
Sequence Characteristics and Expression Analysis of the Gene Encoding Sedoheptulose-1,7-Bisphosphatase, an Important Calvin Cycle Enzyme in Upland Cotton (Gossypium hirsutum L.)

Abstract
:1. Introduction
2. Results
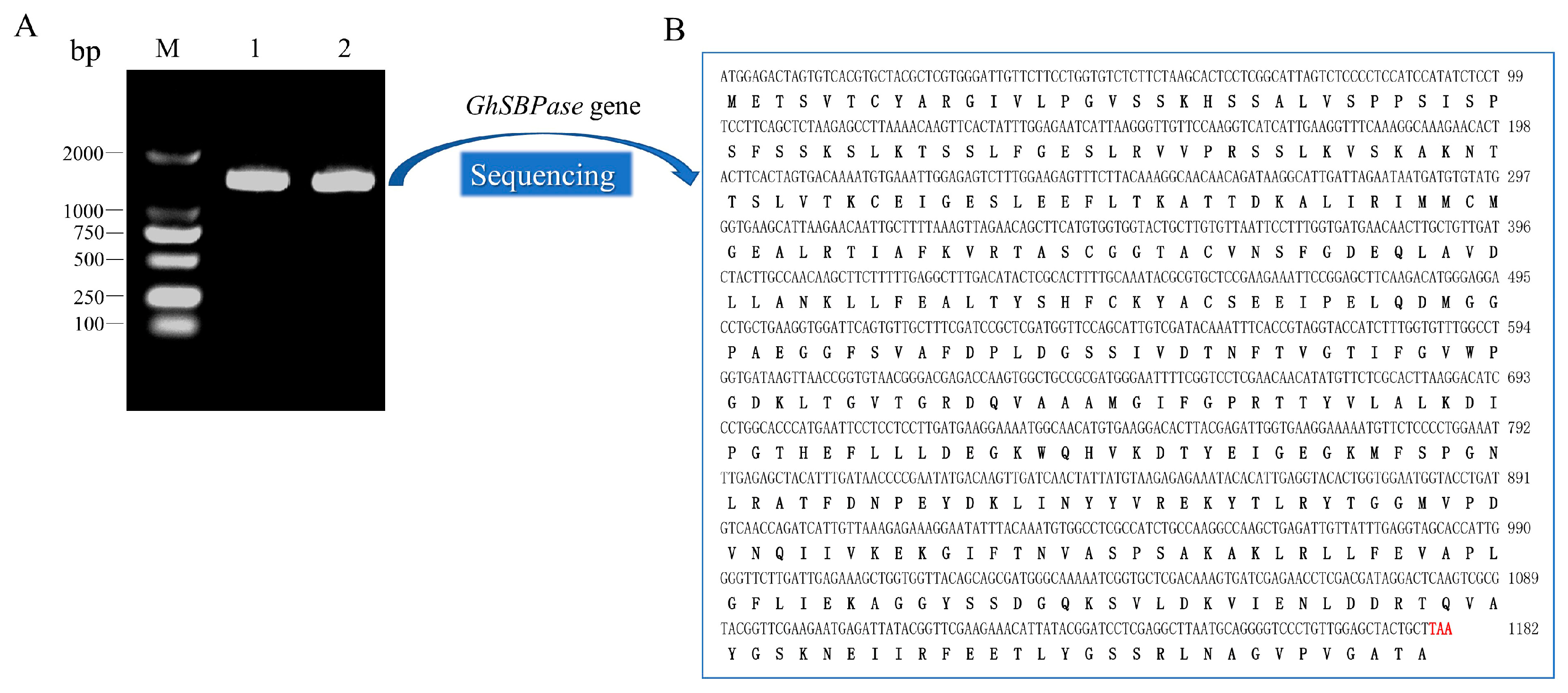
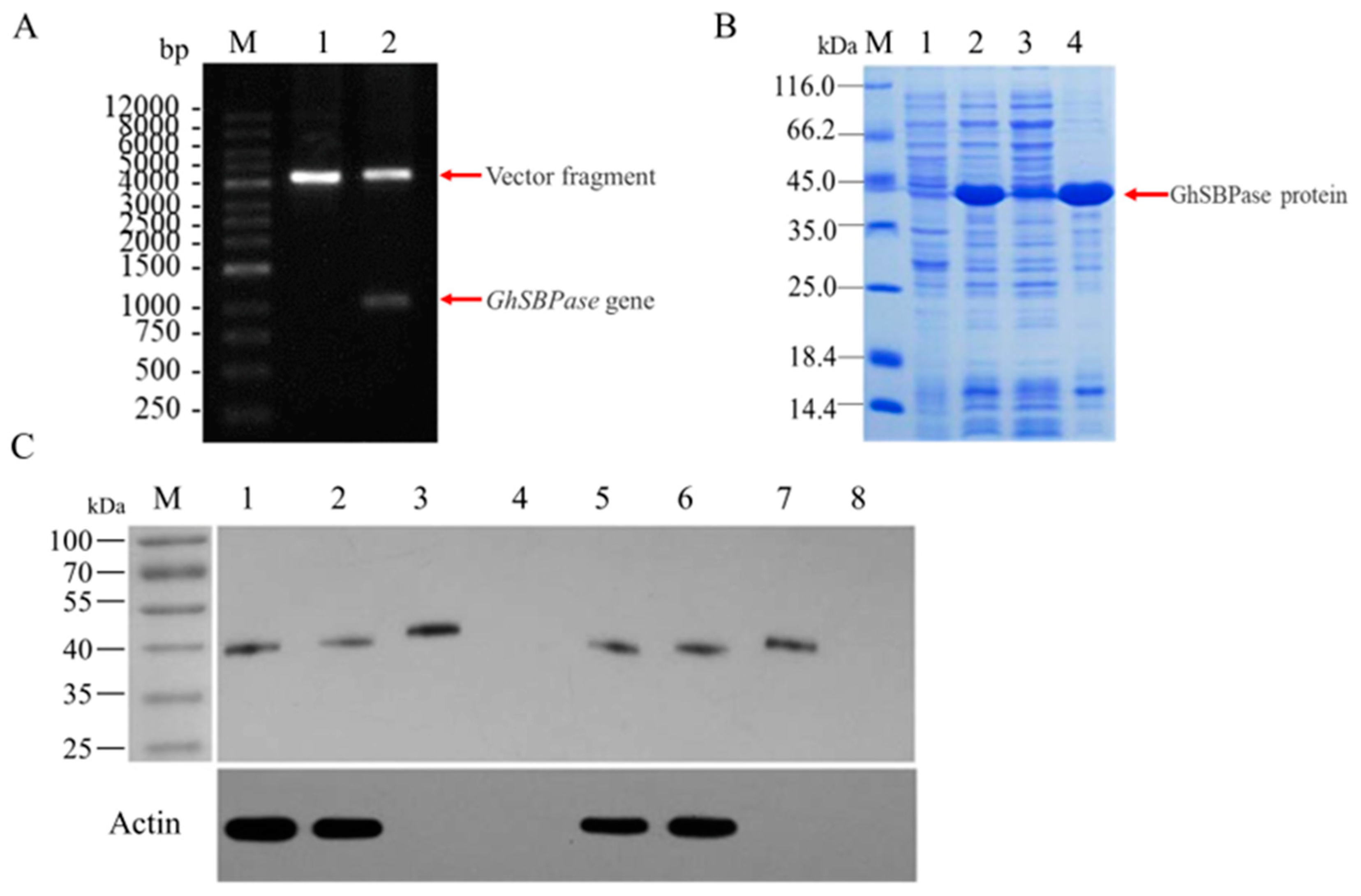
2.1. Identification and Cloning of GhSBPase Gene
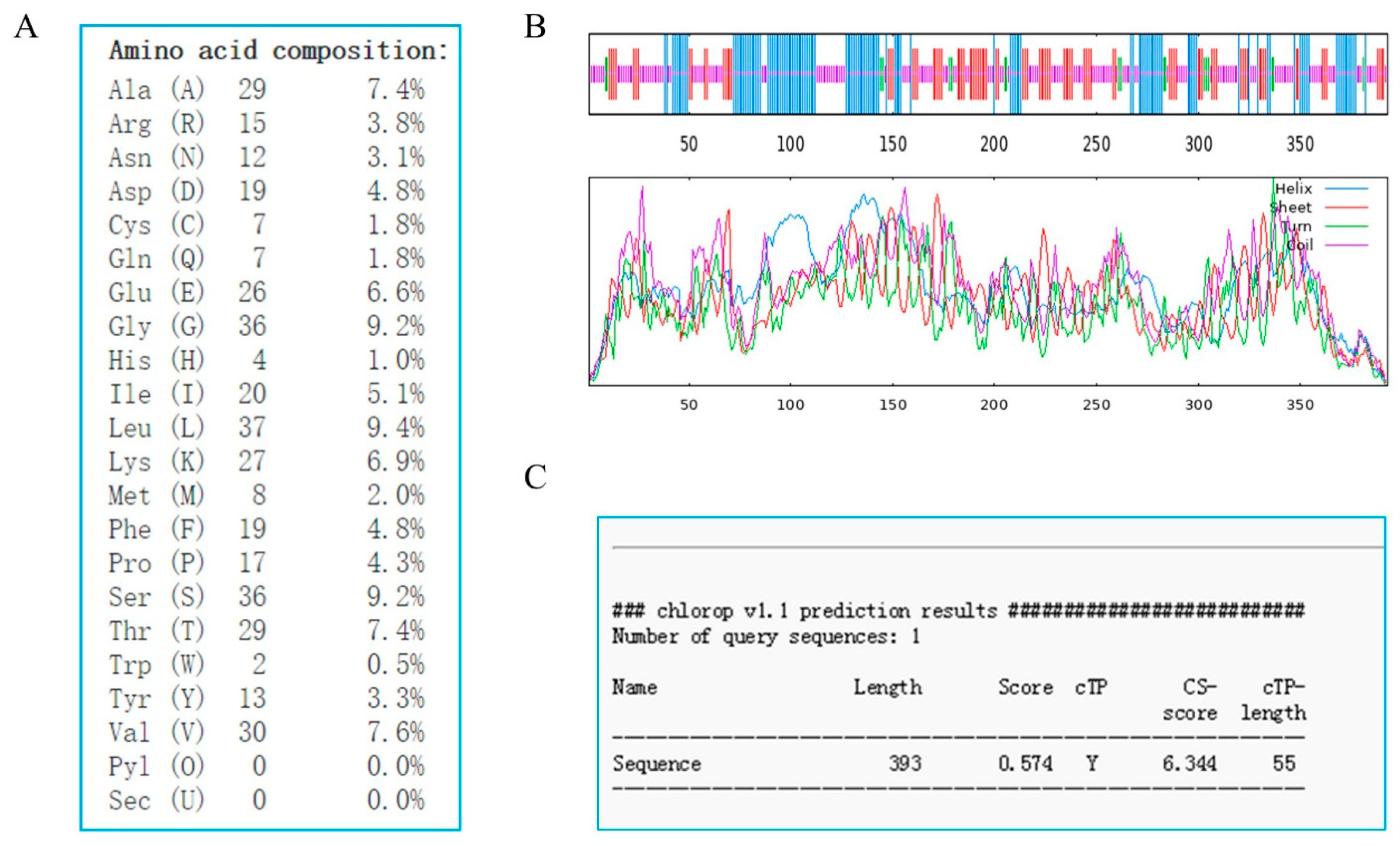
2.2. Sequence Characteristics of GhSBPase Protein
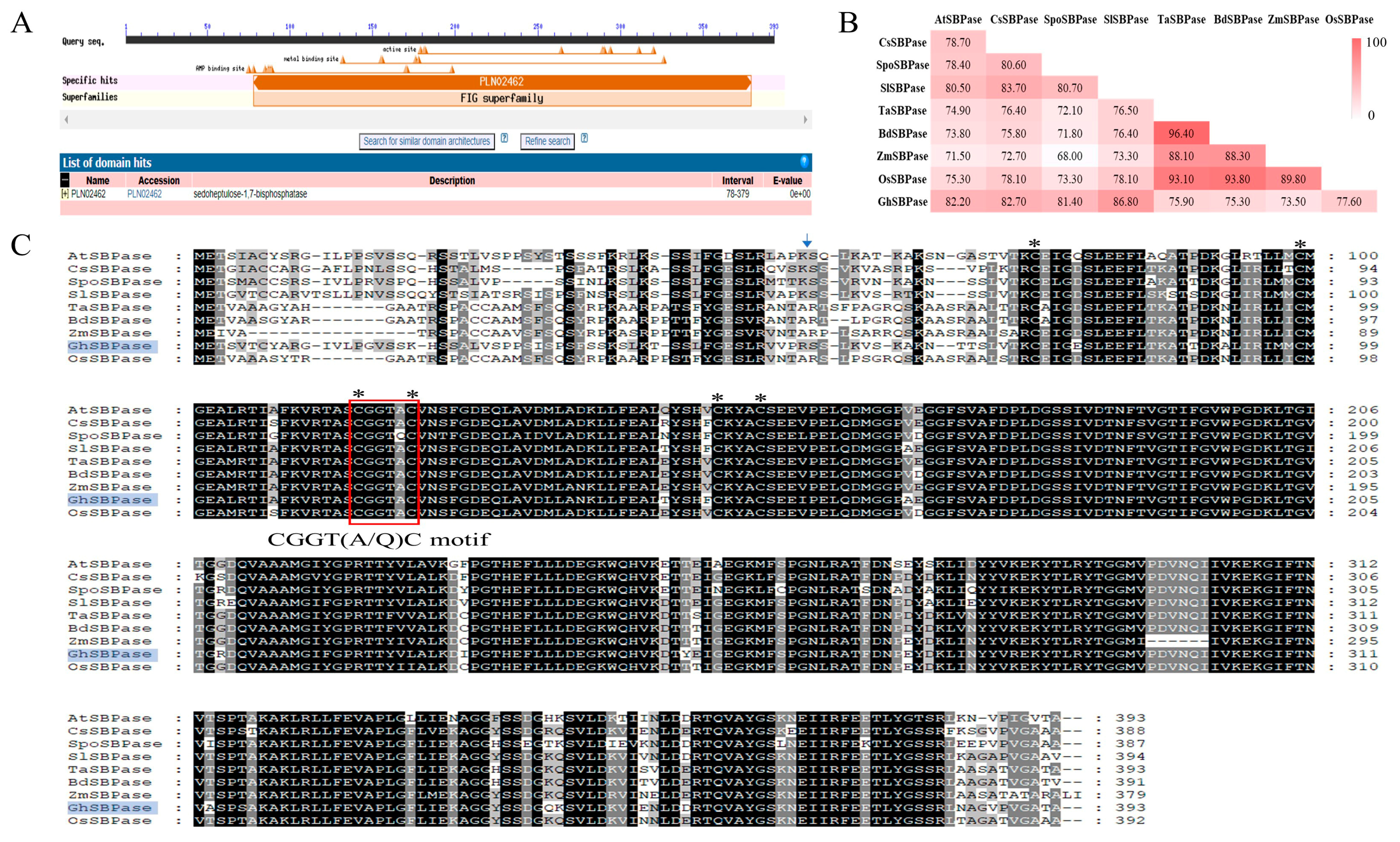
2.3. Conserved Domain Analysis of GhSBPase Protein
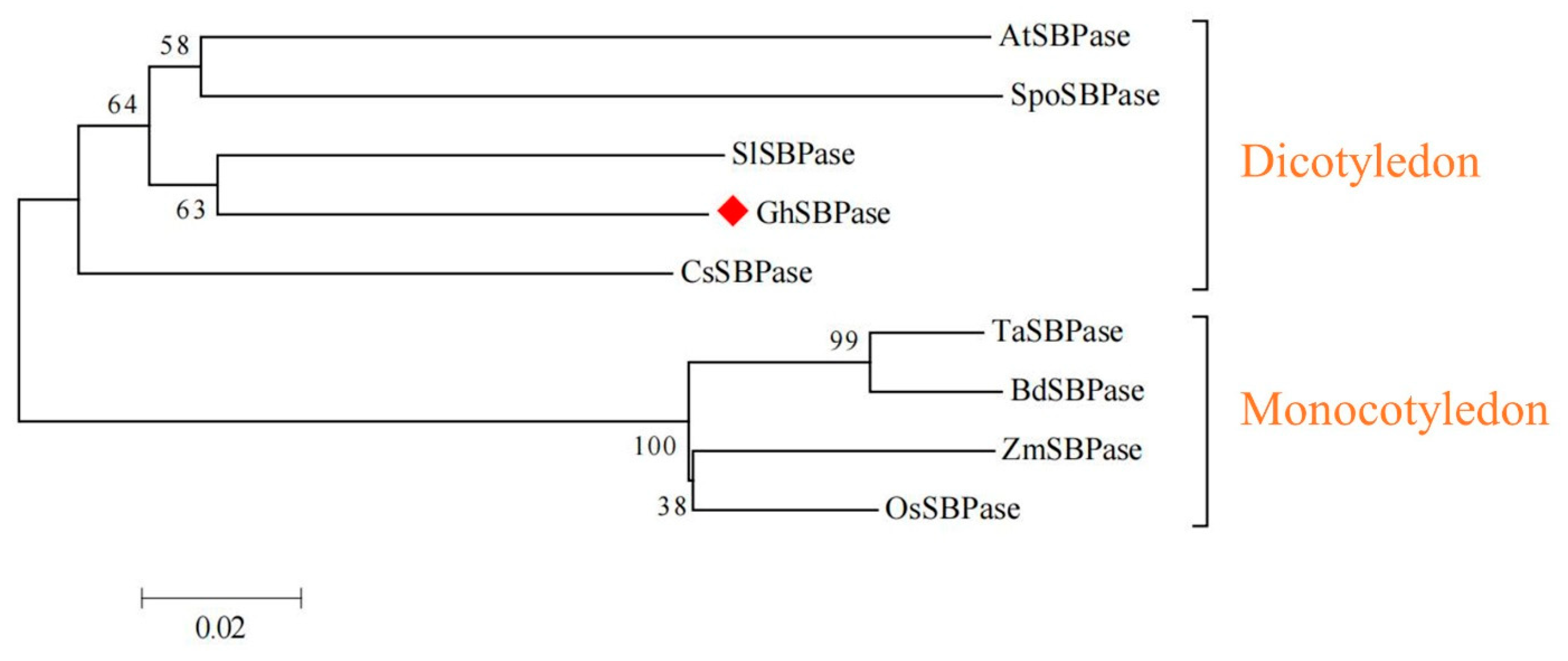
2.4. Evolutionary Relationship Analysis of GhSBPase Protein
2.5. Western-Blot Analysis of GhSBPase Protein
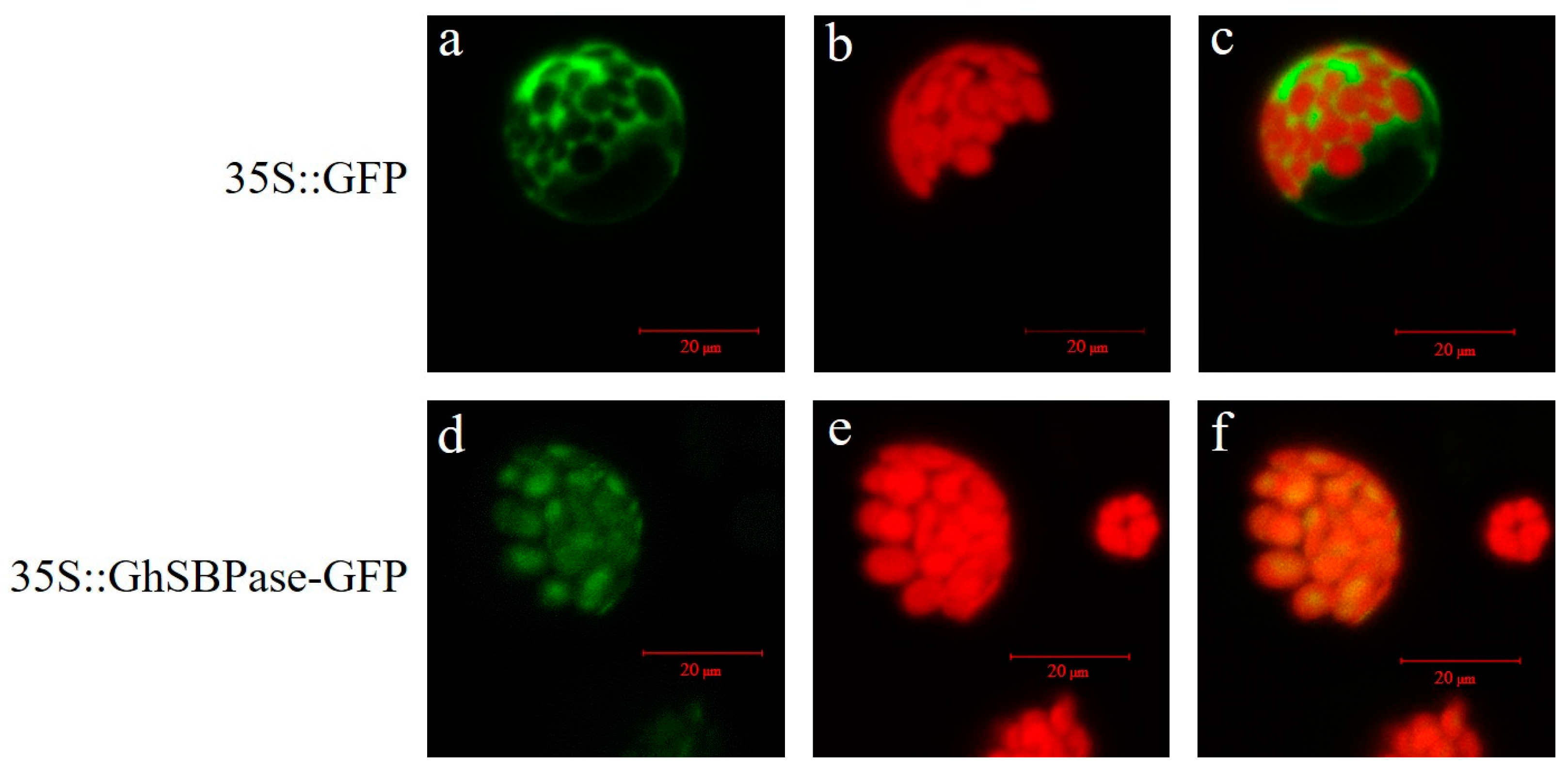
2.6. Subcellular Localization of GhSBPase Protein
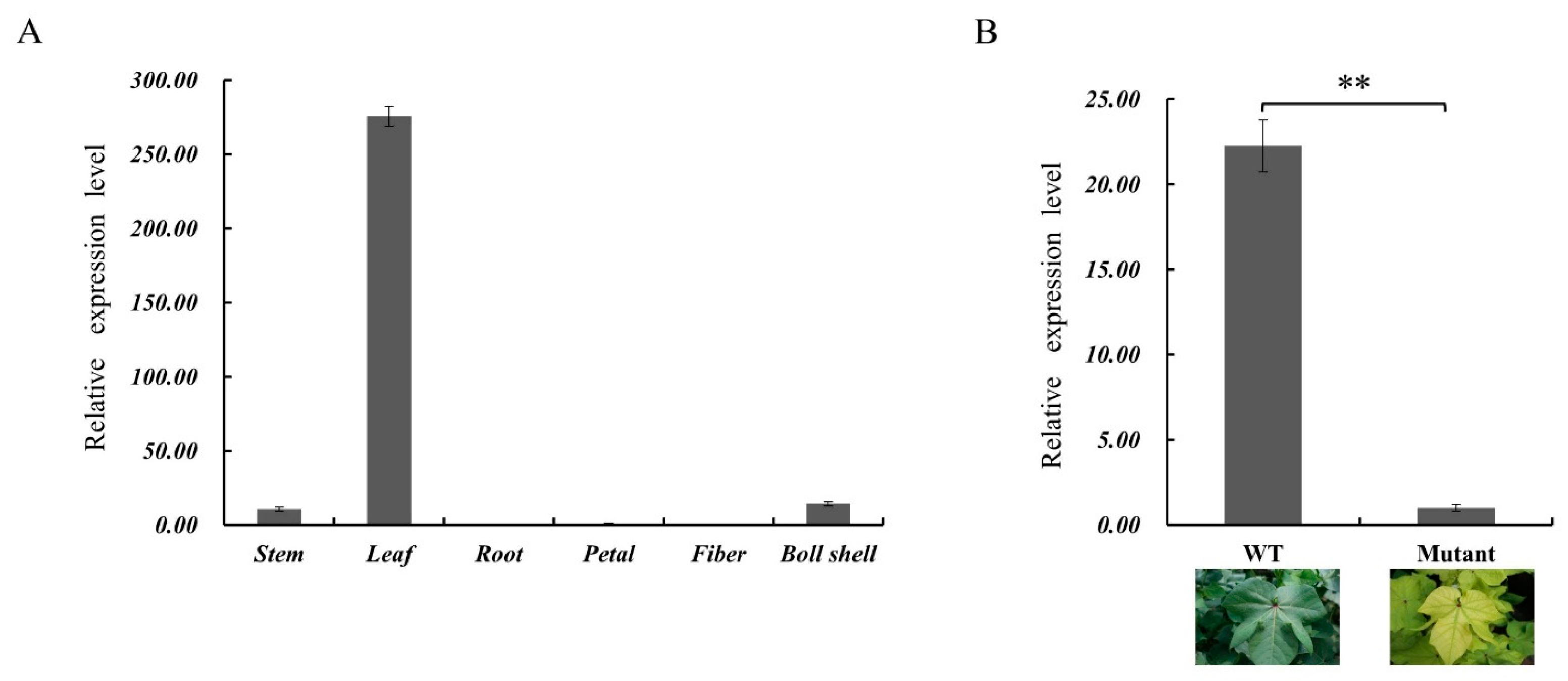
2.7. Tissue Expression Analysis GhSBPase Gene
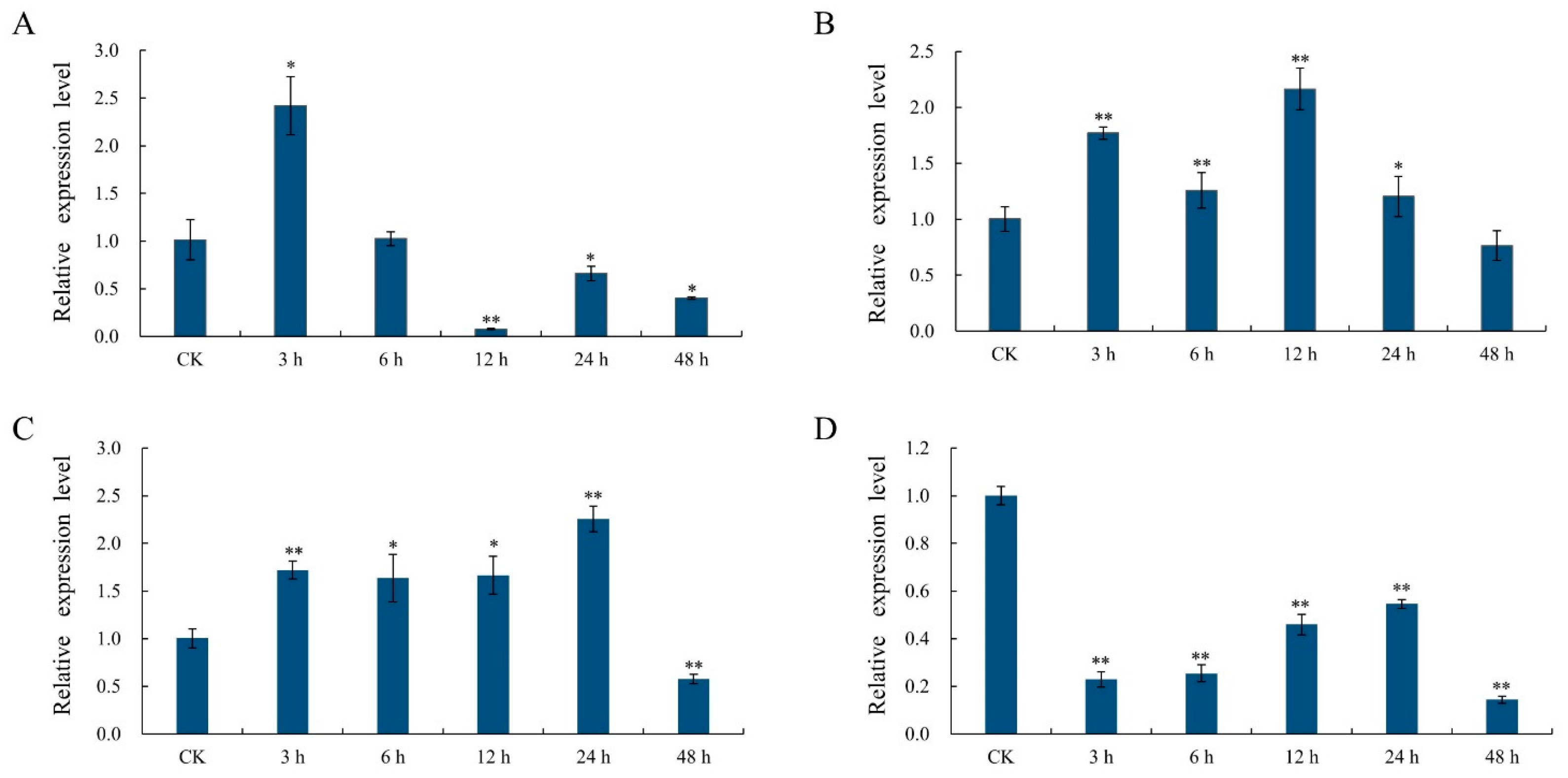
2.8. Analysis of the Response of the GhSBPase Gene to Abiotic Stress
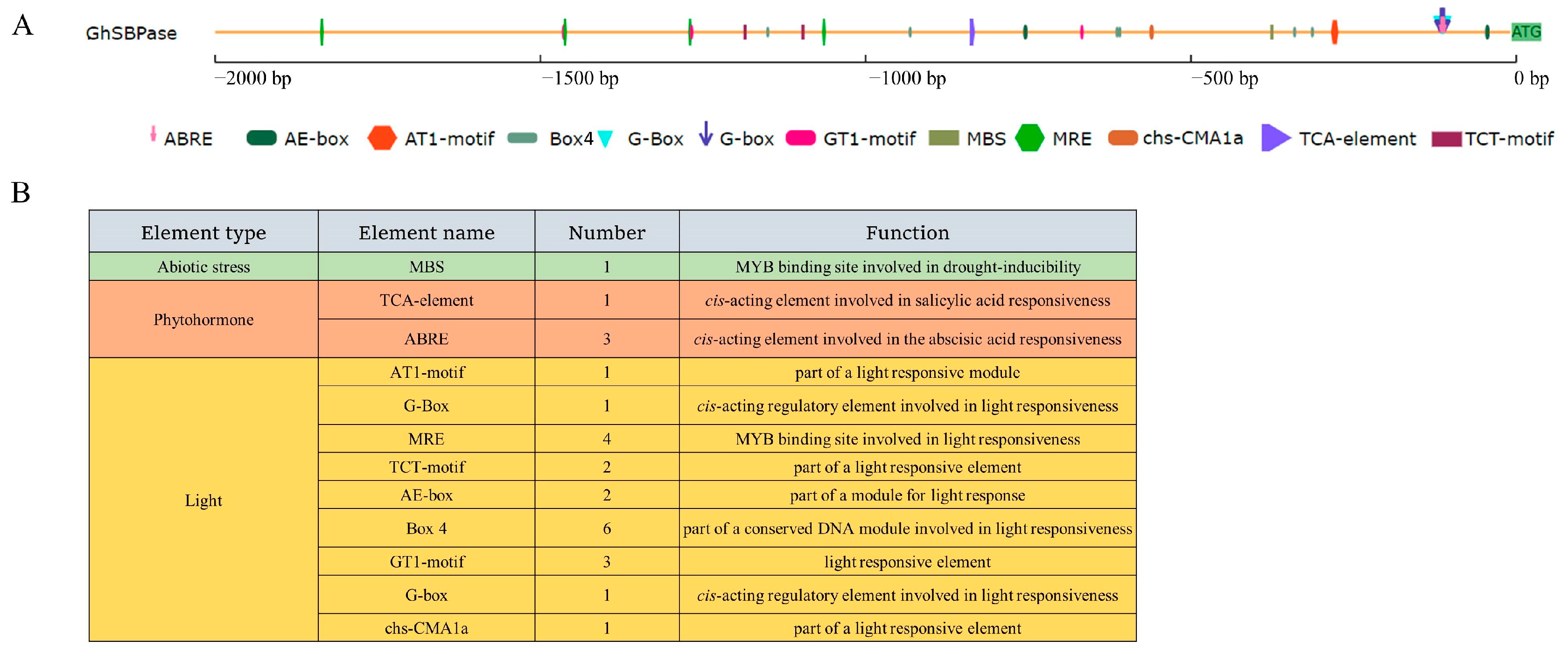
2.9. Cis-Acting Elements in GhSBPase Gene Promoter
2.10. Correlation between GhSBPase Gene Expression and Chlorophyll Fluorescence Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions and Stress Treatment
4.2. RNA Extraction and cDNA Synthesis
4.3. Isolation of GhSBPase Gene from Cotton Leaf
4.4. Sequence Analysis of GhSBPase Protein
4.5. SDS-PAGE and Immunological Analysis
4.6. Subcellular Localization
4.7. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.8. Cis-Acting Elements Analysis in the Promoter Region
4.9. Trait Measurement and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adams IIIW, W.; Cohu, C.M.; Muller, O.; Demmig-Adams, B. Foliar phloem infrastructure in support of photosynthesis. Front. Plant Sci. 2013, 4, 194. [Google Scholar] [CrossRef] [Green Version]
- Dunwell, J.M. Transgenic approaches to crop improvement. J. Exp. Bot. 2000, 51 (Suppl. S1), 487–496. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Purcell, L.C.; Sneller, C.H. Crop transformation and the challenge to increase yield potential. Trends Plant Sci. 2004, 9, 70–75. [Google Scholar] [CrossRef]
- Austin, R.B.; Ford, M.A.; Morgan, C.L. Genetic improvement in the yield of winter wheat: A further evaluation. J. Agric. Sci. 1989, 112, 295–301. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, A.L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Jin, J.; Liu, X.; Wang, G.; Mi, L.; Shen, Z.; Chen, X.; Herbert, S.J. Agronomic and physiological contributions to the yield improvement of soybean cultivars released from 1950 to 2006 in Northeast China. Field Crops Res. 2010, 115, 116–123. [Google Scholar] [CrossRef]
- Raines, C.A. Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: Current and future strategies. Plant Physiol. 2011, 155, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, S.; Ohkubo, S.; San, N.S.; Yamamoto, T. Genetic determination for source capacity to support breeding of high-yielding rice (Oryza sativa). Mol. Breed. 2020, 40, 1226–1235. [Google Scholar] [CrossRef]
- Batista-Silva, W.; da Fonseca-Pereira, P.; Martins, A.O.; Zsögön, A.; Nunes-Nesi, A.; Araújo, W.L. Engineering improved photosynthesis in the era of synthetic biology. Plant Commun. 2020, 1, 100032. [Google Scholar] [CrossRef]
- Geiger, D.R.; Servaites, J.C. Diurnal regulation of photosynthetic carbon metabolism in C3 plants. Annu. Rev. Plant Biol. 1994, 45, 235–256. [Google Scholar] [CrossRef]
- Raines, C.A.; Lloyd, J.C.; Dyer, T.A. New insights into the structure and function of sedoheptulose-1, 7-bisphosphatase; an important but neglected Calvin cycle enzyme. J. Exp. Bot. 1999, 50, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Raines, C.A. The Calvin cycle revisited. Photosynth. Res. 2003, 75, 1–10. [Google Scholar] [CrossRef]
- Devos, K.M.; Atkinson, M.D.; Chinoy, C.N.; Lloyd, J.C.; Raines, C.A.; Dyer, T.A.; Gale, M.D. The coding sequence for sedoheptulose-1,7-bisphosphatase detects multiple homologues in wheat genomic DNA. Theor. Appl. Genet. 1992, 85, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Willingham, N.M.; Lloyd, J.C.; Raines, C.A. Molecular cloning of the Arabidopsis thaliana sedoheptulose-1,7-biphosphatase gene and expression studies in wheat and Arabidopsis thaliana. Plant Mol. Biol. 1994, 26, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Cadet, F.; Meunier, J.C. Spinach (Spinacia oleracea) chloroplast sedoheptulose-1,7-bisphosphatase. Activation and deactivation, and immunological relationship to fructose-1,6-bisphosphatase. Biochem. J. 1988, 253, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Gai, Y.; Ma, J.; Zheng, C.; Mu, Z. Molecular cloning and characterization of a cDNA encoding sedoheptulose-1, 7-bisphosphatase from mulberry (Morus alba var. multicaulis). Silvae Genet. 2008, 57, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Bi, H.; Liu, P.; Ai, X. Molecular cloning and expression analysis of the gene encoding sedoheptulose-1, 7-bisphosphatase from Cucumis sativus. Sci. Hortic. 2011, 129, 414–420. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, J.; Yu, T.; Li, X.; Li, S.; Hua, Y.; Li, Y.; Zhu, Y. Molecular cloning and characterization of rice sedoheptulose-1, 7-bisphosphatase gene that is regulated by environmental stresses. J. Plant Biochem. Biotechnol. 2004, 13, 93–99. [Google Scholar] [CrossRef]
- Harrison, E.P.; Willingham, N.M.; Lloyd, J.C.; Raines, C.A. Reduced sedoheptulose-1, 7-bisphosphatase levels in transgenic tobacco lead to decreased photosynthetic capacity and altered carbohydrate accumulation. Planta. 1997, 204, 27–36. [Google Scholar] [CrossRef]
- Raines, C.A.; Harrison, E.P.; Ölçer, H.; Lloyd, J.C. Investigating the role of the thiol-regulated enzyme sedoheptulose-1, 7-bisphosphatase in the control of photosynthesis Physiol. Plant 2000, 110, 303–308. [Google Scholar] [CrossRef]
- Olçer, H.; Lloyd, J.C.; Raines, C.A. Photosynthetic capacity is differentially affected by reductions in sedoheptulose-1,7-bisphosphatase activity during leaf development in transgenic tobacco plants. Plant Physiol. 2001, 125, 982–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, T.; Bryant, B.; Lefebvre, S.; Lloyd, J.C.; Raines, C.A. Decreased SBPase activity alters growth and development in transgenic tobacco plants. Plant Cell Environ. 2006, 29, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Li, H.; Jiao, J.; Li, D.; Zhou, L.; Wan, J.; Li, Y. Reduction in SBPase activity by antisense RNA in transgenic rice plants: Effect on photosynthesis, growth, and biomass allocation at different nitrogen levels. J. Plant Biol. 2009, 52, 382–394. [Google Scholar] [CrossRef]
- Lefebvre, S.; Lawson, T.; Zakhleniuk, O.V.; Lloyd, J.C.; Raines, C.A.; Fryer, M. Increased sedoheptulose-1,7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 2005, 138, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Raines, C.A. Transgenic approaches to manipulate the environmental responses of the C3 carbon fixation cycle. Plant Cell Environ. 2006, 29, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, L.; Liang, C.; Meng, Z.; Tahira, S.; Guo, S.; Zhang, R. Overexpression of Brassica napus cytosolic fructose-1, 6-bisphosphatase and sedoheptulose-1, 7-bisphosphatase genes significantly enhanced tobacco growth and biomass. J. Integr. Agric. 2022, 21, 49–59. [Google Scholar] [CrossRef]
- Driever, S.M.; Simkin, A.J.; Alotaibi, S.; Fisk, S.J.; Madgwick, P.J.; Sparks, C.A.; Jones, H.D.; Lawson, T.; Parry MA, J.; Raines, C.A. Increased SBPase activity improves photosynthesis and grain yield in wheat grown in greenhouse conditions. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160384. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.Y.; Li, Y.; Fang, G.; Hu, D.H.; Jin, W.B.; Wang, Z.H.; Li, Y.S. Transgenic rice expressing Ictb and FBP/Sbpase derived from Cyanobacteria exhibits enhanced photosynthesis and mesophyll conductance to CO2. PLoS ONE 2015, 10, e0140928. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.Y.; Li, Y.; Fang, G.; Hu, D.H.; Wang, Z.H.; Yu, G.H.; Li, Y.S. ccaA and FBP/SBPase derived from cyanobacteria improved photosynthetic capacity in rice. Plant Mol. Biol. Report. 2017, 35, 619–633. [Google Scholar] [CrossRef]
- Feng, L.; Han, Y.; Liu, G.; An, B.; Yang, J.; Yang, G.; Li, Y.; Zhu, Y. Overexpression of sedoheptulose-1,7-bisphosphatase enhances photosynthesis and growth under salt stress in transgenic rice plants. Funct. Plant Biol. 2007, 34, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, K.; Li, Y.; Tan, Y.; Kong, J.; Li, H.; Li, Y.; Zhu, Y. Overexpression of SBPase enhances photosynthesis against high temperature stress in transgenic rice plants. Plant Cell Rep. 2007, 26, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wang, M.; Zhang, S.; Ai, X. Changes in SBPase activity influence photosynthetic capacity, growth, and tolerance to chilling stress in transgenic tomato plants. Sci. Rep. 2016, 6, 32741. [Google Scholar] [CrossRef] [Green Version]
- Ding, F.; Wang, M.; Zhang, S. Overexpression of a Calvin cycle enzyme SBPase improves tolerance to chilling-induced oxidative stress in tomato plants. Sci. Hortic. 2017, 214, 27–33. [Google Scholar] [CrossRef]
- Köhler, I.H.; Ruiz-Vera, U.M.; VanLoocke, A.; Thomey, M.L.; Clemente, T.; Long, S.P.; Ort, D.R.; Bernacchi, C.J. Expression of cyanobacterial FBP/SBPase in soybean prevents yield depression under future climate conditions. J. Exp. Bot. 2017, 68, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants FACE the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef]
- Gao, H.; Li, N.; Li, J.; Khan, A.; Ahmad, I.; Wang, Y.; Wang, F.; Luo, H. Improving boll capsule wall, subtending leaves anatomy and photosynthetic capacity can increase seed cotton yield under limited drip irrigation systems. Ind. Crops Prod. 2021, 161, 113214. [Google Scholar] [CrossRef]
- Simkin, A.J.; López-Calcagno, P.E.; Raines, C.A. Feeding the world: Improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 2019, 70, 1119–1140. [Google Scholar] [CrossRef] [Green Version]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean photosynthesis and crop yield are improved by accelerating recovery from photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef]
- Raines, C.A. Improving plant productivity by re-tuning the regeneration of RuBP in the Calvin-Benson-Bassham cycle. New Phytol. 2022, 236, 350–356. [Google Scholar] [CrossRef]
- Li, C.; Li, N.; Huang, R.; Chen, C.; Guo, J.; Yang, X.; Zhang, X.; Sun, C.; Deng, X.; Wang, P. A single nucleotide substitution at the 3'-end of SBPase gene involved in Calvin cycle severely affects plant growth and grain yield in rice. BMC Plant Biol. 2020, 20, 345. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Yu, H.D.; Guan, Y.; Li, J.K.; Guo, F.Q. Carbonylation and loss-of-function analyses of SBPase reveal its metabolic interface role in oxidative stress, carbon assimilation, and multiple aspects of growth and development in Arabidopsis. Mol. Plant 2012, 5, 1082–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, I.C.; Lee, K.H.; Nahm, B.H.; Kim, J.K. Chloroplast targeting signal of a rice rbcS gene enhances transgene expression. Mol. Breed. 2002, 9, 81–91. [Google Scholar] [CrossRef]
- Wong, E.Y.; Hironaka, C.M.; Fischhoff, D.A. Arabidopsis thaliana small subunit leader and transit peptide enhance the expression of Bacillus thuringiensis proteins in transgenic plants. Plant Mol. Biol. 1992, 20, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Potts, S.C.; Willingham, N.M.; Raines, C.A.; Lloyd, J.C. A light- and developmentally-regulated DNA-binding interaction is common to the upstream sequences of the wheat Calvin cycle bisphosphatase genes. Plant Mol. Biol. 1993, 22, 507–516. [Google Scholar] [CrossRef]
- Jones, P.G.; Lloyd, J.C.; Raines, C.A. Glucose feeding of intact wheat plants represses the expression of a number of Calvin cycle genes. Plant Cell Environ. 1996, 19, 231–236. [Google Scholar] [CrossRef]
- Alotaibi, S.S. Developing specific leaf promoters tools for genetic use in transgenic plants towards food security. Saudi J. Biol. Sci. 2021, 28, 5187–5192. [Google Scholar] [CrossRef] [PubMed]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Sekhar, S.; Panda, D.; Kumar, J.; Mohanty, N.; Biswal, M.; Baig, M.J.; Kumar, A.; Umakanta, N.; Samantaray, S.; Pradhan, S.K.; et al. Comparative transcriptome profiling of low light tolerant and sensitive rice varieties induced by low light stress at active tillering stage. Sci. Rep. 2019, 9, 5753. [Google Scholar] [CrossRef] [Green Version]
- Ding, F.; Wang, M.; Zhang, S. Sedoheptulose-1,7-Bisphosphatase is involved in methyl jasmonate- and dark-induced leaf senescence in tomato plants. Int. J. Mol. Sci. 2018, 19, 3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alotaibi, S.; Alyasi, H.; El-Shehawi, A.; Gaber, A.; Hassan, M.; Aljuaid, B.; Simkin, A.; Raines, C. Functional analysis of SBPase gene promoter in transgenic wheat under abiotic stresses. Biotechnology 2019, 18, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76–85. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Zhang, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat. Plant Growth Regul. 2020, 90, 109–121. [Google Scholar] [CrossRef]
- Sagervanshi, A.; Naeem, A.; Geilfus, C.M.; Kaiser, H.; Mühling, K.H. One-time abscisic acid priming induces long-term salinity resistance in Vicia faba: Changes in key transcripts, metabolites, and ionic relations. Physiol Plant 2021, 172, 146–161. [Google Scholar] [CrossRef]
- Sherin, G.; Aswathi, K.P.R.; Puthur, J.T. Photosynthetic functions in plants subjected to stresses are positively influenced by priming. Plant Stress 2022, 4, 100079. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, L.; Zhou, J.; Zhong, X.; Gu, F.; Liu, Q.; Li, H.; Guo, R. iTRAQ protein profile analysis of young and old leaves of cotton (Gossypium hirsutum L.) differing in response to alkali stress. Plant Physiol. Biochem. 2019, 141, 370–379. [Google Scholar] [CrossRef]
- Song, J.; Liu, X.; Li, X.; Wang, H.; Chu, R.; Qu, F.; Zhang, S.; Li, Q. Transcriptome analysis reveals genes and pathways associated with salt tolerance during seed germination in Suaeda liaotungensis. Int. J. Mol. Sci. 2022, 23, 12229. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.D.; Jian, G.H.; Wang, X.N.; Mo, Z.G.; Peng, Q.Y.; Chen, S.; Mo, Y.J.; Zhou, S.X.; Huang, Y.X.; Ling, Y. High temperature-mediated disturbance of carbohydrate metabolism and gene expressional regulation in rice: A review. Plant Signal. Behav. 2021, 16, 1862564. [Google Scholar] [CrossRef]
- Xie, Q.; Zhang, Y.; Cheng, Y.; Tian, Y.; Luo, J.; Hu, Z.; Chen, G. The role of melatonin in tomato stress response, growth and development. Plant Cell Rep. 2022, 41, 1631–1650. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Salvucci, M.E. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc Natl Acad Sci. USA 2000, 97, 13430–13435. [Google Scholar] [CrossRef] [Green Version]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of photosynthesis in a C4 plant, maize, to heat stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierling, E.; Key, J.L. Ribulose 1,5-bisphosphate carboxylase synthesis during heat shock. Plant Physiol. 1985, 78, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Law, R.D.; Crafts-Brandner, S.J. High temperature stress increases the expression of wheat leaf ribulose-1,5-bisphosphate carboxylase/oxygenase activase protein. Arch. Biochem. Biophys. 2001, 386, 261–267. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. 1994, 41, 95–98. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, Y.; Chen, P.; Zhou, J.; Zhang, C.; Song, Z.; Huo, X.; Du, Z.; Gong, J.; Zhao, C.; et al. Genome-wide association study reveals novel quantitative trait loci and candidate genes of lint percentage in upland cotton based on the CottonSNP80K array. Theor. Appl. Genet. 2022, 135, 2279–2295. [Google Scholar] [CrossRef]
- Yin, Z.; Meng, F.; Song, H.; He, X.; Xu, X.; Yu, D. Mapping quantitative trait loci associated with chlorophyll a fluorescence parameters in soybean (Glycine max (L.) Merr.). Planta 2010, 231, 875–885. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | ΦPSII | qP | NPQ |
|---|---|---|---|
| GhSBPase expression | 0.076 | 0.248 ** | −0.126 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, M.; Hu, G.; Dong, J.; Chen, Y.; Fu, Y.; Zhang, J.; Wang, Q. Sequence Characteristics and Expression Analysis of the Gene Encoding Sedoheptulose-1,7-Bisphosphatase, an Important Calvin Cycle Enzyme in Upland Cotton (Gossypium hirsutum L.). Int. J. Mol. Sci. 2023, 24, 6648. https://doi.org/10.3390/ijms24076648
Chao M, Hu G, Dong J, Chen Y, Fu Y, Zhang J, Wang Q. Sequence Characteristics and Expression Analysis of the Gene Encoding Sedoheptulose-1,7-Bisphosphatase, an Important Calvin Cycle Enzyme in Upland Cotton (Gossypium hirsutum L.). International Journal of Molecular Sciences. 2023; 24(7):6648. https://doi.org/10.3390/ijms24076648
Chicago/Turabian StyleChao, Maoni, Genhai Hu, Jie Dong, Yu Chen, Yuanzhi Fu, Jinbao Zhang, and Qinglian Wang. 2023. "Sequence Characteristics and Expression Analysis of the Gene Encoding Sedoheptulose-1,7-Bisphosphatase, an Important Calvin Cycle Enzyme in Upland Cotton (Gossypium hirsutum L.)" International Journal of Molecular Sciences 24, no. 7: 6648. https://doi.org/10.3390/ijms24076648
APA StyleChao, M., Hu, G., Dong, J., Chen, Y., Fu, Y., Zhang, J., & Wang, Q. (2023). Sequence Characteristics and Expression Analysis of the Gene Encoding Sedoheptulose-1,7-Bisphosphatase, an Important Calvin Cycle Enzyme in Upland Cotton (Gossypium hirsutum L.). International Journal of Molecular Sciences, 24(7), 6648. https://doi.org/10.3390/ijms24076648