
The Role and Mechanism of Hydrogen-Rich Water in the Cucumis sativus Response to Chilling Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
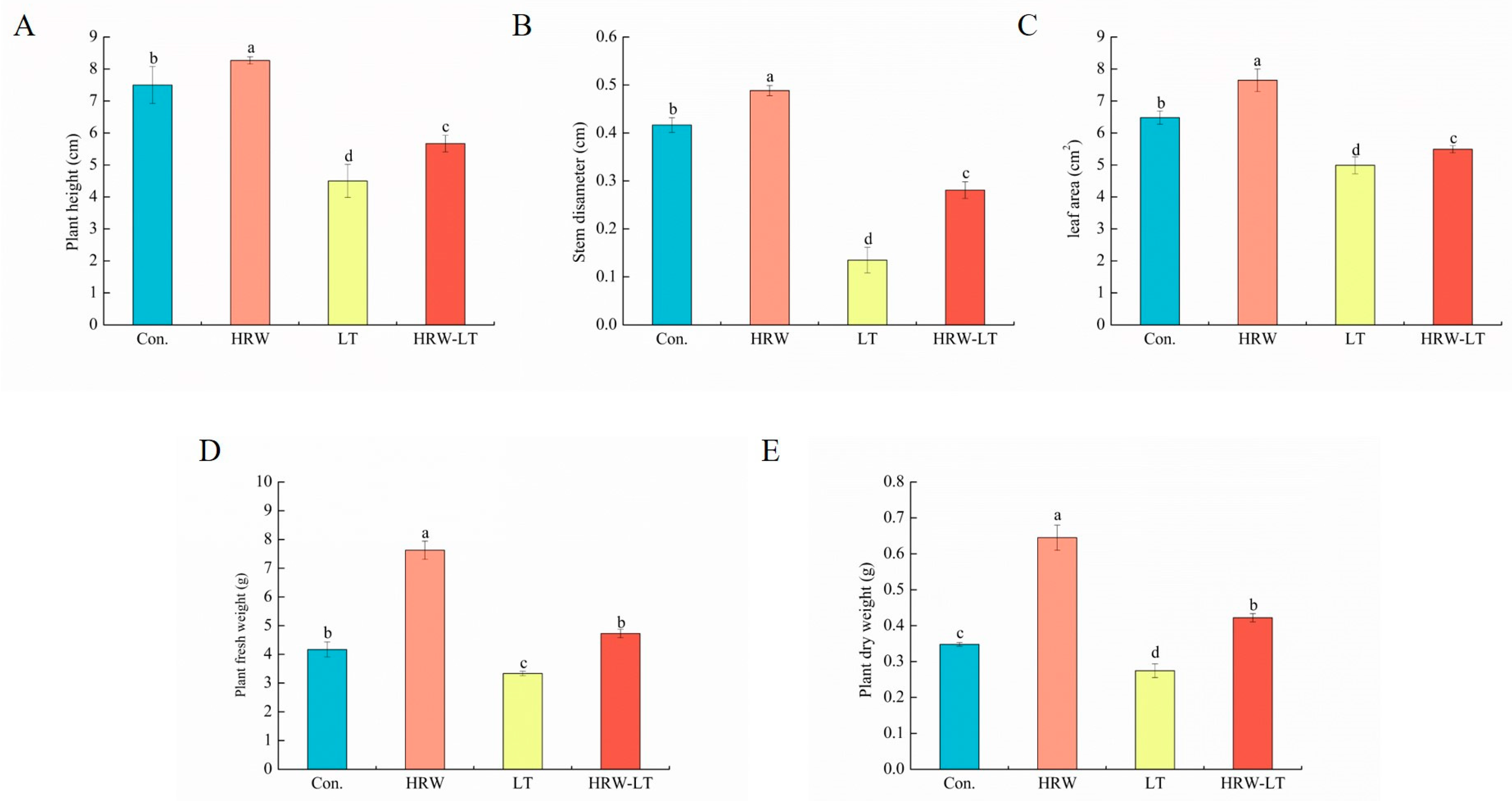
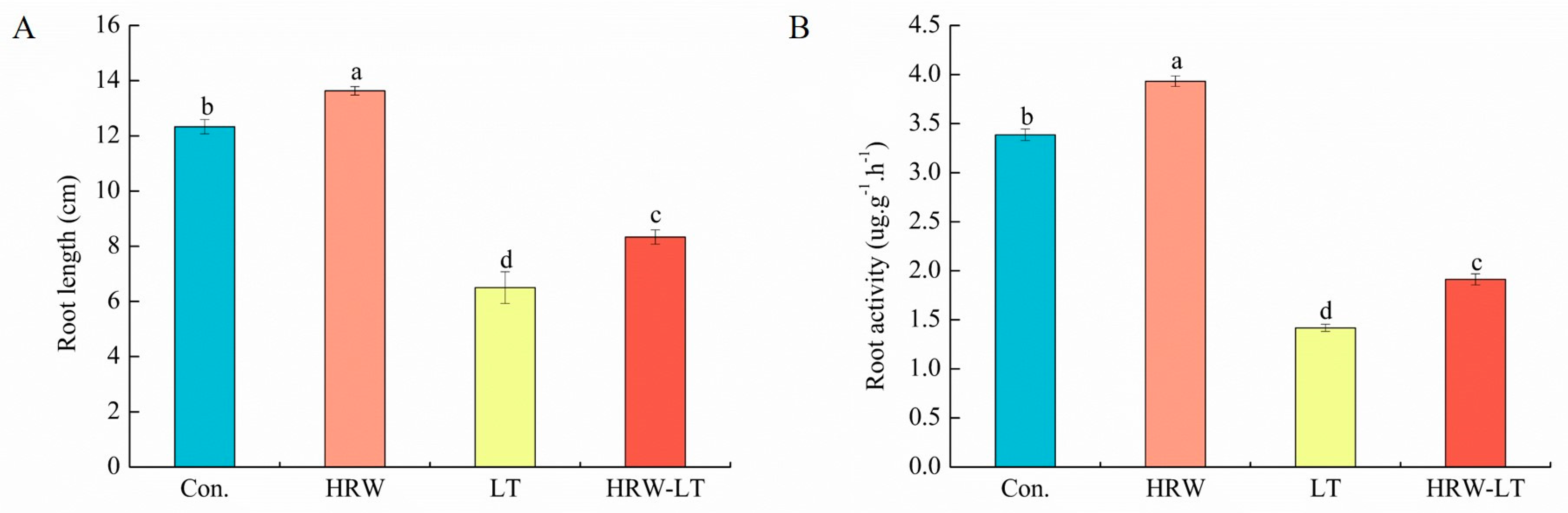
2.1. HRW Can Enhance Cucumber Growth and Development under Chilling Stress
2.2. Effect of HRW on Photosynthesis of Cucumber Seedlings under Chilling Stress
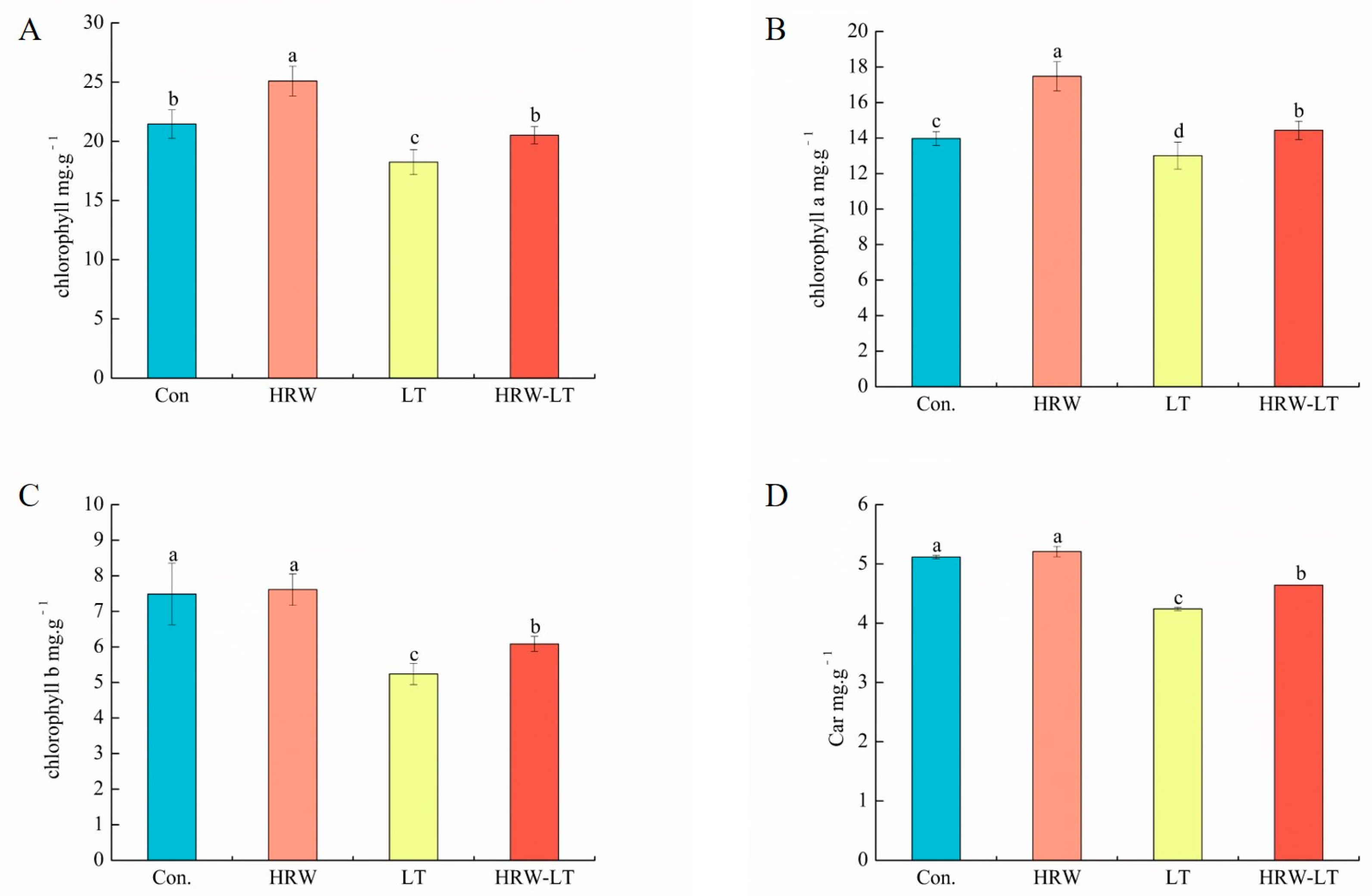
2.2.1. Effect on Pigment Content
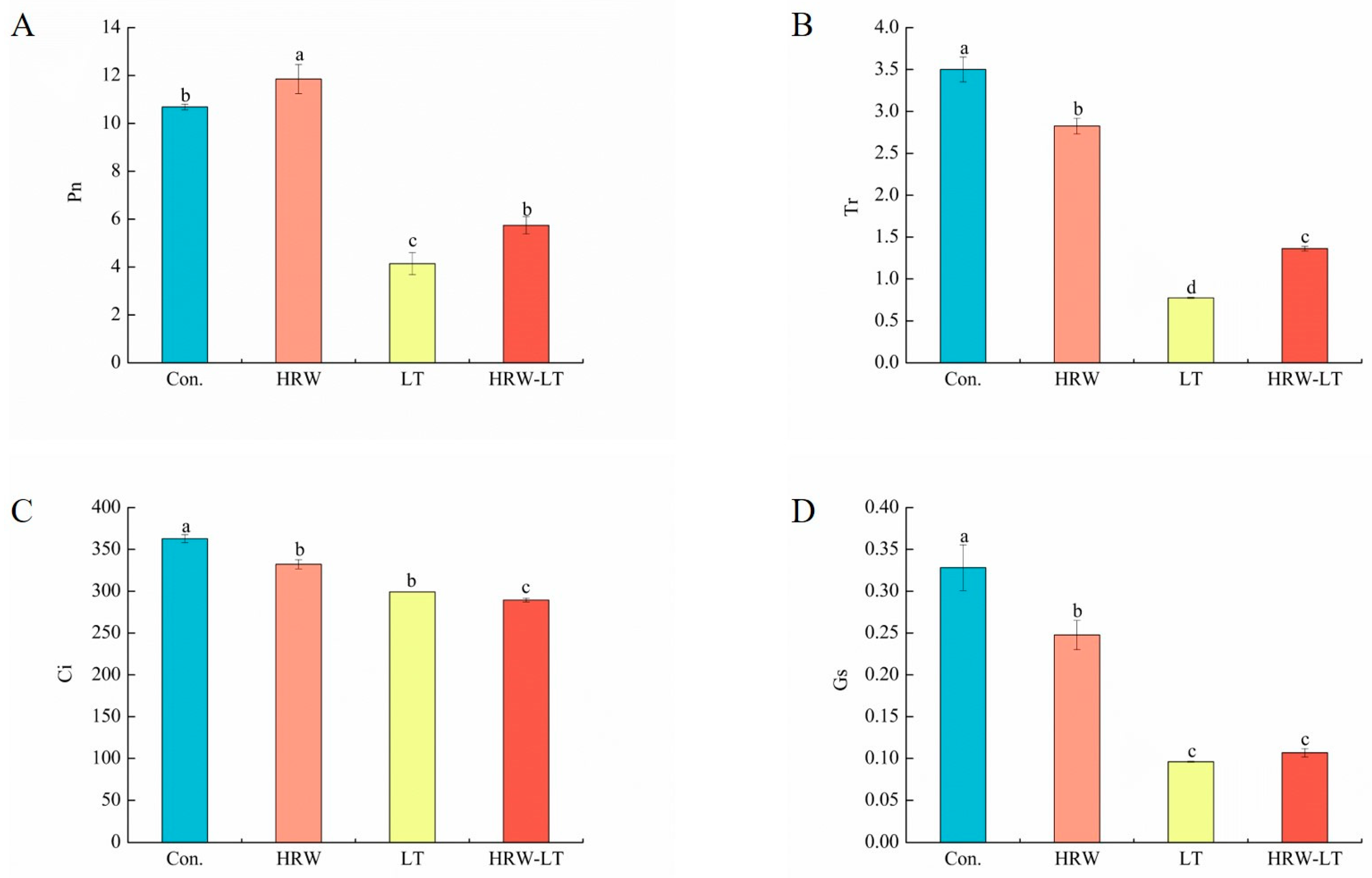
2.2.2. Effect of Gas Exchange Parameters
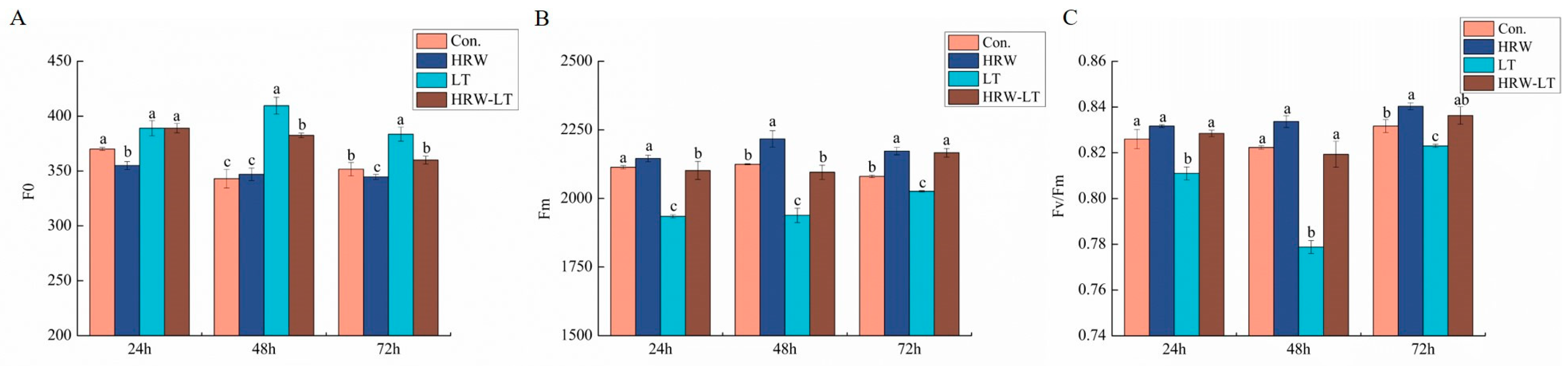
2.3. Effect of HRW on the Fluorescence Parameters of Cucumber Seedlings under Chilling Stress
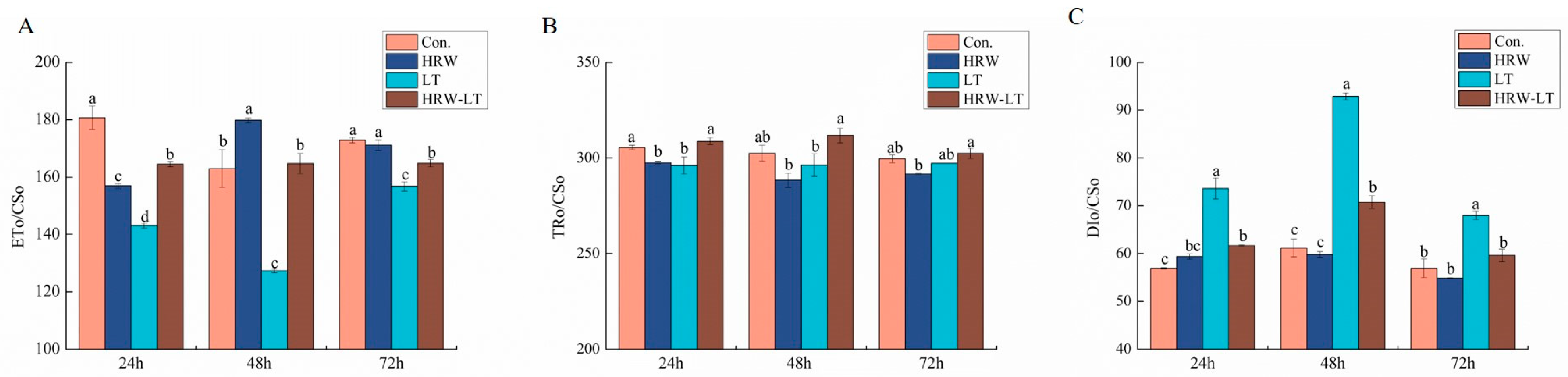
2.4. Effect of HRW on Energy Flow Parameters of Cucumber Seedlings under Chilling Stress
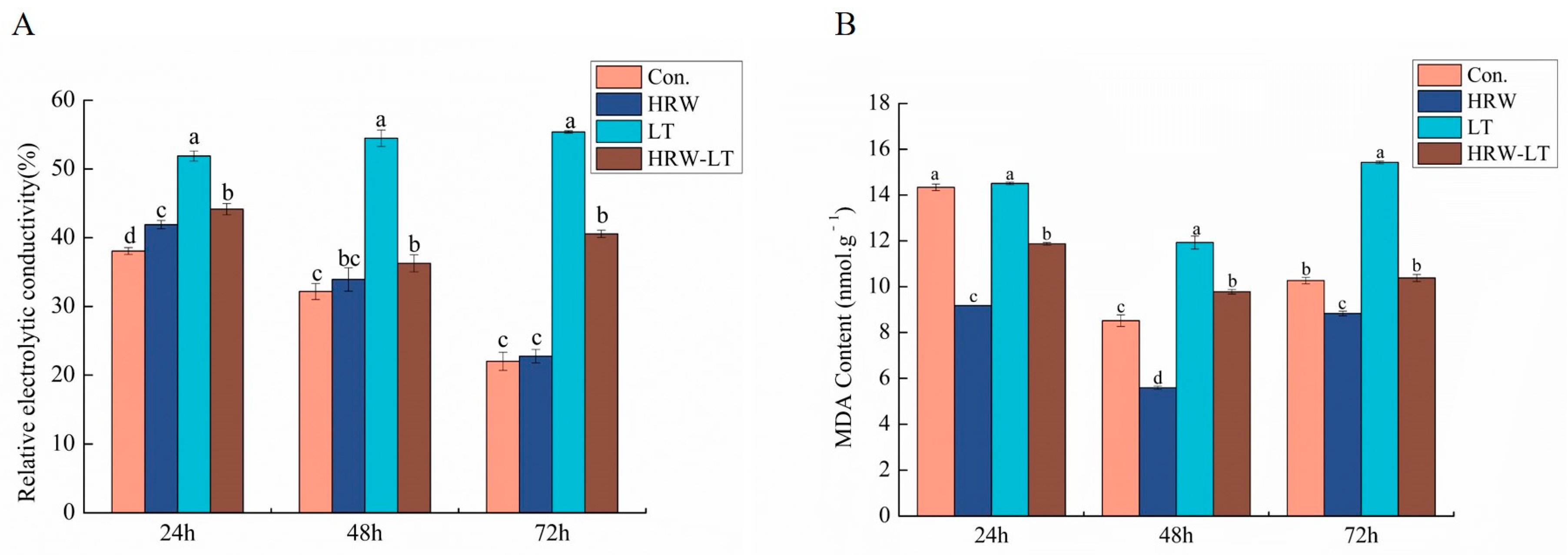
2.5. HRW Reduced Cell Membrane Damage in Cucumber Seedlings under Chilling Stress
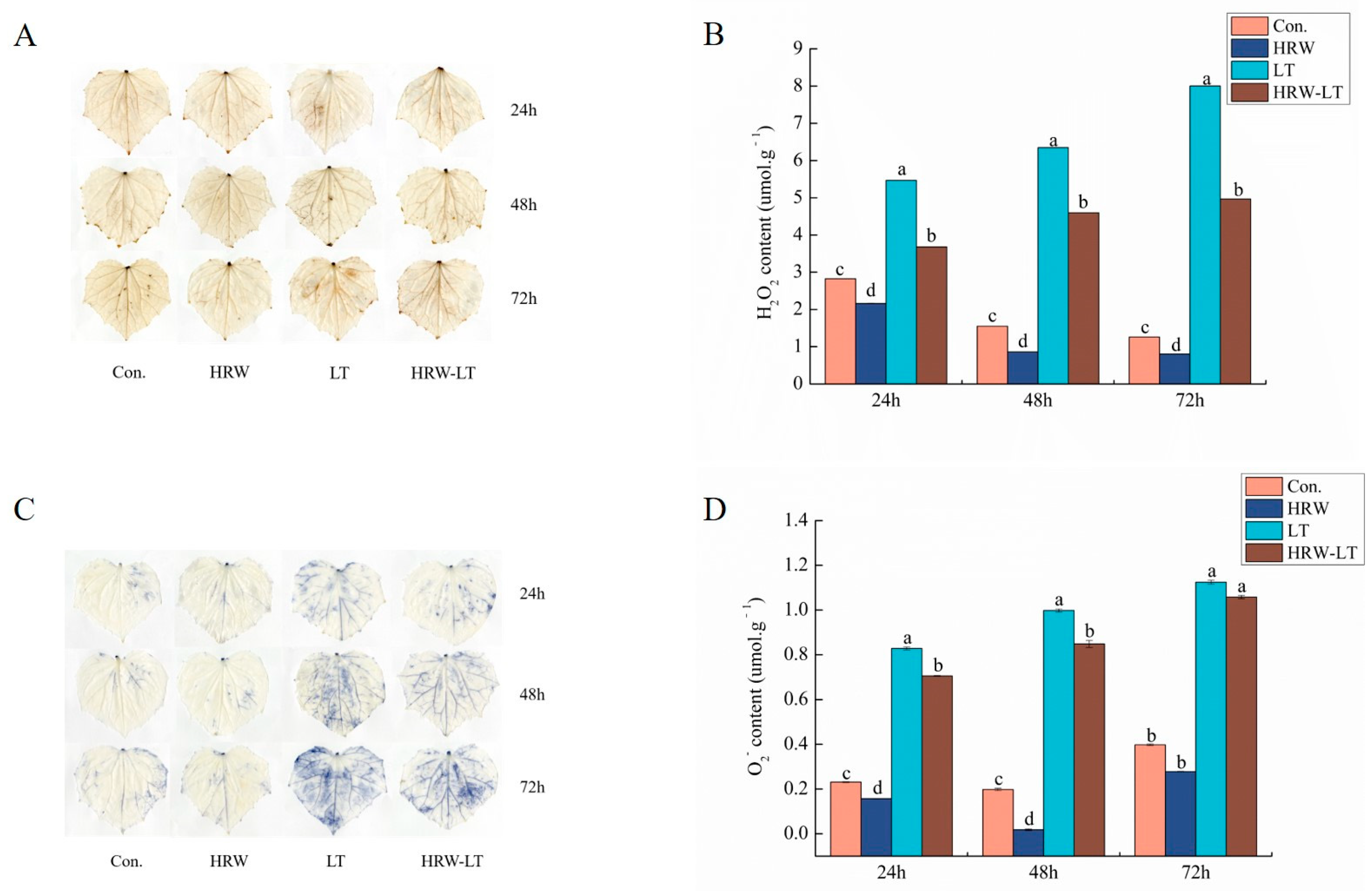
2.6. Effect of HRW on the Antioxidation System of Cucumber Seedlings under Chilling Stress
2.6.1. Effect of HRW on the Enzymatic System of Cucumber Seedlings under Chilling Stress
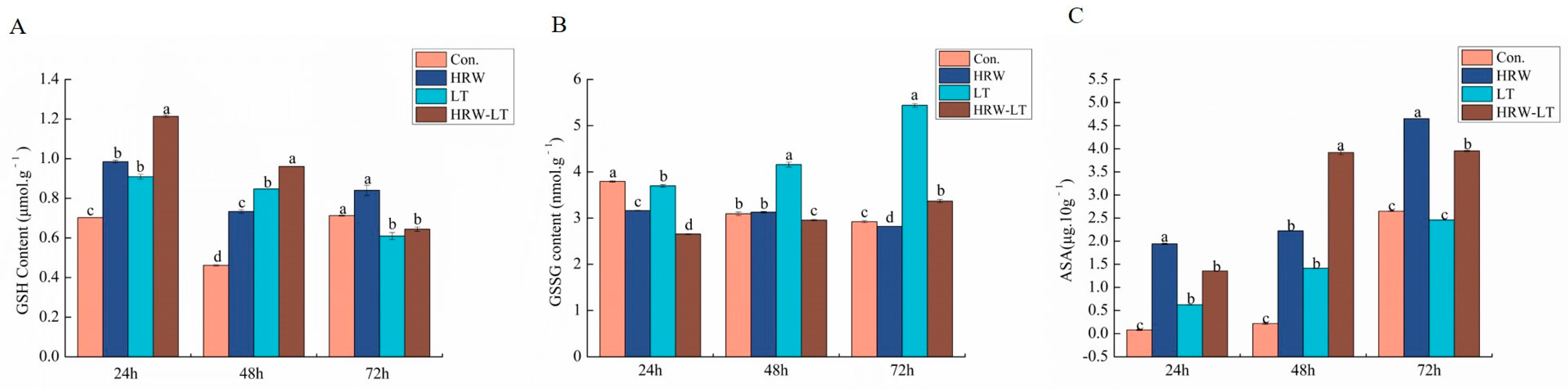
2.6.2. Effect of HRW on the Nonenzymatic System of Cucumber Seedlings under Chilling Stress
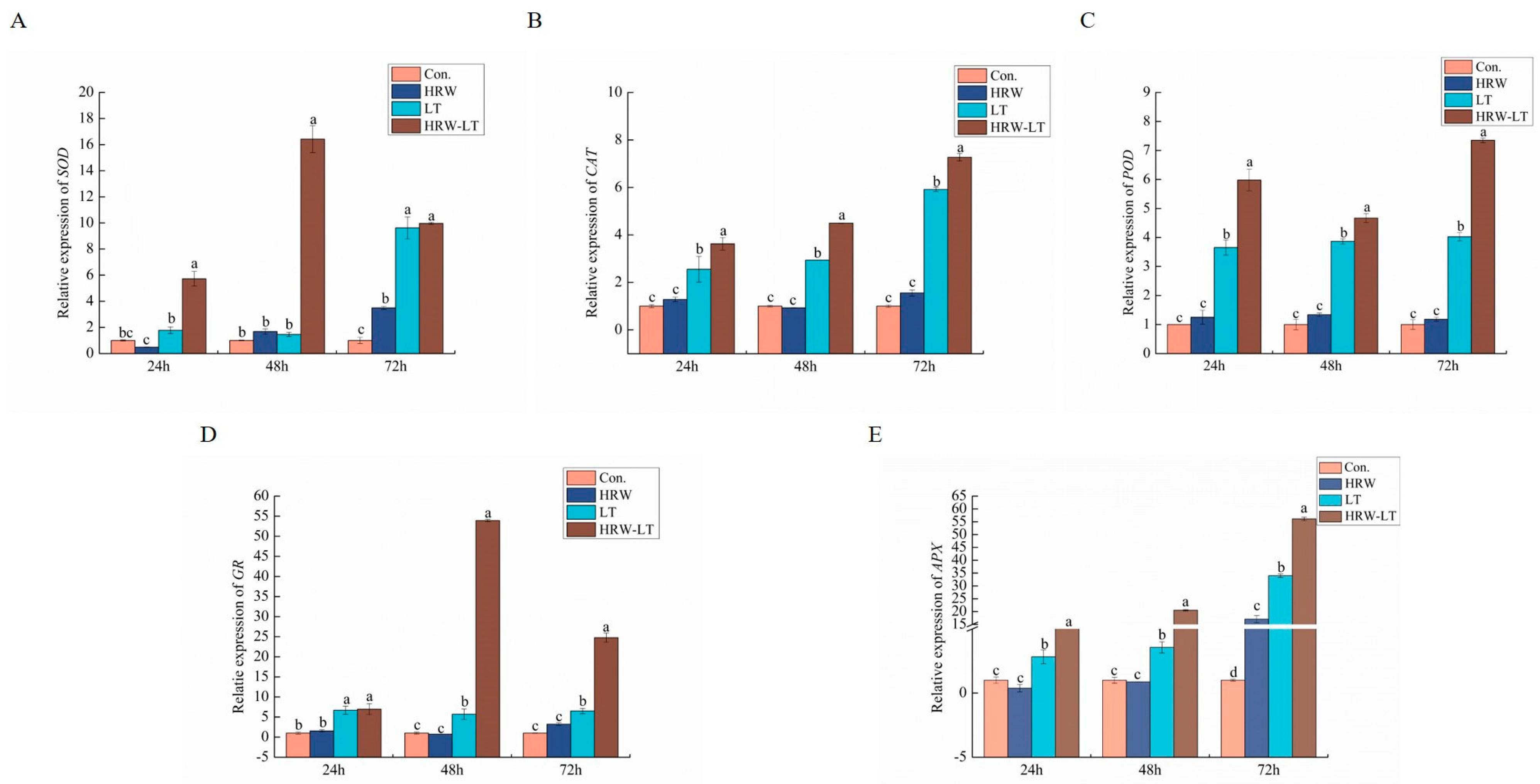
2.6.3. Effect of HRW on Antioxidant-Related Enzyme Gene Expression in Cucumber Seedlings under Chilling Stress
3. Discussion
4. Materials and Methods
4.1. Preparation of Hydrogen-Rich Water
4.2. Plant Material
4.3. Growth and Development Indicator Measurements
4.4. Measurement of Gas Exchange Parameters
4.5. Determination of Chlorophyll Content and Chlorophyll Fluorescent Parameters
4.6. MDA Content and REC Measurement
4.7. H2O2 and O2− Measurement
4.8. Antioxidant System Measurement
4.9. Primer Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sakina, A.; Wani, W.; Mushtaq, M.; Wani, S.H.; Shikari, A.B. Omics approaches for cold stress tolerance in plants. In Recent Approaches in Omics for Plant Resilience to Climate Change; Springer: Cham, Swizterland, 2019; pp. 331–356. [Google Scholar]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Fariduddin, Q.; Yusuf, M.; Chalkoo, S.; Hayat, S.; Ahmad, A. 28-homobrassinolide improves growth and photosynthesis in Cucumis sativus L. through an enhanced antioxidant system in the presence of chilling stress. Photosynthetica 2011, 49, 55–64. [Google Scholar] [CrossRef]
- Goldstein, I.; Chastre, J.; Rouby, J.-J. Novel and Innovative Strategies to Treat Ventilator-Associated Pneumonia: Optimizing the Duration of Therapy and Nebulizing Antimicrobial Agents. Semin. Respir. Crit. Care Med. 2006, 27, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Vargas, S.R.; Dos Santos, P.V.; Giraldi, L.A.; Zaiat, M.; Calijuri, M.D.C. Anaerobic phototrophic processes of hydrogen production by different strains of microalgae Chlamydomonas sp. FEMS Microbiol. Lett. 2018, 365, fny073. [Google Scholar] [CrossRef] [Green Version]
- Hemschemeier, A.; Fouchard, S.; Cournac, L.; Peltier, G.; Happe, T. Hydrogen production by Chlamydomonas reinhardtii: An elaborate interplay of electron sources and sinks. Planta 2007, 227, 397–407. [Google Scholar] [CrossRef]
- Ohta, S. Recent Progress Toward Hydrogen Medicine: Potential of Molecular Hydrogen for Preventive and Therapeutic Applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Renwick, G.M.; Giumarro, C.; Siegel, S.M. Hydrogen Metabolism in Higher Plants. Plant Physiol. 1964, 39, 303–306. [Google Scholar] [CrossRef]
- Li, J.; Zhang, B.; Chen, X.; Yu, C.; He, Y.; Cao, J.; Song, Z.; Huang, Z. Effects of hydrogen rich water on seed germination and seedling growth of vegetables. J. Chang. Veg. 2022, 8, 10–14. [Google Scholar]
- Wu, Q.; Su, N.; Huang, X.; Ling, X.; Yu, M.; Cui, J.; Shabala, S. Hydrogen-rich water promotes elongation of hypocotyls and roots in plants through mediating the level of endogenous gibberellin and auxin. Funct. Plant Biol. 2020, 47, 771–778. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, M.; Hu, L.; Liao, W.; Dawuda, M.M.; Li, C. Carbon Monoxide Is Involved in Hydrogen Gas-Induced Adventitious Root Development in Cucumber under Simulated Drought Stress. Front. Plant Sci. 2017, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Qi, N.; Wang, C.; Li, C.; Huang, D.; Li, Y.; Wang, N.; Liao, W. Hydrogen-rich water promotes the formation of bulblets in Lilium davidii var. unicolor through regulating sucrose and starch metabolism. Planta 2021, 254, 106. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef]
- Cortés, A.J.; Blair, M.W. Genotyping by Sequencing and Genome-Environment Associations in Wild Common Bean Predict Widespread Divergent Adaptation to Drought. Front. Plant Sci. 2018, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; Chavarro, C.M.; Madriñán, S.; This, D.; Blair, M.W. Molecular ecology and selection in the drought-related Asr gene polymorphisms in wild and cultivated common bean (Phaseolus vulgaris L.). BMC Genet. 2012, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Cheng, P.; Zhang, T.; Abdalmegeed, D.; Xu, S.; Shen, W. Hydrogen-rich water prepared by ammonia borane can enhance rapeseed (Brassica napus L.) seedlings tolerance against salinity, drought or cadmium. Ecotoxicol. Environ. Saf. 2021, 224, 112640. [Google Scholar] [CrossRef]
- Hu, H.; Li, P.; Shen, W. Preharvest application of hydrogen-rich water not only affects daylily bud yield but also contributes to the alleviation of bud browning. Sci. Hortic. 2021, 287, 110267. [Google Scholar] [CrossRef]
- Wu, M.; Xie, X.; Wang, Z.; Zhang, J.; Luo, Z.; Shen, W.; Yang, J. Hydrogen-rich water alleviates programmed cell death induced by GA in wheat aleurone layers by modulation of reactive oxygen species metabolism. Plant Physiol. Biochem. 2021, 163, 317–326. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.-K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, N.Y.; Kim, S.; Kang, N.Y.; Novák, O.; Ku, S.-J.; Cho, C.; Lee, D.J.; Lee, E.-J.; Strnad, M.; et al. A Subset of Cytokinin Two-component Signaling System Plays a Role in Cold Temperature Stress Response in Arabidopsis. J. Biol. Chem. 2010, 285, 23371–23386. [Google Scholar] [CrossRef] [Green Version]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146, dev151894. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Båga, M.; Chibbar, R.N. Brassinosteroid receptor mutation influences starch granule size distribution in barley grains. Plant Physiol. Biochem. 2020, 154, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Eremina, M.; Unterholzner, S.J.; Rathnayake, A.I.; Castellanos, M.; Khan, M.; Kugler, K.G.; May, S.T.; Mayer, K.F.X.; Rozhon, W.; Poppenberger, B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. Proc. Natl. Acad. Sci. USA 2016, 113, E5982–E5991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Kumar, A.; Bhardwaj, R. Plant steroidal hormone epibrassinolide regulate—Heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ. Exp. Bot. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Wilson, H.R.; Veal, D.; Whiteman, M.; Hancock, J.T. Hydrogen gas and its role in cell signalling. CAB Rev. Perspect. Agric. Veter-Sci. Nutr. Nat. Resour. 2017, 2017, 1–3. [Google Scholar] [CrossRef]
- Russell, G.; Zulfiqar, F.; Hancock, J.T. Hydrogenases and the Role of Molecular Hydrogen in Plants. Plants 2020, 9, 1136. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, M.; Sun, X. Molecular Hydrogen Is Involved in Phytohormone Signaling and Stress Responses in Plants. PLoS ONE 2013, 8, e71038. [Google Scholar] [CrossRef]
- Schumacher, I.; Menghini, D.; Ovinnikov, S.; Hauenstein, M.; Fankhauser, N.; Zipfel, C.; Hörtensteiner, S.; Aubry, S. Evolution of chlorophyll degradation is associated with plant transition to land. Plant J. 2022, 109, 1473–1488. [Google Scholar] [CrossRef]
- Ohashi, K.; Tanaka, A.; Tsuji, H. Formation of the Photosynthetic Electron Transport System during the Early Phase of Greening in Barley Leaves. Plant Physiol. 1989, 91, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Memon, S.A.; Hou, X.; Wang, L.; Li, Y. Promotive effect of 5-aminolevulinic acid on chlorophyll, antioxidative enzymes and photosynthesis of Pakchoi (Brassica campestris ssp. chinensis var. communis Tsen et Lee). Acta Physiol. Plant. 2009, 31, 51–57. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Lidon, F.C.; Loureiro, A.S.; Vieira, D.E.; Bilhó, E.A.; Nobre, P.; Costa, R. Photoinhibition in Chilling Stressed Wheat and Maize. Photosynthetica 2001, 39, 161–166. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the Relationship Between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Funct. Plant Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.; Li, J.; Yahya, M. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, X.; Wang, Z.; Shen, W.; Xu, X. Protective effects of hydrogen-rich water on the photosynthetic apparatus of maize seedlings (Zea mays L.) as a result of an increase in antioxidant enzyme activities under high light stress. Plant Growth Regul. 2015, 77, 43–56. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. Xanthophyll cycle and light stress in nature: Uniform response to excess direct sunlight among higher plant species. Planta 1996, 198, 460–470. [Google Scholar] [CrossRef]
- Ali, B.; Tao, Q.; Zhou, Y.; Gill, R.A.; Ali, S.; Rafiq, M.T.; Xu, L.; Zhou, W. 5-Aminolevolinic acid mitigates the cadmium-induced changes in Brassica napus as revealed by the biochemical and ultra-structural evaluation of roots. Ecotoxicol. Environ. Saf. 2013, 92, 271–280. [Google Scholar] [CrossRef]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Bai, L.; Miao, L.; Liu, Y.; Li, S.; Yu, X.; Li, Y. 24-Epibrassinolide Ameliorates Endogenous Hormone Levels to Enhance Low-Temperature Stress Tolerance in Cucumber Seedlings. Int. J. Mol. Sci. 2018, 19, 2497. [Google Scholar] [CrossRef] [Green Version]
- Su, N.; Wu, Q.; Liu, Y.Y.; Cai, J.; Shen, W.B.; Xia, K.; Cui, J. Hydrogen-Rich Water Reestablishes ROS Homeostasis but Exerts Differential Effects on Anthocyanin Synthesis in Two Varieties of Radish Sprouts under UV-A Irradiation. J. Agric. Food Chem. 2014, 62, 6454–6462. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.S.; Jin, Z.L.; Wan, G.L.; Liu, D.; Liu, H.B.; Yoneyama, K.; Zhou, W.J. 5-Aminolevulinic acid improves photosynthetic gas exchange capacity and ion uptake under salinity stress in oilseed rape (Brassica napus L.). Plant Soil 2010, 332, 405–415. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Kong, L.; Gui, R.; Yang, X.; Zhang, J.; Gong, Q.; Qin, D.; Zhuang, M.; Ashraf, U.; Mo, Z. Application of hydrogen-rich water modulates physio-biochemical functions and early growth of fragrant rice under Cd and Pb stress. Environ. Sci. Pollut. Res. Int. 2021, 28, 58558–58569. [Google Scholar] [CrossRef]
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef]
- Li, L.; Li, N.; Jiang, S.; Leng, J.; Wang, X. Experimental Guidance of Plant Physiology Module, 1st ed.; Science Press: Beijing, China, 2009. [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; An, Z.; Liao, J.; Ran, N.; Zhu, Y.; Ren, S.; Meng, X.; Cui, N.; Yu, Y.; Fan, H. The Role and Mechanism of Hydrogen-Rich Water in the Cucumis sativus Response to Chilling Stress. Int. J. Mol. Sci. 2023, 24, 6702. https://doi.org/10.3390/ijms24076702
Wang X, An Z, Liao J, Ran N, Zhu Y, Ren S, Meng X, Cui N, Yu Y, Fan H. The Role and Mechanism of Hydrogen-Rich Water in the Cucumis sativus Response to Chilling Stress. International Journal of Molecular Sciences. 2023; 24(7):6702. https://doi.org/10.3390/ijms24076702
Chicago/Turabian StyleWang, Xue, Zhonghui An, Jiameng Liao, Nana Ran, Yimeng Zhu, Shufeng Ren, Xiangnan Meng, Na Cui, Yang Yu, and Haiyan Fan. 2023. "The Role and Mechanism of Hydrogen-Rich Water in the Cucumis sativus Response to Chilling Stress" International Journal of Molecular Sciences 24, no. 7: 6702. https://doi.org/10.3390/ijms24076702
APA StyleWang, X., An, Z., Liao, J., Ran, N., Zhu, Y., Ren, S., Meng, X., Cui, N., Yu, Y., & Fan, H. (2023). The Role and Mechanism of Hydrogen-Rich Water in the Cucumis sativus Response to Chilling Stress. International Journal of Molecular Sciences, 24(7), 6702. https://doi.org/10.3390/ijms24076702