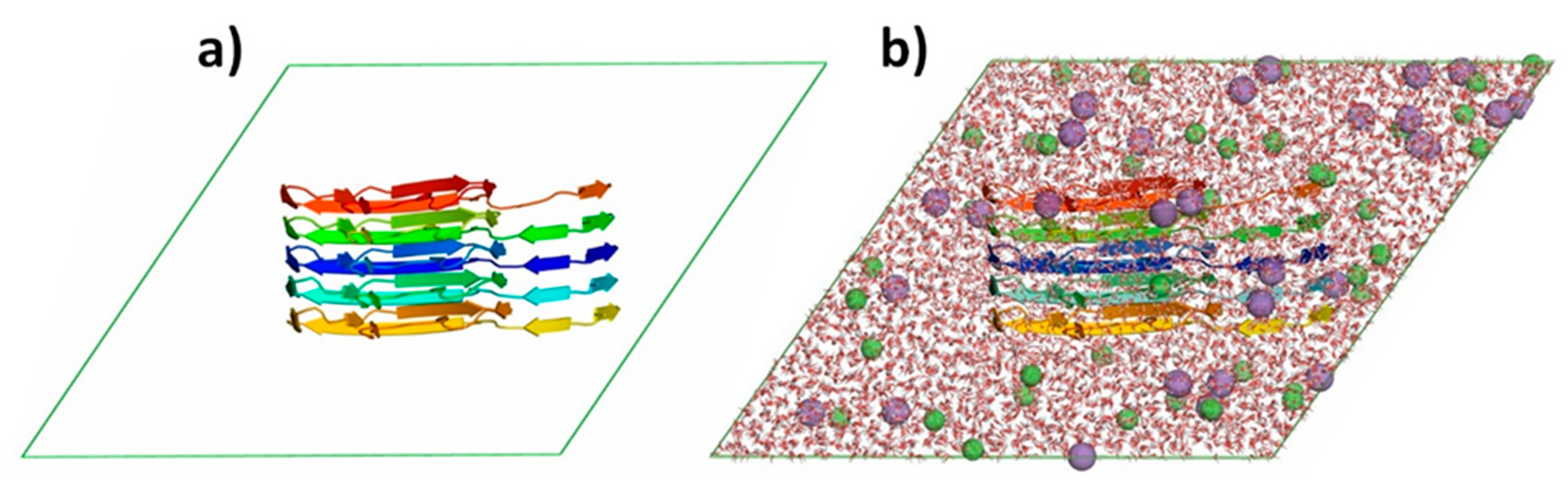
3. Methods and Materials
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Baev, A.Y.; Vinokurov, A.Y.; Novikova, I.N.; Dremin, V.V.; Potapova, E.V.; Abramov, A.Y. Interaction of Mitochondrial Calcium and ROS in Neurodegeneration. Cells 2022, 11, 706. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M. Alpha-synuclein and neurodegenerative diseases. Nat. Rev. Neurosci. 2001, 2, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Westman, E.; Ballard, C.; Aarsland, D. Cognitive impairment in patients with Parkinson’s disease: Diagnosis, biomarkers, and treatment. Lancet Neurol. 2012, 11, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Caughey, B.; Lansbury, P.T. Protofibrils, pores, fibrils, and neurodegeneration: Separating the responsible protein aggregates from the innocent bystanders. Annu. Rev. Neurosci. 2003, 26, 267–298. [Google Scholar] [CrossRef]
- Pieri, L.; Madiona, K.; Bousset, L.; Melki, R. Fibrillar alpha-synuclein and huntingtin exon 1 assemblies are toxic to the cells. Biophys. J. 2012, 102, 2894–2905. [Google Scholar] [CrossRef] [Green Version]
- Cookson, M.R. The biochemistry of Parkinson’s disease. Annu. Rev. Biochem. 2005, 74, 29–52. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, J.T.; Lansbury, P.T. Seeding “one-dimensional crystallization” of amyloid: A pathogenic mechanism in Alzheimer’s disease and scrapie? Cell 1993, 73, 1055–1058. [Google Scholar] [CrossRef]
- Uversky, V.N. A protein-chameleon: Conformational plasticity of alpha-synuclein, a disordered protein involved in neurodegenerative disorders. J. Biomol. Struct. Dyn. 2003, 21, 211–234. [Google Scholar] [CrossRef]
- Cole, N.B.; Murphy, D.D. The cell biology of α-synuclein. NeuroMol. Med. 2002, 1, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Wong, Y.C.; Krainc, D. alpha-synuclein toxicity in neurodegeneration: Mechanism and therapeutic strategies. Nat. Med. 2017, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Moretto, E.; Stuart, S.; Surana, S.; Vargas, J.N.S.; Schiavo, G. The Role of Extracellular Matrix Components in the Spreading of Pathological Protein Aggregates. Front. Cell Neurosci. 2022, 16, 844211. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Guerra, F.; Calvani, R.; Coelho-Junior, H.J.; Bucci, C.; Marzetti, E. Circulating extracellular vesicles: Friends and foes in neurodegeneration. Neural Regen. Res. 2022, 17, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, D.; Cisternas-Olmedo, M.; Arcos, J.; Nassif, M.; Vidal, R.L. Contribution of Autophagy-Lysosomal Pathway in the Exosomal Secretion of Alpha-Synuclein and Its Impact in the Progression of Parkinson’s Disease. Front. Mol. Neurosci. 2022, 15, 805087. [Google Scholar] [CrossRef]
- Uversky, V.N. Neuropathology, biochemistry, and biophysics of alpha-synuclein aggregation. J. Neurochem. 2007, 103, 17–37. [Google Scholar] [CrossRef]
- Burre, J. The Synaptic Function of alpha-Synuclein. J. Park. Dis. 2015, 5, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Ludtmann, M.H.; Angelova, P.R.; Ninkina, N.N.; Gandhi, S.; Buchman, V.L.; Abramov, A.Y. Monomeric Alpha-Synuclein Exerts a Physiological Role on Brain ATP Synthase. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 10510–10521. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Yang, D.; Li, X.Q.; Liu, J.; Back, T.C.; Trivett, A.; Karim, B.; Barbut, D.; Zasloff, M.; Oppenheim, J.J. Alpha synuclein, the culprit in Parkinson disease, is required for normal immune function. Cell Rep. 2022, 38, 110090. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. α-Synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Creekmore, B.C.; Chang, Y.W.; Lee, E.B. The Cryo-EM Effect: Structural Biology of Neurodegenerative Disease Aggregates. J. Neuropathol. Exp. Neurol. 2021, 80, 514–529. [Google Scholar] [CrossRef]
- Tarutani, A.; Adachi, T.; Akatsu, H.; Hashizume, Y.; Hasegawa, K.; Saito, Y.; Robinson, A.C.; Mann, D.M.A.; Yoshida, M.; Murayama, S.; et al. Ultrastructural and biochemical classification of pathogenic tau, alpha-synuclein and TDP-43. Acta Neuropathol. 2022, 143, 613–640. [Google Scholar] [CrossRef] [PubMed]
- Koopman, M.B.; Ferrari, L.; Rudiger, S.G.D. How do protein aggregates escape quality control in neurodegeneration? Trends Neurosci. 2022, 45, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C. Conformational strains of pathogenic amyloid proteins in neurodegenerative diseases. Nat. Rev. Neurosci. 2022, 23, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Armstrong, S.; Hamzeh, A.; Visanji, N.P.; Sardi, S.P.; Tandon, A. Alpha-Synuclein Targeting Therapeutics for Parkinson’s Disease and Related Synucleinopathies. Front. Neurol. 2022, 13, 852003. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, J.T.; Kelly, L.; Verma, A.; Carare, R.O.; Nicoll, J.A.R.; Dodart, J.C. Amyloid-beta and alpha-Synuclein Immunotherapy: From Experimental Studies to Clinical Trials. Front. Neurosci. 2021, 15, 733857. [Google Scholar] [CrossRef]
- Magalhaes, P.; Lashuel, H.A. Opportunities and challenges of alpha-synuclein as a potential biomarker for Parkinson’s disease and other synucleinopathies. NPJ Park. Dis. 2022, 8, 93. [Google Scholar] [CrossRef]
- Minnaar, C.A.; Szasz, A. Forcing the Antitumor Effects of HSPs Using a Modulated Electric Field. Cells 2022, 11, 1838. [Google Scholar] [CrossRef]
- Hartmann, C.J.; Fliegen, S.; Groiss, S.J.; Wojtecki, L.; Schnitzler, A. An update on best practice of deep brain stimulation in Parkinson’s disease. Adv. Neurol. Disord. 2019, 12, 1756286419838096. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.S.; Suresh, A.; Pirogova, E. Effects of oscillating electric fields on conotoxin peptide conformation: A molecular dynamic simulation study. J. Mol. Graph. Model. 2021, 103, 107799. [Google Scholar] [CrossRef]
- Della Valle, E.; Marracino, P.; Pakhomova, O.; Liberti, M.; Apollonio, F. Nanosecond pulsed electric signals can affect electrostatic environment of proteins below the threshold of conformational effects: The case study of SOD1 with a molecular simulation study. PLoS ONE 2019, 14, e0221685. [Google Scholar] [CrossRef] [PubMed]
- Astrakas, L.; Gousias, C.; Tzaphlidou, M. Electric field effects on chignolin conformation. J. Appl. Phys. 2011, 109, 094702. [Google Scholar] [CrossRef]
- Lu, Y.; Shi, X.F.; Salsbury, F.R., Jr.; Derreumaux, P. Small static electric field strength promotes aggregation-prone structures in amyloid-beta(29-42). J. Chem. Phys. 2017, 146, 145101. [Google Scholar] [CrossRef] [PubMed]
- Todorova, N.; Bentvelzen, A.; Yarovsky, I. Electromagnetic field modulates aggregation propensity of amyloid peptides. J. Chem. Phys. 2020, 152, 035104. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shi, X.F.; Salsbury, F.R., Jr.; Derreumaux, P. Influence of electric field on the amyloid-beta(29-42) peptides embedded in a membrane bilayer. J. Chem. Phys. 2018, 148, 045105. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.K.; Mitra, S.; Chakraborty, M.; Ghosh, S.; Sen, S.; Dasgupta, S.; DasGupta, S. Disruption of human serum albumin fibrils by a static electric field. J. Phys. D Appl. Phys. 2014, 47, 148570. [Google Scholar] [CrossRef]
- Olloqui-Sariego, J.L.; Perez-Mejias, G.; Marquez, I.; Guerra-Castellano, A.; Calvente, J.J.; De la Rosa, M.A.; Andreu, R.; Diaz-Moreno, I. Electric field-induced functional changes in electrode-immobilized mutant species of human cytochrome c. Biochim Biophys. Acta Bioenerg. 2022, 1863, 148570. [Google Scholar] [CrossRef]
- Samaranayake, C.P.; Sastry, S.K. Molecular dynamics evidence for nonthermal effects of electric fields on pectin methylesterase activity. Phys. Chem. Chem. Phys. 2021, 23, 14422–14432. [Google Scholar] [CrossRef]
- Favi, P.M.; Zhang, Q.; O’Neill, H.; Mamontov, E.; Diallo, S.O. Dynamics of lysozyme and its hydration water under an electric field. J. Biol. Phys. 2014, 40, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Liao, C.; Zhou, J. Effects of external electric fields on lysozyme adsorption by molecular dynamics simulations. Biophys. Chem. 2013, 179, 26–34. [Google Scholar] [CrossRef]
- Basu, S.; Gorai, B.; Basu, B.; Maiti, P.K. Electric Field-Mediated Fibronectin-Hydroxyapatite Interaction: A Molecular Insight. J. Phys. Chem. B 2021, 125, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; You, L.; Dou, W.; Sun, T.; Xu, P. Effects of an Electric Field on the Conformational Transition of the Protein: A Molecular Dynamics Simulation Study. Polymers 2019, 11, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscat, S.; Stojceski, F.; Danani, A. Elucidating the effect of static electric field on amyloid beta 1–42 supramolecular assembly. J. Mol. Graph. Model. 2020, 96, 107535. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, Y.; Wei, G.; Ding, F.; Sun, Y. Molecular Insights into the Misfolding and Dimerization Dynamics of the Full-Length α-Synuclein from Atomistic Discrete Molecular Dynamics Simulations. ACS Chem. Neurosci. 2022, 13, 3126–3137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Tian, Y.; Li, Z.; Liu, S.; Hu, X.; Yang, Z.; Ling, X.; Liu, S.; Zhang, J. Modeling. Molecular dynamics study to investigate the dimeric structure of the full-length α-synuclein in aqueous solution. J. Chem. Inf. Model. 2017, 57, 2281–2293. [Google Scholar] [CrossRef]
- Lindsay, R.J.; Mansbach, R.A.; Gnanakaran, S.; Shen, T. Effects of pH on an IDP conformational ensemble explored by molecular dynamics simulation. Biophys. Chem. 2021, 271, 106552. [Google Scholar] [CrossRef]
- Dedmon, M.M.; Lindorff-Larsen, K.; Christodoulou, J.; Vendruscolo, M.; Dobson, C.M. Mapping long-range interactions in α-synuclein using spin-label NMR and ensemble molecular dynamics simulations. J. Am. Chem. Soc. 2005, 127, 476–477. [Google Scholar] [CrossRef]
- Han, J.Y.; Choi, T.S.; Kim, H.I. Molecular role of Ca2+ and hard divalent metal cations on accelerated fibrillation and interfibrillar aggregation of α-synuclein. Sci. Rep. 2018, 8, 1895. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Zacharopoulou, M.; Schierle, G.S.K. The cellular environment affects monomeric α-synuclein structure. Trends Biochem. Sci. 2019, 44, 453–466. [Google Scholar] [CrossRef]
- Li, B.; Ge, P.; Murray, K.A.; Sheth, P.; Zhang, M.; Nair, G.; Sawaya, M.R.; Shin, W.S.; Boyer, D.R.; Ye, S. Cryo-EM of full-length α-synuclein reveals fibril polymorphs with a common structural kernel. Nat. Commun. 2018, 9, 3609. [Google Scholar] [CrossRef]
- Hou, L.; Zagorski, M.G. Sorting out the driving forces for parallel and antiparallel alignment in the Aβ peptide fibril structure. Biophys. J. 2004, 86, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.W.; Drakulic, S.; Deas, E.; Ouberai, M.; Aprile, F.A.; Arranz, R.; Ness, S.; Roodveldt, C.; Guilliams, T.; De-Genst, E.J. Structural characterization of toxic oligomers that are kinetically trapped during α-synuclein fibril formation. Proc. Natl. Acad. Sci. USA 2015, 112, E1994–E2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzokov, J.; Naderi, S.; Van Der Schoot, P. Prediction of the structure of a silk-like protein in oligomeric states using explicit and implicit solvent models. Soft Matter 2014, 10, 5362–5374. [Google Scholar] [CrossRef] [PubMed]
- Razzokov, J.; Naderi, S.; Van Der Schoot, P. Nanoscale insight into silk-like protein self-assembly: Effect of design and number of repeat units. Phys. Biol. 2018, 15, 066010. [Google Scholar] [CrossRef] [Green Version]
- Marshall, K.E.; Morris, K.L.; Charlton, D.; O’Reilly, N.; Lewis, L.; Walden, H.; Serpell, L.C. Hydrophobic, aromatic, and electrostatic interactions play a central role in amyloid fibril formation and stability. Biochemistry 2011, 50, 2061–2071. [Google Scholar] [CrossRef]
- Müller, W.A.; Sarkis, J.R.; Marczak, L.D.F.; Muniz, A.R. Molecular dynamics study of the effects of static and oscillating electric fields in ovalbumin. Innov. Food Sci. Emerg. Technol. 2022, 75, 102911. [Google Scholar] [CrossRef]
- Kuang, Z.; Luginsland, J.; Thomas, R.J.; Dennis, P.B.; Kelley-Loughnane, N.; Roach, W.P.; Naik, R.R. Molecular dynamics simulations explore effects of electric field orientations on spike proteins of SARS-CoV-2 virions. Sci. Rep. 2022, 12, 12986. [Google Scholar] [CrossRef]
- Boyer, D.R.; Li, B.; Sun, C.; Fan, W.; Zhou, K.; Hughes, M.P.; Sawaya, M.R.; Jiang, L.; Eisenberg, D.S. The α-synuclein hereditary mutation E46K unlocks a more stable, pathogenic fibril structure. Proc. Natl. Acad. Sci. USA 2020, 117, 3592–3602. [Google Scholar] [CrossRef]
- Yao, Y.; Tang, Y.; Wei, G. Epigallocatechin Gallate Destabilizes α-Synuclein Fibril by Disrupting the E46–K80 Salt-Bridge and Inter-protofibril Interface. ACS Chem. Neurosci. 2020, 11, 4351–4361. [Google Scholar] [CrossRef]
- Razzokov, J.; Yusupov, M.; Bogaerts, A. Oxidation destabilizes toxic amyloid beta peptide aggregation. Sci. Rep. 2019, 9, 5476. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Margreitter, C.; Reif, M.M.; Oostenbrink, C. Update on phosphate and charged post-translationally modified amino acid parameters in the GROMOS force field. J. Comput. Chem. 2017, 38, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.J.; Postma, J.P.; van Gunsteren, W.F.; Hermans, J. Interaction models for water in relation to protein hydration. In Intermolecular Forces; Springer: Berlin/Heidelberg, Germany, 1981; pp. 331–342. [Google Scholar]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Maiorov, V.N.; Crippen, G.M. Size-independent comparison of protein three-dimensional structures. Proteins Struct. Funct. Bioinform. 1995, 22, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Eisenhaber, F.; Lijnzaad, P.; Argos, P.; Sander, C.; Scharf, M. The double cubic lattice method: Efficient approaches to numerical integration of surface area and volume and to dot surface contouring of molecular assemblies. J. Comput. Chem. 1995, 16, 273–284. [Google Scholar] [CrossRef]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Cryst. 2002, 40, 82–92. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Guzmán, J.N.; Sánchez-Padilla, J.; Goldberg, J.A. What causes the death of dopaminergic neurons in Parkinson’s disease? Prog. Brain Res. 2010, 183, 59–77. [Google Scholar]
- Kuncel, A.M.; Grill, W.M. Selection of stimulus parameters for deep brain stimulation. Clin. Neurophysiol. 2004, 115, 2431–2441. [Google Scholar] [CrossRef]
- Deas, E.; Cremades, N.; Angelova, P.R.; Ludtmann, M.H.; Yao, Z.; Chen, S.; Horrocks, M.H.; Banushi, B.; Little, D.; Devine, M.J.; et al. Alpha-Synuclein Oligomers Interact with Metal Ions to Induce Oxidative Stress and Neuronal Death in Parkinson’s Disease. Antioxid. Redox Signal. 2016, 24, 376–391. [Google Scholar] [CrossRef] [Green Version]
- Vaikath, N.; Sudhakaran, I.; Abdi, I.; Gupta, V.; Majbour, N.; Ghanem, S.; Abdesselem, H.; Vekrellis, K.; El-Agnaf, O. Structural and Biophysical Characterization of Stable Alpha-Synuclein Oligomers. Int. J. Mol. Sci. 2022, 23, 14630. [Google Scholar] [CrossRef] [PubMed]
- Ludtmann, M.H.R.; Angelova, P.R.; Horrocks, M.H.; Choi, M.L.; Rodrigues, M.; Baev, A.Y.; Berezhnov, A.V.; Yao, Z.; Little, D.; Banushi, B.; et al. alpha-synuclein oligomers interact with ATP synthase and open the permeability transition pore in Parkinson’s disease. Nat. Commun. 2018, 9, 2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremades, N.; Cohen, S.I.; Deas, E.; Abramov, A.Y.; Chen, A.Y.; Orte, A.; Sandal, M.; Clarke, R.W.; Dunne, P.; Aprile, F.A.; et al. Direct observation of the interconversion of normal and toxic forms of alpha-synuclein. Cell 2012, 149, 1048–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EF (V/nm) | α-Helix | 3 10-Helix | β-Sheet | β-Bridge | Bend | Turn | Coil |
|---|---|---|---|---|---|---|---|
| 0.0 | 0.0 | 0.0 | 49 | 2 | 11 | 6 | 32 |
| 0.05 | 0.0 | 0.0 | 40 | 3 | 11 | 7 | 38 |
| 0.10 | 0.0 | 1.0 | 40 | 2 | 14 | 5 | 39 |
| 0.15 | 0.0 | 1.0 | 40 | 2 | 11 | 6 | 39 |
| 0.20 | 1.0 | 1.0 | 38 | 2 | 12 | 5 | 40 |
| 0.25 | 0.0 | 1.0 | 32 | 2 | 13 | 7 | 44 |
| 0.30 | 0.0 | 1.0 | 19 | 6 | 20 | 4 | 49 |
| 0.35 | 4.0 | 0.0 | 20 | 2 | 15 | 4 | 54 |
| 0.40 | 3.0 | 1.0 | 13 | 5 | 10 | 3 | 64 |
| EF (V/nm) | SASA (nm2) | R Gyration (Å) | h-Bond/Chain |
|---|---|---|---|
| 0.00 | 139.58 ± 2.38 | 2.013 ± 0.02 | 48.20 ± 1.75 |
| 0.05 | 140.52 ± 2.42 | 2.028 ± 0.01 | 47.08 ± 1.55 |
| 0.10 | 153.90 ± 2.65 | 2.129 ± 0.01 | 44.08 ± 1.47 |
| 0.15 | 153.27 ± 2.44 | 2.142 ± 0.01 | 45.63 ± 1.52 |
| 0.20 | 150.69 ± 2.75 | 2.136 ± 0.01 | 44.73 ± 1.50 |
| 0.25 | 152.02 ± 2.65 | 2.214 ± 0.01 | 45.68 ± 1.44 |
| 0.30 | 164.08 ± 4.01 | 3.150 ± 0.03 | 40.03 ± 1.65 |
| 0.35 | 166.54 ± 4.33 | 3.398 ± 0.02 | 40.30 ± 1.58 |
| 0.40 | 184.36 ± 4.32 | 4.179 ± 0.02 | 35.32 ± 1.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razzokov, J.; Fazliev, S.; Makhkamov, M.; Marimuthu, P.; Baev, A.; Kurganov, E. Effect of Electric Field on α-Synuclein Fibrils: Revealed by Molecular Dynamics Simulations. Int. J. Mol. Sci. 2023, 24, 6312. https://doi.org/10.3390/ijms24076312
Razzokov J, Fazliev S, Makhkamov M, Marimuthu P, Baev A, Kurganov E. Effect of Electric Field on α-Synuclein Fibrils: Revealed by Molecular Dynamics Simulations. International Journal of Molecular Sciences. 2023; 24(7):6312. https://doi.org/10.3390/ijms24076312
Chicago/Turabian StyleRazzokov, Jamoliddin, Sunnatullo Fazliev, Mukhriddin Makhkamov, Parthiban Marimuthu, Artyom Baev, and Erkin Kurganov. 2023. "Effect of Electric Field on α-Synuclein Fibrils: Revealed by Molecular Dynamics Simulations" International Journal of Molecular Sciences 24, no. 7: 6312. https://doi.org/10.3390/ijms24076312
APA StyleRazzokov, J., Fazliev, S., Makhkamov, M., Marimuthu, P., Baev, A., & Kurganov, E. (2023). Effect of Electric Field on α-Synuclein Fibrils: Revealed by Molecular Dynamics Simulations. International Journal of Molecular Sciences, 24(7), 6312. https://doi.org/10.3390/ijms24076312