
Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies

 , and
, and
Abstract
:1. Introduction
1.1. COVID-19 Vaccines and Immunoserology
1.2. Natural (Auto)Antibodies (nAAbs)
1.3. Citrate Synthase (CS) as a Natural Autoantibody
1.4. Nonspecific Effects of Vaccines (NSEs)
1.5. Heat Shock Protein 60 and 70 (HSP60 and HSP70)
1.6. COVID-19 and Autoimmunity
1.7. Serologic Response to COVID-19 Vaccines in Patients Treated with Biological Therapies
2. Results
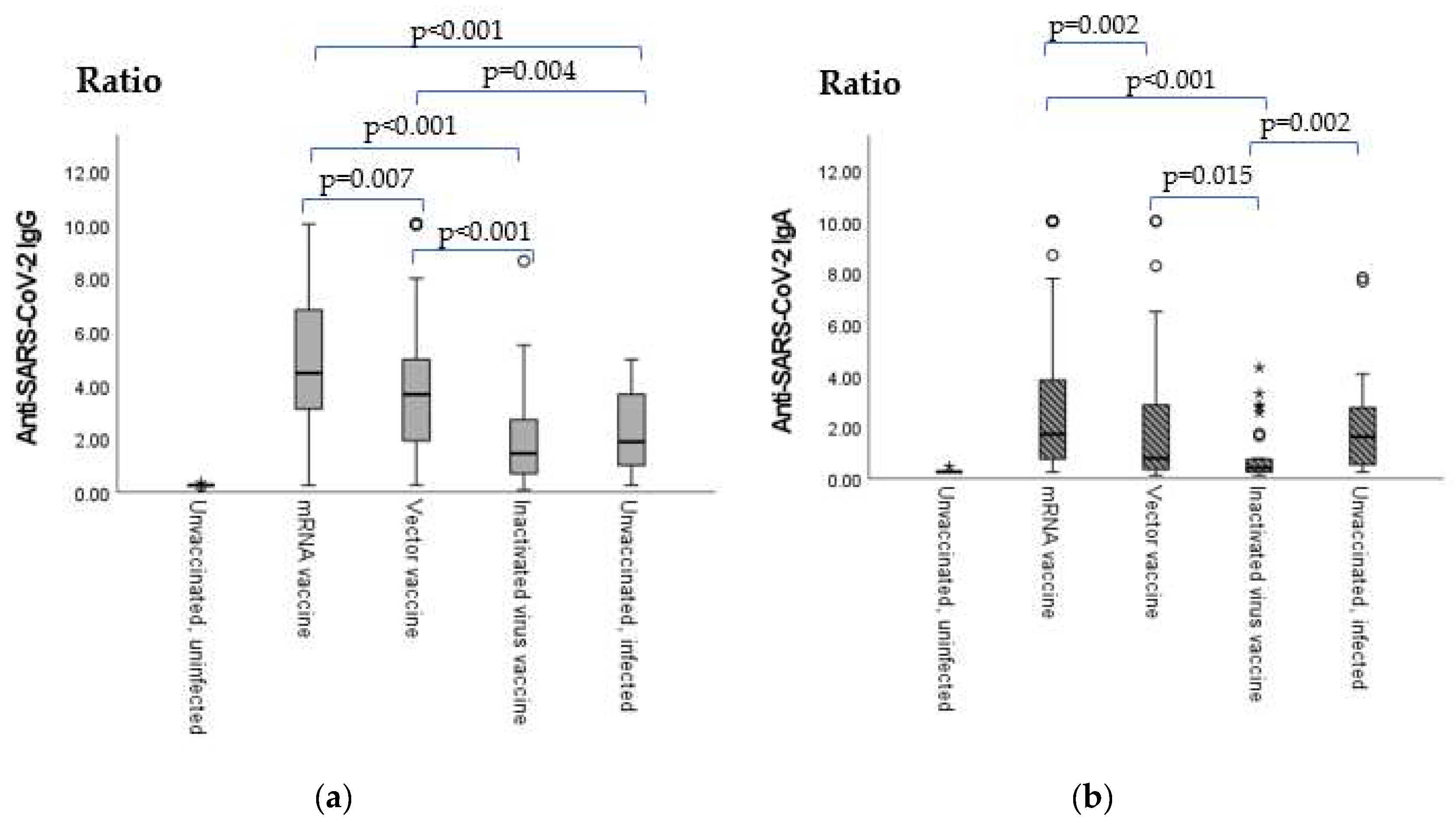
2.1. Anti-SARS-CoV-2 Vaccination- or Infection-Induced IgG and IgA Antibody Levels
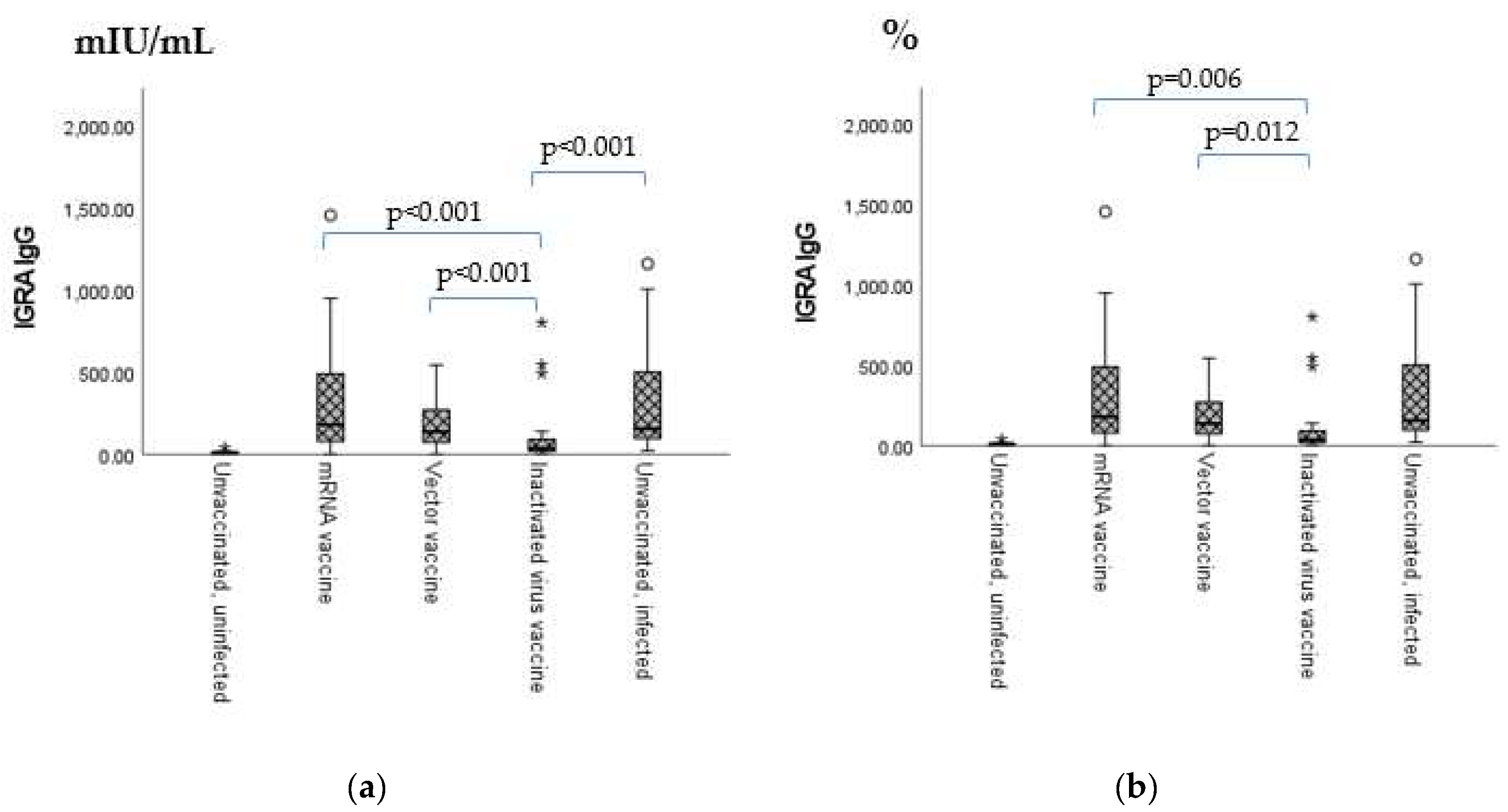
2.2. Anti-SARS-CoV-2 Neutralizing Antibody Titers and Anti-IFN-γ (IGRA) Levels
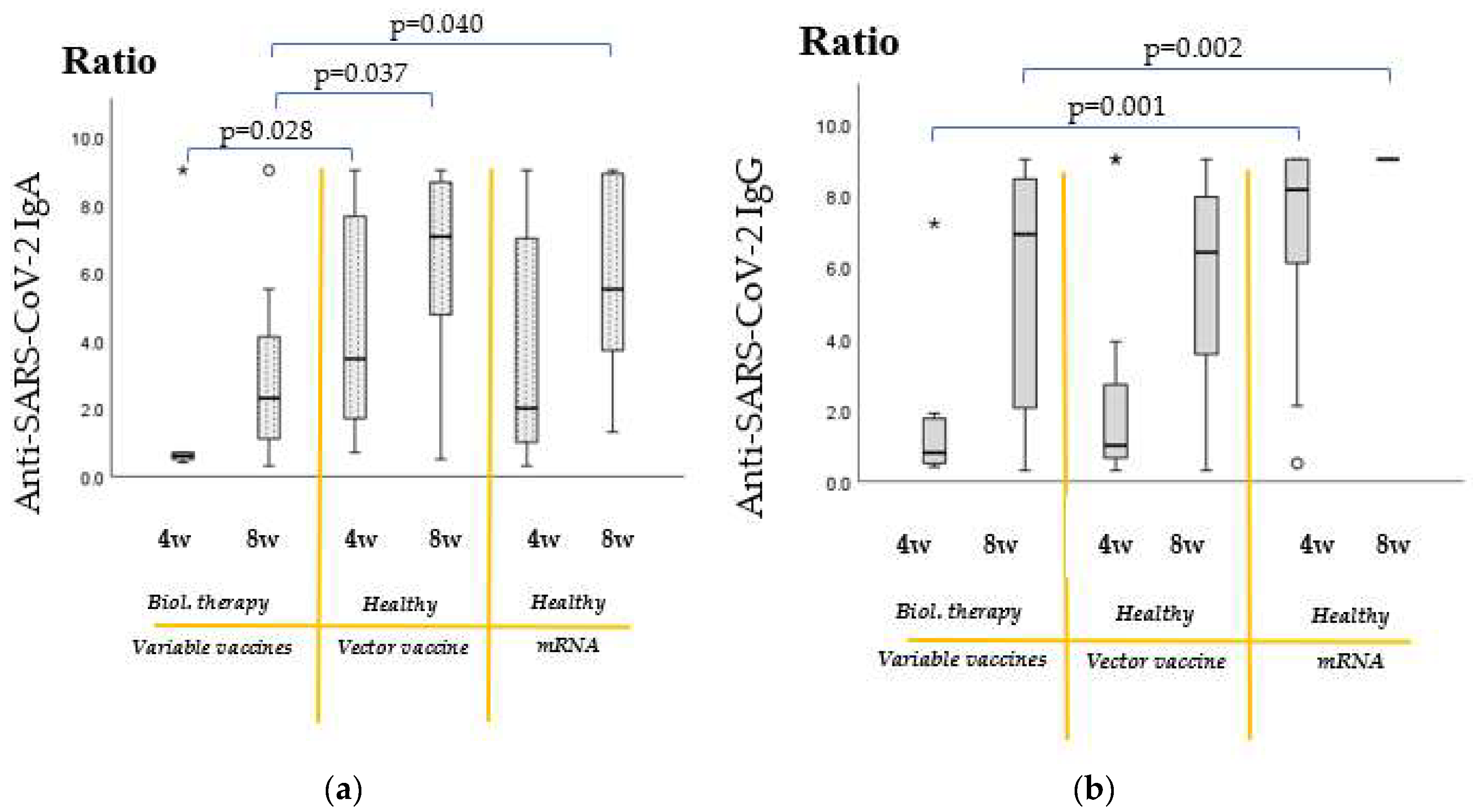
2.3. Increase in Antibody Titers of Biological Therapy-Treated Patients from the 4th to 8th Week Post-Vaccination
2.4. Comparison of Vaccine Efficacy Results of Recipients of Immunomodulatory Therapies with Untreated, Healthy Individuals
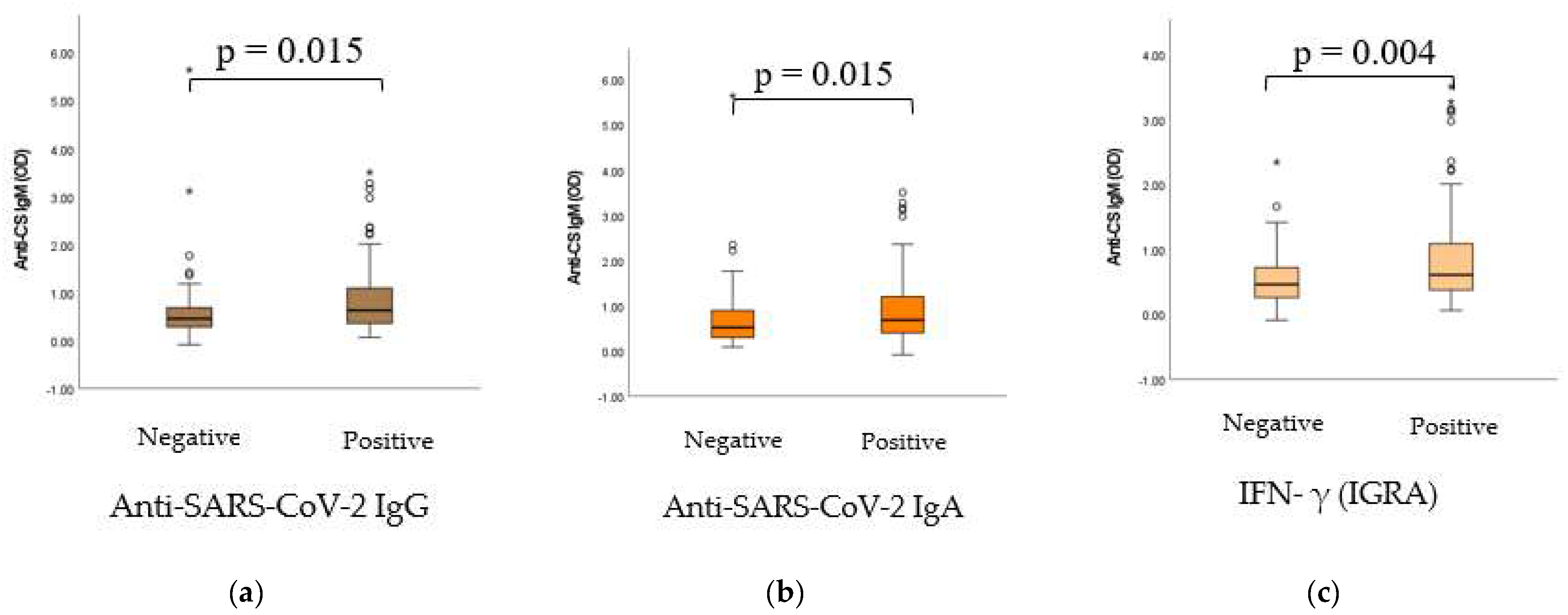
2.5. Dynamic Adaptation of IgM Isotype Anti-CS Natural Autoantibodies (nAAbs)
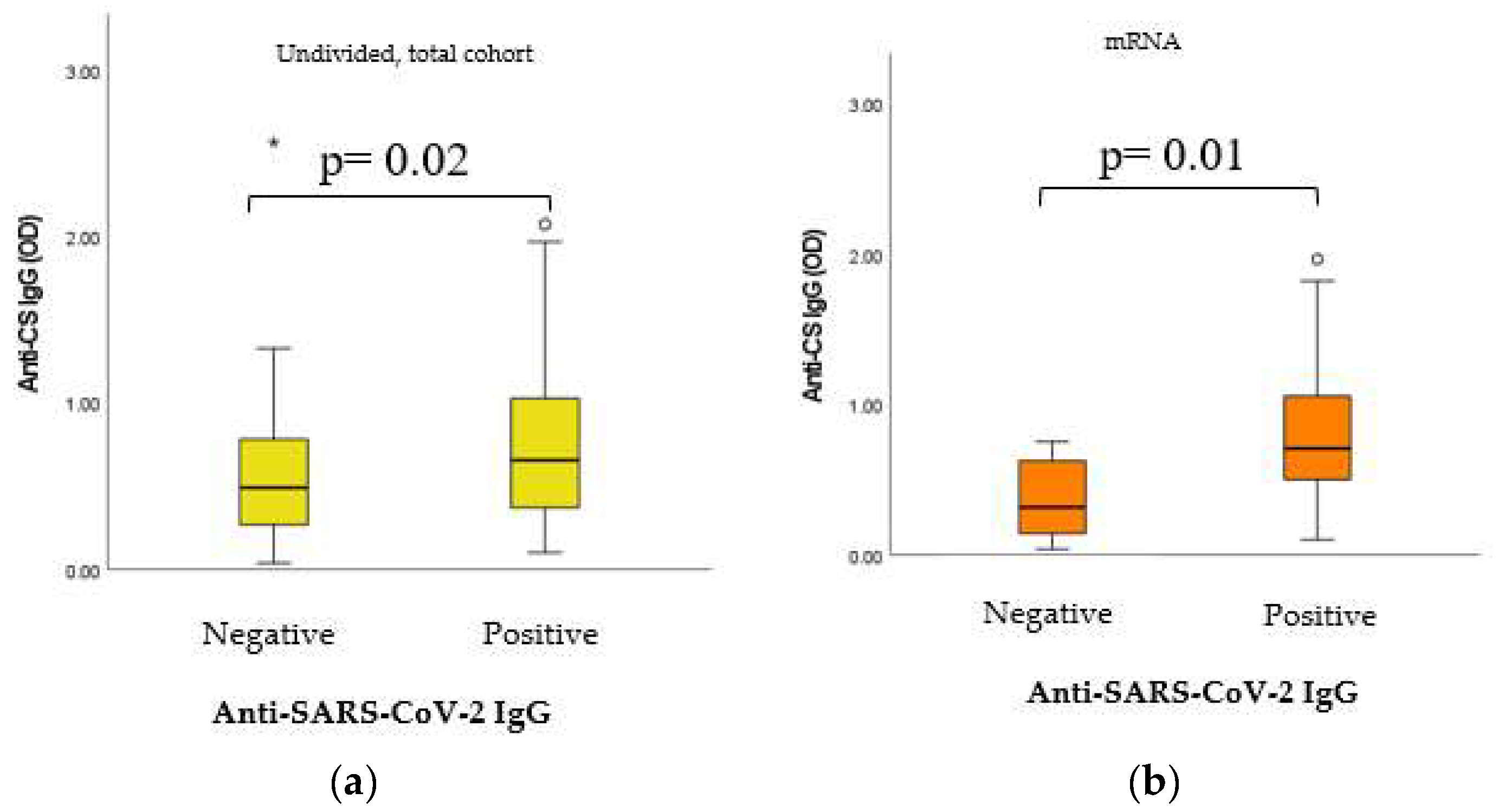
2.6. Dynamic Adaptation of IgG Isotype Anti-CS Natural Autoantibodies (nAAbs)
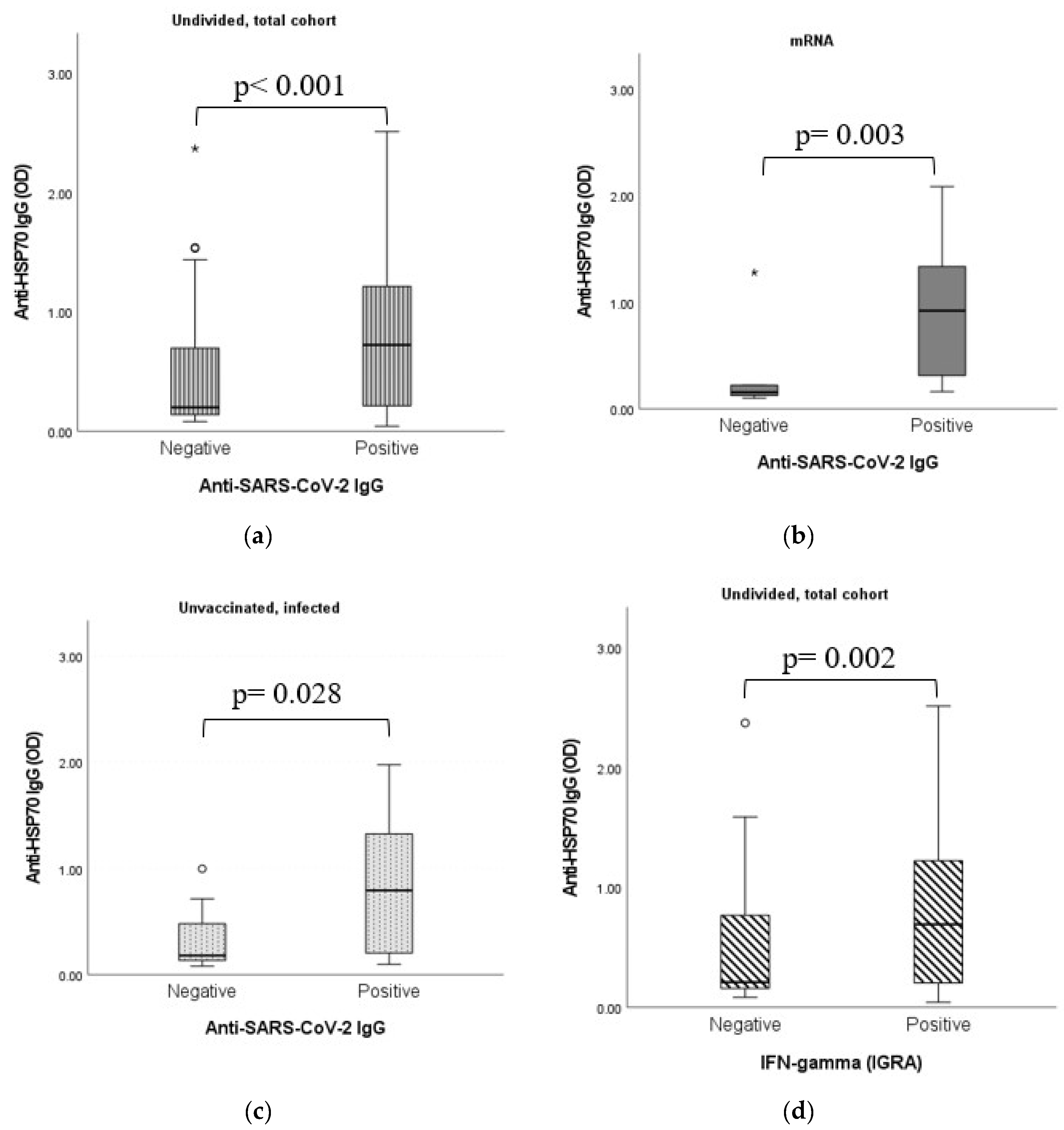
2.7. Dynamic Adaptation of Anti-HSP70 IgG Antibodies
2.8. Dynamic Adaptation of Anti-HSP70 IgG Antibodies
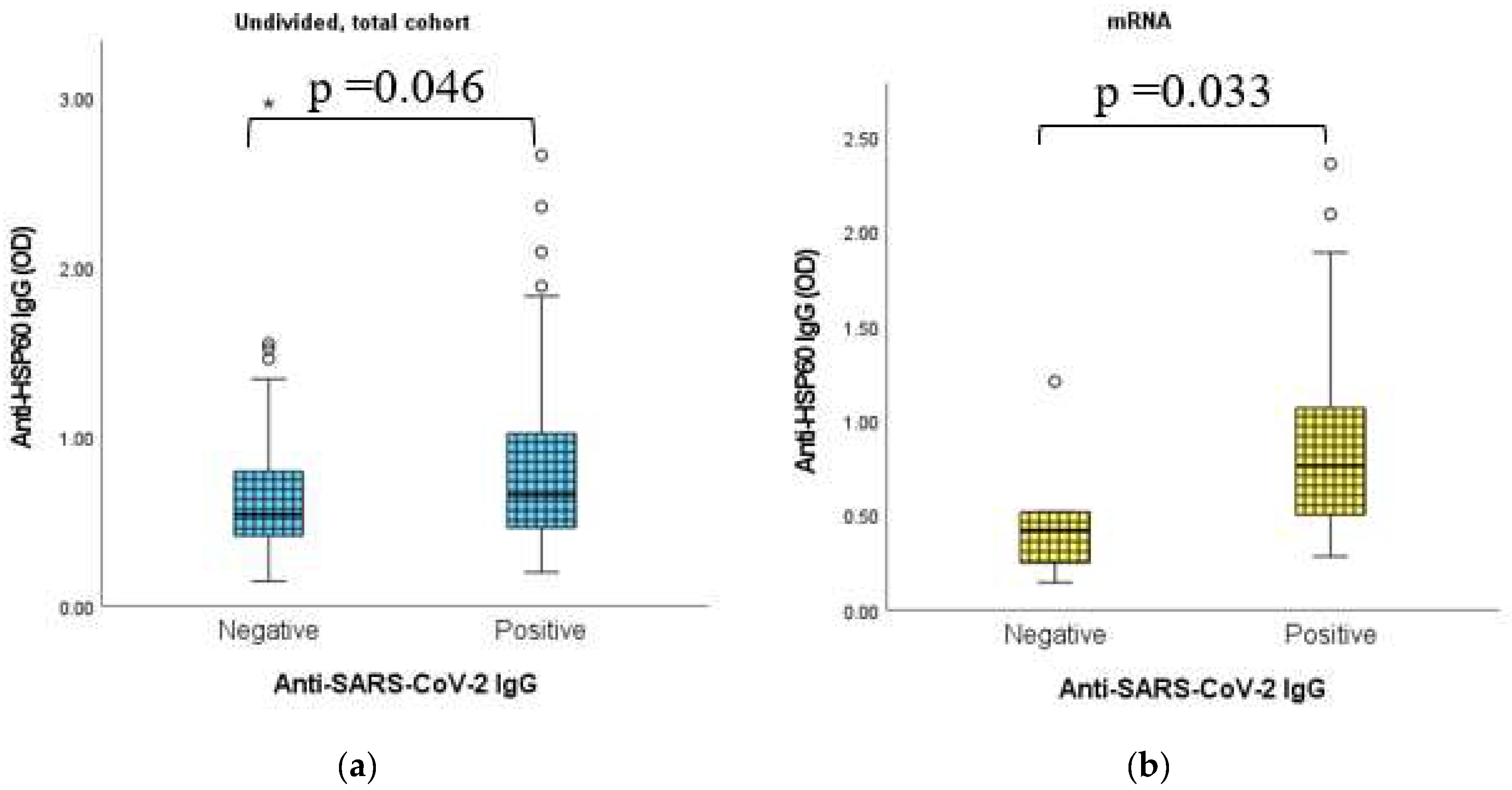
2.9. Dynamic Adaptation of Anti-HSP60 IgG Antibodies
3. Discussion
3.1. Vaccine Efficacy
3.2. Potential Interference of Biological Therapies (Anti-TNFα and Anti-IL17) with Vaccine Efficacy
3.3. Vaccines (or Infections) and nAAbs
4. Materials and Methods
4.1. Human Blood Samples
4.2. Human Serum Samples for the Comparison of Healthy Individuals versus Autoimmune Patients Receiving Biological Therapies
4.3. Hsp60/Hsp70 IgG and IgM In-House ELISA Assays
4.4. Citrate Synthase (CS) IgG and IgM In-House ELISA Assays
4.5. Neutralizing Antibody Measurement—Virus Neutralization ELISA (sVNT)
4.6. Interferon-γ ELISA
4.7. Anti-SARS-CoV-2 ELISA (IgG, IgA)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE2 | Human angiotensin-converting enzyme 2 |
| BCG | Bacillus calmette–guérin |
| CS | Citrate synthase |
| DTP | Diphtheria, tetanus and pertussis vaccine |
| HRP | Horseradish peroxidase |
| HSP | Heat shock protein |
| Ig | Immunoglobulin |
| IGRA | Interferon gamma release assay |
| IL | Interleukin |
| MMR | Measles, mumps and rubella vaccine |
| MS | Microsoft |
| MV | Measles vaccine |
| nAAb | Natural autoantibody |
| nAb | Natural antibody |
| NSE | Nonspecific side effect |
| PBS | Phosphate-buffered saline |
| PVA | Polyvinyl alcohol |
| RBD | Receptor-binding domain |
| RT | Room temperature |
| S1 | Spike glycoprotein 1 |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| TMB | 3,3′,5,5′-tetramethylbenzidine |
| TNF-α | Tumor necrosis factor alpha |
| URIs | Upper respiratory infections |
| WB | Washing buffer |
References
- Keshavarz, B.; Richards, N.E.; Workman, L.J.; Patel, J.; Muehling, L.M.; Canderan, G.; Murphy, D.D.; Brovero, S.G.; Ailsworth, S.M.; Eschenbacher, W.H.; et al. Trajectory of IgG to SARS-CoV-2 After Vaccination with BNT162b2 or MRNA-1273 in an Employee Cohort and Comparison with Natural Infection. Front. Immunol. 2022, 13, 1144. [Google Scholar] [CrossRef]
- Appelgren, D.; Eriksson, P.; Ernerudh, J.; Segelmark, M. Marginal-Zone B-Cells Are Main Producers of IgM in Humans, and Are Reduced in Patients with Autoimmune Vasculitis. Front. Immunol. 2018, 9, 2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czömpöly, T.; Olasz, K.; Simon, D.; Nyárády, Z.; Pálinkás, L.; Czirják, L.; Berki, T.; Németh, P. A possible new bridge between innate and adaptive immunity: Are the anti-mitochondrial citrate synthase autoantibodies components of the natural antibody network? Mol. Immunol. 2006, 43, 1761–1768. [Google Scholar] [CrossRef]
- Reyneveld, I.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Holodick, N.E.; Rodríguez-Zhurbenko, N.; Hernández, A.M. Defining Natural Antibodies. Front. Immunol. 2017, 8, 872. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, H.R.; Gong, J.; Gyles, C.L.; Hayes, M.A.; Zhou, H.; Sanei, B.; Chambers, J.R.; Sharif, S. Probiotics Stimulate Production of Natural Antibodies in Chickens. Clin. Vaccine Immunol. 2006, 13, 975–980. [Google Scholar] [CrossRef] [Green Version]
- Beinart, D.; Ren, D.; Pi, C.; Poulton, S.; Holzknecht, Z.E.; Swanson, C.; Parker, W. Immunization enhances the natural antibody repertoire. EXCLI J. 2017, 16, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Bachi, A.L.; Suguri, V.M.; Ramos, L.R.; Mariano, M.; Vaisberg, M.; Lopes, J.D.; Bachi, A.L.; Suguri, V.M.; Ramos, L.R.; Mariano, M.; et al. Increased production of autoantibodies and specific antibodies in response to influenza virus vaccination in physically active older individuals. Results Immunol. 2013, 3, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Maddur, M.S.; Lacroix-Desmazes, S.; Dimitrov, J.D.; Kazatchkine, M.D.; Bayry, J.; Kaveri, S.V. Natural Antibodies: From First-Line Defense Against Pathogens to Perpetual Immune Homeostasis. Clin. Rev. Allergy Immunol. 2019, 58, 213–228. [Google Scholar] [CrossRef]
- Kohler, H.; Bayry, J.; Nicoletti, A.; Kaveri, S.V. Natural Autoantibodies as Tools to Predict the Outcome of Immune Response? Scand. J. Immunol. 2003, 58, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Czömpöly, T.; Olasz, K.; Nyárády, Z.; Simon, D.; Bovári, J.; Németh, P. Detailed analyses of antibodies recognizing mitochondrial antigens suggest similar or identical mechanism for production of natural antibodies and natural autoantibodies. Autoimmun. Rev. 2008, 7, 463–467. [Google Scholar] [CrossRef]
- Simon, D.; Czömpöly, T.; Berki, T.; Minier, T.; Peti, A.; Tóth, E.; Czirják, L.; Németh, P. Naturally occurring and disease-associated auto-antibodies against topoisomerase I: A fine epitope mapping study in systemic sclerosis and systemic lupus erythematosus. Int. Immunol. 2009, 21, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, D.; Gilicze, O.; Farkas, N.; Najbauer, J.; Németh, P.; Lénárd, L.; Berki, T. Correlation of natural autoantibodies and cardiovascular disease-related anti-bacterial antibodies in pericardial fluid of cardiac surgery patients. Clin. Exp. Immunol. 2018, 193, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönwall, C.; Akhter, E.; Oh, C.; Burlingame, R.W.; Petri, M.; Silverman, G.J. IgM autoantibodies to distinct apoptosis-associated antigens correlate with protection from cardiovascular events and renal disease in patients with SLE. Clin. Immunol. 2012, 142, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Benn, C.S.; Netea, M.G.; Selin, L.K.; Aaby, P. A small jab—A big effect: Nonspecific immunomodulation by vaccines. Trends Immunol. 2013, 34, 431–439. [Google Scholar] [CrossRef]
- Aaby, P.; Samb, B.; Simondon, F.; Seck, A.M.C.; Knudsen, K.; Whittle, H. Non-specific beneficial effect of measles immunisation: Analysis of mortality studies from developing countries. BMJ 1995, 311, 481–485. [Google Scholar] [CrossRef] [Green Version]
- Aaby, P.; Benn, C. Developing the concept of beneficial non-specific effect of live vaccines with epidemiological studies. Clin. Microbiol. Infect. 2019, 25, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Soares-Weiser, K.; A López-López, J.; Kakourou, A.; Chaplin, K.; Christensen, H.; Martin, N.K.; Sterne, J.A.C.; Reingold, A.L. Association of BCG, DTP, and measles containing vaccines with childhood mortality: Systematic review. BMJ 2016, 355, i5170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørup, S.; Jensen, A.K.G.; Aaby, P.; Benn, C.S. Revaccination with Measles-Mumps-Rubella Vaccine and Infectious Disease Morbidity: A Danish Register-based Cohort Study. Clin. Infect. Dis. 2018, 68, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Sørup, S.; Benn, C.S.; Poulsen, A.; Krause, T.G.; Aaby, P.; Ravn, H. Live Vaccine Against Measles, Mumps, and Rubella and the Risk of Hospital Admissions for Nontargeted Infections. JAMA 2014, 311, 826–835. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, R.; Voysey, M.; McQuaid, F.; De Nie, K.; Ryan, R.; Orr, O.; Uhlig, U.; Sande, C.; O’Connor, D.; Pollard, A.J. Non-specific immunological effects of selected routine childhood immunisations: Systematic review. BMJ 2016, 355, i5225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, I.R. Autoantibody repertoires, natural biomarkers, and system controllers. Trends Immunol. 2013, 34, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Makhoba, X.H.; Makumire, S. The capture of host cell’s resources: The role of heat shock proteins and polyamines in SARS-COV-2 (COVID-19) pathway to viral infection. Biomol. Concepts 2022, 13, 220–229. [Google Scholar] [CrossRef]
- Ko, S.-H.; Huang, L.-M.; Tarn, W.-Y. The Host Heat Shock Protein MRJ/DNAJB6 Modulates Virus Infection. Front. Microbiol. 2019, 10, 2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Liu, Q.; Liang, C.; Zhou, L. Structural and functional analysis of the Hsp70/Hsp40 chaperone system. Protein Sci. 2019, 29, 378–390. [Google Scholar] [CrossRef]
- Lubkowska, A.; Pluta, W.; Strońska, A.; Lalko, A. Role of Heat Shock Proteins (HSP70 and HSP90) in Viral Infection. Int. J. Mol. Sci. 2021, 22, 9366. [Google Scholar] [CrossRef] [PubMed]
- Iyer, K.; Chand, K.; Mitra, A.; Trivedi, J.; Mitra, D. Diversity in heat shock protein families: Functional implications in virus infection with a comprehensive insight of their role in the HIV-1 life cycle. Cell Stress Chaperon 2021, 26, 743–768. [Google Scholar] [CrossRef]
- Jakovac, H. COVID-19 and hypertension: Is the HSP60 culprit for the severe course and worse outcome? Am. J. Physiol. Circ. Physiol. 2020, 319, H793–H796. [Google Scholar] [CrossRef]
- Wan, Q.; Song, D.; Li, H.; He, M.-L. Stress proteins: The biological functions in virus infection, present and challenges for target-based antiviral drug development. Signal Transduct. Target. Ther. 2020, 5, 1–40. [Google Scholar] [CrossRef]
- Muhammad, J.S.; Siddiqui, R.; Khan, N.A. COVID-19: Does SARS-CoV-2 Modulate Acanthamoeba Epigenetics to Enhance Survival and Transmission in the Environment? ACS Pharmacol. Transl. Sci. 2021, 4, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, J.S.; Sharif-Askari, N.S.; Cui, Z.-G.; Hamad, M.; Halwani, R. SARS-CoV-2 Infection-Induced Promoter Hypomethylation as an Epigenetic Modulator of Heat Shock Protein A1L (HSPA1L) Gene. Front. Genet. 2021, 12, 622271. [Google Scholar] [CrossRef]
- Hallberg, P.; Smedje, H.; Eriksson, N.; Kohnke, H.; Daniilidou, M.; Öhman, I.; Yue, Q.-Y.; Cavalli, M.; Wadelius, C.; Magnusson, P.K.; et al. Pandemrix-induced narcolepsy is associated with genes related to immunity and neuronal survival. Ebiomedicine 2019, 40, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, Y.; Shoenfeld, Y. Vaccine-induced autoimmunity: The role of molecular mimicry and immune crossreaction. Cell. Mol. Immunol. 2018, 15, 586–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nellore, A.; Randall, T.D. Narcolepsy and influenza vaccination—The inappropriate awakening of immunity. Ann. Transl. Med. 2016, 4, S29. [Google Scholar] [CrossRef] [Green Version]
- Ehrenfeld, M.; Tincani, A.; Andreoli, L.; Cattalini, M.; Greenbaum, A.; Kanduc, D.; Alijotas-Reig, J.; Zinserling, V.; Semenova, N.; Amital, H.; et al. Covid-19 and autoimmunity. Autoimmun. Rev. 2020, 19, 102597. [Google Scholar] [CrossRef]
- Kasperkiewicz, M. Covid-19, heat shock proteins, and autoimmune bullous diseases: A potential link deserving further attention. Cell Stress Chaperon 2020, 26, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Gammazza, A.M.; Dieli, F.; Macario, D.; Macario, A.J. Does SARS-CoV-2 Trigger Stress-Induced Autoimmunity by Molecular Mimicry? A Hypothesis. J. Clin. Med. 2020, 9, 2038. [Google Scholar] [CrossRef]
- Price, K.N.; Frew, J.W.; Hsiao, J.L.; Shi, V.Y. COVID-19 and immunomodulator/immunosuppressant use in dermatology. J. Am. Acad. Dermatol. 2020, 82, e173–e175. [Google Scholar] [CrossRef]
- Lebwohl, M.; Rivera-Oyola, R.; Murrell, D.F. Should biologics for psoriasis be interrupted in the era of COVID-19? J. Am. Acad. Dermatol. 2020, 82, 1217–1218. [Google Scholar] [CrossRef]
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Wang, S.; Chen, L.; Shen, Z. Biologics for Psoriasis During the COVID-19 Pandemic. Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; E Gudjonsson, J.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Rabinowitz, K.M.; Navon, M.; Edelman-Klapper, H.; Zittan, E.; Shitrit, A.B.-G.; Goren, I.; Avni-Biron, I.; Ollech, J.E.; Lichtenstein, L.; Banai-Eran, H.; et al. Anti-TNFα Treatment Impairs Long-Term Immune Responses to COVID-19 mRNA Vaccine in Patients with Inflammatory Bowel Diseases. Vaccines 2022, 10, 1186. [Google Scholar] [CrossRef]
- Nagy, A.; Mátrai, P.; Hegyi, P.; Alizadeh, H.; Bajor, J.; Czopf, L.; Gyöngyi, Z.; Kiss, Z.; Márta, K.; Simon, M.; et al. The effects of TNF-alpha inhibitor therapy on the incidence of infection in JIA children: A meta-analysis. Pediatr. Rheumatol. 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Ablamunits, V.; Lepsy, C. Blocking TNF signaling may save lives in COVID-19 infection. Mol. Biol. Rep. 2022, 49, 2303–2309. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.; Lado, A.; Blatt, N. Application of Monoclonal Antibody Drugs in Treatment of COVID-19: A Review. Bionanoscience 2022, 12, 1436–1454. [Google Scholar] [CrossRef]
- Bascones-Martinez, A.; Mattila, R.; Gomez-Font, R.; Meurman, J. Immunomodulatory drugs: Oral and systemic adverse effects. Med. Oral Patol. Oral Cir. Bucal 2014, 19, e24–e31. [Google Scholar] [CrossRef]
- Pacha, O.; Sallman, M.A.; Evans, S.E. COVID-19: A case for inhibiting IL-17? Nat. Rev. Immunol. 2020, 20, 345–346. [Google Scholar] [CrossRef]
- A Collier, D.; Ferreira, I.A.T.M.; Kotagiri, P.; Datir, R.; Lim, E.; Touizer, E.; Meng, B.; Abdullahi, A.; CITIID-NIHR BioResource COVID-19 Collaboration; Elmer, A.; et al. Age-related immune response heterogeneity to SARS-CoV-2 vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef]
- Guimarães, L.E.; Baker, B.; Perricone, C.; Shoenfeld, Y. Vaccines, adjuvants and autoimmunity. Pharmacol. Res. 2015, 100, 190–209. [Google Scholar] [CrossRef]
- Chang, S.E.; Feng, A.; Meng, W.; Apostolidis, S.A.; Mack, E.; Artandi, M.; Barman, L.; Bennett, K.; Chakraborty, S.; Chang, I.; et al. New-onset IgG autoantibodies in hospitalized patients with COVID-19. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef]
- Böröcz, K.; Simon, D.; Erdő-Bonyár, S.; Kovács, K.T.; Tuba, É.; Czirják, L.; Németh, P.; Berki, T. Relationship between natural and infection-induced antibodies in systemic autoimmune diseases (SAD): SLE, SSc and RA. Clin. Exp. Immunol. 2020, 203, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Nagele, E.P.; Han, M.; Acharya, N.K.; DeMarshall, C.; Kosciuk, M.C.; Nagele, R.G. Natural IgG Autoantibodies Are Abundant and Ubiquitous in Human Sera, and Their Number Is Influenced by Age, Gender, and Disease. PLoS ONE 2013, 8, e60726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S. Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity. Int. J. Mol. Sci. 2020, 21, 5298. [Google Scholar] [CrossRef] [PubMed]
- Marino Gammazza, A.; Légaré, S.; Lo Bosco, G.; Fucarino, A.; Angileri, F.; Conway de Macario, E.; Macario, A.J.; Cappello, F. Human Molecular Chaperones Share with SARS-CoV-2 Antigenic Epitopes Potentially Capable of Eliciting Autoimmunity against Endothelial Cells: Possible Role of Molecular Mimicry in COVID-19. Cell Stress Chaperones 2020, 25, 737–741. [Google Scholar] [CrossRef]
- Androvitsanea, A.; Stylianou, K.; Drosataki, E.; Petrakis, I. The Pathophysiological Role of Heat Shock Response in Autoimmunity: A Literature Review. Cells 2021, 10, 2626. [Google Scholar] [CrossRef]
- Euroimmun SARS-CoV-2 NeutraLISA Technical Data 2020. EUROIMMUN Product Catalogue 2022. Available online: https://www.euroimmun.com/documents/Catalogue/en/index.html (accessed on 29 March 2023).
- NIBSC. 20/136 NIBSC Medicines and Healthcare Products Regulatory Agency. NIBSC 2020, 8, 147–154. [Google Scholar]
- Euroimmun Test Principle Quan-T-Cell SARS-CoV-2: Stimulation Tube Set Quan-T-Cell ELISA. EUROIMMUN Product Catalogue 2022. Available online: https://www.euroimmun.com/documents/Catalogue/en/index.html (accessed on 29 March 2023).
- NIBSC 87/586, N. NIBSC Medicines and Healthcare Products Regulatory Agency. NIBSC 2013, 16, 2013. [Google Scholar]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J.; et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci. Immunol. 2023, 8. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sero-Positivity Ratios | mRNA Vaccine | Vector Vaccine | Inactivated Virus Vaccine | Unvaccinated and Infected |
|---|---|---|---|---|
| Anti-SARS-CoV-2 IgG | 95.88% | 87.67% | 69.23% | 58.06% |
| Anti-SARS-CoV-2 IgA | 65.38% | 45.21% | 22.58% | 64.95% |
| Sero-Positivity Ratios | mRNA Vaccine | Vector Vaccine | Inactivated Virus Vaccine | Unvaccinated Infected |
|---|---|---|---|---|
| Anti-IFN-γ (IGRA) IgG | 88.12% | 87.01% | 42.42% | 93.10% |
| Neutralizing antibody (NeutraLisa IgA/G/M) | 52.17% | 53.95% | 44.83% | 44.83% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Böröcz, K.; Kinyó, Á.; Simon, D.; Erdő-Bonyár, S.; Németh, P.; Berki, T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. Int. J. Mol. Sci. 2023, 24, 6439. https://doi.org/10.3390/ijms24076439
Böröcz K, Kinyó Á, Simon D, Erdő-Bonyár S, Németh P, Berki T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. International Journal of Molecular Sciences. 2023; 24(7):6439. https://doi.org/10.3390/ijms24076439
Chicago/Turabian StyleBöröcz, Katalin, Ágnes Kinyó, Diana Simon, Szabina Erdő-Bonyár, Péter Németh, and Timea Berki. 2023. "Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies" International Journal of Molecular Sciences 24, no. 7: 6439. https://doi.org/10.3390/ijms24076439
APA StyleBöröcz, K., Kinyó, Á., Simon, D., Erdő-Bonyár, S., Németh, P., & Berki, T. (2023). Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. International Journal of Molecular Sciences, 24(7), 6439. https://doi.org/10.3390/ijms24076439