
A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak

 ,
,  ,
,  ,
,  ,
, 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
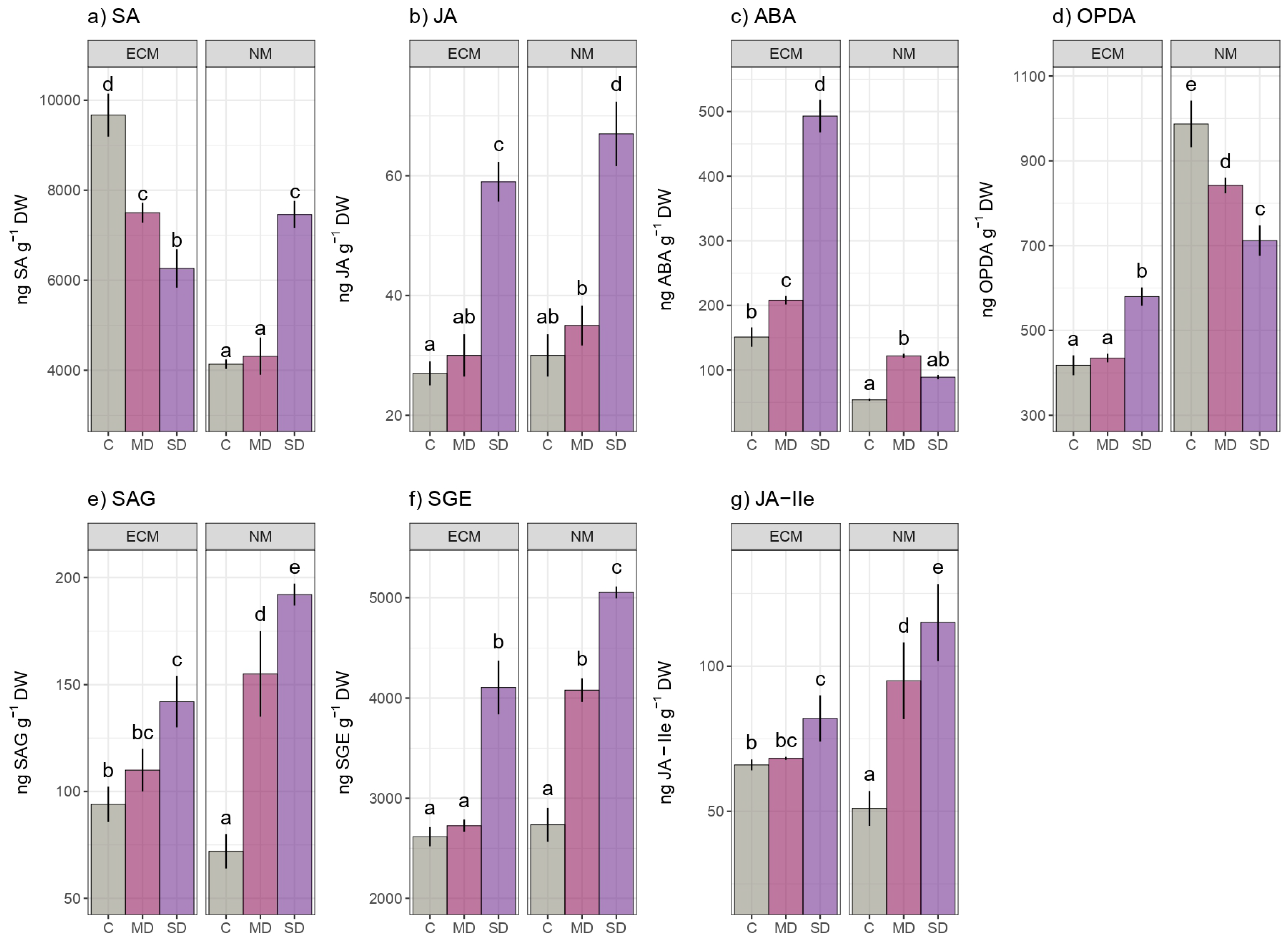
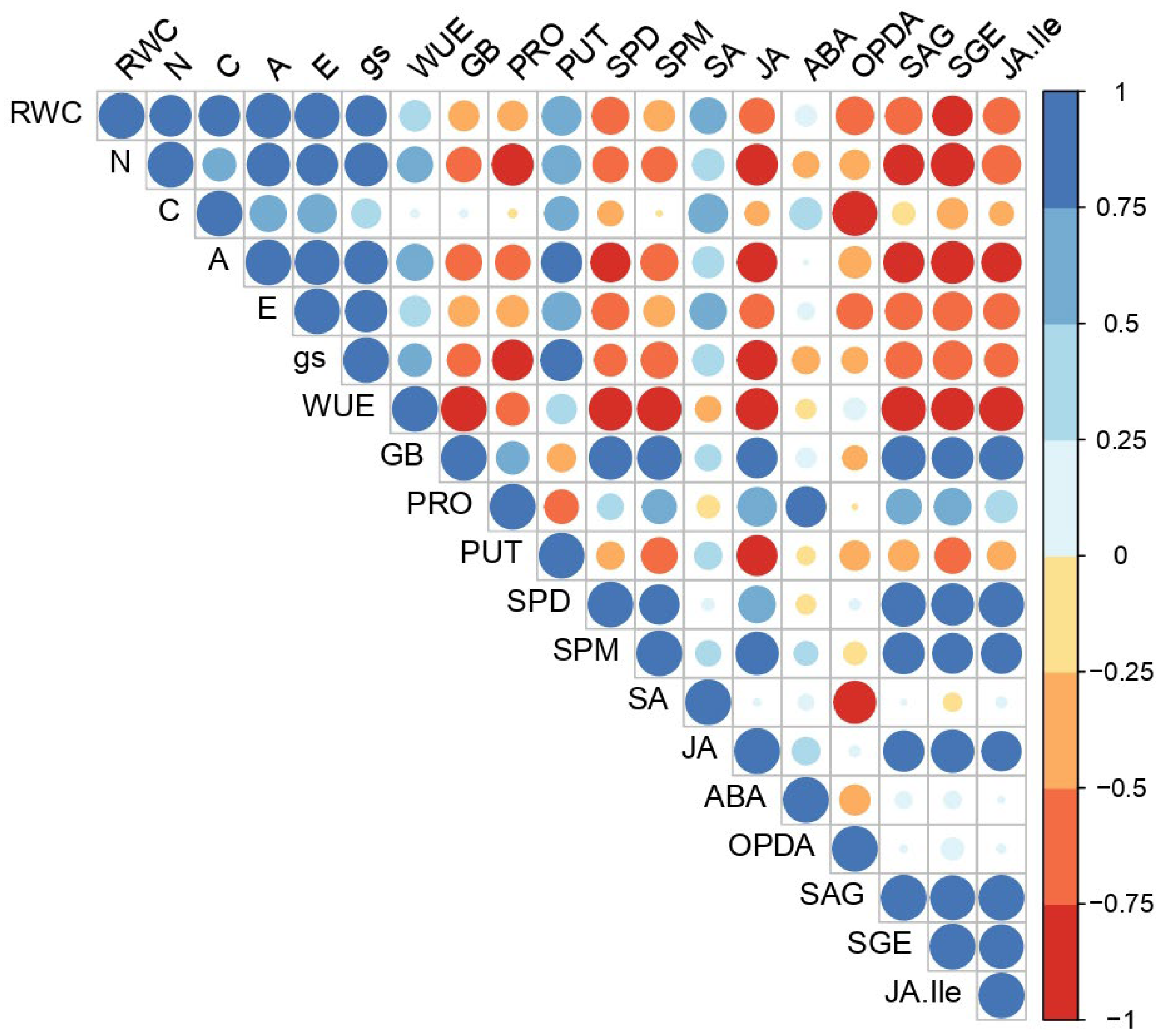
2. Results
2.1. Drought Effect on Carbon, Nitrogen, and Leaf-Relative Water Contents in ECM and NM Seedlings
2.2. Drought Effect on Osmolyte Levels in the Presence and Absence of ECM Fungi
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Physiological Measurements
4.3. Quantification of Osmolytes’ Accumulation
- Polyamines (putrescine (Put), spermidine (Spd), and spermine (Spm)) were extracted from freeze-dried oak leaves (approx. 20 mg DW) with 2 mL of perchloric acid (4% v/v), homogenized in a ball mill, and then centrifuged at 15,000× g for 30 min. The polyamines were derivatized using dansyl-chloride, as previously described by Scaramagli et al. [118]. The separation of different polyamine dansyl-derivateives was performed using a reverse phase C18 column (Spherisorb ODS2, 5-μm particle diameter, 4.6 × 250 mm, Waters, Wexford, Ireland) by high-performance liquid chromatography (HPLC), coupled with fluorescent detection (Shimadzu, Kyoto, Japan) by employing the acetonitrile-water gradient as previously described [118].
- After dyeing with ninhydrin reagent, the free proline was determined spectrophotometrically according to Bates et al. [119] with minor modifications. Around 20 mg of freeze-dried powdered leaf material was homogenized with 1 mL of sulfosalicylic acid (3% w/v) and centrifuged for 10 min at 4000 rpm. A total of 0.7 mL of the supernatant was mixed with 0.7 mL of acid ninhydrin solution (2.5% ninhydrin in glacial acetic acid-distilled water-85% orthophosphoric acid (6:3:1)) and 0.7 mL of glacial acetic acid and the reaction mixture was boiled at +95 °C for 1 h and then transferred to ice bath. The pinkish proline-ninhydrin complex compound was extracted with 2 mL of toluene with vigorous vortexing and the absorbances were recorded at 520 nm by using a MultiScan GO (ThermoScientific, Bremen, Germany) spectrophotometer.
- The concentration of glycine-betaine (GB), as the predominant quaternary ammonium compound (QAC), was determined spectrophotometrically using the precipitation method of QAC-periodide complexes in an acid medium given by Grieve and Grattan [120].
4.4. Plant Hormones Analysis
4.5. Carbon and Nitrogen Elemental Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stojanović, D.; Matović, B.; Orlović, S.; Kržič, A.; Trudić, B.; Galić, Z.; Stojnić, S.; Pekeč, S. Future of the Main Important Forest Tree Species in Serbia from the Climate Change Perspective. South-East Eur. For. 2014, 5, 117–124. [Google Scholar] [CrossRef]
- Kostić, S.; Levanič, T.; Orlović, S.; Matović, B.; Stojanović, D.B. Turkey Oak (Quercus cerris L.) is More Drought Tolerant and Better Reflects Climate Variations Compared to Pedunculate Oak (Quercus robur L.) in Lowland Mixed Forests in Northwestern Serbia: A Stable Carbon Isotope Ratio (Δ13C) and Radial Growth Approach. Ecol. Indic. 2022, 142, 109242. [Google Scholar] [CrossRef]
- Kebert, M.; Vuksanović, V.; Stefels, J.; Bojović, M.; Horák, R.; Kostić, S.; Kovačević, B.; Orlović, S.; Neri, L.; Magli, M.; et al. Species-Level Differences in Osmoprotectants and Antioxidants Contribute to Stress Tolerance of Quercus robur L., and Q. cerris L. Seedlings under Water Deficit and High Temperatures. Plants 2022, 11, 1744. [Google Scholar] [CrossRef]
- Ali, S.; Tyagi, A.; Park, S.; Mir, R.A.; Mushtaq, M.; Bhat, B.; Mahmoudi, H.; Bae, H. Deciphering the Plant Microbiome to Improve Drought Tolerance: Mechanisms and Perspectives. Environ. Exp. Bot. 2022, 201, 104933. [Google Scholar] [CrossRef]
- Stojanović, D.B.; Orlović, S.; Zlatković, M.; Kostić, S.; Vasić, V.; Miletić, B.; Kesić, L.; Matović, B.; Božanić, D.; Pavlović, L.; et al. Climate Change within Serbian Forests: Current State and Future Perspectives. Topola 2021, 208, 39–56. [Google Scholar] [CrossRef]
- Rađević, V.; Pap, P.; Vasić, V. Management of the Common Oak Forests in Ravni Srem: Yesterday, Today, Tomorrow. Topola 2020, 206, 41–52. [Google Scholar] [CrossRef]
- Jokanović, D.; Desimirović, I.; Živanović, K. Taxonomy and Phytogeography Analysis of Medicinal Plants Within Forest Complex of Pedunculate Oak at the Area of Gornji Srem. Topola 2022, 210, 5–14. [Google Scholar] [CrossRef]
- Hermans, S.M.; Lear, G.; Case, B.S.; Buckley, H.L. The Soil Microbiome: An Essential, but Neglected, Component of Regenerative Agroecosystems. iScience 2023, 26, 106028. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Keel, C. Signaling in the Rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef]
- Yu, R.-P.; Lambers, H.; Callaway, R.M.; Wright, A.J.; Li, L. Belowground Facilitation and Trait Matching: Two or Three to Tango? Trends Plant Sci. 2021, 26, 1227–1235. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, Y.; Zhang, N.; Xun, W.; Feng, H.; Miao, Y.; Shao, J.; Shen, Q.; Zhang, R. Chemical Communication in Plant–Microbe Beneficial Interactions: A Toolbox for Precise Management of Beneficial Microbes. Curr. Opin. Microbiol. 2023, 72, 102269. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Pati, D.; Mohapatra, R.; Sahu, B.B.; Singh, P. The Impact of Microbes in Plant Immunity and Priming Induced Inheritance: A Sustainable Approach for Crop Protection. Plant. Stress 2022, 4, 100072. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E.; Qiu, Z.; Singh, B.K. Microbiome-Mediated Stress Resistance in Plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef]
- Bogati, K.; Walczak, M. The Impact of Drought Stress on Soil Microbial Community, Enzyme Activities and Plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–Microbiome Interactions under a Changing World: Responses, Consequences and Perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Leski, T.; Pietras, M.; Rudawska, M. Ectomycorrhizal Fungal Communities of Pedunculate and Sessile Oak Seedlings from Bare-Root Forest Nurseries. Mycorrhiza 2010, 20, 179–190. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Čapelja, E.; Vuksanović, V.; Stojnić, S.; Gavranović Markić, A.; Zlatković, M.; Milović, M.; Galović, V.; Orlović, S. Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content. Plants 2022, 11, 3360. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Zlatković, M.; Stojnić, S.; Čapelja, E.; Zorić, M.; Kiprovski, B.; Budakov, D.; Orlović, S. Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L. Forests 2022, 13, 1491. [Google Scholar] [CrossRef]
- Tarkka, M.T.; Grams, T.E.E.; Angay, O.; Kurth, F.; Maboreke, H.R.; Mailänder, S.; Bönn, M.; Feldhahn, L.; Fleischmann, F.; Ruess, L.; et al. Ectomycorrhizal Fungus Supports Endogenous Rhythmic Growth and Corresponding Resource Allocation in Oak during Various Below- and Aboveground Biotic Interactions. Sci. Rep. 2021, 11, 23680. [Google Scholar] [CrossRef] [PubMed]
- Garbaye, J.; Churin, J.-L. Growth Stimulation of Young Oak Plantations Inoculated with the Ectomycorrhizal Fungus Paxillus Involutus with Special Reference to Summer Drought. Ecol. Manag. 1997, 98, 221–228. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Khan, M.A.R. Understanding the Roles of Osmolytes for Acclimatizing Plants to Changing Environment: A Review of Potential Mechanism. Plant Signal. Behav. 2021, 16, 1913306. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its Actions during the Drought Stress in Plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Salim, B.B.M.; El-Yazied, A.A. Effect of Mycorrhiza on Growth, Biochemical Constituents and Yield of Snap Bean Plants under Water Deficit Conditions. J. Ornam. Hortic. 2015, 7, 131–140. [Google Scholar]
- Malhi, G.S.; Kaur, M.; Kaushik, P.; Alyemeni, M.N.; Alsahli, A.A.; Ahmad, P. Arbuscular Mycorrhiza in Combating Abiotic Stresses in Vegetables: An Eco-Friendly Approach. Saudi J. Biol. Sci. 2021, 28, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.M.; Maiale, S.J.; Rossi, F.R.; Marina, M.; Ruíz, O.A.; Gárriz, A. Polyamine Metabolism Responses to Biotic and Abiotic Stress. In Polyamines: Methods and Protocols; Alcázar, R., Tiburcio, A.F., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; pp. 37–49. ISBN 978-1-4939-7398-9. [Google Scholar]
- Kebert, M.; Rapparini, F.; Neri, L.; Bertazza, G.; Orlović, S.; Biondi, S. Copper-Induced Responses in Poplar Clones are Associated with Genotype- and Organ-Specific Changes in Peroxidase Activity and Proline, Polyamine, ABA, and IAA Levels. J. Plant Growth Regul. 2017, 36, 131–147. [Google Scholar] [CrossRef]
- Velikova, V.B.; Edreva, A.M.; Tsonev, T.D.; Jones, H.G. Singlet Oxygen Quenching by Phenylamides and Their Parent Compounds. Z. Nat. C 2007, 62, 833–838. [Google Scholar] [CrossRef]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Polyamines Increase Nitric Oxide and Reactive Oxygen Species in Guard Cells of Arabidopsis Thaliana during Stomatal Closure. Protoplasma 2018, 255, 153–162. [Google Scholar] [CrossRef]
- Gayatri, G.; Agurla, S.; Raghavendra, A. Nitric Oxide in Guard Cells as an Important Secondary Messenger during Stomatal Closure. Front. Plant Sci. 2013, 4, 425. [Google Scholar] [CrossRef]
- Cvikrová, M.; Gemperlová, L.; Martincová, O.; Vanková, R. Effect of Drought and Combined Drought and Heat Stress on Polyamine Metabolism in Proline-over-Producing Tobacco Plants. Plant. Physiol. Biochem. 2013, 73, 7–15. [Google Scholar] [CrossRef]
- Sharma, K.; Gupta, S.; Thokchom, S.D.; Jangir, P.; Kapoor, R. Arbuscular Mycorrhiza-Mediated Regulation of Polyamines and Aquaporins During Abiotic Stress: Deep Insights on the Recondite Players. Front. Plant Sci. 2021, 12, 642101. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Elucidating the Mechanisms Underlying Enhanced Drought Tolerance in Plants Mediated by Arbuscular Mycorrhizal Fungi. Front. Microbiol. 2021, 12, 809473. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones Enhanced Drought Tolerance in Plants: A Coping Strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Goh, D.M.; Cosme, M.; Kisiala, A.B.; Mulholland, S.; Said, Z.M.F.; Spíchal, L.; Emery, R.J.N.; Declerck, S.; Guinel, F.C. A Stimulatory Role for Cytokinin in the Arbuscular Mycorrhizal Symbiosis of Pea. Front. Plant Sci. 2019, 10, 262. [Google Scholar] [CrossRef]
- Pavlović, I.; Petřík, I.; Tarkowská, D.; Lepeduš, H.; Vujčić Bok, V.; Radić Brkanac, S.; Novák, O.; Salopek-Sondi, B. Correlations between Phytohormones and Drought Tolerance in Selected Brassica Crops: Chinese Cabbage, White Cabbage and Kale. Int. J. Mol. Sci. 2018, 19, 2866. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant Hormone-Mediated Regulation of Stress Responses. BMC Plant. Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- Fernández, J.; Moreno, L. Mechanisms for Tolerance to Water-Deficit Stress in Plants Inoculated with Arbuscular Mycorrhizal Fungi. A Review. Agron. Colomb. 2016, 34, 179–189. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef]
- Salvi, P.; Manna, M.; Kaur, H.; Thakur, T.; Gandass, N.; Bhatt, D.; Muthamilarasan, M. Phytohormone Signaling and Crosstalk in Regulating Drought Stress Response in Plants. Plant. Cell Rep. 2021, 40, 1305–1329. [Google Scholar] [CrossRef] [PubMed]
- Rapparini, F.; Peñuelas, J. Mycorrhizal Fungi to Alleviate Drought Stress on Plant Growth. In Use of Microbes for the Alleviation of Soil Stresses; Miransari, M., Ed.; Springer: New York, NY, USA, 2014; Volume 1, pp. 21–42. ISBN 978-1-4614-9466-9. [Google Scholar]
- Stuart, E.K.; Plett, K.L. Digging Deeper: In Search of the Mechanisms of Carbon and Nitrogen Exchange in Ectomycorrhizal Symbioses. Front. Plant Sci. 2020, 10, 1658. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.; Kim, D.-G.; Li, J.; Liu, Y.; Hai, X.; Liu, Q.; Huang, C.; Shangguan, Z.; Kuzyakov, Y. Drought Effects on Soil Carbon and Nitrogen Dynamics in Global Natural Ecosystems. Earth Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]
- Han, M.-L.; Lv, Q.-Y.; Zhang, J.; Wang, T.; Zhang, C.-X.; Tan, R.-J.; Wang, Y.-L.; Zhong, L.-Y.; Gao, Y.-Q.; Chao, Z.-F.; et al. Decreasing Nitrogen Assimilation under Drought Stress by Suppressing DST-Mediated Activation of Nitrate Reductase 1.2 in Rice. Mol. Plant. 2022, 15, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Yadollahi, A.; Arzani, K.; Imani, A.; Aghaalikhani, M. Gas-Exchange Response of Almond Genotypes to Water Stress. Photosynthetica 2015, 53, 29–34. [Google Scholar] [CrossRef]
- Yan, W.; Zhong, Y.; Shangguan, Z. A Meta-Analysis of Leaf Gas Exchange and Water Status Responses to Drought. Sci. Rep. 2016, 6, 20917. [Google Scholar] [CrossRef]
- Xiong, S.; Wang, Y.; Chen, Y.; Gao, M.; Zhao, Y.; Wu, L. Effects of Drought Stress and Rehydration on Physiological and Biochemical Properties of Four Oak Species in China. Plants 2022, 11, 679. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Nguyen, H.T.; Holaday, A.S. Leaf Water Content and Gas-Exchange Parameters of Two Wheat Genotypes Differing in Drought Resistance. Crop. Sci. 1990, 30, 105–111. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Yamanaka, T.; Yoshimura, K.; Kosugi, Y. Ectomycorrhizal-Fungal Colonization Induces Physio-Morphological Changes in Quercus serrata Leaves and Roots. J. Plant Nutr. Soil. Sci. 2012, 175, 900–906. [Google Scholar] [CrossRef]
- Heinonsalo, J.; Juurola, E.; Linden, A.; Pumpanen, J. Ectomycorrhizal Fungi Affect Scots Pine Photosynthesis through Nitrogen and Water Economy, Not Only through Increased Carbon Demand. Environ. Exp. Bot. 2015, 109, 103–112. [Google Scholar] [CrossRef]
- Wright, D.P.; Scholes, J.D.; Read, D.J.; Rolfe, S.A. Changes in Carbon Allocation and Expression of Carbon Transporter Genes in Betula pendula Roth. Colonized by the Ectomycorrhizal Fungus Paxillus involutus (Batsch) Fr. Plant. Cell Environ. 2000, 23, 39–49. [Google Scholar] [CrossRef]
- Hu, B.; Simon, J.; Rennenberg, H. Drought and Air Warming Affect the Species-Specific Levels of Stress-Related Foliar Metabolites of Three Oak Species on Acidic and Calcareous Soil. Tree Physiol. 2013, 33, 489–504. [Google Scholar] [CrossRef]
- Hu, B.; Simon, J.; Günthardt-Goerg, M.S.; Arend, M.; Kuster, T.M.; Rennenberg, H. Changes in the Dynamics of Foliar N Metabolites in Oak Saplings by Drought and Air Warming Depend on Species and Soil Type. PloS ONE 2015, 10, e0126701. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant Survival under Drought Stress: Implications, Adaptive Responses, and Integrated Rhizosphere Management Strategy for Stress Mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, Salt, and Temperature Stress-Induced Metabolic Rearrangements and Regulatory Networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Niinemets, Ü. Uncovering the Hidden Facets of Drought Stress: Secondary Metabolites Make the Difference. Tree Physiol. 2015, 36, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Mukarram, M.; Khan, M.M.A.; Zehra, A.; Petrik, P.; Kurjak, D. Suffer or Survive: Decoding Salt-Sensitivity of Lemongrass and its Implication on Essential Oil Productivity. Front. Plant Sci. 2022, 13, 903954. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.E.; Savouré, A.; Szabados, L. Proline Metabolism as Regulatory Hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of Proline Biosynthesis, Degradation, Uptake and Transport in Higher Plants: Its Implications in Plant Growth and Abiotic Stress Tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Yoshiba, Y.; Kiyosue, T.; Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of Levels of Proline as an Osmolyte in Plants under Water Stress. Plant. Cell Physiol. 1997, 38, 1095–1102. [Google Scholar] [CrossRef]
- Wu, H.-H.; Zou, Y.-N.; Rahman, M.M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas Alter Sucrose and Proline Metabolism in Trifoliate Orange Exposed to Drought Stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef]
- Aganchich, B.; Wahbi, S.; Yaakoubi, A.; El-Aououad, H.; Bota, J. Effect of Arbuscular Mycorrhizal Fungi Inoculation on Growth and Physiology Performance of Olive Trees under Regulated Deficit Irrigation and Partial Rootzone Drying. S. Afr. J. Bot. 2022, 148, 1–10. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-um, S. Arbuscular Mycorrhizal Fungi (AMF) Improved Water Deficit Tolerance in Two Different Sweet Potato Genotypes Involves Osmotic Adjustments via Soluble Sugar and Free Proline. Sci. Hortic. 2016, 198, 107–117. [Google Scholar] [CrossRef]
- Karunanantham, K.; Lakshminarayanan, S.P.; Ganesamurthi, A.K.; Ramasamy, K.; Rajamony, V.R. Chapter 10–Arbuscular Mycorrhiza—A Health Engineer for Abiotic Stress Alleviation. In Rhizosphere Engineering; Dubey, R.C., Kumar, P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 171–198. ISBN 978-0-323-89973-4. [Google Scholar]
- Hagh-Doust, N.; Färkkilä, S.M.A.; Hosseyni Moghaddam, M.S.; Tedersoo, L. Symbiotic Fungi as Biotechnological Tools: Methodological Challenges and Relative Benefits in Agriculture and Forestry. Fungal. Biol. Rev. 2022, 42, 34–55. [Google Scholar] [CrossRef]
- Cao, X.; Wu, L.; Wu, M.; Zhu, C.; Jin, Q.; Zhang, J. Abscisic Acid Mediated Proline Biosynthesis and Antioxidant Ability in Roots of Two Different Rice Genotypes under Hypoxic Stress. BMC Plant Biol. 2020, 20, 198. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycinebetaine: An Effective Protectant against Abiotic Stress in Plants. Trends Plant. Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef]
- He, C.; Zhang, W.; Gao, Q.; Yang, A.; Hu, X.; Zhang, J. Enhancement of Drought Resistance and Biomass by Increasing the Amount of Glycine Betaine in Wheat Seedlings. Euphytica 2011, 177, 151–167. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine Betaine Counteracts the Inhibitory Effects of Waterlogging on Growth, Photosynthetic Pigments, Oxidative Defence System, Nutrient Composition, and Fruit Quality in Tomato. J. Hortic. Sci. Biotechnol. 2018, 93, 385–391. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Aslanpour, M.; Doulati Baneh, H.; Tehranifar, A.; Shoor, M. The Effect of Mycorrhizal Fungi on the Amount of Glycine Betaine, Soluble Sugar, Proline, Leaf Water Content and Leaf Chlorophyll of the White Seedless Grape under Drought Stress Conditions. Int. J. Adv. Biotechnol. Res. 2016, 7, 1119–1133. [Google Scholar]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines Are Important in Abiotic Stress Signaling. Plant. Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant. Scie 2019, 9, 1945. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Liu, K.; Wang, Z.; Liu, L. Involvement of Polyamines in the Drought Resistance of Rice. J. Exp. Bot. 2007, 58, 1545–1555. [Google Scholar] [CrossRef]
- Liu, M.; Chen, J.; Guo, Z.; Lu, S. Differential Responses of Polyamines and Antioxidants to Drought in a Centipedegrass Mutant in Comparison to Its Wild Type Plants. Front. Plant. Sci. 2017, 8, 792. [Google Scholar] [CrossRef] [PubMed]
- Lawas, L.M.F.; Li, X.; Erban, A.; Kopka, J.; Jagadish, S.V.K.; Zuther, E.; Hincha, D.K. Metabolic Responses of Rice Cultivars with Different Tolerance to Combined Drought and Heat Stress under Field Conditions. GigaScience 2019, 8, giz050. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.; Guo, J. Arbuscular Mycorrhizal Fungi Improve Plant Growth of Ricinus communis by Altering Photosynthetic Properties and Increasing Pigments under Drought and Salt Stress. Ind. Crops Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Liang, S.-M.; Zheng, F.-L.; Wu, Q.-S. Elucidating the Dialogue between Arbuscular Mycorrhizal Fungi and Polyamines in Plants. World J. Microbiol. Biotechnol. 2022, 38, 159. [Google Scholar] [CrossRef]
- Majumdar, R.; Shao, L.; Minocha, R.; Long, S.; Minocha, S.C. Ornithine: The Overlooked Molecule in the Regulation of Polyamine Metabolism. Plant Cell Physiol. 2013, 54, 990–1004. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Ying-Ning, Z.; Liu, C.-Y.; Xiao, J.; Li, Y. Arbuscular Mycorrhizas Alter Root System Architecture of Citrus Tangerine through Regulating Metabolism of Endogenous Polyamines. Plant Growth Regul. 2012, 68, 27–35. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Lee, D.-J. Exogenously Applied Polyamines Increase Drought Tolerance of Rice by Improving Leaf Water Status, Photosynthesis and Membrane Properties. Acta Physiol. Plant 2009, 31, 937–945. [Google Scholar] [CrossRef]
- Ullah, A.; Ihsan, M.; Laiq, M.; Nisar, M.; Hazrat, A.; Ullah, S.I.; Ullah, S.; Ullah, A. 8–Role of Nitric Oxide in Drought Stress. In Nitric Oxide in Plant Biology; Pratap Singh, V., Singh, S., Tripathi, D.K., Romero-Puertas, M.C., Sandalio, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 197–210. ISBN 978-0-12-818797-5. [Google Scholar]
- Yamasaki, H.; Cohen, M.F. NO Signal at the Crossroads: Polyamine-Induced Nitric Oxide Synthesis in Plants? Trends Plant Sci. 2006, 11, 522–524. [Google Scholar] [CrossRef]
- Nandy, S.; Das, T.; Tudu, C.K.; Mishra, T.; Ghorai, M.; Gadekar, V.S.; Anand, U.; Kumar, M.; Behl, T.; Shaikh, N.K.; et al. Unravelling the Multi-Faceted Regulatory Role of Polyamines in Plant Biotechnology, Transgenics and Secondary Metabolomics. Appl. Microbiol. Biotechnol. 2022, 106, 905–929. [Google Scholar] [CrossRef]
- Liao, D.; Wang, S.; Cui, M.; Liu, J.; Chen, A.; Xu, G. Phytohormones Regulate the Development of Arbuscular Mycorrhizal Symbiosis. Int. J. Mol. Sci. 2018, 19, 3146. [Google Scholar] [CrossRef]
- MacLean, A.; Bravo, A.; Harrison, M. Plant Signaling and Metabolic Pathways Enabling Arbuscular Mycorrhizal Symbiosis. Plant. Cell 2017, 29, 2319–2335. [Google Scholar] [CrossRef]
- Sebastiana, M.; Duarte, B.; Monteiro, F.; Malhó, R.; Caçador, I.; Matos, A. The Leaf Lipid Composition of Ectomycorrhizal Oak Plants Shows a Drought-Tolerance Signature. Plant. Physiol. Biochem. 2019, 144, 157–165. [Google Scholar] [CrossRef]
- Sebastiana, M.; Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Monteiro, F.; Figueiredo, A.; Maia, M.; Nascimento, R.; Silva, M.S.; Ferreira, A.N.; et al. Metabolomics and Transcriptomics to Decipher Molecular Mechanisms Underlying Ectomycorrhizal Root Colonization of an Oak Tree. Sci. Rep. 2021, 11, 8576. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling Mycorrhiza-Induced Resistance. Curr. Opin Plant. Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic Acid-Induced Abiotic Stress Tolerance and Underlying Mechanisms in Plants. Front. Plant. Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed]
- Fayez, K.A.; Bazaid, S.A. Improving Drought and Salinity Tolerance in Barley by Application of Salicylic Acid and Potassium Nitrate. J. Saudi Soc. Agric. Sci. 2013, 13, 45–55. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Zamarreño, Á.M.; García-Mina, J.M.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Symbiosis and Salicylic Acid Regulate Aquaporins and Root Hydraulic Properties in Maize Plants Subjected to Drought. Agric. Water Manag. 2018, 202, 271–284. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Vlot, A.C.; Wildermuth, M.C.; Klessig, D.F. Salicylic Acid Biosynthesis and Metabolism. Arab. Book 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed]
- Vaca, E.; Behrens, C.; Theccanat, T.; Choe, J.-Y.; Dean, J.V. Mechanistic Differences in the Uptake of Salicylic Acid Glucose Conjugates by Vacuolar Membrane-Enriched Vesicles Isolated from Arabidopsis Thaliana. Physiol. Plant 2017, 161, 322–338. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Peñuelas, J. Photo- and Antioxidative Protection, and a Role for Salicylic Acid during Drought and Recovery in Field-Grown Phillyrea angustifolia Plants. Planta 2003, 217, 758–766. [Google Scholar] [CrossRef]
- Bandurska, H.; Niedziela, J.; Chadzinikolau, T. Separate and Combined Responses to Water Deficit and UV-B Radiation. Int. J. Plant Sci. 2013, 213, 98–105. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Blake, P.S.; Clay, H.A. Desiccation Stress in Recalcitrant Quercus robur L. Seeds Results in Lipid Peroxidation and Increased Synthesis of Jasmonates and Abscisic Acid. J. Exp. Bot. 1996, 47, 661–667. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and Plant Responses to Salinity Stress: A Review. Plant. Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Methyl Jasmonate-Induced Alteration in Lipid Peroxidation, Antioxidative Defence System and Yield in Soybean Under Drought. J. Agron. Crop. Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Kotrade, P.; Sehr, E.M.; Brüggemann, W. Expression Profiles of 12 Drought Responsive Genes in Two European (Deciduous) Oak Species during a Two-Year Drought Experiment with Consecutive Drought Periods. Plant. Gene 2019, 19, 100193. [Google Scholar] [CrossRef]
- Torres, N.; Goicoechea, N.; Zamarreño, A.M.; Carmen Antolín, M. Mycorrhizal Symbiosis Affects ABA Metabolism during Berry Ripening in Vitis vinifera L. Cv. Tempranillo Grown under Climate Change Scenarios. Plant. Sci. 2018, 274, 383–393. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF Inoculation and Phosphorus Supplementation Alleviates Drought Induced Growth and Photosynthetic Decline in Nicotiana Tabacum by Up-Regulating Antioxidant Metabolism and Osmolyte Accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Armada, E.; Zamarreño, A.M.; García-Mina, J.M.; Aroca, R. Local Root ABA/Cytokinin Status and Aquaporins Regulate Poplar Responses to Mild Drought Stress Independently of the Ectomycorrhizal Fungus Laccaria Bicolor. J. Exp. Bot. 2019, 70, 6437–6446. [Google Scholar] [CrossRef]
- Hill, R.A.; Wong-Bajracharya, J.; Anwar, S.; Coles, D.; Wang, M.; Lipzen, A.; Ng, V.; Grigoriev, I.V.; Martin, F.; Anderson, I.C.; et al. Abscisic Acid Supports Colonization of Eucalyptus Grandis Roots by the Mutualistic Ectomycorrhizal Fungus Pisolithus microcarpus. New Phytol. 2022, 233, 966–982. [Google Scholar] [CrossRef]
- Jin, W.; Peng, L.; Zhang, X.; Sun, H.; Yuan, Z. Effects of Endophytic and Ectomycorrhizal Basidiomycetes on Quercus virginiana Seedling Growth and Nutrient Absorption. J. Sustain. For. 2019, 38, 457–470. [Google Scholar] [CrossRef]
- Jimenez Aleman, G.H.; Thirumalaikumar, V.P.; Jander, G.; Fernie, A.R.; Skirycz, A. OPDA, More than Just a Jasmonate Precursor. Phytochem 2022, 204, 113432. [Google Scholar] [CrossRef]
- De Domenico, S.; Bonsegna, S.; Horres, R.; Pastor, V.; Taurino, M.; Poltronieri, P.; Imtiaz, M.; Kahl, G.; Flors, V.; Winter, P.; et al. Transcriptomic Analysis of Oxylipin Biosynthesis Genes and Chemical Profiling Reveal an Early Induction of Jasmonates in Chickpea Roots under Drought Stress. Plant Physiol. Biochem. 2012, 61, 115–122. [Google Scholar] [CrossRef]
- Liu, W.; Park, S.-W. 12-Oxo-Phytodienoic Acid: A Fuse and/or Switch of Plant Growth and Defense Responses? Front. Plant Sci. 2021, 12, 724079. [Google Scholar] [CrossRef]
- León Morcillo, R.J.; Ocampo, J.A.; García Garrido, J.M. Plant 9-Lox Oxylipin Metabolism in Response to Arbuscular Mycorrhiza. Plant Signal. Behav. 2012, 7, 1584–1588. [Google Scholar] [CrossRef]
- Pedranzani, H.; Rodríguez-Rivera, M.; Gutiérrez, M.; Porcel, R.; Hause, B.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Symbiosis Regulates Physiology and Performance of Digitaria Eriantha Plants Subjected to Abiotic Stresses by Modulating Antioxidant and Jasmonate Levels. Mycorrhiza 2016, 26, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Stumpe, M.; Carsjens, J.-G.; Stenzel, I.; Göbel, C.; Lang, I.; Pawlowski, K.; Hause, B.; Feussner, I. Lipid Metabolism in Arbuscular Mycorrhizal Roots of Medicago Truncatula. Phytochem 2005, 66, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Vasic, V.; Konstantinovic, B.; Orlovic, S. Application of Post-Emergence Herbicides in the Regeneration of Pedunculate Oak (Quercus robur L.) Forests. Forestry 2014, 87, 407–415. [Google Scholar] [CrossRef]
- Scaramagli, S.; Biondi, S.; Capitani, F.; Gerola, P.; Altamura, M.M.; Torrigiani, P. Polyamine Conjugate Levels and Ethylene Biosynthesis: Inverse Relationship with Vegetative Bud Formation in Tobacco Thin Layers. Physiol. Plant. 1999, 105, 366–375. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant. Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid Assay for Determination of Water Soluble Quaternary Ammonium Compounds. Plant. Soil. 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Sánchez-Bel, P.; Sanmartín, N.; Pastor, V.; Mateu, D.; Cerezo, M.; Vidal-Albalat, A.; Pastor-Fernández, J.; Pozo, M.J.; Flors, V. Mycorrhizal Tomato Plants Fine Tunes the Growth-Defence Balance upon N Depleted Root Environments. Plant. Cell Environ. 2018, 41, 406–420. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Kumaravel, S. Study on Phenolic Content, Antioxidant Activity and CHNS Elemental Analysis of Amorphophallus sylvaticus. Int. J. Agric. Life Sci. 2016, 2, 12–17. [Google Scholar]
- Kassambara, A. Pipe-Friendly Framework for Basic Statistical Tests, [R Package Rstatix, Version 0.7.0]; Free Software Foundation Inc.: Boston, MA, USA, 2021.
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kebert, M.; Kostić, S.; Stojnić, S.; Čapelja, E.; Markić, A.G.; Zorić, M.; Kesić, L.; Flors, V. A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak. Int. J. Mol. Sci. 2023, 24, 7510. https://doi.org/10.3390/ijms24087510
Kebert M, Kostić S, Stojnić S, Čapelja E, Markić AG, Zorić M, Kesić L, Flors V. A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak. International Journal of Molecular Sciences. 2023; 24(8):7510. https://doi.org/10.3390/ijms24087510
Chicago/Turabian StyleKebert, Marko, Saša Kostić, Srđan Stojnić, Eleonora Čapelja, Anđelina Gavranović Markić, Martina Zorić, Lazar Kesić, and Victor Flors. 2023. "A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak" International Journal of Molecular Sciences 24, no. 8: 7510. https://doi.org/10.3390/ijms24087510
APA StyleKebert, M., Kostić, S., Stojnić, S., Čapelja, E., Markić, A. G., Zorić, M., Kesić, L., & Flors, V. (2023). A Fine-Tuning of the Plant Hormones, Polyamines and Osmolytes by Ectomycorrhizal Fungi Enhances Drought Tolerance in Pedunculate Oak. International Journal of Molecular Sciences, 24(8), 7510. https://doi.org/10.3390/ijms24087510