
The Complete Chloroplast Genomes of Nine Smilacaceae Species from Hong Kong: Inferring Infra- and Inter-Familial Phylogeny

 , , ,
, , , 
Abstract
:1. Introduction
1.1. The Family Smilacaceae
1.2. The Argument on the Genus Heterosmilax
1.3. Adulteration of Medicinally Valuable Smilacaceae Species and How Chloroplast Genomes Would Help in Authentication
1.4. Chloroplast Genomes of Smilacaceae and Its Potential Application
2. Results
2.1. Genome Size, Structure, and Order
2.2. Genome Content and Structural Analysis
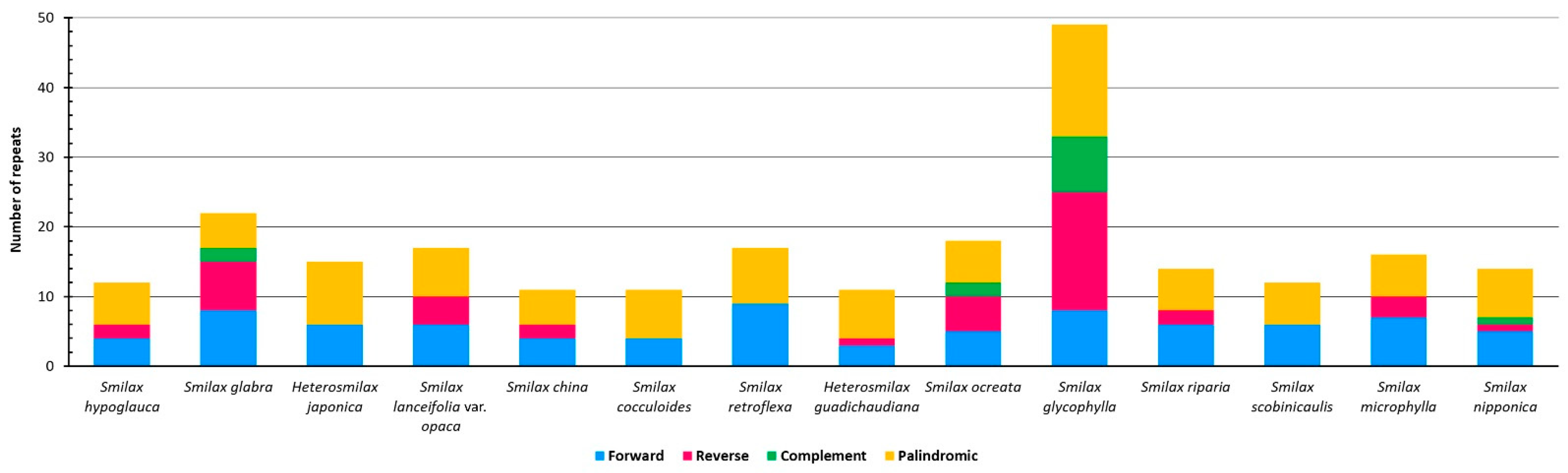
2.2.1. Simple Sequence Repeats (SSRs)
2.2.2. Long Tandem Repeats (LTRs)
2.3. Codon Usage Bias
2.4. Selection Pressure Analysis
2.5. Boundary and Structural Variation
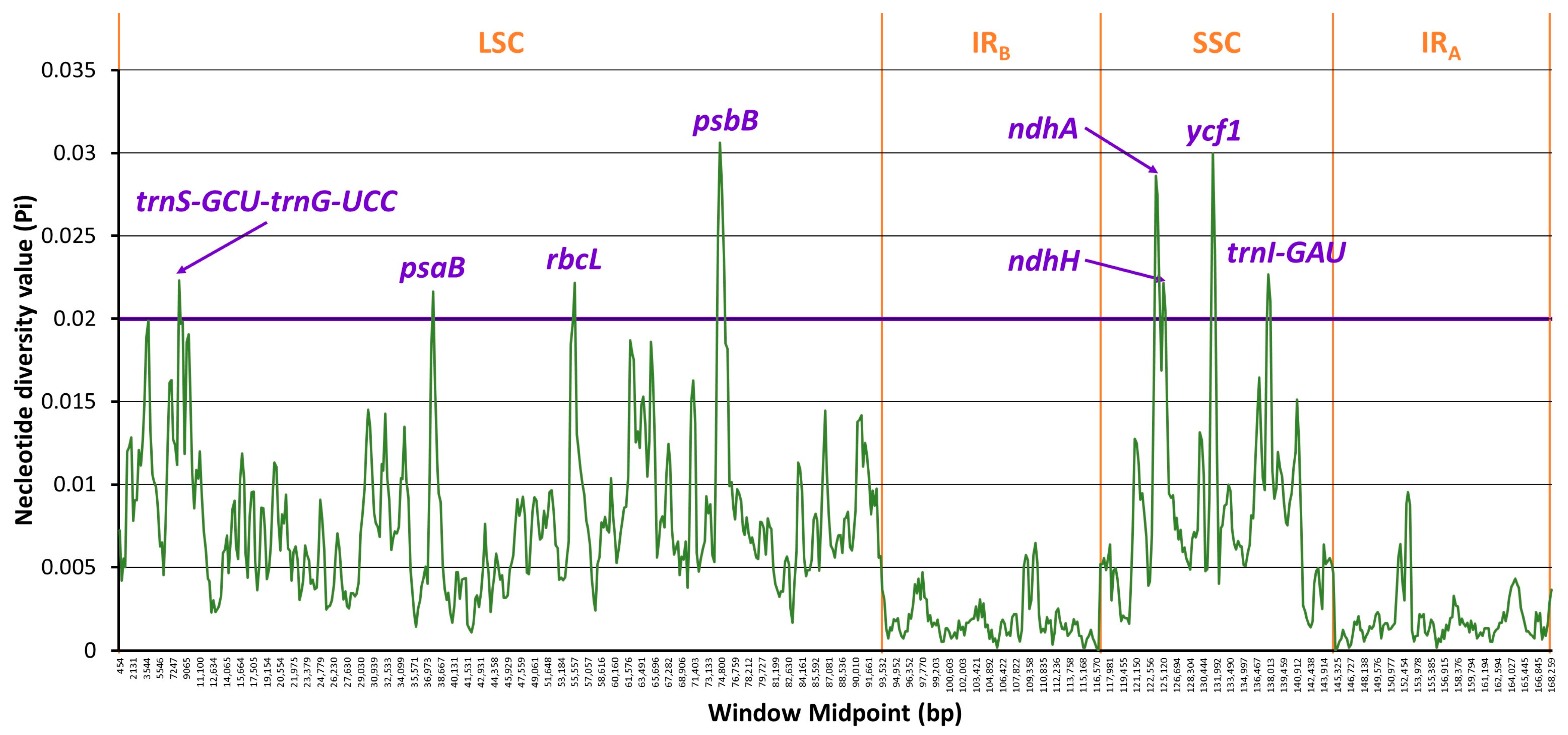
2.6. Divergence Hotspots
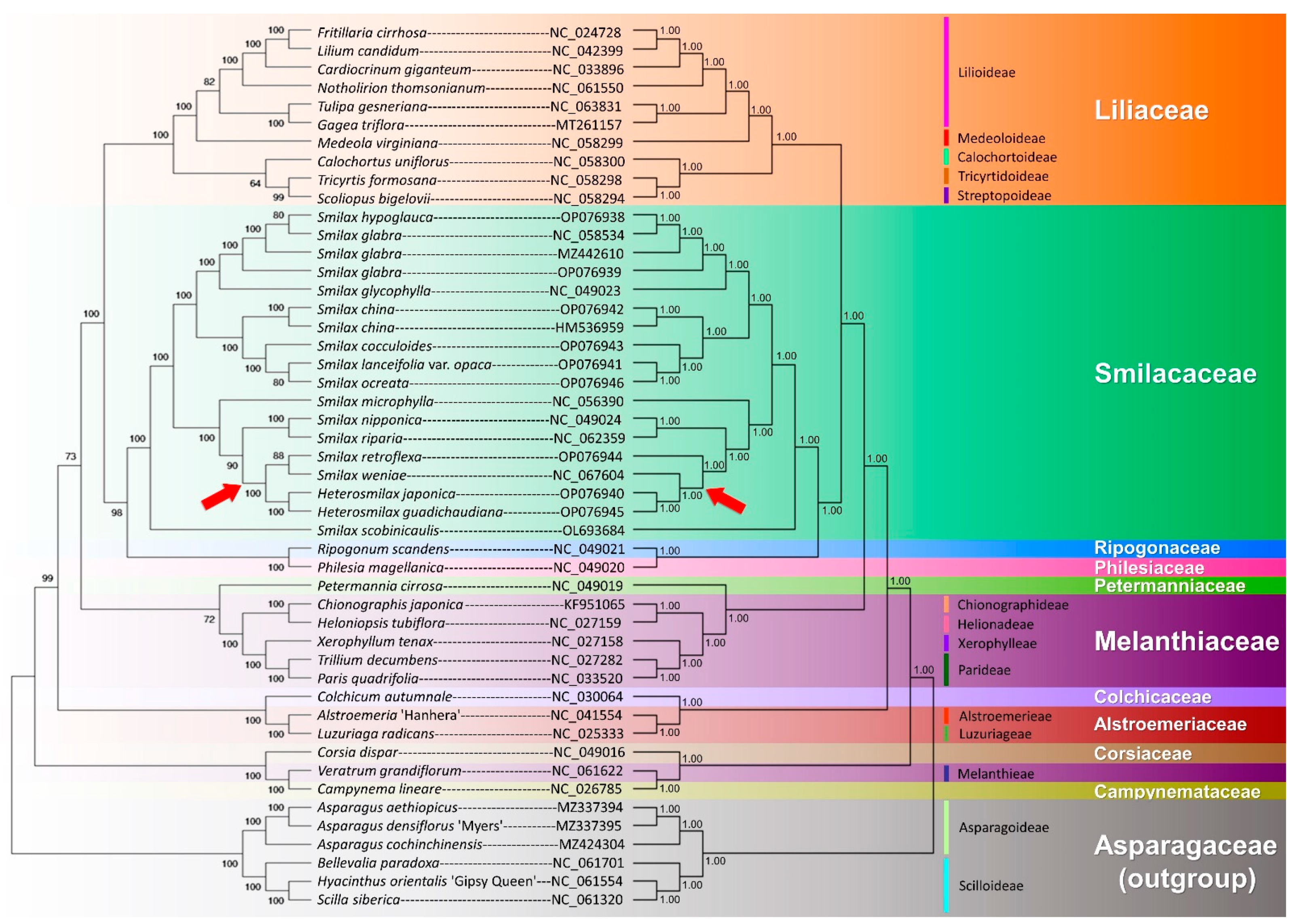
2.7. Phylogenomic Analysis
2.7.1. Family Level
2.7.2. Order Level
3. Discussion
3.1. The Chloroplast Genomes of Smilacaceae
3.1.1. Selection Pressure of PCGs and the Correlation with Environmental Factors
3.1.2. Identification of Divergence Hotspots
3.2. The Status and Taxonomical Revision of Heterosmilax
3.3. The Phylogenomics of Liliales
4. Methods
4.1. Plant Materials and DNA Extraction
4.2. Genome Sequencing, Assembly and Annotation
4.3. Genome Content and Structural Analysis
4.3.1. Sequence Repeats Analysis
4.3.2. Boundary and Structural Variation Analysis
4.3.3. Codon Usage Bias and Selection Pressure Analysis
4.3.4. Nucleotide Diversity Analysis
4.4. Phylogenomic Analysis
4.4.1. Family Level
4.4.2. Order Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, K.M.; Fu, C.X. A Nuclear rDNA Phylogeny of Smilax (Smilacaceae). Aliso 2006, 22, 598–605. [Google Scholar] [CrossRef]
- Kong, H.H.; Wang, A.L.; Lee, J.K.; Fu, C.X. Studies of systematic evolution and karyotypic variation in Smilax and Heterosmilax (Smilacaceae). Acta Phytotaxon. Sin. 2007, 45, 257–273. [Google Scholar] [CrossRef]
- Takhtajan, A.L. Order 16. Smilacales Lindley 1833. In Diversity and Classification of Flowering Plants, 1st ed.; Columbia University Press: New York, NY, USA, 1997; pp. 520–523. ISBN 978-0231100984. [Google Scholar]
- Cronquist, A. 14. Family Smilacaceae Ventenat 1799 nom. conserv., the Catbrier Family. In An Integrated System of Classification of Flowering Plants, 1st ed.; Columbia University Press: New York, NY, USA, 1981; pp. 1225–1228. ISBN 0-231-03880-1. [Google Scholar]
- Chen, S.C.; Zhang, X.P.; Ni, S.F.; Fu, C.X.; Cameron, K.M. The systematic value of pollen morphology in Smilacaceae. Plant Syst. Evol. 2006, 259, 19–37. [Google Scholar] [CrossRef]
- Chen, S.C.; Qiu, Y.X.; Wang, A.L.; Cameron, K.M.; Fu, C.X. A phylogenetic analysis of the Smilacaceae based on morphological data. Acta Phytotaxon. Sin. 2006, 44, 113–125. [Google Scholar] [CrossRef]
- Ventenat, É. Order III. Les Smilacées, Smilaceæ. In Tableau du Règne Végétal, Selon la Méthode de Jussieu, 1st ed.; De l’imprimerie de J. Drisonnier: Paris, France, 1799; Tome Second; pp. 146–148. Available online: https://gallica.bnf.fr/ark:/12148/bpt6k6563774q.texteImage (accessed on 28 March 2023).
- Bantham, G.; Hooker, J.D. Liliaceae. In Genera Plantarum: Ad Exemplaria Imprimis in Herberiis Kewensibus Servata Definite, 1st ed.; Lovell Reeve & Co., Limited: London, UK, 1883; Volume 3, pp. 748–836. Available online: https://www.biodiversitylibrary.org/item/186440#page/768/mode/1up (accessed on 28 March 2023).
- Engler, A.; Prantl, K. Liliaceae. In Die Natürlichen Pflanzenfamilien Nebst Ihren Gattungen und Wichtigeren Arten, Insbesondere den Nutzpflanzen, 1st ed.; Verlag von Wilhelm Engelmann: Leipizig, Germany, 1888; Volume II, pp. 10–91. [Google Scholar] [CrossRef]
- Baker, J.G. Liliaceae. In Flora of Tropical Africa, 1st ed.; Hydrocharideae to Liliacea; Thiselton-Dyer, W.T., Ed.; Lovell Reeve & Co., Limited: London, UK, 1898; Volume 7, pp. 421–568. [Google Scholar] [CrossRef]
- Hutchinson, J. 376. Smilacaceae. In The Families of Flowering Plants Arranged According to a New System Based on Their Probable Phylogeny, 2nd ed.; Monocotyledons; Oxford University Press: Oxford, UK, 1959; Volume II, pp. 618–619. [Google Scholar]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Kunth, C.S. Enumeratio Plantarum Omnium Hucusque Cognitarum, Secundum Familias Naturales Disposita, Adjectis Characteribus, Differentiis, et Synonymis, 1st ed.; Tomus Quintus; Sumtibus JG Collae: Stutgardiae et Tubingae, Germany, 1850; Available online: https://bibdigital.rjb.csic.es/records/item/11045-redirection (accessed on 28 March 2023).
- de Candolle, A. Smilacées. In Smilaceæ, Restiaceæ, Meliaceæ, Monographiæ Phanerogamarum, Suites at Prodromus Systematis Naturalis Regni Vegetabilis, 1st ed.; de Candolle, A., de Candolle, C., Eds.; Sumptibus, G. Masson: Paris, France, 1878; Volume Primum, pp. 1–217. Available online: https://bibdigital.rjb.csic.es/records/item/13090-redirection (accessed on 28 March 2023).
- Hutchinson, J. The Families of Flowering Plants: Arranged According to a New System Based on Their Probable Phylogeny, 1st ed.; Monocotyledons; Macmillan and Co., Limited: London, UK, 1934; Volume II, pp. 106–109. [Google Scholar]
- Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. Smilacaceae Ventenat (1799). In The Families of the Monocotyledons: Structure, Evolution, and Taxonomy, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 127–128. ISBN 3-540-13655-X. [Google Scholar]
- Koyama, T. A taxonomic revision of the genus Heterosmilax (Smilacaceae). Brittonia 1984, 36, 184–205. [Google Scholar] [CrossRef]
- Hsu, T.C.; Chung, S.W. Nomenclatural Alterations in the Genus Heterosmilax (Smilacaceae). Taiwan J. Biodivers. 2012, 14, 51–56. [Google Scholar]
- Qi, Z.C.; Li, P.; Fu, C.X. New combinations and a new name in Smilax for species of Heterosmilax in Eastern and Southeast Asian Smilacaceae (Liliales). Phytotaxa 2013, 117, 58–60. Available online: https://mapress.com/phytotaxa/content/2013/f/p00117p060f.pdf (accessed on 28 March 2023). [CrossRef]
- Judd, W. The Smilacaceae in the Southeastern United States. Harv. Pap. Bot. 1998, 3, 147–169. Available online: http://www.jstor.org/stable/41761566 (accessed on 28 March 2023).
- Chen, S.C.; Seine, N.N.K.; Fu, C.X. Seed coat morphology of Smilacaceae and its systematic significance. J. Syst. Evol. 2007, 45, 52–68. [Google Scholar]
- Qi, Z.C.; Cameron, K.M.; Li, P.; Zhao, Y.P.; Chen, S.C.; Chen, G.C.; Fu, C.X. Phylogenetics, character evolution, and distribution patterns of the greenbriers, Smilacaceae (Liliales), a near-cosmopolitan family of monocots. Bot. J. Linn. Soc. 2013, 173, 535–548. [Google Scholar] [CrossRef]
- Pharmacopoeia Commission of the Ministry of Health of the People’s Republic of China. Pharmacopoeia of the People’s Republic of China 2020, 1st ed.; China Medical Science Press: Beijing, China, 2020; Volume 1, ISBN 978-7-5214-1599-5. (In Chinese)
- Kuai, Y.H.; Hai, L.N.; Jian, X.Y.; Zhang, S.J. Progress in Medicinal Plant of Heterosmilax. Zhongguo Shi Yan Fang Ji Xue Za Zhi 2010, 16, 207–209. (In Chinese) [Google Scholar]
- Bai, Y.M.; Hao, J.D. Benchao Textual research and authentication of Tufuling, Baqia and Bixie. China J. Chin. Mater. Med. 2013, 38, 2733–2737. (In Chinese) [Google Scholar]
- Tang, Y.; He, X.C.; Chen, Q.L.; Fan, L.L.; Zhang, J.Y.; Zhao, Z.Z.; Dong, L.S.; Liang, Z.T.; Yi, T.; Chen, H.B. A mixed microscopic method for differentiating seven species of “Bixie”-related Chinese Materia Medica. Microsc. Res. Tech. 2014, 77, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Li, Y.Q.; Zeng, P.; Zhang, W.M.; Jiang, W.; Yang, Z.G. Botanical resource survey of Tufuling. Chin. Tradit. Herb. Drugs 2002, 33, 850–852. (In Chinese) [Google Scholar]
- Li, W.S.; Fang, Q.M.; Chen, X.; Li, B.; Liang, Y. Utlilization and development of medicinal resources of Tufuling. Chin. Arch. Tradit. Chin. Med. 2003, 21, 517–518. (In Chinese) [Google Scholar]
- Yu, J.C.; Wu, C.Z. Identification of Glabrous Greenbrier Rhizome, Smilax glaucochina and Heterosmilax japonica. Strait Pharm. J. 2010, 22, 39–40. (In Chinese) [Google Scholar]
- Yang, D.L.; Yi, Y.J. Authentication research on Tufuling and its adulterants. Pharm. Clin. Res. 1999, 7, 40–42. (In Chinese) [Google Scholar]
- Commission of National Compendium of Chinese Herbal Medicine. National Compendium of Chinese Herbal Medicine, 1st ed.; People’s Medical Publishing House: Beijing, China, 1975. (In Chinese)
- Hunan Medical Products Administration. Standards of Chinese Medicinal Materials in Hunan Province, 1st ed.; Hunan Science & Technology Press: Changsha, China, 2009; ISBN 9787535759184. (In Chinese)
- Jiang, R.L.; Zhao, P.P.; Chen, J.R.; Chen, W.Q.; Zhong, D.H.; Wang, J.S.; Tang, S.P. Monographic study on Tufuling. In Variety Sorting and Quality Research of Commonly Used Chinese Medicinal Materials, 1st ed.; Cooperation Group of the South, Xu, G.J., Xu, L.S., Eds.; Fujian Science & Technology Publishing House: Fujian, China, 1997; Volume 2, ISBN 7-5335-0818-1. (In Chinese) [Google Scholar]
- Xu, S.; Shang, M.Y.; Liu, G.X.; Xu, F.; Wang, X.; Cai, S.Q. High-Performance Liquid Chromatography Fingerprint Analysis of Smilacis Glabrae Rhizoma and Its Adulterants. Chin. Pharm. J. 2017, 52, 1011–1017. (In Chinese) [Google Scholar]
- Yik, M.H.Y.; Kong, B.L.H.; Siu, T.Y.; Lau, D.T.W.; Cao, H.; Shaw, P.C. Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers. Plants 2021, 10, 161. [Google Scholar] [CrossRef]
- Lee, H.O.; Joh, H.J.; Kim, K.; Lee, S.C.; Kim, N.H.; Park, J.Y.; Park, H.S.; Park, M.S.; Kim, S.; Kwak, M.; et al. Dynamic Chloroplast Genome Rearrangement and DNA Barcoding for Three Apiaceae Species Known as the Medicinal Herb “Bang-Poong”. Int. J. Mol. Sci. 2019, 20, 2196. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. Authentication of Herbal Medicines Dipsacus asper and Phlomoides umbrosa Using DNA Barcodes, Chloroplast Genome, and Sequence Characterized Amplified Region (SCAR) Marker. Molecules 2018, 23, 1748. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qi, Z.C.; Zhao, Y.P.; Fu, C.X.; Xiang, Q.Y. Complete cpDNA genome sequence of Smilax china and phylogenetic placement of Liliales—Influences of gene partitions and taxon sampling. Mol. Phylogenetics Evol. 2012, 64, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Do, H.D.K.; Kim, C.; Chase, M.W.; Kim, J.H. Implications of plastome evolution in the true lilies (monocot order Liliales). Mol. Phylogenetics Evol. 2020, 148, 106818. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Liang, D.; Yang, J. The complete chloroplast genome sequence of Smilax microphylla CH Wright. Mitochondrial DNA B Resour. 2021, 6, 2235–2236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Ge, Q.; Han, K.; Shi, C.; Liu, X. The complete chloroplast genome of a shrub species, Smilax glabra (Smilacaceae) from Guangdong, China. Mitochondrial DNA B Resour. 2021, 6, 3064–3066. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Pei, L.; Cui, N. The complete chloroplast genome and phylogenetic analysis of Smilax moranensis (Liliales: Smilacaceae). Mitochondrial DNA B Resour. 2022, 7, 1206–1207. [Google Scholar] [CrossRef]
- Feng, J.Y.; Jin, X.J.; Zhang, S.L.; Yang, J.W.; Fei, S.P.; Huang, Y.S.; Liu, Y.; Qi, Z.C.; Li, P. Smilax weniae, a New Species of Smilacaceae from Limestone Areas Bordering Guizhou and Guangxi, China. Plants 2022, 11, 1032. [Google Scholar] [CrossRef]
- Mehmood, F.; Ubaid, Z.; Shahzadi, I.; Ahmed, I.; Waheed, M.T.; Poczai, P.; Mirza, B. Plastid genomics of Nicotiana (Solanaceae): Insights into molecular evolution, positive selection and the origin of the maternal genome of Aztec tobacco (Nicotiana rustica). PeerJ 2020, 8, e9552. [Google Scholar] [CrossRef]
- Chen, X.Q.; Koyama, T. 11. SMILAX Linnaeus, Sp. Pl. 2: 1028. 1753. In Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louise, MI, USA, 2000; Volume 24, pp. 96–115. ISBN 7-03-008666-X/Q.995. Available online: http://flora.huh.harvard.edu/china/PDF/PDF24/smilax.pdf (accessed on 28 March 2023).
- Li, H.; Xie, D.F.; Chen, J.P.; Zhou, S.D.; He, X.J. Chloroplast genomic comparison of two sister species Allium macranthum and A. fasciculatum provides valuable insights into adaptive evolution. Genes. Genom. 2020, 42, 507–517. [Google Scholar] [CrossRef]
- Yin, K.; Zhang, Y.; Li, Y.; Du, F.K. Different Natural Selection Pressures on the atpF Gene in Evergreen Sclerophyllous and Deciduous Oak Species: Evidence from Comparative Analysis of the Complete Chloroplast Genome of Quercus aquifolioides with Other Oak Species. Int. J. Mol. Sci. 2018, 19, 1042. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Assadi, M.; Naderi, R.; Kafi, M.; Fatahi, R.; Salami, S.A.; Shariati, V. Complete chloroplast genome of Lilium ledebourii (Baker) Boiss and its comparative analysis: Lights into selective pressure and adaptive evolution. Sci. Rep. 2022, 12, 9375. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lou, G.; Cai, X.; Zhang, B.; Cheng, Y.Q.; Wang, H.W. Comparison of the complete plastomes and the phylogenetic analysis of Paulownia species. Sci. Rep. 2020, 10, 2255. [Google Scholar] [CrossRef]
- Casano, L.M.; Zapata, J.M.; Martin, M.; Sabater, B. Chlororespiration and poising of cyclic electron transport. J. Biol. Chem. 2000, 275, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Jia, Z.Y.; Dayananda, B.; Li, J.Q.; Guo, X.L.; Shi, L.; Yuan, X.W.L.; Gao, Y. Analysis of the chloroplast genomes of four Pinus species in Northeast China: Insights into hybrid speciation and identification of DNA molecular markers. J. For. Res. 2022, 33, 1881–1890. [Google Scholar] [CrossRef]
- Stoppel, R.; Meurer, J. Complex RNA metabolism in the chloroplast: An update on the psbB operon. Planta 2013, 237, 441–449. [Google Scholar] [CrossRef]
- Tang, H.; Tang, L.; Shao, S.; Peng, Y.; Li, L.; Luo, Y. Chloroplast genomic diversity in Bulbophyllum section Macrocaulia (Orchidaceae, Epidendroideae, Malaxideae): Insights into species divergence and adaptive evolution. Plant Divers. 2021, 43, 350–361. [Google Scholar] [CrossRef]
- Du, Y.P.; Bi, Y.; Yang, F.P.; Zhang, M.F.; Chen, X.Q.; Xue, J.; Zhang, X.H. Complete chloroplast genome sequences of Lilium: Insights into evolutionary dynamics and phylogenetic analyses. Sci. Rep. 2017, 7, 5751. [Google Scholar] [CrossRef]
- Wang, L.; Liu, B.; Yang, Y.; Zhuang, Q.; Chen, S.; Liu, Y.; Huang, S. The comparative studies of complete chloroplast genomes in Actinidia (Actinidiaceae): Novel insights into heterogenous variation, clpP gene annotation and phylogenetic relationships. Mol. Genet. Genom. 2022, 297, 535–551. [Google Scholar] [CrossRef]
- Wu, H.Y.; Wong, K.H.; Kong, B.L.H.; Siu, T.Y.; But, G.W.C.; Tsang, S.S.K.; Lau, D.T.W.; Shaw, P.C. Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis. Plants 2022, 11, 1109. [Google Scholar] [CrossRef]
- Wong, K.H.; Kong, B.L.H.; Siu, T.Y.; Wu, H.Y.; But, G.W.C.; Shaw, P.C.; Lau, D.T.W. Complete chloroplast genomes of Asparagus aethiopicus L., A. densiflorus (Kunth) Jessop ‘Myers’, and A. cochinchinensis (Lour.) Merr.: Comparative and phylogenetic analysis with congenerics. PLoS ONE 2022, 17, e0266376. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.B.; Lim, C.E.; Kim, J.S.; Kim, K.H.; Lee, J.H.; Yu, H.J.; Mun, J.H. Comparative chloroplast genome analysis of Artemisia (Asteraceae) in East Asia: Insights into evolutionary divergence and phylogenomic implications. BMC Genom. 2020, 21, 415. [Google Scholar] [CrossRef] [PubMed]
- Namgung, J.; Do, H.D.K.; Kim, C.; Choi, H.J.; Kim, J.H. Complete chloroplast genomes shed light on phylogenetic relationships, divergence time, and biogeography of Allioideae (Amaryllidaceae). Sci. Rep. 2021, 11, 3262. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xia, B.; Li, X. The complete chloroplast genome sequences of five pinnate-leaved Primula species and phylogenetic analyses. Sci. Rep. 2020, 10, 20782. [Google Scholar] [CrossRef]
- Mwanzia, V.M.; He, D.X.; Gichira, A.W.; Li, Y.; Ngarega, B.K.; Karichu, M.J.; Kamau, P.W.; Li, Z.Z. The complete plastome sequences of five Aponogeton species (Aponogetonaceae): Insights into the structural organization and mutational hotspots. Plant Divers. 2020, 42, 334–342. [Google Scholar] [CrossRef]
- Brizicky, G.K. Subgeneric and Sectional Names: Their Starting Points and Early Sources. Taxon 1969, 18, 643–660. [Google Scholar] [CrossRef]
- Hong Kong Herbarium; Agriculture, Fisheries and Conservation Department and South China Botanical Garden; Chinese Academy of Sciences. 384. Smilacaceae. In Flora of Hong Kong, 1st ed.; Agriculture, Fisheries and Conservation Department: Hong Kong, China, 2010; Volume 4, pp. 283–287. ISBN 978-988-98253-9-3. [Google Scholar]
- Hong Kong Herbarium, Agriculture, Fisheries and Conservation Department. Hong Kong Plant Database. Available online: https://www.herbarium.gov.hk/en/home/index.html (accessed on 28 March 2023).
- International Plant Names Index. Smilax gaudichaudiana Kunth. Available online: https://www.ipni.org/n/541453-1 (accessed on 28 March 2023).
- Plants of the World Online. Smilax gaudichaudiana Kunth. Plants of the World Online. 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:541453-1 (accessed on 28 March 2023).
- World Flora Online. Smilax gaudichaudiana Kunth. World Flora Online. 2023. Available online: https://wfoplantlist.org/plant-list/taxon/wfo-0000742682-2022-12 (accessed on 28 March 2023).
- Plants of the World Online. Smilax bockii Warb. Plants of the World Online. 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:541312-1 (accessed on 28 March 2023).
- World Flora Online. Smilax bockii Warb. Ex Diels. World Flora Online. 2023. Available online: https://wfoplantlist.org/plant-list/taxon/wfo-0000740749-2022-12?page=1 (accessed on 28 March 2023).
- International Plant Names Index. Smilax goeringii Kladwong, Chantar. & D.A. Simpson. Available online: https://www.ipni.org/n/77190789-1 (accessed on 28 March 2023).
- Li, P.; Fu, X. iPlant.cn. Xiào Bá Qiā, Smilax japonica (Kunth). Available online: https://www.iplant.cn/info/Smilax%20japonica (accessed on 28 March 2023).
- Kladwong, P.; Chantaranothai, P.; Simpson, D.A. Two new names and five lectotypified taxa for the genus Smilax (Smilacaceae), and the transfer of Smilax petiolatumidus to the genus Dioscorea (Dioscoreaceae). Thai For. Bull. 2018, 46, 44–57. [Google Scholar] [CrossRef]
- Gray, A. International Plant Names Index. Smilax japonica (Kunth). Available online: https://www.ipni.org/n/541535-1 (accessed on 28 March 2023).
- Missouri Botanical Garden. Heterosmilax gaudichaudiana (Kunth) Maxim. Tropicos. 2023. Available online: https://www.tropicos.org/name/18407216 (accessed on 28 March 2023).
- Missouri Botanical Garden. Heterosmilax japonica Kunth. Tropicos. 2023. Available online: https://www.tropicos.org/name/18407087 (accessed on 28 March 2023).
- Kim, J.S.; Hong, J.K.; Chase, M.W.; Fay, M.F.; Kim, J.H. Familial relationships of the monocot order Liliales based on a molecular phylogenetic analysis using four plastid loci: matK, rbcL, atpB and atpF-H. Bot. J. Linn. Soc. 2013, 172, 5–21. [Google Scholar] [CrossRef]
- Hutchinson, J. 376. Smilacaceae. In The Families of Flowering Plants Arranged According to a New System Based on Their Probable Phylogeny, 3rd ed.; Oxford University Press: Oxford, UK, 1973; pp. 764–765. [Google Scholar]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Smilacaceae Ventenat. In Plant Systematics: A Phylogenetic Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; pp. 251–253. ISBN 0-87893-403-0. [Google Scholar]
- Conran, J.G.; Clifford, H.T. The taxonomic affinities of the genus Ripogonum. Nord. J. Bot. 1985, 5, 215–219. [Google Scholar] [CrossRef]
- Takhtajan, A. Order 11. Smilacales. In Flowering Plants, 1st ed.; Springer: Berlin, Germany, 2009; pp. 642–644. ISBN 978-1-4020-9608-2. [Google Scholar]
- Thorne, R. Classification and Geography of the Flowering Plants. Bot. Rev. 1992, 58, 225–348. Available online: http://www.jstor.org/stable/4354190 (accessed on 28 March 2023). [CrossRef]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Smilacaceae Ventenat. In Plant Systematics: A Phylogenetic Approach, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2008; pp. 259–260. ISBN 978-0-87893-407-2. [Google Scholar]
- Rudall, P.J.; Stobart, K.L.; Hong, W.P.; Conran, J.G.; Furness, C.A.; Kite, G.C.; Chase, M.W. Consider the lilies: Systematics of Liliales. In Monocots: Systematics and Evolution, 1st ed.; Wilson, K.L., Morrison, D.A., Eds.; CSIRO: Melbourne, Australia, 2000; Volume 1, pp. 347–359. [Google Scholar]
- Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. Order Melanthiales. In The Families of the Monocotyledons: Structure, Evolution, and Taxonomy, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 207–214. ISBN 3-540-13655-X. [Google Scholar]
- Goldblatt, P. The status of R. Dahlgren’s order Lilales and Melanthiales. In Monocotyledons: Systematics and Evolution, 1st ed.; Rudall, R.J., Cribb, P.J., Cutler, D.F., Humphries, C.J., Eds.; Royal Botanical Gardens: Kew, UK, 1995; Volume 1, pp. 181–200. [Google Scholar]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 2003, 85, 531–553. [Google Scholar]
- Neyland, R. A phylogeny inferred from large subunit (26S) ribosomal DNA sequences suggests that Burmanniales is polyphyletic. Aust. Plant Res. 2002, 15, 19–28. [Google Scholar] [CrossRef]
- Fay, M.F.; Chase, M.W.; Rønsted, N.; Devey, D.S.; Pillon, Y.; Pires, J.C.; Petersen, G.; Seberg, O.; Davis, J.I. Phylogenetics of Liliales: Summarized evidence from combined analyses of five plastid and one mitochondrial loci. Aliso 2006, 22, 559–565. [Google Scholar]
- Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. Order Burmanniales. In The Families of the Monocotyledons: Structure, Evolution, and Taxonomy, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 215–220. ISBN 3-540-13655-X. [Google Scholar]
- Chase, M.W.; Soltis, D.E.; Soltis, P.S.; Rudall, P.J.; Fay, M.F.; Hahn, W.H.; Sullivan, S.; Joseph, J.; Molvray, M.; Kores, P.J.; et al. Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification. In Monocots: Systematics and Evolution, 1st ed.; Wilson, K.L., Morrison, D.A., Eds.; CSIRO: Melbourne, Australia, 2000; Volume 1, pp. 3–16. [Google Scholar]
- Neyland, R.; Hennigan, M. A phylogenetic analysis of large-subunit (26S) ribosome DNA sequences suggests that the Corsiaceae are polyphyletic. N. Z. J. Bot. 2003, 41, 1–11. [Google Scholar] [CrossRef]
- Rudall, P.J.; Eastman, A. The questionable affinities of Corsia (Corsiaceae): Evidence from floral anatomy and pollen morphology. Bot. J. Linn. Soc. 2002, 138, 315–324. [Google Scholar] [CrossRef]
- Mennes, C.B.; Lam, V.K.; Rudall, P.J.; Lyon, S.P.; Graham, S.W.; Smets, E.F.; Merckx, V.S. Ancient Gondwana break-up explains the distribution of the mycoheterotrophic family Corsiaceae (Liliales). J. Biogeogr. 2015, 42, 1123–1136. [Google Scholar] [CrossRef]
- Institutum Botanicum Academiae Sincae. Flora Reipublicae Popularis Sinicae; Tomus 15, Angiospermae, Monocotyledonae, Liliaceae (2); Science Press: Beijing, China, 1978. [Google Scholar]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Plants of the World Online. Smilax moranensis M.Martens & Galeotti. Plants of the World Online. 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:541650-1 (accessed on 28 March 2023).
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Plants of the World Online. Smilax scobinicaulis C.H.Wright. Plants of the World Online. 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:541838-1 (accessed on 28 March 2023).
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The Manifold Applications of Repeat Analysis on a Genomic Scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal Alignment: Finding Rear-rangements During Alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, R. FeatureExtract—Extraction of sequence annotation made easy. Nucleic Acids Res. 2005, 33, W567–W569. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sánchez-Gracia, A. DNA Sequence Polymorphism (DnaSP). Version: 6.12.03 (26 February 2019) [software]. Universitat de Barcelona. Available online: http://www.ub.edu/dnasp/ (accessed on 28 March 2023).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef]
- Gernhard, T. The conditioned reconstructed process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef]
- Yule, G.U. II.—A mathematical theory of evolution, based on the conclusions of Dr. JC Willis, F. R. S. Philos. Trans. R. Soc. Lond. B Contain. Pap. Biol. Character 1925, 213, 21–87. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | OP076938 | OP076939 | OP076940 | OP076941 | OP076942 | OP076943 | OP076944 | OP076945 | OP076946 |
|---|---|---|---|---|---|---|---|---|---|
| Information of the vouchers | |||||||||
| Species | Smilax hypoglauca Benth. | Smilax glabra Roxb. | Heterosmilax japonica Kunth | Smilax lanceifolia Roxb. var. opaca A. DC. | Smilax china L. | Smilax cocculoides Warb. | Smilax retroflexa (F. T. Wang & Tang) S. C. Chen | Heterosmilax guadichaudiana (Kunth) Maxim. | Smilax ocreata A. DC. |
| Collector Number | T. Y. Siu 426 | T. Y. Siu 658 | T. Y. Siu 812 | K. H. Wong 150 | K. H. Wong 154 | K. H. Wong 160 | K. H. Wong 162 | K. H. Wong 169 | K. H. Wong 176 |
| Collection date | 1 February 2020 | 4 November 2020 | 12 July 2021 | 4 March 2021 | 4 March 2021 | 16 March 2021 | 16 March 2021 | 7 June 2021 | 16 July 2021 |
| Inventory no. | CUSLSH2478 | CUSLSH2797 | CUSLSH3069 | CUSLSH2953 | CUSLSH2957 | CUSLSH2972 | CUSLSH2974 | CUSLSH3033 | CUSLSH3071 |
| Sheet no. | CUHK5490 | CUHK5496 | CUHK6147 | CUHK6148–CUHK6150 | CUHK6151–CUHK6152 | CUHK6153–CUHK6154 | CUHK6155–CUHK6157 | CUHK6158 | CUHK6159–CUHK6168 |
| Location | Ma On Shan | Keung Shan | Lantau Peak | Tai Mo Shan | Tai Mo Shan | Sunset Peak | Sunset Peak | CUHK | Tsuen Wan |
| Sex | Female | Female | Male | Female | Female | Female | Male | Male | Female |
| Information of the genomic data | |||||||||
| Raw data (GB) | 3.8 | 3.0 | 3.2 | 3.8 | 3.4 | 4.0 | 3.7 | 3.8 | 3.8 |
| Coverage (×) | 456 | 110 | 169 | 155 | 143 | 134 | 192 | 95 | 151 |
| Information of the assembly and annotation | |||||||||
| Genome size (bp) | 158,118 | 158,538 | 158,059 | 158,385 | 158,269 | 158,418 | 159,007 | 157,885 | 158,223 |
| LSC size (bp) | 85,410 | 85,640 | 85,260 | 85,519 | 85,433 | 85,566 | 85,474 | 85,241 | 85,469 |
| SSC size (bp) | 18,352 | 18,556 | 18,373 | 18,458 | 18,524 | 18,478 | 18,577 | 18,364 | 18,370 |
| IR size (bp) | 27,178 | 27,171 | 27,213 | 27,204 | 27,156 | 27,187 | 27,478 | 27,098 | 27,192 |
| Gene number | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 | 132 |
| mRNA | 86 | 86 | 86 | 86 | 86 | 86 | 86 | 86 | 86 |
| tRNA | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 |
| rRNA | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| ORF | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| 1-intron gene | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 |
| 2-intron gene | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Trans-spliced gene | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| GC content | 37.07% | 37.03% | 37.31% | 37.11% | 37.15% | 37.10% | 37.10% | 37.29% | 37.14% |
| Gene Category | Gene Function | Gene Name |
|---|---|---|
| Photosynthesis-related genes | Rubisco | rbcL |
| Photosystem I | psaA, psaB, psaC, psaI, psaJ | |
| Assembly/stability of photosystem I | pafI **, pafII, pbf1 | |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, psbZ | |
| ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | |
| Cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | |
| Cytochrome c synthesis | ccsA | |
| NADPH dehydrogenase | ndhA *, ndhB * (x2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Transcription- and translation-related genes | Transcription | rpoA, rpoB, rpoC1 *, rpoC2 |
| Ribosomal protein | rpl14, rpl16 *, rpl2 * (x2), rpl20, rpl22, rpl23 (x2), rpl32, rpl33, rpl36, rps2, rps3, rps4, rps7 (x2), rps8, rps11, rps12 * (x2, trans-spliced), rps14, rps15, rps16 *, rps18, rps19 (x2) | |
| RNA genes | Transfer RNA | trnA-UGC * (x2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnG-UCC *, trnH-GUG (x2), trnI-CAU (x2), trnI-GAU * (x2) ^, trnK-UUU *, trnL-CAA (x2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU (x2), trnP-UGG, trnQ-UUG, trnR-ACG (x2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC (x2), trnV-UAC *, trnW-CCA, trnY-GUA |
| Ribosomal RNA | rrn16 (x2), rrn23 (x2), rrn4.5 (x2), rrn5 (x2) | |
| Miscellaneous group | Maturase | matK |
| Inner membrane protein | cemA | |
| ATP-dependent protease | clpP1 ** | |
| Acetyl-CoA carboxylase | accD | |
| Unknown functions | ycf1 (x2), ycf2 (x2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, K.-H.; Siu, T.-Y.; Tsang, S.S.-K.; Kong, B.L.-H.; Wu, H.-Y.; But, G.W.-C.; Hui, J.H.-L.; Shaw, P.-C.; Lau, D.T.-W. The Complete Chloroplast Genomes of Nine Smilacaceae Species from Hong Kong: Inferring Infra- and Inter-Familial Phylogeny. Int. J. Mol. Sci. 2023, 24, 7460. https://doi.org/10.3390/ijms24087460
Wong K-H, Siu T-Y, Tsang SS-K, Kong BL-H, Wu H-Y, But GW-C, Hui JH-L, Shaw P-C, Lau DT-W. The Complete Chloroplast Genomes of Nine Smilacaceae Species from Hong Kong: Inferring Infra- and Inter-Familial Phylogeny. International Journal of Molecular Sciences. 2023; 24(8):7460. https://doi.org/10.3390/ijms24087460
Chicago/Turabian StyleWong, Kwan-Ho, Tin-Yan Siu, Stacey Shun-Kei Tsang, Bobby Lim-Ho Kong, Hoi-Yan Wu, Grace Wing-Chiu But, Jerome Ho-Lam Hui, Pang-Chui Shaw, and David Tai-Wai Lau. 2023. "The Complete Chloroplast Genomes of Nine Smilacaceae Species from Hong Kong: Inferring Infra- and Inter-Familial Phylogeny" International Journal of Molecular Sciences 24, no. 8: 7460. https://doi.org/10.3390/ijms24087460
APA StyleWong, K. -H., Siu, T. -Y., Tsang, S. S. -K., Kong, B. L. -H., Wu, H. -Y., But, G. W. -C., Hui, J. H. -L., Shaw, P. -C., & Lau, D. T. -W. (2023). The Complete Chloroplast Genomes of Nine Smilacaceae Species from Hong Kong: Inferring Infra- and Inter-Familial Phylogeny. International Journal of Molecular Sciences, 24(8), 7460. https://doi.org/10.3390/ijms24087460