
RNA-Dependent RNA Targeting by CRISPR-Cas Systems: Characterizations and Applications

Abstract
:1. Introduction
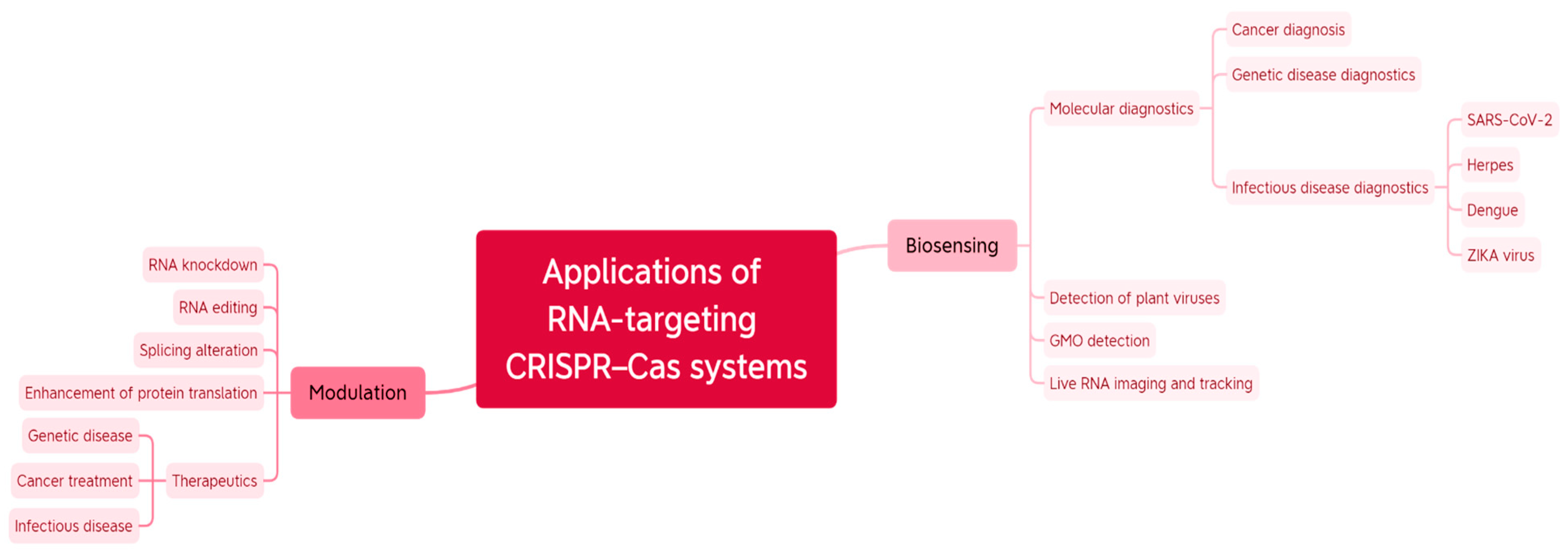
2. Applications of the Versatile CRISPR-Cas RNA Targeting Systems
2.1. Cas-Mediated Biosensing
2.1.1. Bactericidal Agents
2.1.2. CRISPR-Mediated Molecular Diagnostics
2.1.3. Detection of Plant Viruses
2.1.4. CRISPR-Cas Systems in Detection of GMOs
2.2. RNA Imaging and Tracking in Living Cells
2.3. Cancer Diagnosis and Therapy
2.4. RNA-Editing and Regulation of Gene Expression
2.4.1. RNA Knockdown and Gene Silencing
2.4.2. Splicing Alteration
2.4.3. Enhancement of Protein Translation
2.4.4. RNA-Targeting CRISPR-Cas Systems for RNA Editing
3. CRISPR Toolbox
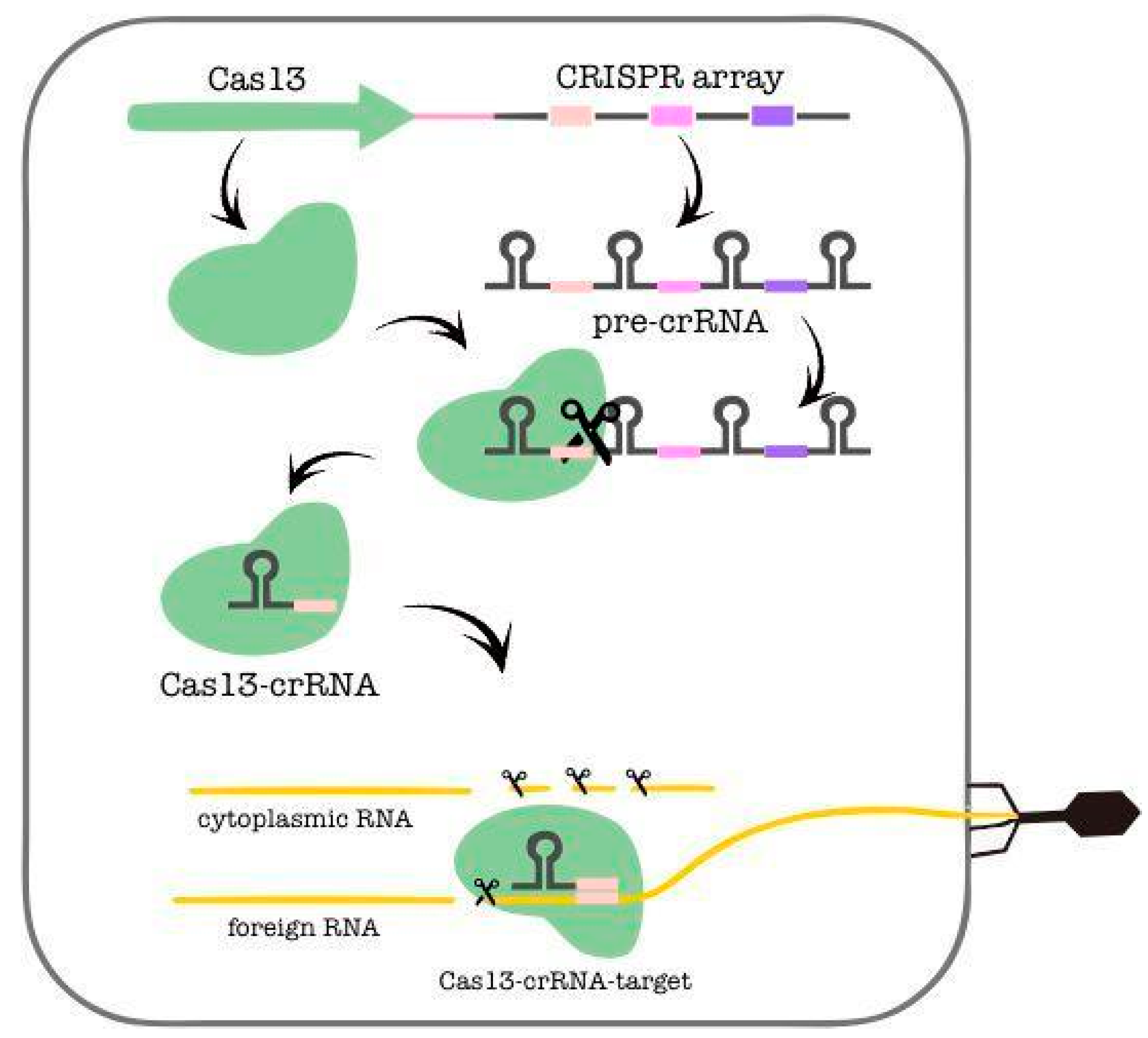
3.1. Type VI CRISPR-Cas
3.2. Type V CRISPR-Cas
3.3. Type III CRISPR-Cas
3.4. Type II CRISPR-Cas

{kind=link}
{kind=link}
| Effector | Characterizations |
|---|---|
| Cas13 | Two HEPN nuclease domains |
| Collateral RNA cleavage No PAM requirements Suffer from severe cytotoxic effects due to trans-cleavage activity Processes own crRNA Single-subunit effector Small enough to fit a single AAV vector with a crRNA expression cassette Spacer length ~ 25 nt Adapted to serve numerous applications Extensively characterized | |
| Cas12g | Collateral RNase and ssDNase activities |
| Single-subunit effector No PAM requirements Compact size Applies two guide RNAs, a tracrRNA and a crRNA Insufficiently explored | |
| Csm | Multiprotein complex No collateral activity |
| No PAM requirements | |
| Powerful tool for eukaryotic RNA knockdown Fewer off-targets than Cas13 RNase and DNase activity crRNA lies at the core of the complex Spacers crRNAs range from ~30 to 45 nt Extensively characterized | |
| Cas9 | Contains two nuclease domains (HNH and RuvC) Collateral activity not identified PAM is required |
| Single-subunit effector Spacer length is 20 nt Low off-target product Target dsDNA and RNA Relies on RNase III to process its crRNA Extensively characterized |
4. Conclusions
Funding
Conflicts of Interest
References
- Redding, S.; Stersnberg, S.H.; Marshall, M.; Gibb, B.; Bhat, P.; Guegler, C.K.; Wiedenheft, B.; Doudna, J.A.; Greene, E.C. Surveillance and Processing of Foreign DNA by the Escherichia Coli CRISPR-Cas System. Cell 2015, 163, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Pausch, P.; Doudna, J.A. Structural Biology of CRISPR–Cas Immunity and Genome Editing Enzymes. Nat. Rev. Microbiol. 2022, 20, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.A.; Utkina, I.; Wolf, Y.I.; Makarova, K.S.; Severinov, K.V.; Koonin, E.V. CRISPR Arrays Away from Cas Genes. CRISPR J. 2020, 3, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Khanzadi, M.N.; Khan, A.A. CRISPR/Cas9: Nature’s Gift to Prokaryotes and an Auspicious Tool in Genome Editing. J. Basic Microbiol. 2020, 60, 91–102. [Google Scholar] [CrossRef]
- Wright, A.V.; Nuñez, J.K.; Doudna, J.A. Biology and Applications of CRISPR Systems: Harnessing Nature’s Toolbox for Genome Engineering. Cell 2016, 164, 29–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenti, M.T.; Serena, M.; Carbonare, L.D.; Zipeto, D. CRISPR/Cas System: An Emerging Technology in Stem Cell Research. World J. Stem Cells 2019, 11, 937–956. [Google Scholar] [CrossRef]
- Gupta, R.; Ghosh, A.; Chakravarti, R.; Singh, R.; Ravichandiran, V.; Swarnakar, S.; Ghosh, D. Cas13d: A New Molecular Scissor for Transcriptome Engineering. Front. Cell Dev. Biol. 2022, 10, 866800. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Jillette, N.; Zhu, J.J.; Li, S.; Cheng, A.W. CRISPR Artificial Splicing Factors. Nat. Commun. 2020, 11, 2973. [Google Scholar] [CrossRef]
- Tang, X.-Z.E.; Tan, S.X.; Hoon, S.; Yeo, G.W. Pre-Existing Adaptive Immunity to the RNA-Editing Enzyme Cas13d in Humans. Nat. Med. 2022, 28, 1372–1376. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Xiao, R.; Han, R.; Chang, L. Cryo-EM Structure of the RNA-Guided Ribonuclease Cas12g. Nat. Chem. Biol. 2021, 17, 387–393. [Google Scholar] [CrossRef]
- Terns, M.P. CRISPR-Based Technologies: Impact of RNA-Targeting Systems. Mol. Cell 2018, 72, 404–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrangou, R.; Doudna, J.A. Applications of CRISPR Technologies in Research and Beyond. Nat. Biotechnol. 2016, 34, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Dugar, G.; Leenay, R.T.; Eisenbart, S.K.; Bischler, T.; Aul, B.U.; Beisel, C.L.; Sharma, C.M. CRISPR RNA-Dependent Binding and Cleavage of Endogenous RNAs by the Campylobacter Jejuni Cas9. Mol. Cell 2018, 69, 893–905.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazlauskiene, M.; Tamulaitis, G.; Kostiuk, G.; Venclovas, Č.; Siksnys, V. Spatiotemporal Control of Type III-A CRISPR-Cas Immunity: Coupling DNA Degradation with the Target RNA Recognition. Mol. Cell 2016, 62, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osawa, T.; Inanaga, H.; Sato, C.; Numata, T. Crystal Structure of the CRISPR-Cas RNA Silencing Cmr Complex Bound to a Target Analog. Mol. Cell 2015, 58, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.X.; Hunnewell, P.; Alfonse, L.E.; Carte, J.M.; Keston-Smith, E.; Sothiselvam, S.; Garrity, A.J.; Chong, S.; Makarova, K.S.; Koonin, E.V.; et al. Functionally Diverse Type V CRISPR-Cas Systems. Science 2019, 363, 88–91. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.R. Molecular Mechanisms of RNA Targeting by Cas13-Containing Type VI CRISPR–Cas Systems. J. Mol. Biol. 2019, 431, 66–87. [Google Scholar] [CrossRef]
- Strutt, S.C.; Torrez, R.M.; Kaya, E.; Negrete, O.A.; Doudna, J.A. RNA-Dependent RNA Targeting by CRISPR-Cas9. eLife 2018, 7, e32724. [Google Scholar] [CrossRef]
- Rousseau, B.A.; Hou, Z.; Gramelspacher, M.J.; Zhang, Y. Programmable RNA Cleavage and Recognition by a Natural CRISPR-Cas9 System from Neisseria Meningitidis. Mol. Cell 2018, 69, 906–914.e4. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.R.; Oakes, B.L.; Sternberg, S.H.; East-Seletsky, A.; Kaplan, M.; Doudna, J.A. Programmable RNA Recognition and Cleavage by CRISPR/Cas9. Nature 2014, 516, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Bauer, D.E.; Chiarle, R. Assessing and Advancing the Safety of CRISPR-Cas Tools: From DNA to RNA Editing. Nat. Commun. 2023, 14, 212. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yin, K.; Zhang, Q.; Gao, C.; Qiu, J.-L. Modulating Chromatin Accessibility by Transactivation and Targeting Proximal DsgRNAs Enhances Cas9 Editing Efficiency in Vivo. Genome Biol. 2019, 20, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Beljouw, S.P.B.; Sanders, J.; Rodríguez-Molina, A.; Brouns, S.J.J. RNA-Targeting CRISPR–Cas Systems. Nat. Rev. Microbiol. 2023, 21, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 Is a Single-Component Programmable RNA-Guided RNA-Targeting CRISPR Effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two Distinct RNase Activities of CRISPR-C2c2 Enable Guide-RNA Processing and RNA Detection. Nature 2016, 538, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Bikard, D.; Euler, C.W.; Jiang, W.; Nussenzweig, P.M.; Goldberg, G.W.; Duportet, X.; Fischetti, V.A.; Marraffini, L.A. Exploiting CRISPR-Cas Nucleases to Produce Sequence-Specific Antimicrobials. Nat. Biotechnol. 2014, 32, 1146–1150. [Google Scholar] [CrossRef] [Green Version]
- Collias, D.; Beisel, C.L. CRISPR Technologies and the Search for the PAM-Free Nuclease. Nat. Commun. 2021, 12, 555. [Google Scholar] [CrossRef]
- Kiga, K.; Tan, X.-E.; Ibarra-Chávez, R.; Watanabe, S.; Aiba, Y.; Sato’o, Y.; Li, F.-Y.; Sasahara, T.; Cui, B.; Kawauchi, M.; et al. Development of CRISPR-Cas13a-Based Antimicrobials Capable of Sequence-Specific Killing of Target Bacteria. Nat. Commun. 2020, 11, 2934. [Google Scholar] [CrossRef]
- Kordyś, M.; Sen, R.; Warkocki, Z. Applications of the Versatile CRISPR-Cas13 RNA Targeting System. WIREs RNA 2022, 13, e1694. [Google Scholar] [CrossRef]
- Song, Z.; Yu, Y.; Bai, X.; Jia, Y.; Tian, J.; Gu, K.; Zhao, M.; Zhou, C.; Zhang, X.; Wang, H.; et al. Pathogen-Specific Bactericidal Method Mediated by Conjugative Delivery of CRISPR-Cas13a Targeting Bacterial Endogenous Transcripts. Microbiol. Spectr. 2022, 10, e01300-22. [Google Scholar] [CrossRef]
- Pardee, K.; Green, A.A.; Takahashi, M.K.; Braff, D.; Lambert, G.; Lee, J.W.; Ferrante, T.; Ma, D.; Donghia, N.; Fan, M.; et al. Rapid, Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell 2016, 165, 1255–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.X.; Cui, J.Q.; Wu, Z.; Yao, S. Recent Progress in Nucleic Acid Detection with CRISPR. Lab. Chip 2023, 23, 1467–1492. [Google Scholar] [CrossRef]
- Bayoumi, M.; Munir, M. Potential Use of CRISPR/Cas13 Machinery in Understanding Virus–Host Interaction. Front. Microbiol. 2021, 12, 743580. [Google Scholar] [CrossRef] [PubMed]
- Hang, X.-M.; Liu, P.-F.; Tian, S.; Wang, H.-Y.; Zhao, K.-R.; Wang, L. Rapid and Sensitive Detection of Ebola RNA in an Unamplified Sample Based on CRISPR-Cas13a and DNA Roller Machine. Biosens. Bioelectron. 2022, 211, 114393. [Google Scholar] [CrossRef] [PubMed]
- Myhrvold, C.; Freije, C.A.; Gootenberg, J.S.; Abudayyeh, O.O.; Metsky, H.C.; Durbin, A.F.; Kellner, M.J.; Tan, A.L.; Paul, L.M.; Parham, L.A.; et al. Field-Deployable Viral Diagnostics Using CRISPR-Cas13. Science 2018, 360, 444–448. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, X.; Wu, Y.; Guo, M.; Gu, C.; Dai, C.; Kong, D.; Wang, Y.; Zhang, C.; Qu, D.; et al. Rapid and Ultrasensitive Electromechanical Detection of Ions, Biomolecules and SARS-CoV-2 RNA in Unamplified Samples. Nat. Biomed. Eng. 2022, 6, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, T.; Wang, M.; Ledesma-Amaro, R.; Lu, Y.; Hu, X.; Zhang, T.; Yang, M.; Li, Y.; Xiang, J.; et al. CRISPR-Cas13a Cascade-Based Viral RNA Assay for Detecting SARS-CoV-2 and Its Mutations in Clinical Samples. Sens. Actuators B Chem. 2022, 362, 131765. [Google Scholar] [CrossRef]
- Mahas, A.; Aman, R.; Mahfouz, M. CRISPR-Cas13d Mediates Robust RNA Virus Interference in Plants. Genome Biol. 2019, 20, 263. [Google Scholar] [CrossRef] [Green Version]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA Virus Interference via CRISPR/Cas13a System in Plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Ghouri, M.Z.; Munawar, N.; Aftab, S.O.; Ahmad, A. Regulation of CRISPR Edited Food and Feed: Legislation and Future. In GMOs and Political Stance; Elsevier: Amsterdam, The Netherlands, 2023; pp. 261–287. ISBN 978-0-12-823903-2. [Google Scholar]
- Wang, M.; Wang, H.; Li, K.; Li, X.; Wang, X.; Wang, Z. Review of CRISPR/Cas Systems on Detection of Nucleotide Sequences. Foods 2023, 12, 477. [Google Scholar] [CrossRef]
- Yuan, C.; Tian, T.; Sun, J.; Hu, M.; Wang, X.; Xiong, E.; Cheng, M.; Bao, Y.; Lin, W.; Jiang, J.; et al. Universal and Naked-Eye Gene Detection Platform Based on the Clustered Regularly Interspaced Short Palindromic Repeats/Cas12a/13a System. Anal. Chem. 2020, 92, 4029–4037. [Google Scholar] [CrossRef]
- Cao, G.; Dong, J.; Chen, X.; Lu, P.; Xiong, Y.; Peng, L.; Li, J.; Huo, D.; Hou, C. Simultaneous Detection of CaMV35S and T-Nos Utilizing CRISPR/Cas12a and Cas13a with Multiplex-PCR (MPT-Cas12a/13a). Chem. Commun. 2022, 58, 6328–6331. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA Targeting with CRISPR–Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Zhao, B.S.; Myers, S.A.; Carr, S.A.; He, C.; Ting, A.Y. RNA–Protein Interaction Mapping via MS2- or Cas13-Based APEX Targeting. Proc. Natl. Acad. Sci. USA 2020, 117, 22068–22079. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, W.; Shi, T.; Lu, P.; Zhuang, M.; Liu, J.-L. Capturing RNA–Protein Interaction via CRUIS. Nucleic Acids Res. 2020, 48, e52. [Google Scholar] [CrossRef]
- Yi, W.; Li, J.; Zhu, X.; Wang, X.; Fan, L.; Sun, W.; Liao, L.; Zhang, J.; Li, X.; Ye, J.; et al. CRISPR-Assisted Detection of RNA–Protein Interactions in Living Cells. Nat. Methods 2020, 17, 685–688. [Google Scholar] [CrossRef]
- Jiang, W.; Li, H.; Liu, X.; Zhang, J.; zhang, W.; Li, T.; Liu, L.; Yu, X. Precise and Efficient Silencing of Mutant Kras G12D by CRISPR-CasRx Controls Pancreatic Cancer Progression. Theranostics 2020, 10, 11507–11519. [Google Scholar] [CrossRef]
- Cao, C.; Li, A.; Xu, C.; Wu, B.; Liu, J.; Liu, Y. Enhancement of Protein Translation by CRISPR/DCasRx Coupled with SINEB2 Repeat of Noncoding RNAs. Nucleic Acids Res. 2023, gkad010. [Google Scholar] [CrossRef]
- Ruchika; Nakamura, T. Understanding RNA Editing and Its Use in Gene Editing. Gene Genome Ed. 2022, 3, 100021. [Google Scholar] [CrossRef]
- Corey, D.R. Nusinersen, an Antisense Oligonucleotide Drug for Spinal Muscular Atrophy. Nat. Neurosci. 2017, 20, 497–499. [Google Scholar] [CrossRef]
- Smith, I.; Greenside, P.G.; Natoli, T.; Lahr, D.L.; Wadden, D.; Tirosh, I.; Narayan, R.; Root, D.E.; Golub, T.R.; Subramanian, A.; et al. Evaluation of RNAi and CRISPR Technologies by Large-Scale Gene Expression Profiling in the Connectivity Map. PLoS Biol. 2017, 15, e2003213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.E.; Lim, C.K.W.; Krishnan, R.; McCallister, T.X.; Saporito-Magriña, C.; Zeballos, M.A.; McPheron, G.D.; Gaj, T. Targeted Gene Silencing in the Nervous System with CRISPR-Cas13. Sci. Adv. 2022, 8, eabk2485. [Google Scholar] [CrossRef] [PubMed]
- Fricke, T.; Smalakyte, D.; Lapinski, M.; Pateria, A.; Weige, C.; Pastor, M.; Kolano, A.; Winata, C.; Siksnys, V.; Tamulaitis, G.; et al. Targeted RNA Knockdown by a Type III CRISPR-Cas Complex in Zebrafish. CRISPR J. 2020, 3, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Mancilla, A.; Wessels, H.-H.; Legut, M.; Kadina, A.; Mabuchi, M.; Walker, J.; Robb, G.B.; Holden, K.; Sanjana, N.E. Chemically Modified Guide RNAs Enhance CRISPR-Cas13 Knockdown in Human Cells. Cell Chem. Biol. 2022, 29, 321–327.e4. [Google Scholar] [CrossRef]
- Montagud-Martínez, R.; Márquez-Costa, R.; Rodrigo, G. Programmable Regulation of Translation by Harnessing the CRISPR-Cas13 System. Chem. Commun. 2023. [Google Scholar] [CrossRef]
- Xiao, Q.; Xu, Z.; Xue, Y.; Xu, C.; Han, L.; Liu, Y.; Wang, F.; Zhang, R.; Han, S.; Wang, X.; et al. Rescue of Autosomal Dominant Hearing Loss by in Vivo Delivery of Mini DCas13X-Derived RNA Base Editor. Sci. Transl. Med. 2022, 14, eabn0449. [Google Scholar] [CrossRef]
- Tong, H.; Huang, J.; Xiao, Q.; He, B.; Dong, X.; Liu, Y.; Yang, X.; Han, D.; Wang, Z.; Wang, X.; et al. High-Fidelity Cas13 Variants for Targeted RNA Degradation with Minimal Collateral Effects. Nat. Biotechnol. 2023, 41, 108–119. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Qiu, Q.; Zhou, Q.; Ding, J.; Lu, Y.; Liu, P. Alternative Polyadenylation: Methods, Mechanism, Function, and Role in Cancer. J. Exp. Clin. Cancer Res. 2021, 40, 51. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, Y.; Xu, J.; Wang, X.; Luo, S.; Mao, B.; Zhou, Q.; Li, W. Metagenomic Discovery of Novel CRISPR-Cas13 Systems. Cell Discov. 2022, 8, 107. [Google Scholar] [CrossRef]
- Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Dong, X.; Bai, W.; Wang, Y.; et al. Programmable RNA Editing with Compact CRISPR–Cas13 Systems from Uncultivated Microbes. Nat. Methods 2021, 18, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Altae-Tran, H.; Jin, X.; Madigan, V.J.; Oshiro, R.; Makarova, K.S.; Koonin, E.V.; Zhang, F. Compact RNA Editors with Small Cas13 Proteins. Nat. Biotechnol. 2022, 40, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, Z. Recent Advances in Structural Studies of the CRISPR-Cas-Mediated Genome Editing Tools. Natl. Sci. Rev. 2019, 6, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Pei, D.-S. Insights Gained from RNA Editing Targeted by the CRISPR-Cas13 Family. Int. J. Mol. Sci. 2022, 23, 11400. [Google Scholar] [CrossRef] [PubMed]
- Krohannon, A.; Srivastava, M.; Rauch, S.; Srivastava, R.; Dickinson, B.C.; Janga, S.C. CASowary: CRISPR-Cas13 Guide RNA Predictor for Transcript Depletion. BMC Genomics 2022, 23, 172. [Google Scholar] [CrossRef]
- Huynh, N.; Depner, N.; Larson, R.; King-Jones, K. A Versatile Toolkit for CRISPR-Cas13-Based RNA Manipulation in Drosophila. Genome Biol. 2020, 21, 279. [Google Scholar] [CrossRef]
- Ai, Y.; Liang, D.; Wilusz, J.E. CRISPR/Cas13 Effectors Have Differing Extents of off-Target Effects That Limit Their Utility in Eukaryotic Cells. Nucleic Acids Res. 2022, 50, e65. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a Target Binding Unleashes Indiscriminate Single-Stranded DNase Activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Y.; Cheng, Q.-X.; Liu, J.-K.; Nie, X.-Q.; Zhao, G.-P.; Wang, J. CRISPR-Cas12a Has Both Cis- and Trans-Cleavage Activities on Single-Stranded DNA. Cell Res. 2018, 28, 491–493. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.; Collins, J.J.; Zhang, F. Multiplexed and Portable Nucleic Acid Detection Platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Coleman, G.A.; Davín, A.A.; Mahendrarajah, T.A.; Szánthó, L.L.; Spang, A.; Hugenholtz, P.; Szöllősi, G.J.; Williams, T.A. A Rooted Phylogeny Resolves Early Bacterial Evolution. Science 2021, 372, eabe0511. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; Sofos, N.; Montoya, G. Structural Basis of CRISPR-Cas Type III Prokaryotic Defence Systems. Curr. Opin. Struct. Biol. 2020, 65, 119–129. [Google Scholar] [CrossRef]
- Hale, C.R.; Zhao, P.; Olson, S.; Duff, M.O.; Graveley, B.R.; Wells, L.; Terns, R.M.; Terns, M.P. RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex. Cell 2009, 139, 945–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colognori, D.; Trinidad, M.; Doudna, J.A. Precise Transcript Targeting by CRISPR-Csm Complexes. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Kazlauskiene, M.; Kostiuk, G.; Venclovas, Č.; Tamulaitis, G.; Siksnys, V. A Cyclic Oligonucleotide Signaling Pathway in Type III CRISPR-Cas Systems. Science 2017, 357, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Niewoehner, O.; Garcia-Doval, C.; Rostøl, J.T.; Berk, C.; Schwede, F.; Bigler, L.; Hall, J.; Marraffini, L.A.; Jinek, M. Type III CRISPR–Cas Systems Produce Cyclic Oligoadenylate Second Messengers. Nature 2017, 548, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Klompe, S.E.; Vlot, M.; van der Oost, J.; Staals, R.H.J. Shooting the Messenger: RNA-Targetting CRISPR-Cas Systems. Biosci. Rep. 2018, 38, BSR20170788. [Google Scholar] [CrossRef] [Green Version]
- Mogila, I.; Kazlauskiene, M.; Valinskyte, S.; Tamulaitiene, G.; Tamulaitis, G.; Siksnys, V. Genetic Dissection of the Type III-A CRISPR-Cas System Csm Complex Reveals Roles of Individual Subunits. Cell Rep. 2019, 26, 2753–2765.e4. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Ma, J.; Wang, J.; Artamonova, D.; Wang, M.; Liu, L.; Xiang, H.; Severinov, K.; Zhang, X.; Wang, Y. Structure Studies of the CRISPR-Csm Complex Reveal Mechanism of Co-Transcriptional Interference. Cell 2019, 176, 239–253.e16. [Google Scholar] [CrossRef] [Green Version]
- Samai, P.; Pyenson, N.; Jiang, W.; Goldberg, G.W.; Hatoum-Aslan, A.; Marraffini, L.A. Co-Transcriptional DNA and RNA Cleavage during Type III CRISPR-Cas Immunity. Cell 2015, 161, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Tamulaitis, G.; Kazlauskiene, M.; Manakova, E.; Venclovas, Č.; Nwokeoji, A.O.; Dickman, M.J.; Horvath, P.; Siksnys, V. Programmable RNA Shredding by the Type III-A CRISPR-Cas System of Streptococcus Thermophilus. Mol. Cell 2014, 56, 506–517. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, H.T.; Cooper, J.C.; Lo, L.; Potter, J.; Terns, R.M.; Terns, M.P. Programmable Type III-A CRISPR-Cas DNA Targeting Modules. PLoS ONE 2017, 12, e0176221. [Google Scholar] [CrossRef] [Green Version]
- Cameron, P.; Coons, M.M.; Klompe, S.E.; Lied, A.M.; Smith, S.C.; Vidal, B.; Donohoue, P.D.; Rotstein, T.; Kohrs, B.W.; Nyer, D.B.; et al. Harnessing Type I CRISPR–Cas Systems for Genome Engineering in Human Cells. Nat. Biotechnol. 2019, 37, 1471–1477. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, J.; Zhi, S.; Zheng, Q.; Ma, W.; Huang, J.; Liu, Y.; Liu, D.; Liang, P.; Songyang, Z. Repurposing Type I–F CRISPR–Cas System as a Transcriptional Activation Tool in Human Cells. Nat. Commun. 2020, 11, 3136. [Google Scholar] [CrossRef] [PubMed]
- Burmistrz, M.; Krakowski, K.; Krawczyk-Balska, A. RNA-Targeting CRISPR–Cas Systems and Their Applications. Int. J. Mol. Sci. 2020, 21, 1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunitseva, N.; Evteeva, M.; Borisova, A.; Patrushev, M.; Subach, F. RNA-Dependent RNA Targeting by CRISPR-Cas Systems: Characterizations and Applications. Int. J. Mol. Sci. 2023, 24, 6894. https://doi.org/10.3390/ijms24086894
Gunitseva N, Evteeva M, Borisova A, Patrushev M, Subach F. RNA-Dependent RNA Targeting by CRISPR-Cas Systems: Characterizations and Applications. International Journal of Molecular Sciences. 2023; 24(8):6894. https://doi.org/10.3390/ijms24086894
Chicago/Turabian StyleGunitseva, Natalia, Marta Evteeva, Anna Borisova, Maxim Patrushev, and Fedor Subach. 2023. "RNA-Dependent RNA Targeting by CRISPR-Cas Systems: Characterizations and Applications" International Journal of Molecular Sciences 24, no. 8: 6894. https://doi.org/10.3390/ijms24086894
APA StyleGunitseva, N., Evteeva, M., Borisova, A., Patrushev, M., & Subach, F. (2023). RNA-Dependent RNA Targeting by CRISPR-Cas Systems: Characterizations and Applications. International Journal of Molecular Sciences, 24(8), 6894. https://doi.org/10.3390/ijms24086894