
Antiepileptic Properties of Scyllo-Inositol on Pentylenetetrazol-Induced Seizures

 , , and
, , and 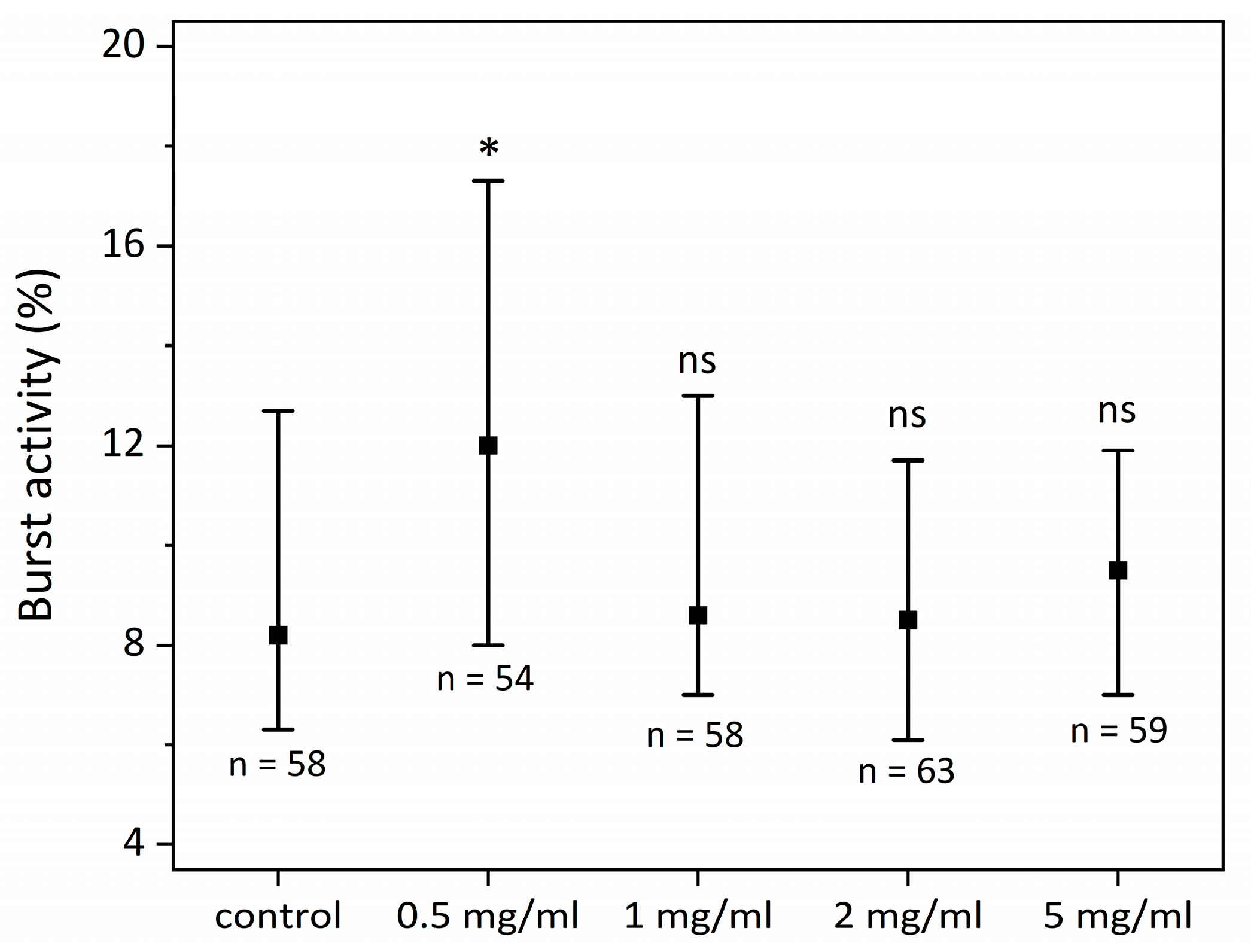{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SCI Effect on Zebrafish Larva Motility
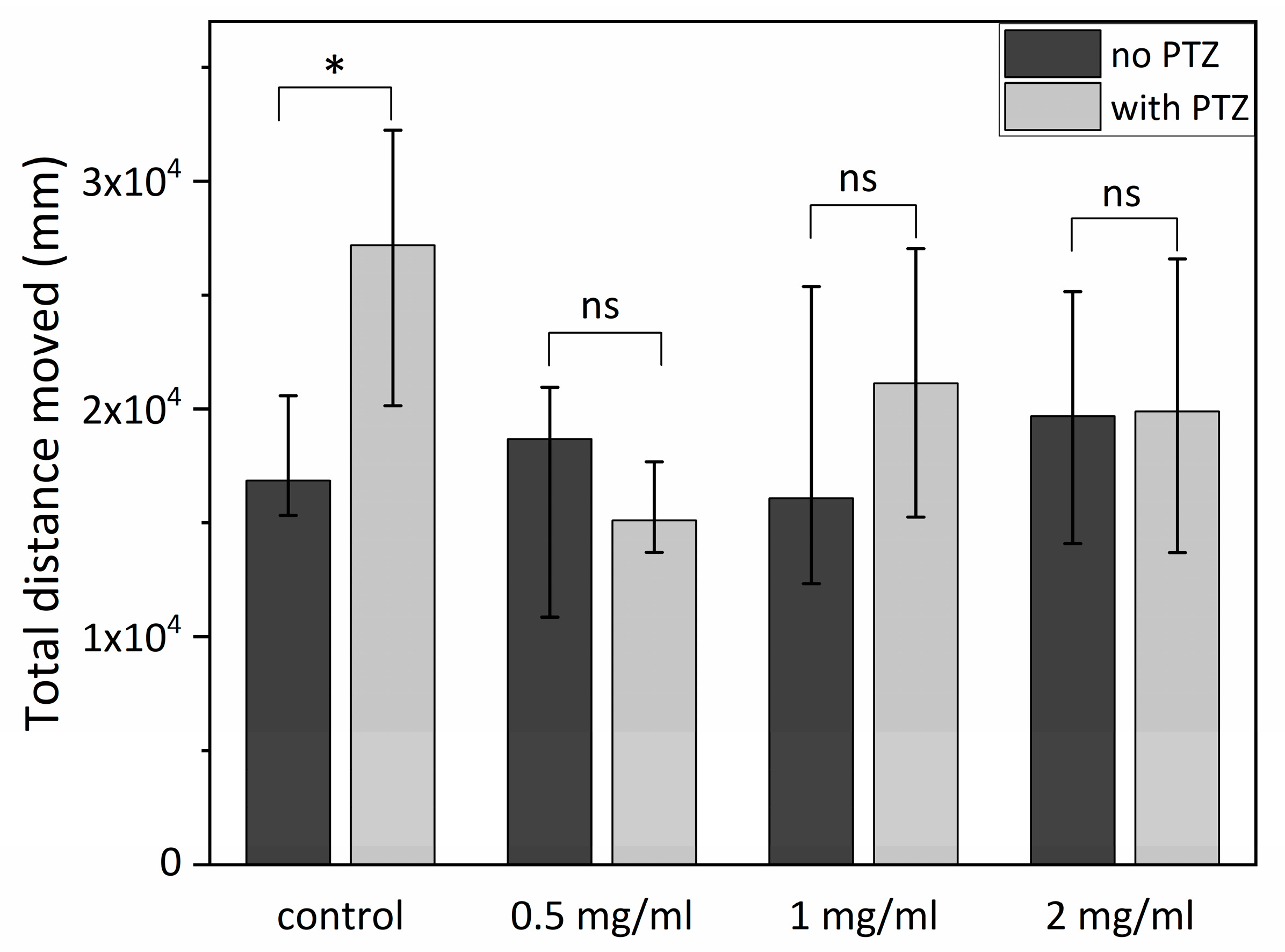
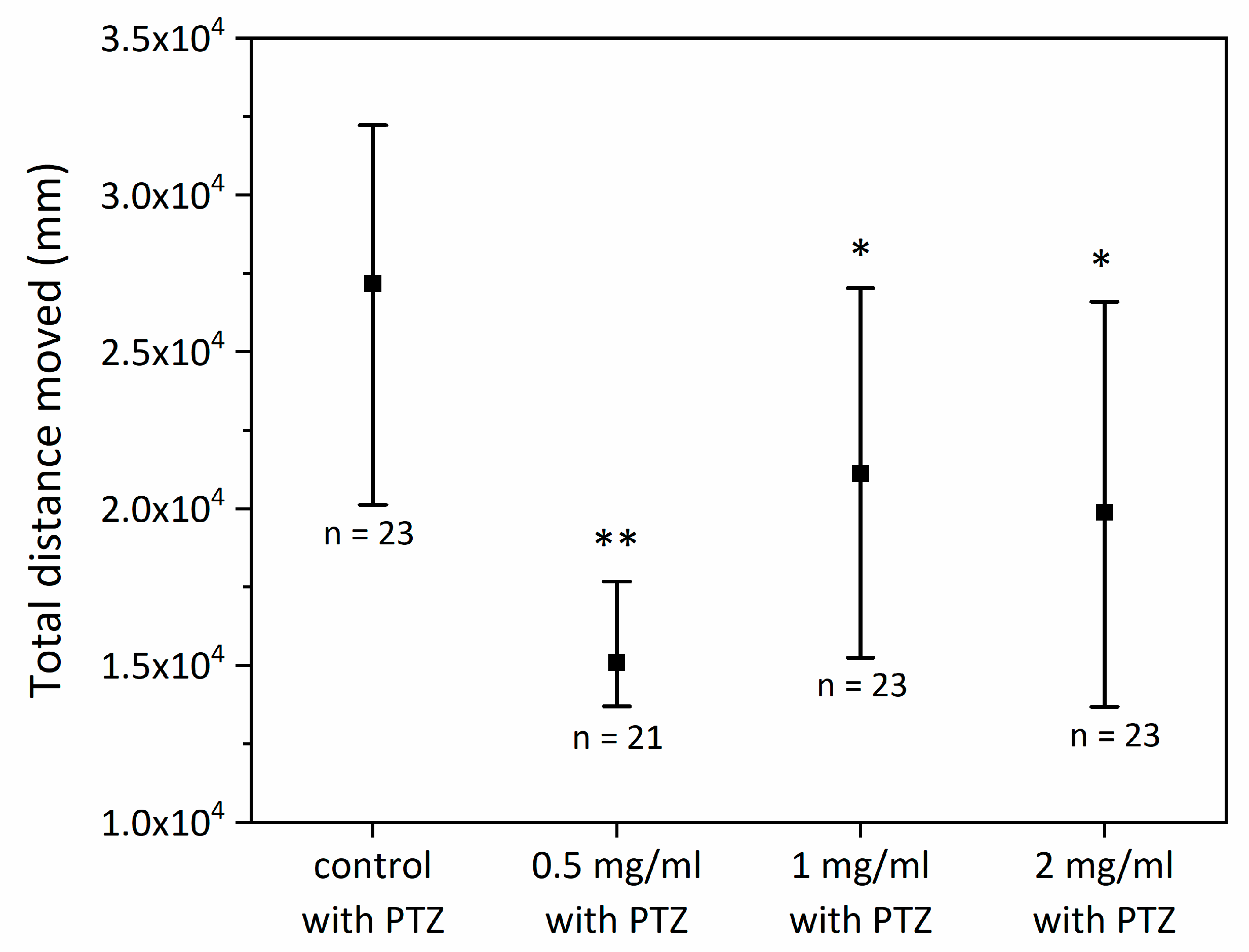
2.2. Antiepileptic Properties of SCI on Zebrafish Larvae—Short-Term Exposure to SCI
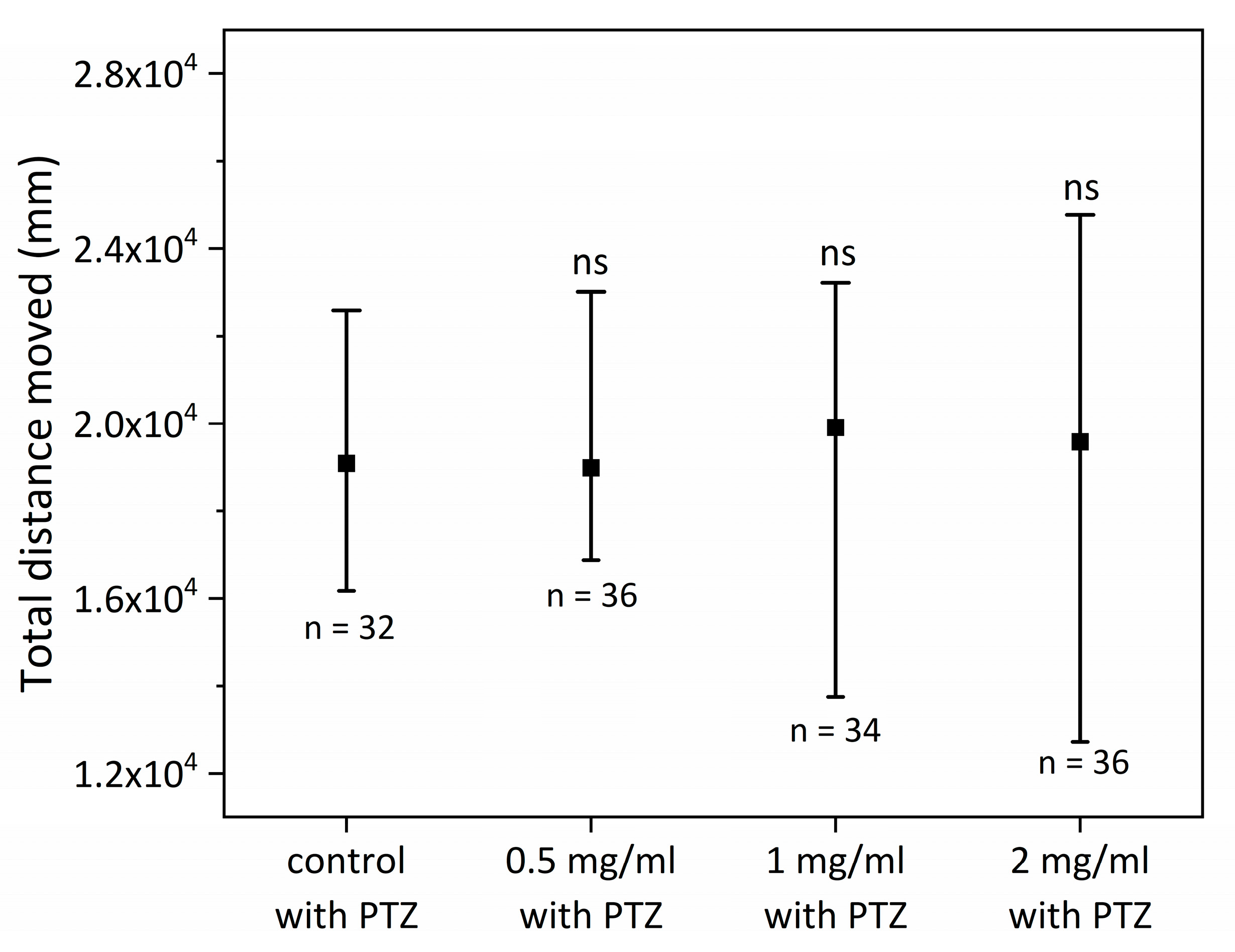
2.3. Antiepileptic Properties of SCI on Zebrafish Larvae—Long-Term Exposure to SCI
3. Discussion
4. Methods and Materials
4.1. SCI Effect on Zebrafish Larvae Activity
4.2. Antiepileptic Properties of SCI on Zebrafish Larvae—Short-Term Exposure to SCI
4.3. Antiepileptic Properties of SCI on Zebrafish Larvae—Long Exposure to SCI
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thijs, R.D.; Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Li, S. Defeating epilepsy: A global public health commitment. Epilepsia Open 2017, 2, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Brodie, M.J.; Liew, D.; Kwan, P. Treatment outcomes in patients with newly diagnosed epilepsy treated with established and new antiepileptic drugs. JAMA Neurol. 2018, 75, 279. [Google Scholar] [CrossRef] [PubMed]
- Carlomagno, G.; Unfer, V. Inositol safety: Clinical evidences. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 931–936. [Google Scholar] [PubMed]
- Antonowski, T.; Osowski, A.; Lahuta, L.; Górecki, R.; Rynkiewicz, A.; Wojtkiewicz, J. Health-promoting properties of selected cyclitols for metabolic syndrome and diabetes. Nutrients 2019, 11, 2314. [Google Scholar] [CrossRef]
- Bizzarri, M.; Fuso, A.; Dinicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Czerwińska, J.; Wygonowska, E.; Kasprowicz-Furmańczyk, M.; Placek, W. D-Chiro-inositol as a treatment in plaque psoriasis: A randomized placebo-controlled clinical trial. Dermatol. Ther. 2020, 34, e14538. [Google Scholar] [CrossRef]
- Mukai, T.; Kishi, T.; Matsuda, Y.; Iwata, N. A meta-analysis of inositol for depression and anxiety disorders. Hum.Psychopharmacol. Clin. Exp. 2013, 29, 55–63. [Google Scholar] [CrossRef]
- Levine, J. Controlled trials of inositol in psychiatry. Eur. Neuropsychopharmacol. 1997, 7, 147–155. [Google Scholar] [CrossRef]
- Genazzani, A.D. Inositol as putative integrative treatment for PCOS. Reprod. BioMedicine Online 2016, 33, 770–780. [Google Scholar] [CrossRef]
- Thomas, M.P.; Mills, S.J.; Potter, B.V. The “other” Inositols and their phosphates: Synthesis, biology, and medicine (with recent advances in myo-inositol chemistry). Angew. Chem. Int. Ed. 2015, 55, 1614–1650. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.Y.; Lan, C.P.; Hasan, S.; Brown, M.E.; McLaurin, J.A. Scyllo-inositol promotes robust mutant huntingtin protein degradation. J. Biol. Chem. 2014, 289, 3666–3676. [Google Scholar] [CrossRef] [PubMed]
- Lien, Y.-H.H.; Michaelis, T.; Moats, R.A.; Ross, B.D. Scyllo-inositol depletion in hepatic encephalopathy. Life Sci. 1994, 54, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Rafii, M.S.; Skotko, B.G.; McDonough, M.E.; Pulsifer, M.; Evans, C.; Doran, E.; Muranevici, G.; Kesslak, P.; Abushakra, S.; Lott, I.T. A randomized, double-blind, placebo-controlled, phase II study of oral ELND005 (scyllo-inositol) in young adults with down syndrome without dementia. J. Alzheimer’s Dis. 2017, 58, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Lodi, R.; Setola, E.; Tonon, C.; Ambrosetto, P.; Franceschi, E.; Crinò, L.; Barbiroli, B.; Cortelli, P. Gliomatosis Cerebri: Clinical, neurochemical and neuroradiological response to temozolomide administration. Magn. Reson. Imaging 2003, 21, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Linking genes to brain, behavior and neurological diseases: What can we learn from zebrafish? Genes Brain Behav. 2004, 3, 63–74. [Google Scholar] [CrossRef]
- Zhdanova, I.V.; Wang, S.Y.; Leclair, O.U.; Danilova, N.P. Melatonin promotes sleep-like state in zebrafish. Brain Res. 2001, 903, 263–268. [Google Scholar] [CrossRef]
- Best, J. Zebrafish: An in vivo model for the study of neurological diseases. Neuropsychiatr. Dis. Treat. 2008, 4, 567–576. [Google Scholar] [CrossRef]
- Tropepe, V.; Sive, H.L. Can zebrafish be used as a model to study the neurodevelopmental causes of autism? Genes Brain Behav. 2003, 2, 268–281. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. Connections of the ventral telencephalon (subpallium) in the zebrafish (danio rerio). Brain Res. 2004, 1011, 206–220. [Google Scholar] [CrossRef]
- Squires, R.F.; Saederup, E.; Crawley, J.N.; Skolnick, P.; Paul, S.M. Convulsant potencies of tetrazoles are highly correlated with actions on GABA/benzodiazepine/picrotoxin receptor complexes in brain. Life Sci. 1984, 35, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef]
- Afrikanova, T.; Serruys, A.-S.K.; Buenafe, O.E.; Clinckers, R.; Smolders, I.; de Witte, P.A.; Crawford, A.D.; Esguerra, C.V. Validation of the zebrafish Pentylenetetrazol seizure model: Locomotor versus electrographic responses to Antiepileptic Drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef] [PubMed]
- Berghmans, S.; Hunt, J.; Roach, A.; Goldsmith, P. Zebrafish offer the potential for a primary screen to identify a wide variety of potential anticonvulsants. Epilepsy Res. 2007, 75, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Baraban, S.C. A zebrafish-centric approach to antiepileptic drug development. Dis. Model. Mech. 2021, 14, dmm049080. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Yamagata, K. Pentylenetetrazole-Induced Kindling Mouse Model. J. Vis. Exp. 2016, 136, e56573. [Google Scholar] [CrossRef]
- Tourov, A.; Ferri, R.; Del Gracco, S.; Elia, M.; Musumeci, S.A.; Stefanini, M.C. Spike morphology in PTZ-induced generalized and cobalt-induced partial experimental epilepsy. Funct. Neurol. 1996, 11, 237–245. [Google Scholar]
- Antonowski, T.; Wiśniewski, K.; Podlasz, P.; Osowski, A.; Wojtkiewicz, J. Study of the potential hepatoprotective effect of myo-inositol and its influence on zebrafish development. Nutrients 2021, 13, 3346. [Google Scholar] [CrossRef]
- DESHUSSES, J.; REBER, G. Assymetry of the myo-inositol transport system in Klebsiella aerogenes. energy is necessary to create the Assymetry of the Transport System. Eur. J. Biochem. 1977, 72, 101–106. [Google Scholar] [CrossRef]
- Nozadze, M.; Mikautadze, E.; Lepsveridze, E.; Mikeladze, E.; Kuchiashvili, N.; Kiguradze, T.; Kikvidze, M.; Solomonia, R. Anticonvulsant activities of myo-inositol and Scyllo-inositol on pentylenetetrazol induced seizures. Seizure 2011, 20, 173–176. [Google Scholar] [CrossRef]
- Richards, M.H.; Belmaker, R.H. Epi-inositol is biochemically active in reversing lithium effects on cytidine monophosphorylphosphatidate (CMP-PA). J. Neural Transm. 1996, 103, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Groenen, P.M.W.; Merkus, H.M.W.M.; Sweep, F.C.G.J.; Wevers, R.A.; Janssen, F.S.M.; Steegers-Theunissen, R.P.M. Kinetics of myo-inositol loading in women of reproductive age. Ann. Clin. Biochem. Int. J. Lab. Med. 2003, 40, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Takenawa, T.; Tsumita, T. Properties of SCYLLITOL transport in rat kidney slices. Biochim. Biophys. Acta (BBA)-Biomembr. 1974, 373, 490–494. [Google Scholar] [CrossRef]
- Solomonia, R.; Kuchiashvili, N.; Berulava, A.; Pkhakadze, V.; Trapaidze, N.; Zhvania, M.; Abesadze, I.; Kojima, H.; Dalakishvili, N. Purification and identification of components of the Aquilegia vulgaris extract fraction exhibiting anti-epileptic activity. J. Biol. Phys. Chem. 2004, 4, 185–192. [Google Scholar]
- Strange, K.; Morrison, R.; Heilig, C.W.; DiPietro, S.; Gullans, S.R. Upregulation of inositol transport mediates inositol accumulation in hyperosmolar brain cells. Am. J. Physiol.-Cell Physiol. 1991, 260, C784–C790. [Google Scholar] [CrossRef]
- Wiese, T.J.; Dunlap, J.A.; Conner, C.E.; Grzybowski, J.A.; Lowe, W.L.; Yorek, M.A. Osmotic regulation of Na-myo-inositol cotransporter mrna level and activity in endothelial and Neural Cells. Am. J. Physiol.-Cell Physiol. 1996, 270, C990–C997. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, M.; Kohmura, E.; Yamashita, T.; Yamauchi, A.; Fujinaka, T.; Yoshimine, T.; Tohyama, M.; Hayakawa, T. Kainic acid-induced seizure upregulates na+/myo-inositol cotransporter mrna in rat brain. Mol. Brain Res. 1999, 70, 179–186. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.T.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicology Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiśniewski, K.; Antonowski, T.; Juranek, J.; Podlasz, P.; Wojtkiewicz, J. Antiepileptic Properties of Scyllo-Inositol on Pentylenetetrazol-Induced Seizures. Int. J. Mol. Sci. 2023, 24, 7598. https://doi.org/10.3390/ijms24087598
Wiśniewski K, Antonowski T, Juranek J, Podlasz P, Wojtkiewicz J. Antiepileptic Properties of Scyllo-Inositol on Pentylenetetrazol-Induced Seizures. International Journal of Molecular Sciences. 2023; 24(8):7598. https://doi.org/10.3390/ijms24087598
Chicago/Turabian StyleWiśniewski, Karol, Tomasz Antonowski, Judyta Juranek, Piotr Podlasz, and Joanna Wojtkiewicz. 2023. "Antiepileptic Properties of Scyllo-Inositol on Pentylenetetrazol-Induced Seizures" International Journal of Molecular Sciences 24, no. 8: 7598. https://doi.org/10.3390/ijms24087598
APA StyleWiśniewski, K., Antonowski, T., Juranek, J., Podlasz, P., & Wojtkiewicz, J. (2023). Antiepileptic Properties of Scyllo-Inositol on Pentylenetetrazol-Induced Seizures. International Journal of Molecular Sciences, 24(8), 7598. https://doi.org/10.3390/ijms24087598