
RanBP1: A Potential Therapeutic Target for Cancer Stem Cells in Lung Cancer and Glioma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
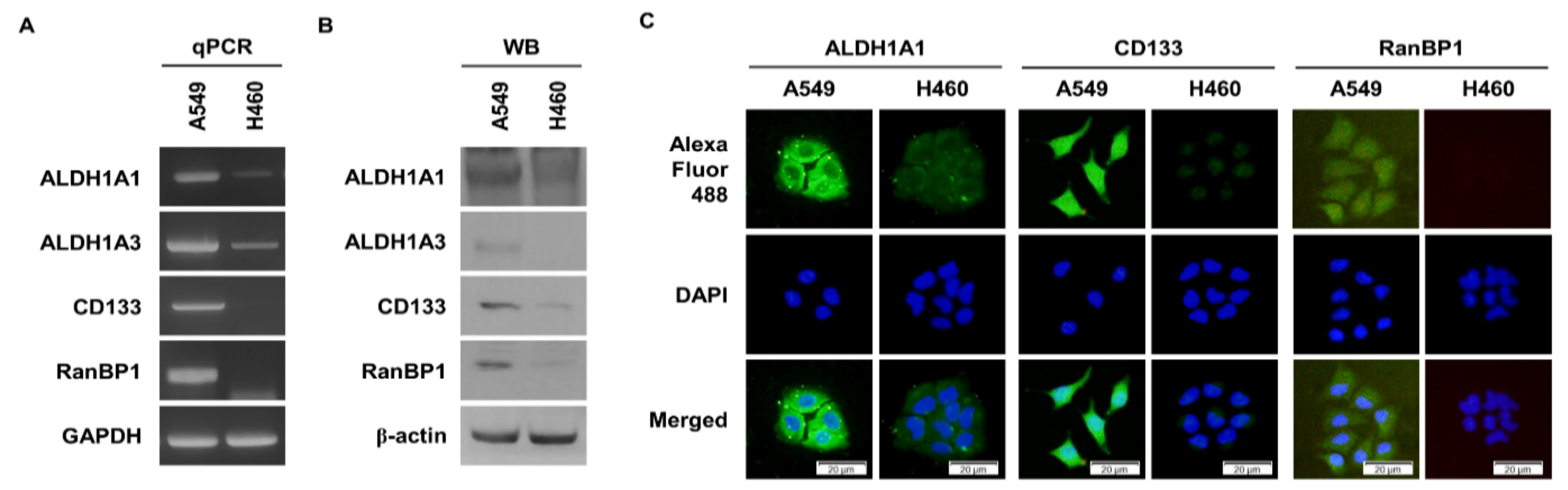
2.1. RanBP1 Expressed in Malignant Lung Cancer Cells
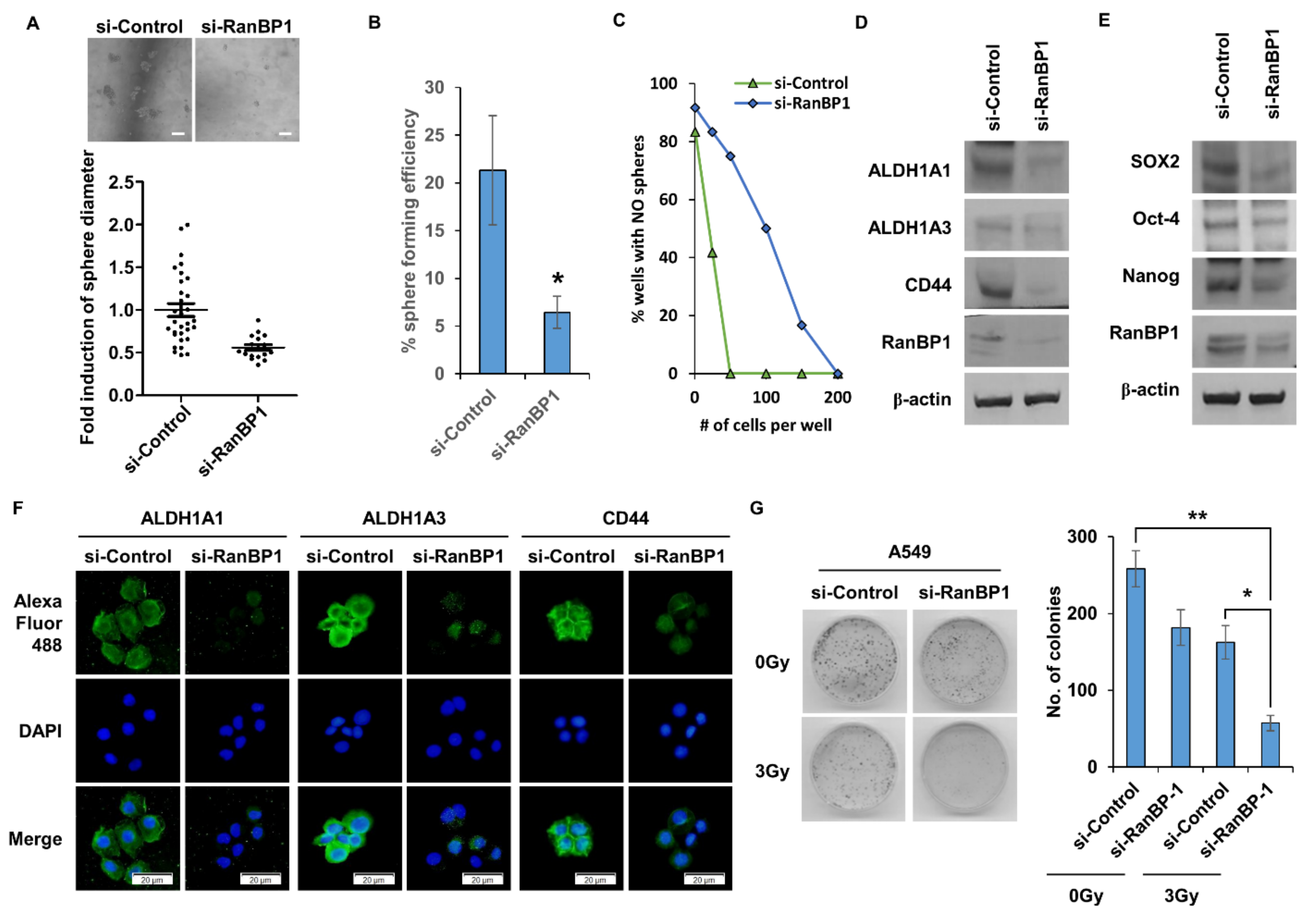
2.2. Lung CSCs Regulated by the Expression of RanBP1
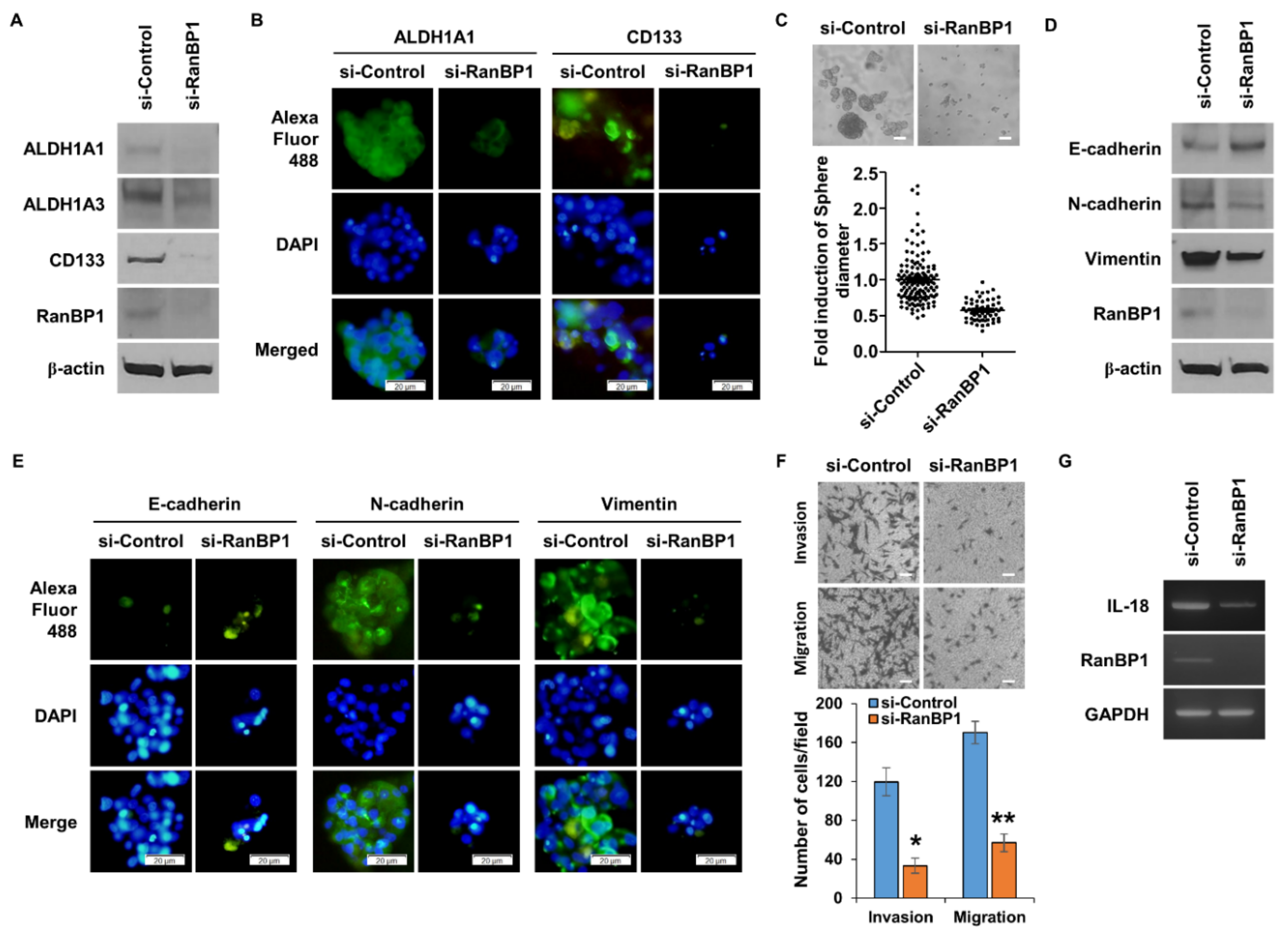
2.3. RanBP1 Involved in Regulating EMT Phenomenon
2.4. Identification of Cytokines Regulated by RanBP1 and Their Impact on LCSCs and EMT
2.5. Regulation of GSCs by RanBP1 Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Sphere-Formation Assays
4.2. Neutralization Assay
4.3. Antibodies
4.4. Small Interfering RNA (siRNA) Mediated Knockdown of RanBP1
4.5. Western Blotting
4.6. RNA Isolation
4.7. RNA Isolation Agarose Gel Electrophoresis
4.8. Immunocytochemistry
4.9. Limited Dilution Assay
4.10. Invasion and Migration Assays
4.11. Wound Healing Assay
4.12. Colony-Formation Assay and Irradiation
4.13. Cytokine Array
4.14. Kaplan−Meier Plotter
4.15. Statistical Analysis
5. Conclusions
- The RanBP1 gene is overexpressed in ALDH1+ lung cancer cells and CD133+ glioma cells.
- The overexpression of RanBP1 regulated the marker proteins and regulatory proteins of lung cancer stem cells and glioma stem cells, and modulated the ability of lung cancer cells to resist irradiation.
- RanBP1 regulated EMT marker proteins and regulatory proteins, and modulated cell migration and invasion ability.
- The cytokine IL-18 was regulated by RanBP1, and IL18 regulated cancer stem cell and EMT characteristics.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dasso, M. The Ran GTPase: Theme and variations. Curr. Biol. 2002, 12, 502–508. [Google Scholar]
- Bischoff, F.R.; Krebber, H.; Smirnova, E.; Dong, W.; Ponstingl, H. Co-activation of RanGTPase and inhibition of GTP dissociation by Ran-GTP binding protein RanBP1. EMBO J. 1995, 14, 705–715. [Google Scholar]
- Yau, K.C.; Arnaoutov, A.; Aksenova, V.; Kaufhold, R.; Chen, S.; Dasso, M. RanBP1 controls the Ran pathway in mammalian cells through regulation of mitotic RCC1 dynamics. Cell Cycle 2020, 19, 1899–1916. [Google Scholar]
- Lu, Y.; Zhao, X.; Li, K.; Luo, G.; Nie, Y.; Shi, Y.; Zhou, Y.; Ren, G.; Feng, B.; Liu, Z.; et al. Thioredoxin-like protein 2 is overexpressed in colon cancer and promotes cancer cell metastasis by interaction with ran. Antioxid. Redox Signal. 2013, 19, 899–911. [Google Scholar]
- Lu, Y.; Meng, Q.; Bai, L.; Wang, R.; Sun, Y.; Li, J.; Fan, J.; Tian, T. LINC00858 stabilizes RAN expression and promotes metastasis of gastric cancer. Biol. Direct 2022, 17, 41. [Google Scholar]
- El-Tanani, M.; Platt-Higgins, A.; Lee, Y.F.; Al Khatib, A.O.; Haggag, Y.; Sutherland, M.; Zhang, S.D.; Aljabali, A.A.A.; Mishra, V.; Serrano-Aroca, Á.; et al. Matrix metalloproteinase 2 is a target of the RAN-GTP pathway and mediates migration, invasion and metastasis in human breast cancer. Life Sci. 2022, 310, 121046. [Google Scholar]
- Che, X.; Liu, M.; Li, D.; Li, Z.; Guo, J.; Jia, R. RAN and YBX1 are required for cell proliferation and IL-4 expression and linked to poor prognosis in oral squamous cell carcinoma. Exp. Cell Res. 2021, 406, 112767. [Google Scholar]
- Chi, N.C.; Adam, E.J.; Adam, S.A. Different binding domains for Ran-GTP and Ran-GDP/RanBP1 on nuclear import factor p97. J. Biol. Chem. 1997, 272, 6818–6822. [Google Scholar]
- Lin, H.C.; Li, J.; Cheng, D.D.; Zhang, X.; Yu, T.; Zhao, F.Y.; Geng, Q.; Zhu, M.X.; Kong, H.W.; Li, H.; et al. Nuclear export protein CSE1L interacts with P65 and promotes NSCLC growth via NF-κB/MAPK pathway. Mol. Ther. -Oncolytics 2021, 21, 23–36. [Google Scholar]
- Okamura, H.; Tsutsi, H.; Komatsu, T.; Yutsudo, M.; Hakura, A.; Tanimoto, T.; Torigoe, K.; Okura, T.; Nukada, Y.; Hattori, K. Cloning of a new cytokine that induces IFN-gamma production by T cells. Nature 1995, 378, 88–91. [Google Scholar]
- Dinarello, C.A. Targeting interleukin 18 with interleukin 18 binding protein. Ann. Rheum. Dis. 2000, 59, i17–i20. [Google Scholar]
- Nolan, K.F.; Greaves, D.R.; Waldmann, H. The human interleukin 18 gene IL18 maps to 11q22.2-q22.3, closely linked to the DRD2 gene locus and distinct from mapped IDDM loci. Genomics 1998, 51, 161–163. [Google Scholar]
- Gupta, S.; Gould, M.P.; DeVecchio, J.; Canaday, D.H.; Auletta, J.J.; Heinzel, F.P. CpG-induced IFNgamma expands TLR4-specific IL-18 responses in vivo. Cell. Immunol. 2006, 243, 75–82. [Google Scholar]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The family of the interleukin-1 receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar]
- Dinarello, C.; Arend, W.; Sims, J.; Smith, D.; Blumberg, H.; O’Neill, L.; Goldbach-Mansky, R.; Pizarro, T.; Hoffman, H.; Bufler, P.; et al. IL-1 family nomenclature. Nat. Immunol. 2010, 11, 973. [Google Scholar]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar]
- Nakamura, K.; Kassem, S.; Cleynen, A.; Chrétien, M.L.; Guillerey, C.; Putz, E.M.; Bald, T.; Förster, I.; Vuckovic, S.; Hill, G.R.; et al. Dysregulated IL-18 Is a Key Driver of Immunosuppression and a Possible Therapeutic Target in the Multiple Myeloma Microenvironment. Cancer Cell 2018, 33, 634–648. [Google Scholar]
- Lopes, R.; Caetano, J.; Ferreira, B.; Barahona, F.; Carneiro, E.A.; João, C. The Immune Microenvironment in Multiple Myeloma: Friend or Foe? Cancers 2021, 13, 625. [Google Scholar]
- Choi, S.I.; Lee, J.H.; Kim, R.K.; Jung, U.; Kahm, Y.J.; Cho, E.W.; Kim, I.G. HSPA1L Enhances Cancer Stem Cell-Like Properties by Activating IGF1Rβ and Regulating β-Catenin Transcription. Int. J. Mol. Sci. 2020, 21, 6957. [Google Scholar]
- Yan, X.; Ma, L.; Yi, D.; Yoon, J.G.; Diercks, A.; Foltz, G.; Price, N.D.; Hood, L.E.; Tian, Q. A CD133-related gene expression signature identifies an aggressive glioblastoma subtype with excessive mutations. Proc. Natl. Acad. Sci. USA 2011, 25, 1591–1596. [Google Scholar]
- Lee, J.H.; Choi, S.I.; Kim, R.K.; Cho, E.W.; Kim, I.G. Tescalcin/c-Src/IGF1Rβ-mediated STAT3 activation enhances cancer stemness and radioresistant properties through ALDH1. Sci. Rep. 2018, 8, 10711. [Google Scholar]
- Yu, Z.; Pestell, T.G.; Lisanti, M.P.; Pestell, R.G. Cancer Stem Cells. Int. J. Biochem. Cell Biol. 2012, 44, 2144–2151. [Google Scholar]
- Chang, J.C. Cancer stem cells: Role in tumor growth, recurrence, metastasis, and treatment resistance. Medicine 2016, 95 (Suppl. S1), S20–S25. [Google Scholar]
- Heerboth, S.; Housman, G.; Leary, M.; Longacre, M.; Byler, S.; Lapinska, K.; Willbanks, A.; Sarkar, S. EMT and tumor metastasis. Clin. Transl. Med. 2015, 4, 6. [Google Scholar]
- Yang, J.; Antin, P.; Berx, G.; Blanpain, C.; Brabletz, T.; Bronner, M.; Campbell, K.; Cano, A.; Casanova, J.; Christofori, G.; et al. EMT International Association (TEMTIA). Guidelines and definitions for research on epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2020, 21, 341–352. [Google Scholar]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar]
- Hutchins, J.R.; Moore, W.J.; Clarke, P.R. Dynamic localisation of Ran GTPase during the cell cycle. BMC Cell Biol. 2009, 10, 66. [Google Scholar]
- Audia, S.; Brescia, C.; Dattilo, V.; D’Antona, L.; Calvano, P.; Iuliano, R.; Trapasso, F.; Perrotti, N.; Amato, R. RANBP1 (RAN Binding Protein 1): The Missing Genetic Piece in Cancer Pathophysiology and Other Complex Diseases. Cancers 2023, 15, 486. [Google Scholar]
- Barriga, E.H.; Alasaadi, D.N.; Mencarelli, C.; Mayor, R.; Pichaud, F. RanBP1 plays an essential role in directed migration of neural crest cells during development. Dev. Biol. 2022, 492, 79–86. [Google Scholar]
- Yuen, H.F.; Chan, K.K.; Platt-Higgins, A.; Dakir, E.H.; Matchett, K.B.; Haggag, Y.A.; Jithesh, P.V.; Habib, T.; Faheem, A.; Dean, F.A.; et al. Ran GTPase promotes cancer progression via Met recepto-rmediated downstream signaling. Oncotarget 2016, 7, 75854–75864. [Google Scholar]
- Zheng, D.; Cao, M.; Zuo, S.; Xia, X.; Zhi, C.; Lin, Y.; Deng, S.; Yuan, X. RANBP1 promotes colorectal cancer progression by regulating pre-miRNA nuclear export via a positive feedback loop with YAP. Oncogene 2022, 41, 930–942. [Google Scholar]
- Wu, C.; Sakorafas, P.; Miller, R.; McCarthy, D.; Scesney, S.; Dixon, R.; Ghayur, T. IL-18 receptor beta-induced changes in the presentation of IL-18 binding sites affect ligand binding and signal transduction. J. Immunol. 2003, 170, 5571–5577. [Google Scholar]
- Lasithiotaki, I.; Tsitoura, E.; Samara, K.D.; Trachalaki, A.; Charalambous, I.; Tzanakis, N.; Antoniou, K.M. NLRP3/Caspase-1 inflammasome activation is decreased in alveolar macrophages in patients with lung cancer. PLoS ONE 2018, 13, e0205242. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahm, Y.-J.; Kim, I.-G.; Kim, R.-K. RanBP1: A Potential Therapeutic Target for Cancer Stem Cells in Lung Cancer and Glioma. Int. J. Mol. Sci. 2023, 24, 6855. https://doi.org/10.3390/ijms24076855
Kahm Y-J, Kim I-G, Kim R-K. RanBP1: A Potential Therapeutic Target for Cancer Stem Cells in Lung Cancer and Glioma. International Journal of Molecular Sciences. 2023; 24(7):6855. https://doi.org/10.3390/ijms24076855
Chicago/Turabian StyleKahm, Yeon-Jee, In-Gyu Kim, and Rae-Kwon Kim. 2023. "RanBP1: A Potential Therapeutic Target for Cancer Stem Cells in Lung Cancer and Glioma" International Journal of Molecular Sciences 24, no. 7: 6855. https://doi.org/10.3390/ijms24076855
APA StyleKahm, Y.-J., Kim, I.-G., & Kim, R.-K. (2023). RanBP1: A Potential Therapeutic Target for Cancer Stem Cells in Lung Cancer and Glioma. International Journal of Molecular Sciences, 24(7), 6855. https://doi.org/10.3390/ijms24076855