
Differential Cytokine Responses and the Clinical Severity of Adult and Pediatric Nephropathia Epidemica
 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
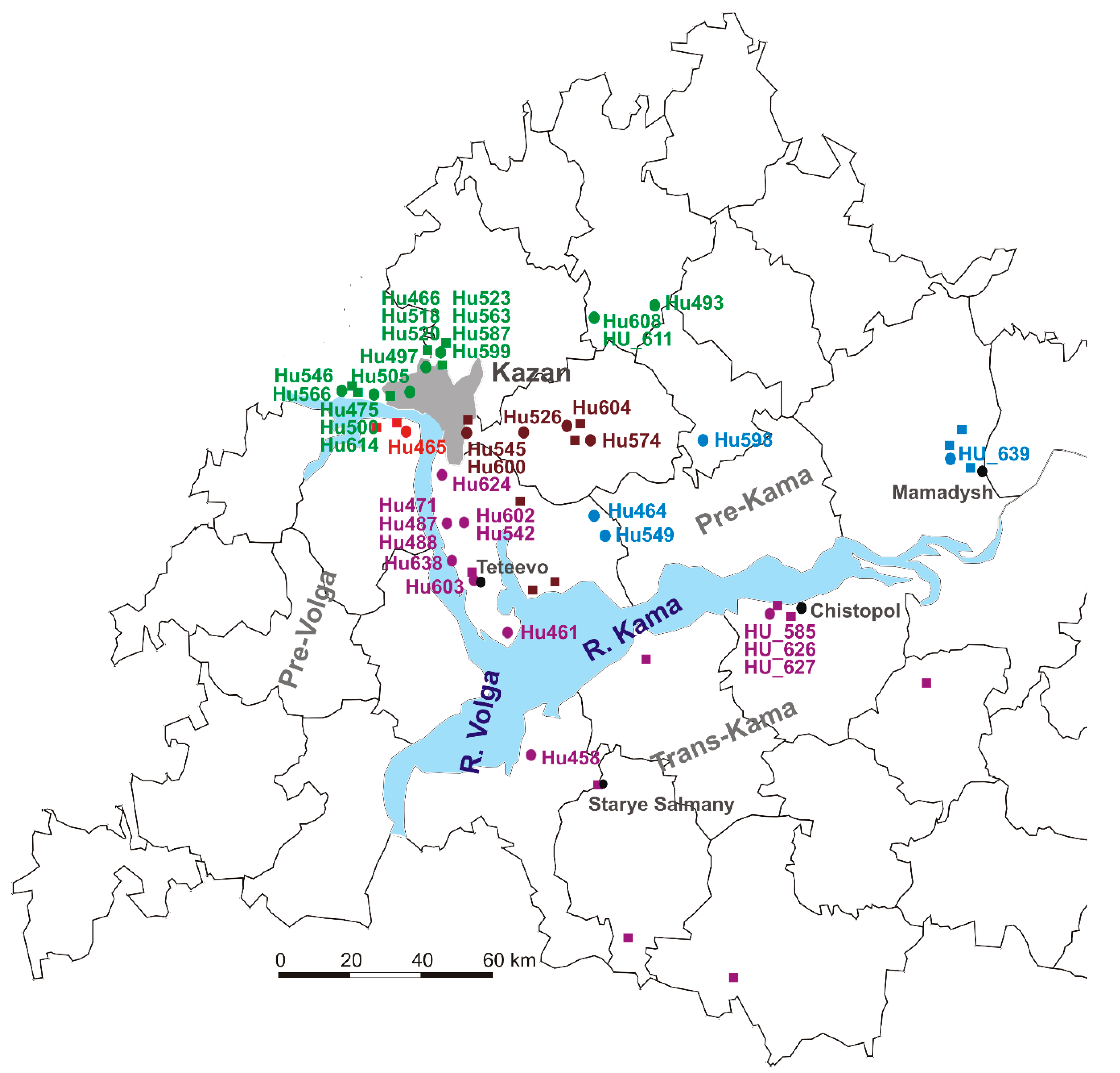
2.1. NE Serum Samples Collection Sites
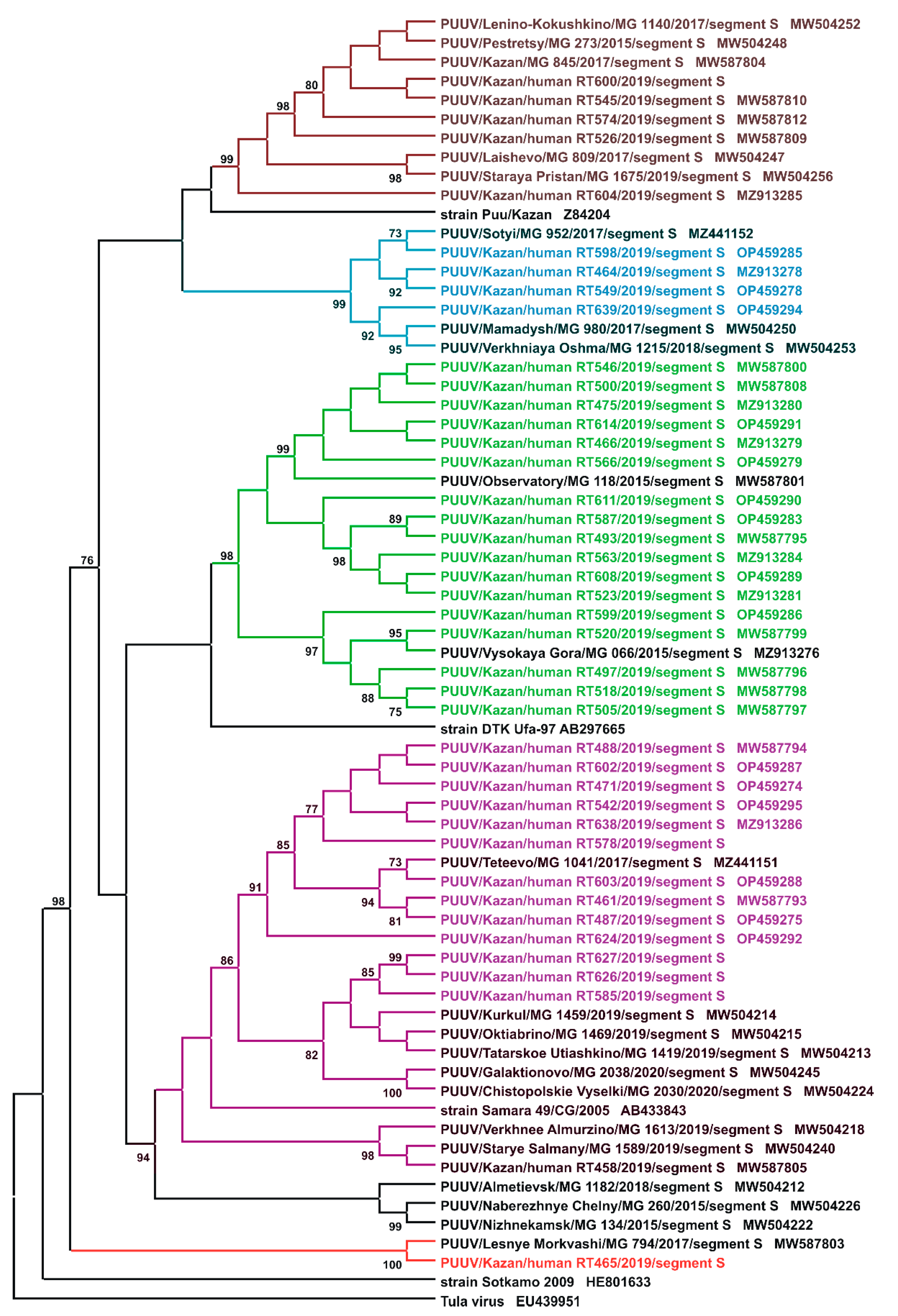
2.2. Sequencing and Phylogenetic Analysis of PUUV Strains from Human and Rodents
2.3. NE Clinical Laboratory Characterization
2.3.1. NE Symptoms and Laboratory Data in Adults
2.3.2. NE Symptoms and Laboratory Data in Children
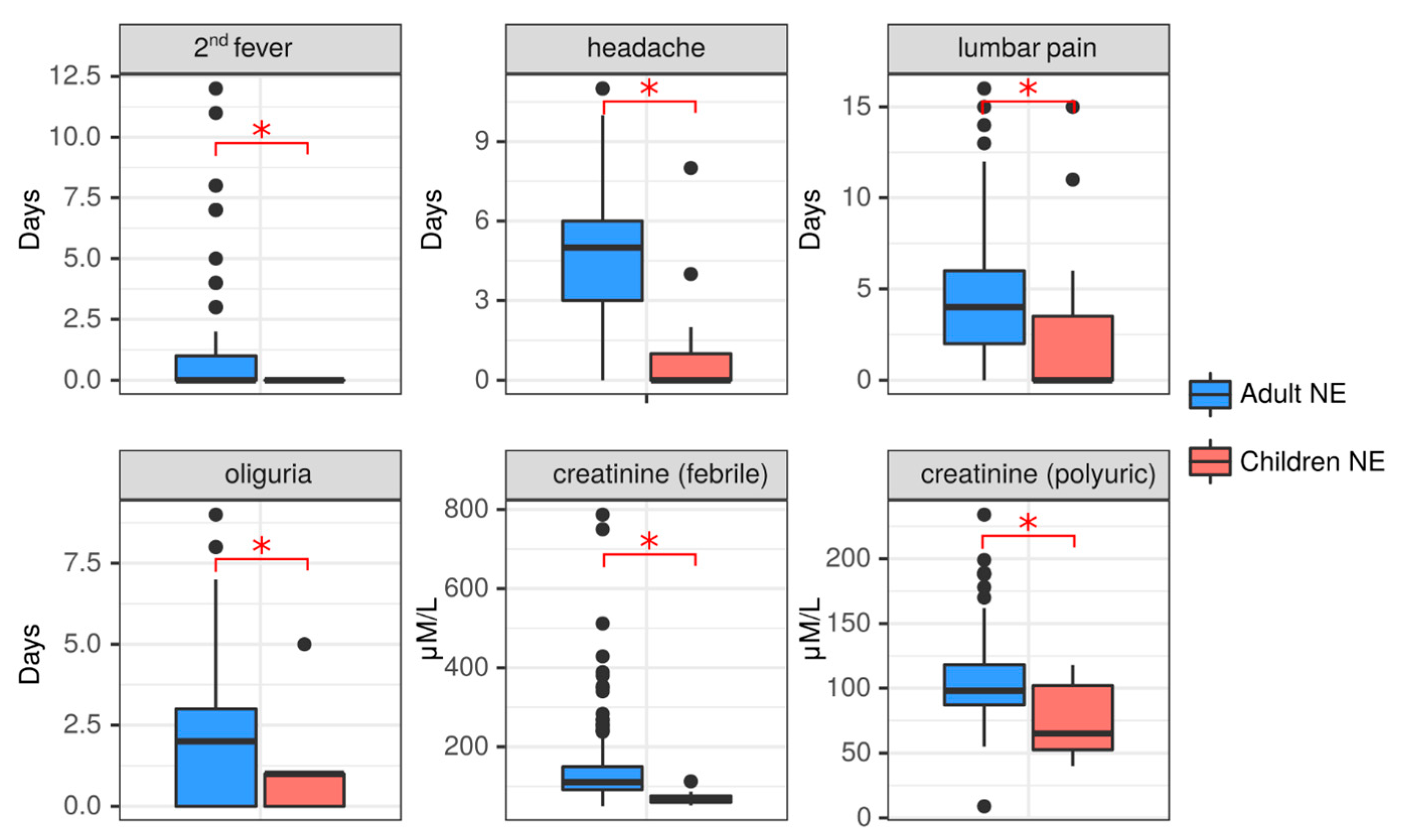
2.3.3. Clinical Comparison of NE in Adult and Pediatric NE Cases
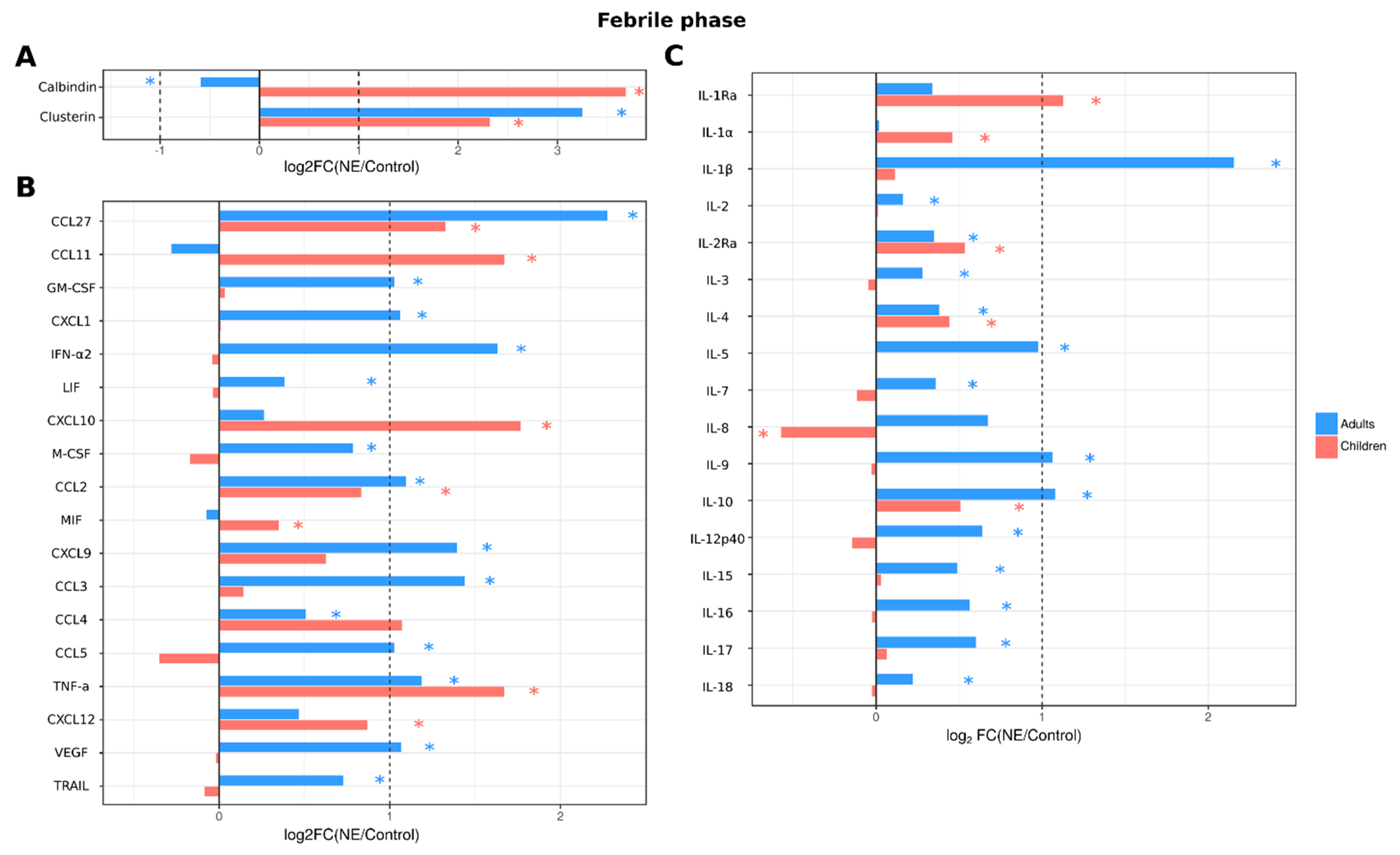
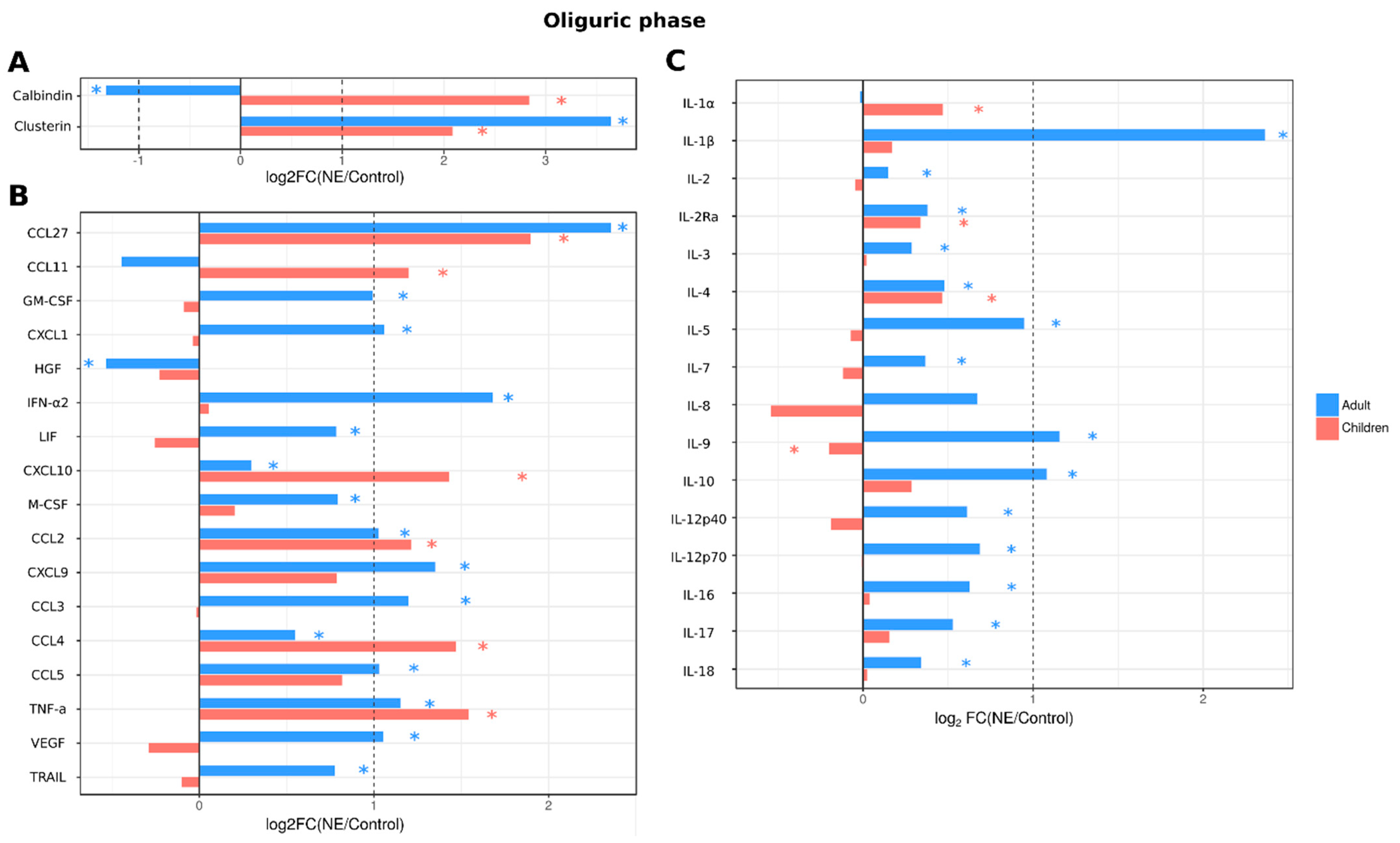
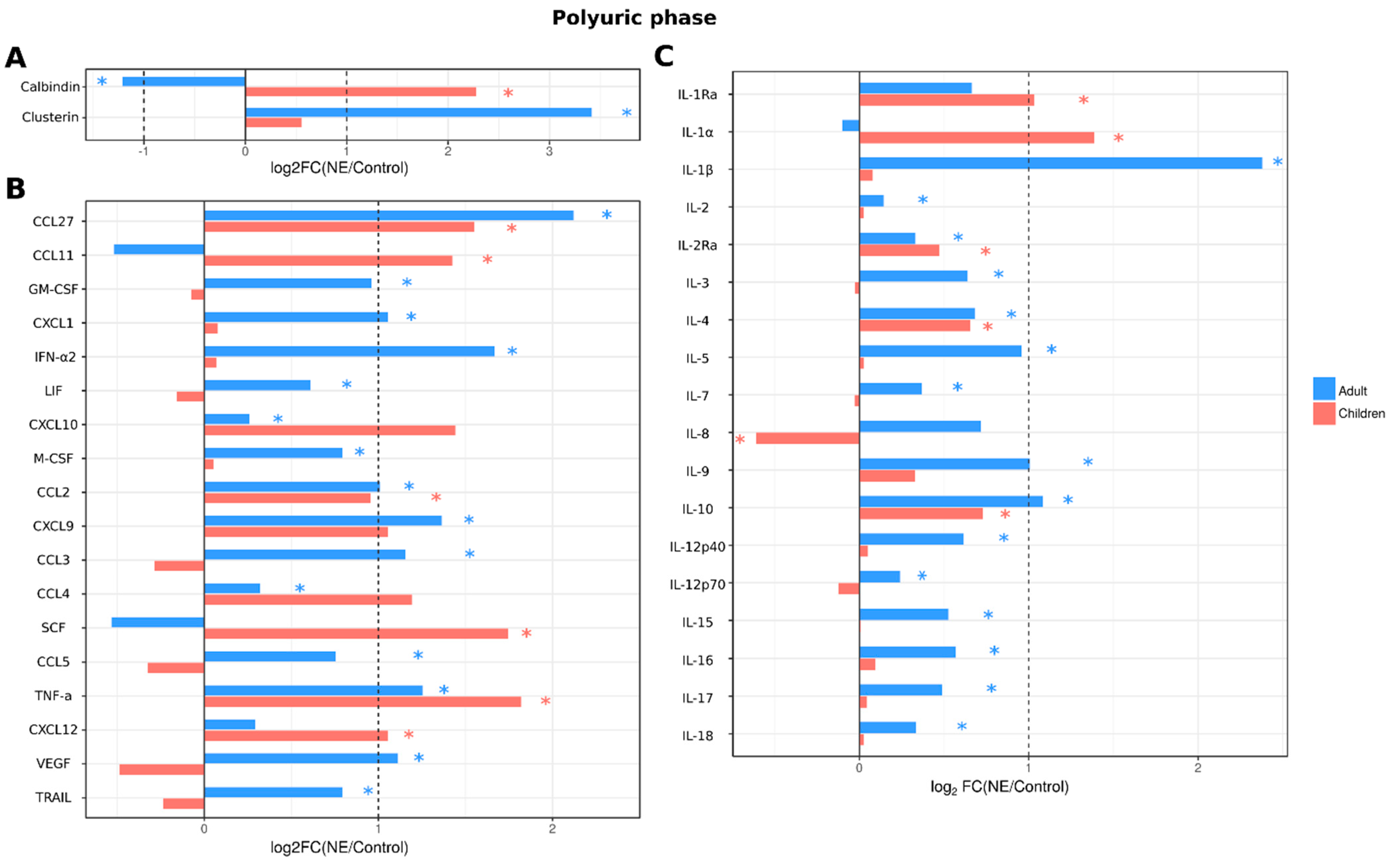
2.3.4. Analysis of Serum Cytokines and Urine Kidney Damage Markers in Adult and Pediatric NE Cases
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Ethics Statement
4.3. Othohantavirus Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. RT-PCR Detection of PUUV Genome
4.5. Phylogenetic Analysis
4.6. Multiplex Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khismatullina, N.A.; Karimov, M.M.; Khaertynov, K.S.; Shuralev, E.A.; Morzunov, S.P.; Khaertynova, I.M.; Ivanov, A.A.; Milova, I.V.; Khakimzyanova, M.B.; Sayfullina, G.; et al. Epidemiological dynamics of nephropathia epidemica in the Republic of Tatarstan, Russia, during the period of 1997–2013. Epidemiol. Infect. 2016, 144, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidyuk, Y.; Shamsutdinov, A.; Kabwe, E.; Ismagilova, R.; Martynova, E.; Belyaev, A.; Shuralev, E.; Trifonov, V.; Savitskaya, T.; Isaeva, G.; et al. Prevalence of the Puumala orthohantavirus Strains in the Pre-Kama Area of the Republic of Tatarstan, Russia. Pathogens 2020, 9, 540. [Google Scholar] [CrossRef] [PubMed]
- Muranyi, W.; Bahr, U.; Zeier, M.; van der Woude, F.J. Hantavirus infection. J. Am. Soc. Nephrol. 2005, 16, 3669–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.A.; Griffen, D.; StatPearls (Internet). Treasure island (FL) StatPearls Publishing; 2022 Jan. Hantavirus Syndrome. (Updated 2022 May 15). Available online: https://www.ncbi.nlm.nih.gov/books/NBK513243 (accessed on 1 November 2020).
- Krautkramer, E.; Zeier, M.; Plyusnin, A. Hantavirus infection: An emerging infectious disease causing acute renal failure. Kidney. Int. 2013, 83, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latus, J.; Schwab, M.; Tacconelli, E.; Pieper, F.M.; Wegener, D.; Rettenmaier, B.; Schwab, A.; Hoffmann, L.; Dippon, J.; Muller, S.; et al. Acute kidney injury and tools for risk-stratification in 456 patients with hantavirus-induced nephropathia epidemica. Nephrol. Dial. Transplant. 2015, 30, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, J.A.; Jorge, S. The RIFLE and AKIN classifications for acute kidney injury: A critical and comprehensive review. Clin. Kidney J. 2013, 6, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanerva, M.; Mustonen, J.; Vaheri, A. Pathogenesis of puumala and other hantavirus infections. Rev. Med. Virol. 1998, 8, 67–86. [Google Scholar] [CrossRef]
- van Ypersele de Strihou, C.; Mery, J.P. Hantavirus-related acute interstitial nephritis in western Europe. Expansion of a world-wide zoonosis. Q. J. Med. 1989, 73, 941–950. [Google Scholar]
- Braun, N.; Haap, M.; Overkamp, D.; Kimmel, M.; Alscher, M.D.; Lehnert, H.; Haas, C.S. Characterization and outcome following Puumala virus infection: A retrospective analysis of 75 cases. Nephrol. Dial. Transplant. 2010, 25, 2997–3003. [Google Scholar] [CrossRef] [Green Version]
- Furberg, M.; Anticona, C.; Schumann, B. Post-infectious fatigue following Puumala virus infection. Infect. Dis. 2019, 51, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Mustonen, J.; Vaheri, A.; Porsti, I.; Makela, S. Long-Term Consequences of Puumala Hantavirus Infection. Viruses 2022, 14, 598. [Google Scholar] [CrossRef] [PubMed]
- Pal, E.; Korva, M.; Resman Rus, K.; Kejzar, N.; Bogovic, P.; Kurent, A.; Avsic-Zupanc, T.; Strle, F. Sequential assessment of clinical and laboratory parameters in patients with hemorrhagic fever with renal syndrome. PLoS ONE 2018, 13, e0197661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peco-Antic, A.; Popovic-Rolovic, M.; Gligic, A.; Popovic, D.; Jovanovic, O.; Kostic, M. Clinical characteristics of haemorrhagic fever with renal syndrome in children. Pediatr. Nephrol. 1992, 6, 335–338. [Google Scholar] [CrossRef]
- Huttunen, N.P.; Makela, S.; Pokka, T.; Mustonen, J.; Uhari, M. Systematic literature review of symptoms, signs and severity of serologically confirmed nephropathia epidemica in paediatric and adult patients. Scand. J. Infect. Dis. 2011, 43, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Makary, P.; Kanerva, M.; Ollgren, J.; Virtanen, M.J.; Vapalahti, O.; Lyytikainen, O. Disease burden of Puumala virus infections, 1995-2008. Epidemiol. Infect. 2010, 138, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, J.; Huttunen, N.P.; Brummer-Korvenkontio, M.; Vaheri, A. Clinical picture of nephropathia epidemica in children. Acta. Paediatr. 1994, 83, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Echterdiek, F.; Kitterer, D.; Alscher, M.D.; Schwenger, V.; Ruckenbrod, B.; Bald, M.; Latus, J. Clinical course of hantavirus-induced nephropathia epidemica in children compared to adults in Germany-analysis of 317 patients. Pediatr. Nephrol. 2019, 34, 1247–1252. [Google Scholar] [CrossRef]
- Lupusoru, G.; Lupusoru, M.; Ailincai, I.; Bernea, L.; Berechet, A.; Spataru, R.; Ismail, G. Hanta hemorrhagic fever with renal syndrome: A pathology in whose diagnosis kidney biopsy plays a major role (Review). Exp. Ther. Med. 2021, 22, 984. [Google Scholar] [CrossRef]
- Outinen, T.K.; Makela, S.; Clement, J.; Paakkala, A.; Porsti, I.; Mustonen, J. Community Acquired Severe Acute Kidney Injury Caused by Hantavirus-Induced Hemorrhagic Fever with Renal Syndrome Has a Favorable Outcome. Nephron 2015, 130, 182–190. [Google Scholar] [CrossRef]
- Satoskar, A.; Nadasdy, T.; Silva, F. Acute postinfectious glomerulonephritis and glomerulonephritis caused by persistent bacterial infection. In Heptinstall’s Pathology of the Kidney, 7th ed.; Jannette, J.G., Olson, J.L., Silva, F.G., D’Agati, V.D., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2015; pp. 678–798. [Google Scholar]
- Krautkrämer, E.; Grouls, S.; Stein, N.; Reiser, J.; Zeier, M. Pathogenic old world hantaviruses infect renal glomerular and tubular cells and induce disassembling of cell-to-cell contacts. J. Virol. 2011, 85, 9811–9823. [Google Scholar] [CrossRef] [Green Version]
- Boehlke, C.; Hartleben, B.; Huber, T.B.; Hopfer, H.; Walz, G.; Neumann-Haefelin, E. Hantavirus infection with severe proteinuria and podocyte foot-process effacement. Am. J. Kidney Dis. 2014, 64, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Hägele, S.; Müller, A.; Nusshag, C.; Reiser, J.; Zeier, M.; Krautkrämer, E. Motility of human renal cells is disturbed by infection with pathogenic hantaviruses. BMC Infect. Dis. 2018, 18, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusshag, C.; Stütz, A.; Hägele, S.; Speer, C.; Kälble, F.; Eckert, C.; Brenner, T.; Weigand, M.A.; Morath, C.; Reiser, J. Glomerular filtration barrier dysfunction in a self-limiting, RNA virus-induced glomerulopathy resembles findings in idiopathic nephrotic syndromes. Sci. Rep. 2020, 10, 19117. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Ushiyama, C.; Takahashi, Y.; Tanaka, A.; Shimada, N.; Ebihara, I.; Koide, H. Effect of dilazep dihydrochloride on urinary albumin excretion in patients with autosomal dominant polycystic kidney disease. Nephron 2001, 88, 80–82. [Google Scholar] [CrossRef]
- Mantula, P.S.; Outinen, T.K.; Clement, J.P.; Huhtala, H.S.; Pörsti, I.H.; Vaheri, A.; Mustonen, J.T.; Mäkelä, S.M. Glomerular proteinuria predicts the severity of acute kidney injury in Puumala hantavirus-induced tubulointerstitial nephritis. Nephron 2017, 136, 193–201. [Google Scholar] [CrossRef]
- Outinen, T.K.; Mantula, P.; Laine, O.K.; Porsti, I.; Vaheri, A.; Makela, S.M.; Mustonen, J. Haematuria is a marker for the severity of acute kidney injury but does not associate with thrombocytopenia in acute Puumala hantavirus infection. Infect. Dis. (Lond.) 2017, 49, 840–846. [Google Scholar] [CrossRef] [Green Version]
- Tietäväinen, J.; Mantula, P.; Outinen, T.; Huhtala, H.; Pörsti, I.H.; Niemelä, O.; Vaheri, A.; Mäkelä, S.; Mustonen, J. Glucosuria predicts the severity of Puumala hantavirus infection. Kidney Int. Rep. 2019, 4, 1296–1303. [Google Scholar] [CrossRef] [Green Version]
- Hansson, M.; Gustafsson, R.; Jacquet, C.; Chebaane, N.; Satchell, S.; Thunberg, T.; Ahlm, C.; Fors Connolly, A.-M. Cystatin C and α-1-microglobulin predict severe acute kidney injury in patients with hemorrhagic fever with renal syndrome. Pathogens 2020, 9, 666. [Google Scholar] [CrossRef]
- Martynova, E.V.; Maksudova, A.N.; Shakirova, V.G.; Abdulkhakov, S.R.; Khaertynova, I.M.; Anokhin, V.A.; Ivanova, V.V.; Abiola, I.M.; Garanina, E.E.; Tazetdinova, L.G. Urinary clusterin is upregulated in nephropathia epidemica. Dis. Markers 2018, 2018, 8658507. [Google Scholar] [CrossRef] [Green Version]
- Dvergsten, J.; Manivel, J.C.; Correa-Rotter, R.; Rosenberg, M.E. Expression of clusterin in human renal diseases. Kidney Int. 1994, 45, 828–835. [Google Scholar] [CrossRef] [Green Version]
- Correa-Rotter, R.; Ibarra-Rubio, M.E.; Schwochau, G.; Cruz, C.; Silkensen, J.R.; Pedraza-Chaverri, J.; Chmielewski, D.; Rosenberg, M.E. Induction of clusterin in tubules of nephrotic rats. J. Am. Soc. Nephrol. 1998, 9, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Davidyuk, Y.N.; Kabwe, E.; Shakirova, V.G.; Martynova, E.V.; Ismagilova, R.K.; Khaertynova, I.M.; Khaiboullina, S.F.; Rizvanov, A.A.; Morzunov, S.P. Characterization of the Puumala orthohantavirus strains in the northwestern region of the Republic of Tatarstan in relation to the clinical manifestations in hemorrhagic fever with renal syndrome patients. Front. Pharmacol. 2019, 10, 970. [Google Scholar] [CrossRef] [PubMed]
- Davidyuk, Y.N.; Kabwe, E.; Shamsutdinov, A.F.; Knyazeva, A.V.; Martynova, E.V.; Ismagilova, R.K.; Trifonov, V.A.; Savitskaya, T.A.; Isaeva, G.S.; Urbanowicz, R.A. The Distribution of Puumala orthohantavirus Genome Variants Correlates with the Regional Landscapes in the Trans-Kama Area of the Republic of Tatarstan. Pathogens 2021, 10, 1169. [Google Scholar] [CrossRef]
- Kellum, J.; Lameire, N.; Aspelin, P.; Barsoum, R.; Burdmann, E.; Goldstein, S. KDIGO AKI guidelines. Kidney Int. 2012, 2, 1–138. [Google Scholar]
- Van de Veerdonk, F.L.; Netea, M.G.; Dinarello, C.A.; Joosten, L.A. Inflammasome activation and IL-1beta and IL-18 processing during infection. Trends Immunol. 2011, 32, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Martynova, E.; Rizvanov, A.; Urbanowicz, R.A.; Khaiboullina, S. Inflammasome Contribution to the Activation of Th1, Th2, and Th17 Immune Responses. Front. Microbiol. 2022, 13, 851835. [Google Scholar] [CrossRef]
- Hatscher, L.; Lehmann, C.H.K.; Purbojo, A.; Onderka, C.; Liang, C.; Hartmann, A.; Cesnjevar, R.; Bruns, H.; Gross, O.; Nimmerjahn, F.; et al. Select hyperactivating NLRP3 ligands enhance the T(H)1-and T(H)17-inducing potential of human type 2 conventional dendritic cells. Sci. Signal. 2021, 14, eabe1757. [Google Scholar] [CrossRef]
- Okamura, H.; Tsutsi, H.; Komatsu, T.; Yutsudo, M.; Hakura, A.; Tanimoto, T.; Torigoe, K.; Okura, T.; Nukada, Y.; Hattori, K.; et al. Cloning of a new cytokine that induces IFN-gamma production by T cells. Nature 1995, 378, 88–91. [Google Scholar] [CrossRef]
- Van Den Eeckhout, B.; Tavernier, J.; Gerlo, S. Interleukin-1 as Innate Mediator of T Cell Immunity. Front. Immunol. 2020, 11, 621931. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 regulates both Th1 and Th2 responses. Annu. Rev. Immunol. 2001, 19, 423–474. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, B.; Zhuang, R.; Zhang, Y.; Liu, B.; Zhang, C.; Yu, H.; Yi, J.; Yang, A.; Jin, B. Hantaan virus infection induces both Th1 and ThGranzyme B+ cell immune responses that associated with viral control and clinical outcome in humans. PLoS Pathog. 2015, 11, e1004788. [Google Scholar] [CrossRef] [PubMed]
- Van Epps, H.L.; Schmaljohn, C.S.; Ennis, F.A. Human memory cytotoxic T-lymphocyte (CTL) responses to Hantaan virus infection: Identification of virus-specific and cross-reactive CD8(+) CTL epitopes on nucleocapsid protein. J. Virol. 1999, 73, 5301–5308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terajima, M.; Van Epps, H.L.; Li, D.; Leporati, A.M.; Juhlin, S.E.; Mustonen, J.; Vaheri, A.; Ennis, F.A. Generation of recombinant vaccinia viruses expressing Puumala virus proteins and use in isolating cytotoxic T cells specific for Puumala virus. Virus Res. 2002, 84, 67–77. [Google Scholar] [CrossRef]
- Kilpatrick, E.D.; Terajima, M.; Koster, F.T.; Catalina, M.D.; Cruz, J.; Ennis, F.A. Role of specific CD8+ T cells in the severity of a fulminant zoonotic viral hemorrhagic fever, hantavirus pulmonary syndrome. J. Immunol. 2004, 172, 3297–3304. [Google Scholar] [CrossRef] [Green Version]
- Kallio-Kokko, H.; Vapalahti, O.; Lundkvist, A.; Vaheri, A. Evaluation of Puumala virus IgG and IgM enzyme immunoassays based on recombinant baculovirus-expressed nucleocapsid protein for early nephropathia epidemica diagnosis. Clin. Diagn. Virol. 1998, 10, 83–90. [Google Scholar] [CrossRef]
- Lundkvist, A.; Hukic, M.; Horling, J.; Gilljam, M.; Nichol, S.; Niklasson, B. Puumala and Dobrava viruses cause hemorrhagic fever with renal syndrome in Bosnia-Herzegovina: Evidence of highly cross-neutralizing antibody responses in early patient sera. J. Med. Virol. 1997, 53, 51–59. [Google Scholar] [CrossRef]
- Bharadwaj, M.; Nofchissey, R.; Goade, D.; Koster, F.; Hjelle, B. Humoral immune responses in the hantavirus cardiopulmonary syndrome. J. Infect. Dis. 2000, 182, 43–48. [Google Scholar] [CrossRef]
- Carson, W.E.; Dierksheide, J.E.; Jabbour, S.; Anghelina, M.; Bouchard, P.; Ku, G.; Yu, H.; Baumann, H.; Shah, M.H.; Cooper, M.A.; et al. Coadministration of interleukin-18 and interleukin-12 induces a fatal inflammatory response in mice: Critical role of natural killer cell interferon-gamma production and STAT-mediated signal transduction. Blood 2000, 96, 1465–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, M.A.; Chadwick, L.J.; Gattone, V.H.; Calvet, J.P. The SGP-2 gene is developmentally regulated in the mouse kidney and abnormally expressed in collecting duct cysts in polycystic kidney disease. Dev. Biol. 1991, 146, 483–490. [Google Scholar] [CrossRef]
- Schwochau, G.B.; Nath, K.A.; Rosenberg, M.E. Clusterin protects against oxidative stress in vitro through aggregative and nonaggregative properties. Kidney Int. 1998, 53, 1647–1653. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jun, H.O.; Yu, Y.S.; Min, B.H.; Park, K.H.; Kim, K.W. Protective effect of clusterin from oxidative stress-induced apoptosis in human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Jun, H.-O.; Kim, D.-h.; Lee, S.-W.; Lee, H.S.; Seo, J.H.; Kim, J.H.; Kim, J.H.; Yu, Y.S.; Min, B.H.; Kim, K.-W. Clusterin protects H9c2 cardiomyocytes from oxidative stress-induced apoptosis via Akt/GSK-3β signaling pathway. Exp. Mol. Med. 2011, 43, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, M.E.; Silkensen, J. Clusterin: Physiologic and pathophysiologic considerations. Int. J. Biochem. Cell Biol. 1995, 27, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Nguan, C.Y.; Guan, Q.; Gleave, M.E.; Du, C. Promotion of cell proliferation by clusterin in the renal tissue repair phase after ischemia-reperfusion injury. Am. J. Physiol. Ren. Physiol. 2014, 306, F724–F733. [Google Scholar] [CrossRef] [Green Version]
- Lambers, T.T.; Mahieu, F.; Oancea, E.; Hoofd, L.; de Lange, F.; Mensenkamp, A.R.; Voets, T.; Nilius, B.; Clapham, D.E.; Hoenderop, J.G.; et al. Calbindin-D28K dynamically controls TRPV5-mediated Ca2+ transport. EMBO J. 2006, 25, 2978–2988. [Google Scholar] [CrossRef] [Green Version]
- Saha, H.; Mustonen, J.; Pietila, K.; Pasternack, A. Metabolism of calcium and vitamin D3 in patients with acute tubulointerstitial nephritis: A study of 41 patients with nephropathia epidemica. Nephron 1993, 63, 159–163. [Google Scholar] [CrossRef]
- Yushchuk, N.D.; Vegerov, Y.Y. Lectures on Infectious Diseases. In Publishing House; GEOTAR-Media: Moscow, Russia, 2001; p. 532. [Google Scholar]
- Yushchuk, N. Infectious Diseases: National Guidelines; Vengerov, Y.Y., Ed.; GEOTAR-Media: Moscow, Russia, 2019; Volume 2, p. 1104. [Google Scholar]
- Dzagurova, T.K.; Ivanov, A.P.; Korotina, N.A.; Malkin, A.E.; Sinyugina, A.A.; Sotskova, S.E.; Ishmukhametov, A.A.; Tkachenko, E.A. Development of Drug Laboratory Methods and Manufacture Technology for Specific Diagnosis of Hemorrhagic Fever with Renal Syndrome. Remedium 2015, 1, 42–47. [Google Scholar] [CrossRef]
- DNASTAR, Inc. Lasergene: Expert Sequence Analysis Software, Version 5; DNASTAR, Inc.: Madison, WI, USA, 2003.
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sign/Symptom | % |
|---|---|
| Fever | 129/129 (100%) |
| 2nd fever | 37/129 (28.68%) |
| Headache | 103/129 (79.84%) |
| Lumbar pain | 110/129 (85.27%) |
| Oliguria | 92/129 (71.32%) |
| Anuria | 5/129 (3.88%) |
| Abdominal pain | 49/129 (37.98%) |
| Nausea | 53/129 (41.09%) |
| Vomiting | 38/129 (29.46%) |
| Diarrhea | 43/129 (33.33%) |
| Cough | 19/129 (14.73%) |
| Hemorrhages | 9/129 (6.98%) |
| Fog in eyes | 49/129 (37.98%) |
| Analyte | Phase | Adult NE (Mean ± SD) | Adult Control (Mean ± SD) | p Value |
|---|---|---|---|---|
| Urea, mM/L | Febrile | 8.39 ± 6.03 | 4.29 ± 0.35 | <0.0001 * |
| Oliguric | 10.13 ± 7.50 | <0.0001 * | ||
| Polyuric | 6.44 ± 8.87 | 0.004 * | ||
| sCr, µM/L | Febrile | 148.85 ± 112.36 | 78.84 ± 10.50 | <0.0001 * |
| Oliguric | 172.64 ± 141.60 | <0.0001 * | ||
| Polyuric | 103.60 ± 31.93 | <0.0001 * | ||
| K, mEq/L | Febrile | 4.04 ± 0.43 | 3.51 ± 0.94 | <0.0001 * |
| Oliguric | 5.43 ± 11.91 | <0.0001 * | ||
| Thrombocytes, cells/μL | Febrile | 90.71 ± 56.30 | 216.31 ± 11.92 | <0.0001 * |
| Oliguric | 143.37 ± 82.75 | <0.0001 * | ||
| Polyuric | 256.71 ± 83.75 | 0.068 |
| Sign/Symptom | Occurrence (%) |
|---|---|
| Fever | 11/11 (100%) |
| 2nd fever | 0/11 (0%) |
| Headache | 3/11 (27.27%) |
| Lumbar pain | 4/11 (36.36%) |
| Oliguria | 7/11 (63.64%) |
| Anuria | 0/11 (0%) |
| Abdominal pain | 5/11 (45.45%) |
| Nausea | 5/11 (54.55%) |
| Vomiting | 5/11 (45.45%) |
| Diarrhea | 1/11 (9.09%) |
| Cough | 0/11 (0%) |
| Hemorrhages | 1/11 (9.09%) |
| Fog in eyes | 3/11 (27.27%) |
| Analyte | Phase | Children NE (Mean ± SD) | Children Control (Mean ± SD) | p Value |
|---|---|---|---|---|
| Urea, mM/L | Febrile | 5.18 ± 1.99 | 2.99 ± 0.51 | <0.0001 * |
| Oliguric | 10.29 ± 9.95 | 0.001 * | ||
| Polyuric | 5.28 ± 1.11 | 0.0002 * | ||
| sCr, µM/L | Febrile | 70.55 ± 17.33 | 43.52 ± 8.72 | 0.21 |
| Oliguric | 144.64 ± 118.82 | 0.003 * | ||
| Polyuric | 75.18 ± 28.85 | 0.018 * | ||
| K, mEq/L | Febrile | 4.06 ± 0.28 | 4.19 ± 0.50 | 0.92 |
| Oliguric | 4.40 ± 0.50 | 0.43 | ||
| Thrombocytes, cells/μL | Febrile | 91.73 ± 51.39 | 235.73 ± 44.10 | 0.0001 * |
| Oliguric | 150.45 ± 69.70 | 0.03 * | ||
| Polyuric | 266.55 ± 68.31 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martynova, E.; Stott-Marshall, R.J.; Shakirova, V.; Saubanova, A.; Bulatova, A.; Davidyuk, Y.N.; Kabwe, E.; Markelova, M.; Khaertynova, I.; Foster, T.L.; et al. Differential Cytokine Responses and the Clinical Severity of Adult and Pediatric Nephropathia Epidemica. Int. J. Mol. Sci. 2023, 24, 7016. https://doi.org/10.3390/ijms24087016
Martynova E, Stott-Marshall RJ, Shakirova V, Saubanova A, Bulatova A, Davidyuk YN, Kabwe E, Markelova M, Khaertynova I, Foster TL, et al. Differential Cytokine Responses and the Clinical Severity of Adult and Pediatric Nephropathia Epidemica. International Journal of Molecular Sciences. 2023; 24(8):7016. https://doi.org/10.3390/ijms24087016
Chicago/Turabian StyleMartynova, Ekaterina, Robert J. Stott-Marshall, Venera Shakirova, Albina Saubanova, Asiya Bulatova, Yuriy N. Davidyuk, Emmanuel Kabwe, Maria Markelova, Ilseyar Khaertynova, Toshana L. Foster, and et al. 2023. "Differential Cytokine Responses and the Clinical Severity of Adult and Pediatric Nephropathia Epidemica" International Journal of Molecular Sciences 24, no. 8: 7016. https://doi.org/10.3390/ijms24087016
APA StyleMartynova, E., Stott-Marshall, R. J., Shakirova, V., Saubanova, A., Bulatova, A., Davidyuk, Y. N., Kabwe, E., Markelova, M., Khaertynova, I., Foster, T. L., & Khaiboullina, S. (2023). Differential Cytokine Responses and the Clinical Severity of Adult and Pediatric Nephropathia Epidemica. International Journal of Molecular Sciences, 24(8), 7016. https://doi.org/10.3390/ijms24087016