1. Introduction
Rice (
Oryza sativa L.) is one of the top three staple crops worldwide, which feeds over half of the global population. Like yield, grain quality is one of the most concerning key agronomic traits for rice consumers, sellers, processors, producers, and breeders, which largely determines the taste, nutritional value, and price of rice. The rice grain quality is mainly composed of milling quality, appearance quality, cooking and sensory quality, and nutrition quality [
1], which is comprehensively controlled by a series of key regulatory genes and environmental factors. Thus, mapping and cloning key genes regulating grain quality from important rice germplasm resources, studying and uncovering the corresponding regulatory mechanisms, and subsequent application of these genes in breeding for rice grain quality are indispensable and helpful for scientists and breeders to develop superior-quality rice varieties.
Some transcription factors, such as MADS-box family members known to regulate inflorescence and spikelet development in rice, are also thought to be related to grain quality regulation, but the detailed underlying regulatory mechanisms are still to be explored. It is interesting that some MADS-box genes such as
OsMADS1,
OsMADS6, and
OsMADS7 are specifically and highly expressed both in panicles and seeds [
2], suggesting these genes could also be related to the regulation of seed development and grain quality formation. This deduction was partly confirmed by the abnormal phenotypes in
OsMADS6 mutant with both abnormal floral organs and shriveled seeds in which the grain quality, starch filling, and expressions of ADP-glucose pyrophosphorylase genes were decreased, but the seed protein content was increased [
3]. In addition, another MADS-box gene,
OsMADS29, was found to be specifically expressed in seeds, including nucellus, dorsal-vascular trace, embryo, and endosperm. Os
MADS29-RNAi (Os
MADS29-RNA interference) plants exhibited suppressed starch biosynthesis and reduced grain-filling rate, loosely packed starch granules, and shrunken grains. Expression of
OsMADS29 is induced by auxin, and OsMADS29 transcription factor regulates degradation of maternal tissues, including ovule, by directly binding to the downstream programmed cell death genes in these maternal tissues, thereby controlling subsequent seed development and affecting the grain quality formation [
4]. In addition, Zhang et al. [
5] reported that high temperature at the early filling stage greatly induced the expression of
OsMADS7 and suppression of it in rice endosperm stabilized amylose content, possibly by maintaining a relatively low filling rate and high expression of the encoding gene of GBSS I (granule-bound starch synthase I) under high-temperature stress.
Furthermore, extensive studies have revealed that another class E floral homeotic gene of the MADS-box family,
OsMADS1, plays important roles in regulating floral organ development in rice by regulating the floral meristem specification and floral organ identity [
6,
7,
8,
9]. Several recent studies further found out that OsMADS1 also controls grain shape by interactions with G-protein (guanine nucleotide-binding protein) subunits, including DEP1 (dense and erect panicle 1) and GS3 (grain size 3) to collaboratively regulate expressions of underlying downstream target genes related to grain shape and development [
10,
11,
12,
13].
After the first
OsMADS1 mutant,
lhs1 (
leafy hull sterile1) was reported by Kinoshita et al. [
14], at least 11 mutants and four NILs (Near-isogenic lines) of
OsMADS1 including
lhs1 [
7,
14],
nsr (
naked seed rice) [
8],
NF1019,
ND2920,
NE3043 and
NG778 [
6],
osmads1-z [
9],
afo (
abnormal floral organs) [
15],
ohms1 (
open hull and male sterile 1) [
16],
cy15 [
17] and Olr (Oat-like rice) [
10], NIL (SLG) [
13], WYJ7 (Wuyungeng No. 7)
-lgy3-dep1–1, RD23
-lgy3-gs3 and PA64S (Peiai 64S)/9311 (Yangdao No. 6)
-lgy3-gs3 [
11] have been reported so far. Thereinto, Olr is a spontaneous and severe
OsMADS1 mutant, which was named for its unique grain shape, which highly resembles oat grains. Olr displayed abnormal floral organs, open hulls formed by remarkably elongated leafy lemmas and paleae, occasionally formed conjugated twin brown rice, aberrant grain shape, low seed setting rate, slow grain-filling rate, low 1000-brown rice weight, and extremely low yield [
10].
However, it’s interesting that the grain quality was only analyzed in NIL (SLG) [
13], WYJ7
-lgy3-dep1–1, RD23
-lgy3-gs3, PA64S/9311
-lgy3-gs3 [
11], which may be partly due to unavailable seeds caused by sterility or extremely low fertility of these mutants. These four rice NILs carry the same mutated allele of
OsMADS1,
OsMADS1lgy3/OsLG3bSLG, but the effect of this allele on the grain quality of these NILs was divided into two distinct categories. There is no significant difference in chalkiness between grains of NIL (SLG) and its receptor parent Nipponbare, which indicates that
OsLG3bSLG did not affect grain quality in Nipponbare [
13]. However, WYJ7
-lgy3-dep1–1, RD23
-lgy3-gs3, and PA64S/9311
-lgy3-gs3 grains all displayed lower chalkiness compared with their respective receptor parent, WYJ7, RD23, and LYPJ (Liangyoupeijiu, PA64S/9311), which indicates that
OsMADS1lgy3 affected and improved grain quality in WYJ7, RD23, and LYPJ [
11]. Interestingly, the question of whether
OsMADS1 regulates grain quality is still contradictory and obscure.
Thus, it is necessary to analyze the grain quality of more mutants or accessions carrying different mild, moderate, or severe OsMADS1 alleles to confirm whether OsMADS1 regulates grain quality and further uncover the regulatory mechanism. In this study, we analyzed the grain quality of the OsMADS1Olr mutant (Olr) and identified a correlation between the mutation in the OsMADS1 gene and the chalkiness phenotype. The Olr mutant displays poor grain quality with chalky endosperms, abnormal morphology, and loose arrangement of starch granules, with lower starch content but higher protein content in grains. Moreover, OsMADS1 has a crucial role in regulating rice quality by coordinating gene expressions and regulatory networks of starch and storage protein metabolisms in rice grains. Therefore, this study also highlighted the application potentials of OsMADS1 and the target gene seed-specific-RNAi system in rice breeding to develop superior-quality rice varieties.
3. Discussion
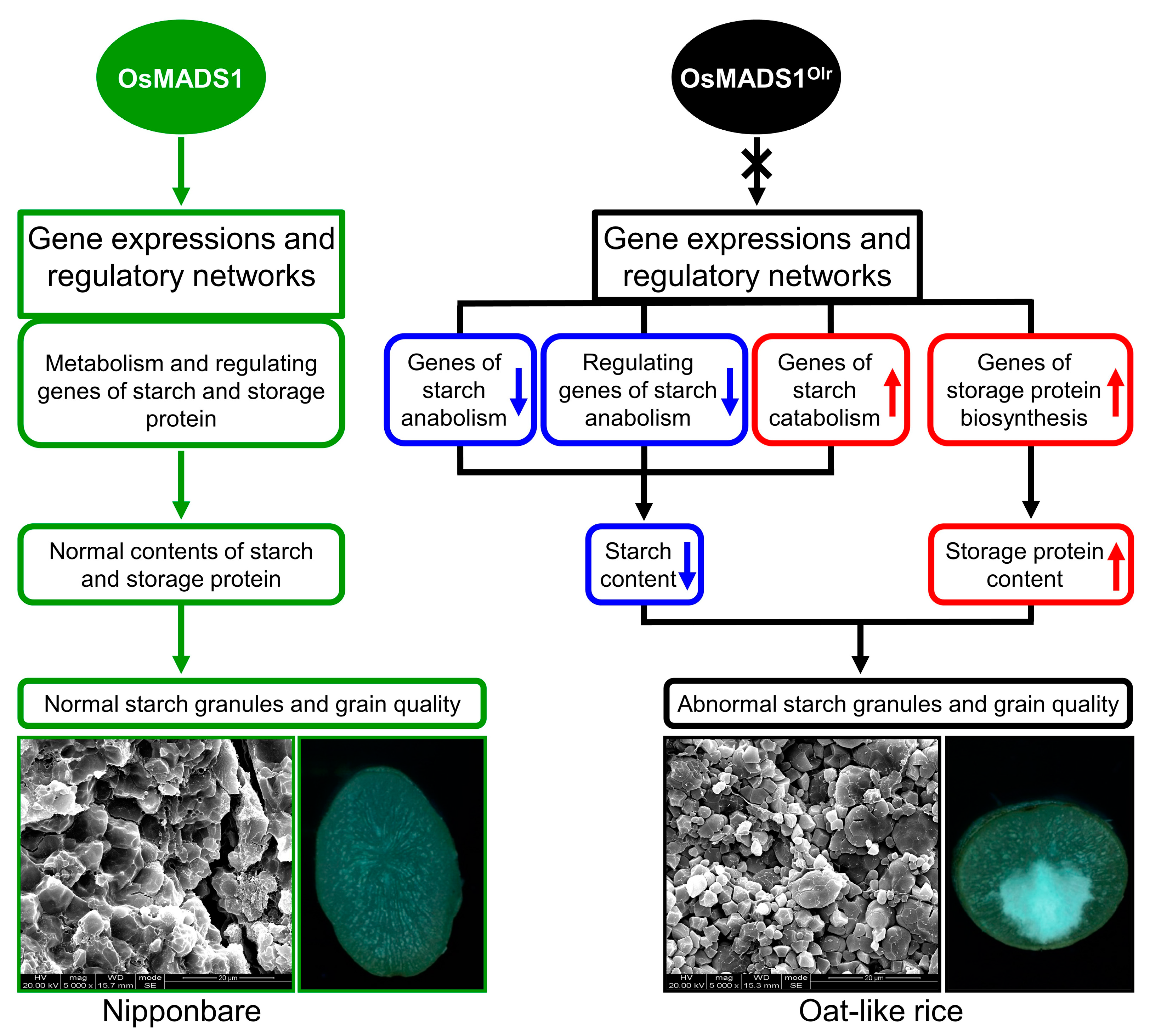
In this study, we preliminarily reveal that
OsMADS1 regulates grain quality, gene expressions, and regulatory networks of starch and storage protein metabolisms in rice (
Figure 9). However, it is very interesting that the functions of
OsMADS1 in regulating grain quality are contradictory and obscure in previous studies. The report of Liu et al. [
11] indicates that wild-type
OsMADS1 negatively regulates grain quality in WYJ7, RD23, and LYPJ. By contrast, the report of Yu et al. [
13] indicates that
OsMADS1 does not affect grain quality. On the other hand, our results in this study showed that
OsMADS1 positively regulates grain quality (
Figure 1 and
Figure 3).
However, it is very interesting that the regulatory role of
OsMADS1 in grain quality is seemingly contrary between our results (
Figure 1 and
Figure 3) and the report of Liu et al. [
11]. Firstly, these seemingly contradictory results may be partly derived from the differences in mutation types and resulted in effects on the gene functions between mutated
OsMADS1lgy3 and
OsMADS1Olr alleles. The OsMADS1 protein contains a MADS-box domain located in the N-terminal region of the protein and is responsible for the DNA binding with target genes, I region (Intervening region) and K-box domain (Keratin-like domain) mainly involved in dimerization and protein-protein interactions, and the C-terminal region implicated in transcriptional activation and higher-order complex formation [
2]. The mutated
OsMADS1lgy3/
OsLG3bSLG allele encodes an alternatively spliced mutated protein OsMADS1
lgy3, in which the terminal 37 residues were truncated, and an additional 5 residues were added to its predicted C domain [
11,
12,
13]. This mutation in the C-terminal region of OsMADS1
lgy3 did not affect the interactions with its interacting proteins, such as DEP1, and the DNA-binding affinity to its target genes, such as
OsMADS55 and
OsPIN1a, but largely abolished the transactivation activity to its target genes such as
OsMADS55 and
OsPIN1a [
11], which could thereby change and affect the regulatory functions of OsMADS1
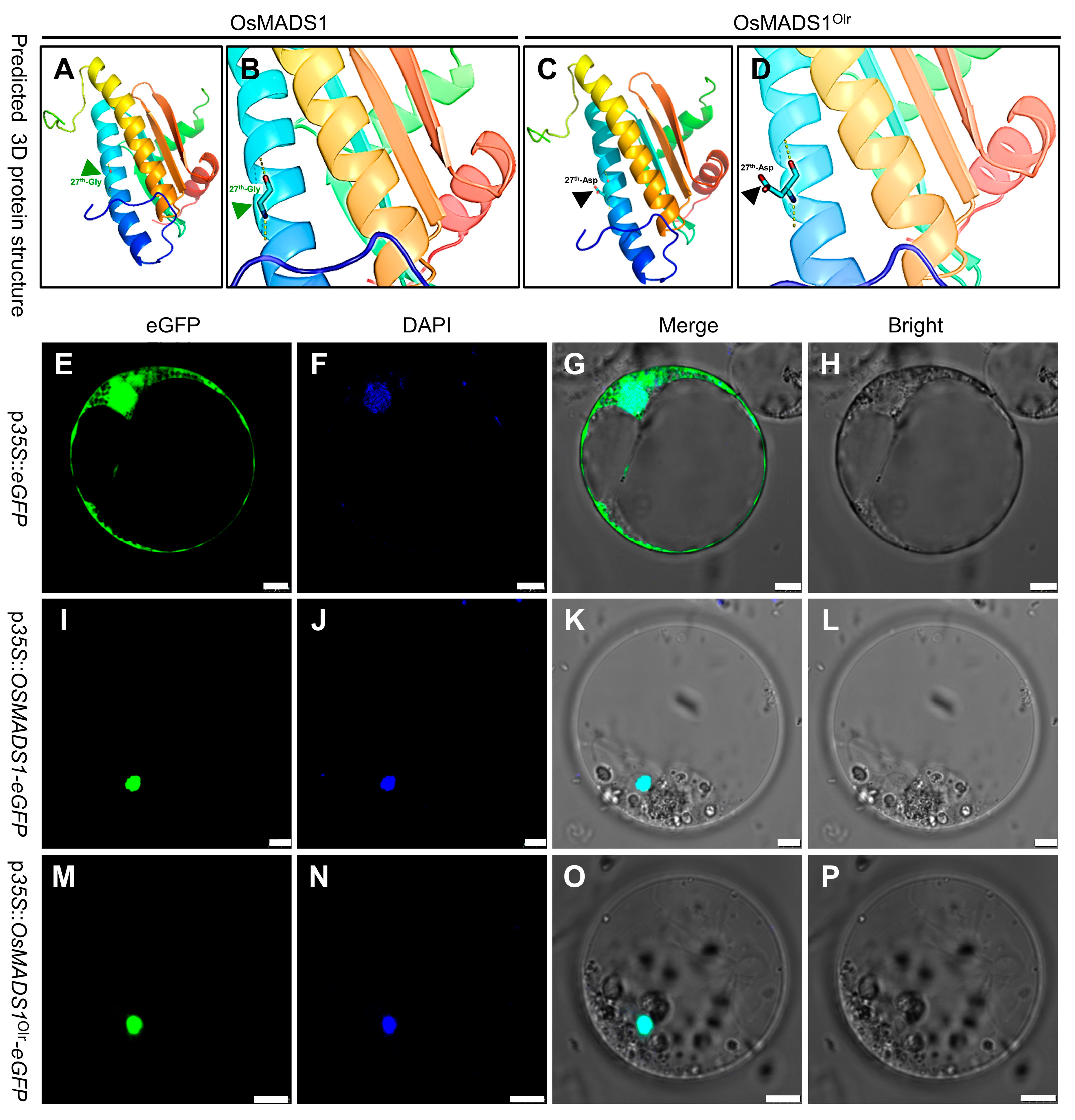
lgy3 to grain development and quality. On the other hand, the I region, K-box domain, and C-terminal region were normal, but the 27th conserved glycine in the MADS-box domain was mutated to aspartic acid in OsMADS1
Olr protein (
Figure 4D), which could affect its 3D protein structure (
Figure 5A–D) and the DNA-binding affinity to its target genes and thereby change and affect the regulatory functions of OsMADS1
Olr to spikelet and grain development as well as grain quality. Thus, the differences in mutation types and resulted in effects on the gene functions between mutated
OsMADS1lgy3 and
OsMADS1Olr alleles may partly cause the differences in grain quality between the three NILs of
OsMADS1lgy3 (WYJ7
-lgy3-dep1–1, RD23
-lgy3-gs3 and PA64S/9311
-lgy3-gs3) and Olr mutant harboring the
OsMADS1Olr.
In the regulation processes of grain quality by OsMADS1lgy3 or OsMADS1Olr allele, it needs other cofactors or interacting proteins to form the transcription complex to regulate expressions of a large number of downstream grain quality-related target genes. The OsMADS1lgy3 and OsMADS1Olr are individually carried by the three NILs and Olr with different genetic backgrounds. Therefore, it is likely that OsMADS1lgy3 and OsMADS1Olr have some different cofactors, interacting proteins, and downstream target genes, and some genes encoding these cofactors and interacting proteins may also have different spatial and temporal expression patterns in different backgrounds. Therefore, the differences in components and their contents between OsMADS1lgy3 and OsMADS1Olr transcription complex in different backgrounds could result in the different effects of gene functions in regulating grain quality between OsMADS1lgy3 and OsMADS1Olr alleles.
The three NILs of
OsMADS1lgy3 displayed improved grain quality accompanied by normal spikelet fertility, longer grain length, and increased yield. On the other hand, the Olr mutant harboring
OsMADS1Olr exhibited decreased grain quality and the abnormal grain shape. Furthermore,
OsMADS1lgy3 is a useful allele in rice breeding, but
OsMADS1Olr is not likely to be used in rice breeding directly. These results revealed that compared with the mild
OsMADS1lgy3 allele,
OsMADS1Olr is a more severe allele. In addition, this study and previous results indicate that
OsMADS1 plays critical and dual roles in regulating grain quality and grain shape [
10,
11,
12,
13].
Moreover, it also plays balanced regulatory roles in controlling grain shape, which regulates the specification of the lemma and palea and simultaneously acts as an inhibitor of overdevelopment of the lemma, palea, and grain length to maintain the balanced development of rice grains. Overexpression of wild-type
OsMADS1 in Nip resulted in decreased grain length and smaller grain size but suppression of
OsMADS1 expression in Nip caused increased grain length and bigger grain size [
11]. The dual and balanced regulatory roles of
OsMADS1 may also exist in the regulation of both grain quality and grain shape in rice and could be partly realized by coordinating the regulation of grain quality and grain shape. Li et al. [
19] reported that the appearance quality attribute, grain chalkiness, is positively correlated with the grain shape attribute, grain width. Therefore, the compromise in gene function in the mild mutated allele
OsMADS1lgy3 may result in the re-establishment and maintenance of a better balance and homeostasis to regulate grain quality and grain shape, which finally leads to the improvement of grain quality and longer grain length in WYJ7
-lgy3-dep1–1, RD23
-lgy3-gs3 and PA64S/9311
-lgy3-gs3 NILs. Conversely, a serious impact on gene function in the severely mutated allele
OsMADS1Olr may damage the normal regulatory roles of
OsMADS1 and break down the balance and homeostasis to regulate grain quality and grain shape, which finally causes the poor grain quality and abnormal grain shape in Olr.
In conclusion, the opposite and seemingly contradictory effects between OsMADS1lgy3 and OsMADS1Olr alleles on the grain quality in NIL (SLG), WYJ7-lgy3-dep1–1, RD23-lgy3-gs3, PA64S/9311-lgy3-gs3 and Olr can be partly explained and may be caused by the differences of mutation types and resulted in effects on the gene functions of OsMADS1 alleles, influences of different genetic backgrounds to the gene functions of OsMADS1 alleles, dual and balanced regulatory roles of OsMADS1 alleles in regulating grain quality and the grain shape.
Our results showed that
OsMADS1 controls grain quality by regulating starch and seed storage protein metabolism in rice. However, the detailed molecular regulatory mechanism still remains to be explored. Bello et al. [
20] reported that two NF-Y type transcription factors, OsNF-YB1 and NF-YC12 (nuclear transcription factor Y subunit C-12), bind to each other and sequentially bind to another Helix-loop-helix transcription factor bHLH144 (basic helix-loop-helix protein 144) to form a heterotrimer complex NF-YB1-YC12-bHLH144, in which NF-YC12 and bHLH144 maintain NF-YB1 stability from the degradation mediated by ubiquitin/26S proteasome, while NF-YB1 directly binds to the G-box (CACGTG) domain of
Wx (
Waxy, granule-bound starch synthase I gene) promoter and activates
Wx transcription, hence to regulate rice grain quality. There is also the possibility that OsMADS1 and other transcription factors can form heterodimerization and transcription complex to jointly regulate transcriptions of some downstream grain quality-related target genes to regulate grain quality. Our results showed that concurrently and overall decreased expressions of
OsMADS1,
OsNF-YB1,
OsNF-YC12,
bHLH144, and starch biosynthesizing genes in the developing grains of Olr is consistent with the reduced starch content in Olr grains, which partly supports this possibility.
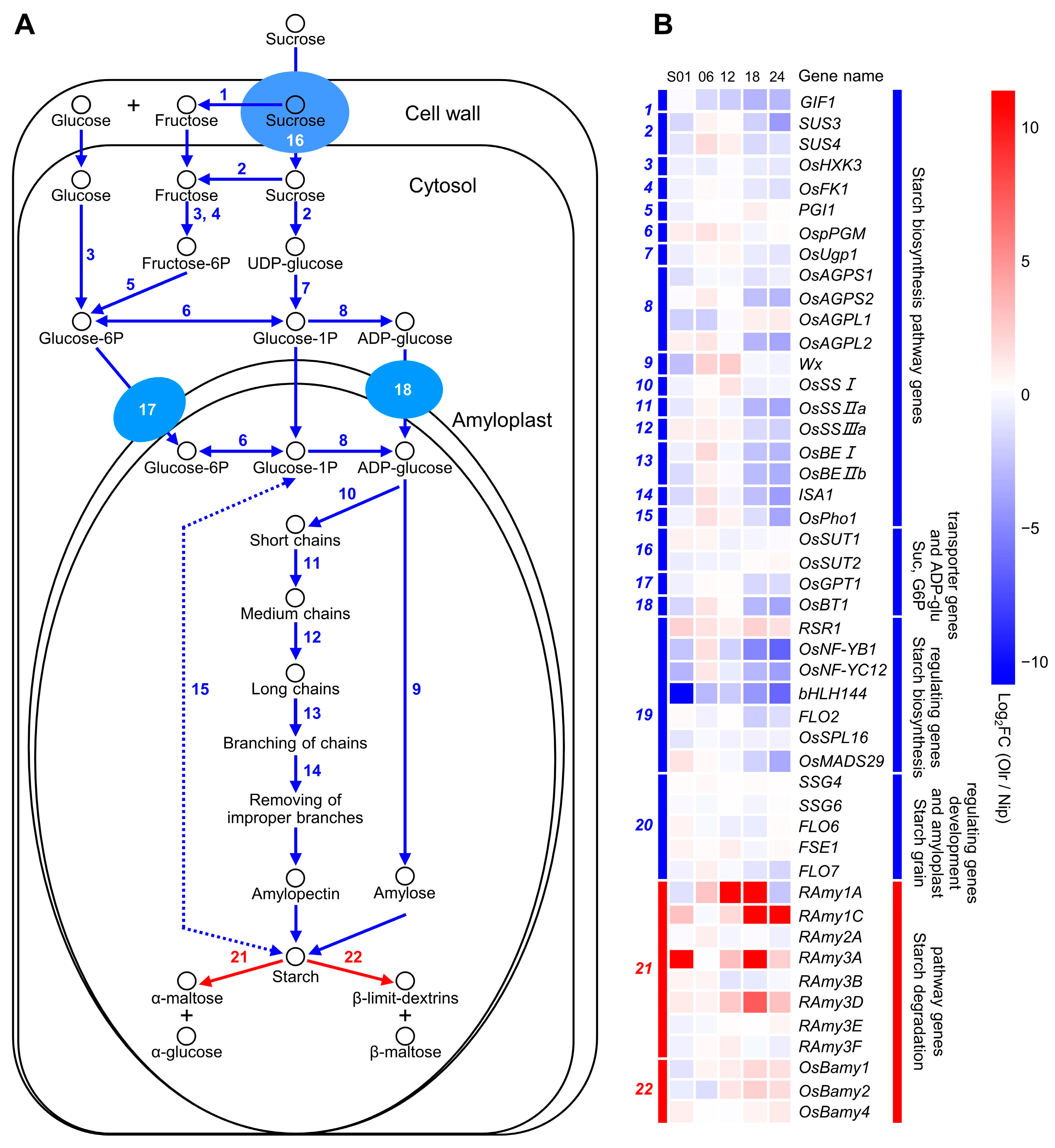
Furthermore, in contrast to the positive regulators of starch biosynthesis, OsNF-YB1, OsNF-YC12, and bHLH144, another transcription factor RSR1 was reported to be a negative regulator of starch biosynthesis, and its expressions were obviously reduced in developing grains of Olr [
21] (
Figure 6). These results suggest that OsMADS1 may have the dual regulatory roles of activating or/and suppressing expressions of different starch and seed storage protein-related genes in grains by binding or interacting with some of the positive or/and negative regulators such as OsNF-YB1, OsNF-YC12, bHLH144, and RSR1 to accurately regulate starch and seed storage protein biosynthesis, hereafter accurately controlling grain quality. Our results showed that in developing grains of Olr, expressions of starch biosynthesizing and regulating genes were suppressed, but expressions of starch-degrading genes were promoted as a whole, and the seed storage protein biosynthesis regulating genes were suppressed at 1 DAF, 18 DAF, and 24 DAF but promoted at 6 DAF and 12 DAF in general (
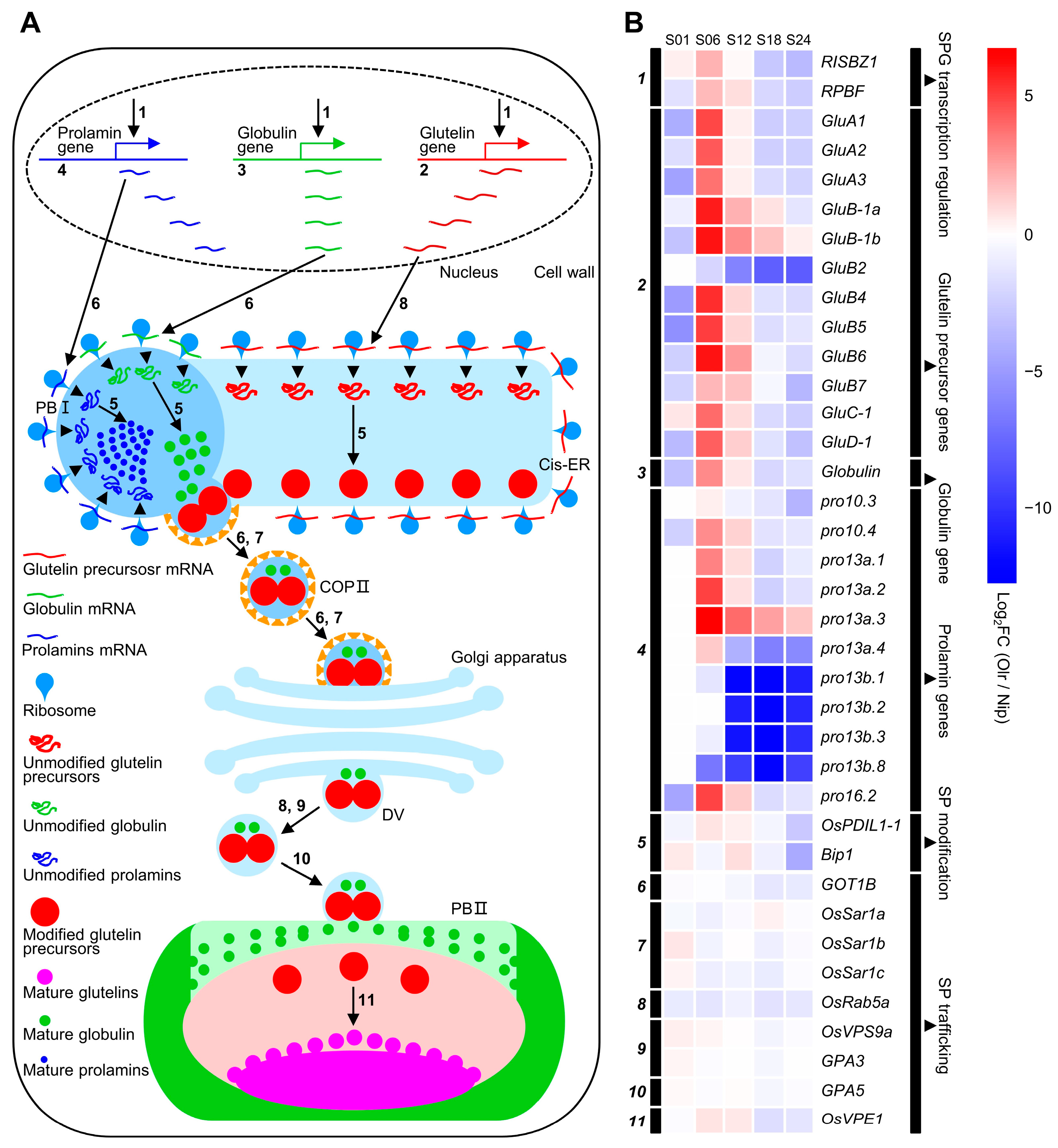
Figure 6 and
Figure 7), which is consistent with this deduction.
On the other hand, RISBZ1 and RPBF encode a rice basic leucine zipper transcription factor and a rice prolamin box binding factor, respectively, which not only interact with each other to regulate expressions of seed storage protein genes through GCN4 Motif and thereby regulate storage protein biosynthesis but also regulate expressions of some of the starch biosynthesizing genes such as
Wx [
22,
23]. Thus, similar to RISBZ1 and RPBF, there is also the possibility that OsMADS1 can regulate expressions of both starch, seed storage protein metabolizing, and related genes to collaboratively regulate starch and seed storage protein metabolism, thereby controlling grain quality.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
The ‘Olr (Oat-like rice)’ is a stably hereditable and recessive mutant caused by a spontaneous mutation, which was originally discovered in the paddy field in 2001 [
10]. The
japonica cv. Nip (Nipponbare) was used as a control of Olr in the comparative phenotypic and transcriptomic analyses during grain development because its original wild type is unknown. Nip was also used as the transgenic recipient and wild-type control in the
OsMADS1 seed-specific RNAi (p
OsTip3::
OsMADS1-RNAi) transgenic experiment. The detailed method for constructing the p
OsTip3::
OsMADS1-RNAi transgenic lines was described in the study of Li et al. [
10]. Olr, Nip, the F
2 and F
3 progeny plants derived from a cross combination of Olr/Nip were used for the determination of starch and protein contents in grains. Field-grown rice plants were grown under normal paddy conditions during natural growing seasons in an isolated experimental plot in Chengdu city, Sichuan province of China.
4.2. Measurement of Starch, Amylose, Amylopectin, and Protein Content
Brown rice from Nip, Olr, F
2, and F
3 progeny plants of Olr and Nip were milled into fine flour by using a flour mill. The fine flour from each of the samples was subsequently used for the determination of the starch, amylose, amylopectin, and protein contents. Starch content, amylose content, and amylopectin contents were determined by using the starch assay kits Megazyme K-TSTA and K-AMYL (Megazyme, Wicklow, Ireland) as described by Wei et al. [
24]. The protein content of the samples was determined following the method described by Kang et al. [
25].
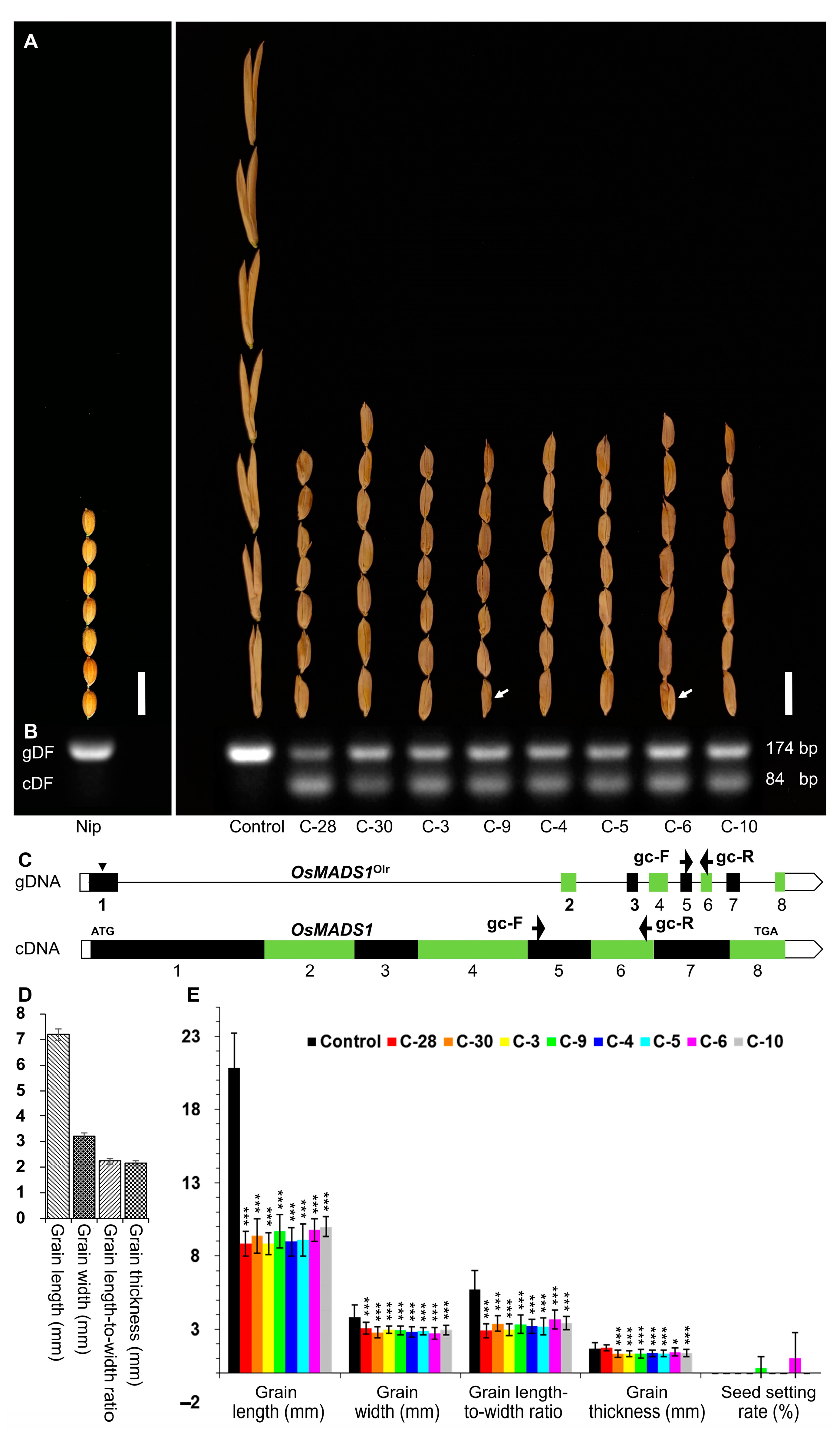
4.3. Vector Construction and Rice Transformation
For the construction of the p
Ubi::
OsMADS1 vector used to express the wild-type
OsMADS1 gene in Olr to rescue its poor grain quality, the wild-type
OsMADS1 cDNA was cloned from Nip and was inserted into the binary vector pCUbi1390 [
26] at the BamH I site and the Spe I site. Then, the p
Ubi::
OsMADS1 vector was transformed into the Olr calli to obtain T
0 transgenic plants by
Agrobacterium tumefaciens (EHA105)-mediated transformation [
27]. The corresponding primers developed for constructing the p
Ubi::
OsMADS1 vector are listed in
Table S8.
4.4. Transcriptome Samples Collection and Treatment
Nip and Olr grains at representative time points during grain development consisting of 1 DAF, 6 DAF, 12 DAF, 18 DAF, and 24 DAF were collected from field-grown rice plants. For all grain samples at each time point, three biological replicates were harvested from Nip and Olr plants. Harvested samples were immediately frozen in liquid nitrogen and stored at −80 °C. Then all samples frozen by drikold were shipped to Basebio Biotechnology Co., Ltd. (Chengdu 610041, China) for sample quality testing, cDNA library construction, transcriptome sequencing, and initial data analysis.
4.5. Transcriptome Data Analysis
The sequencing quality of obtained raw data with the fastq format of each sample was assessed using the FastQC tool (
https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 27 April 2023). Subsequently, the clean reads of each sample with high quality were aligned to the rice reference genome sequence (Nipponbare rice genome IRGSP-1.0) using the Tophat2 software (v2.1.1, Center for Bioinformatics and Computational Biology, University of Maryland, College Park, MD, USA) [
28]. For transcript quantification, HTSeq-count script (v2.0.2,
https://pypi.org/project/HTSeq/, accessed on 27 April 2023) was then used to obtain the calculated reads count and FPKM (fragments per kilobase of transcript per million mapped reads) value for every single transcript of every gene in each sample. To test the correlation of different samples and the similarity of biological replicates, the PCA (Principal Component Analysis) and sample-to-sample correlation analysis were used. DEGs testing was analyzed using DESeq R packages according to the manual. Raw count data was prepared by custom perl script based on the results of eXpress software and was imported into the DESeq framework. The FDRs (False Discovery Rates) were controlled using the Benjamini–Hochberg method at an FDR of 5% [
29]. For analyzing the overlapping DEGs, Venn diagram analysis was employed [
30]. Functional enrichment analysis of DEGs is based on the hypergeometric test. The obtained DEGs between Olr and Nip grain samples were annotated by the databases of GO and KEGG and subsequently subjected to GO and KEGG significant enrichment analysis to identify starch and storage protein-related GO functional categories and KEGG pathways [
31,
32]. SOMs were constructed to identify and compare the expression patterns of DEGs between Olr and Nip grains by using SOM R packages. Subsequently, temporal expression profiles of the obtained robust clusters were constructed based on the extracted characteristic values of the gene expression levels in the robust clustering maps by the SOMs analysis [
33].
4.6. Grain Appearance Quality Analysis and Microscope Observation
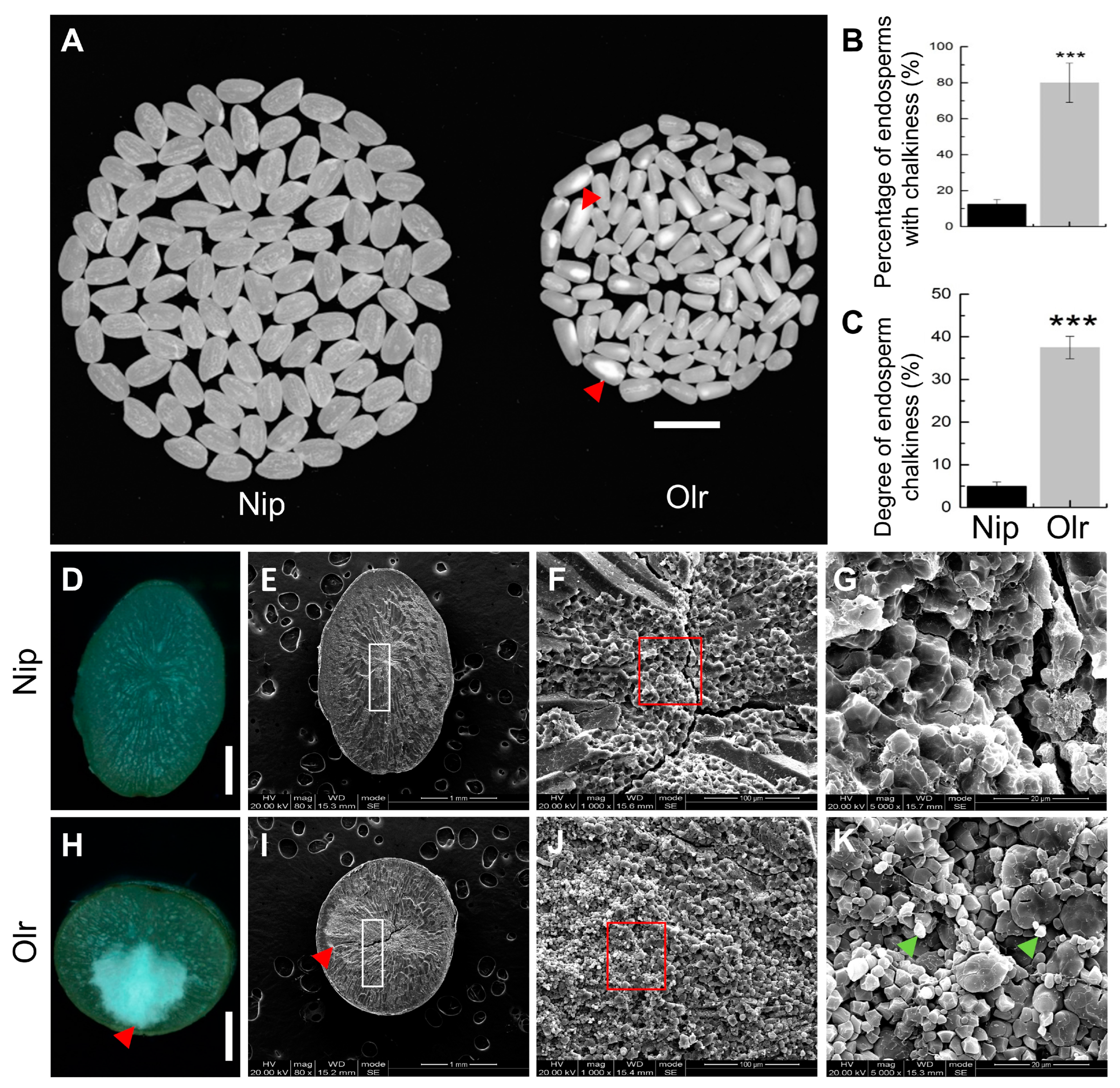
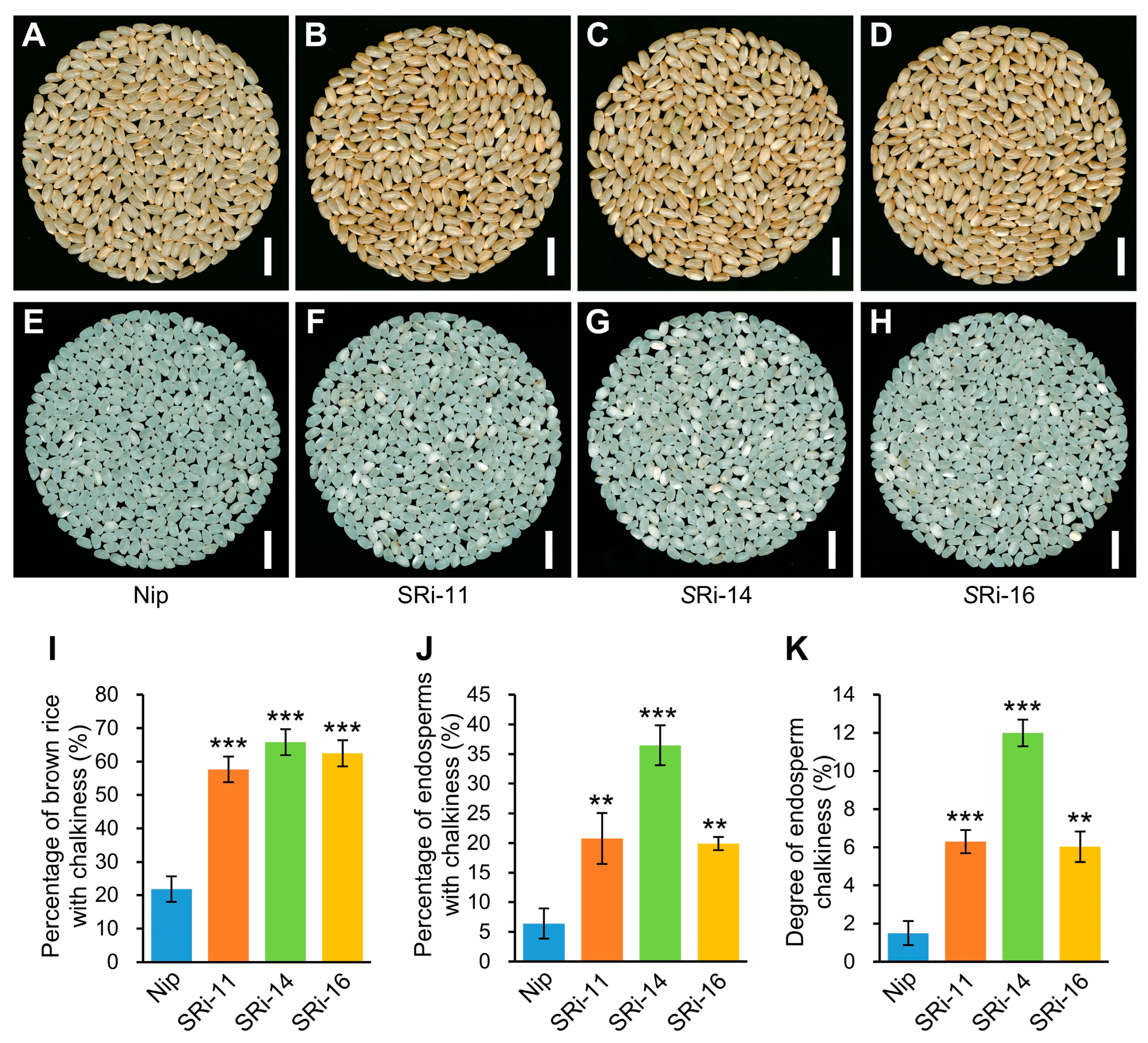
Mature grains were harvested and naturally dried, then stored at room temperature. Grains of Nip and Olr were dehulled using a rice huller or manually. Then brown rice of Nip and Olr was ground into white rice (endosperms) using a rice polisher. The appearance quality of these white rice, including the percentage of endosperms with chalkiness and degree of endosperm chalkiness, was analyzed using a rice quality analyzer (TPMZ-A, Zhejiang Top Cloud-agri Technology Co., Ltd., Hangzhou, China).
Brown rice grains of Nip and Olr were transversely cut in the middle using a dissecting blade. Then the corresponding half-brown rice grains were mounted on stubs, and the transverse sections of these samples were first observed under a dissecting microscope (SMZ745T, Nikon, Tokyo, Japan). Then each of the same observed samples was mounted on a stub, and the transverse section was coated with gold and subsequently observed under an SEM (Inspect S50, FEI Company, Hillsboro, OR, USA) with an accelerating voltage of 20.00 kV. The transverse sections of the brown rice grains with or without chalkiness and the corresponding starch granules were observed and photographed under the SEM.
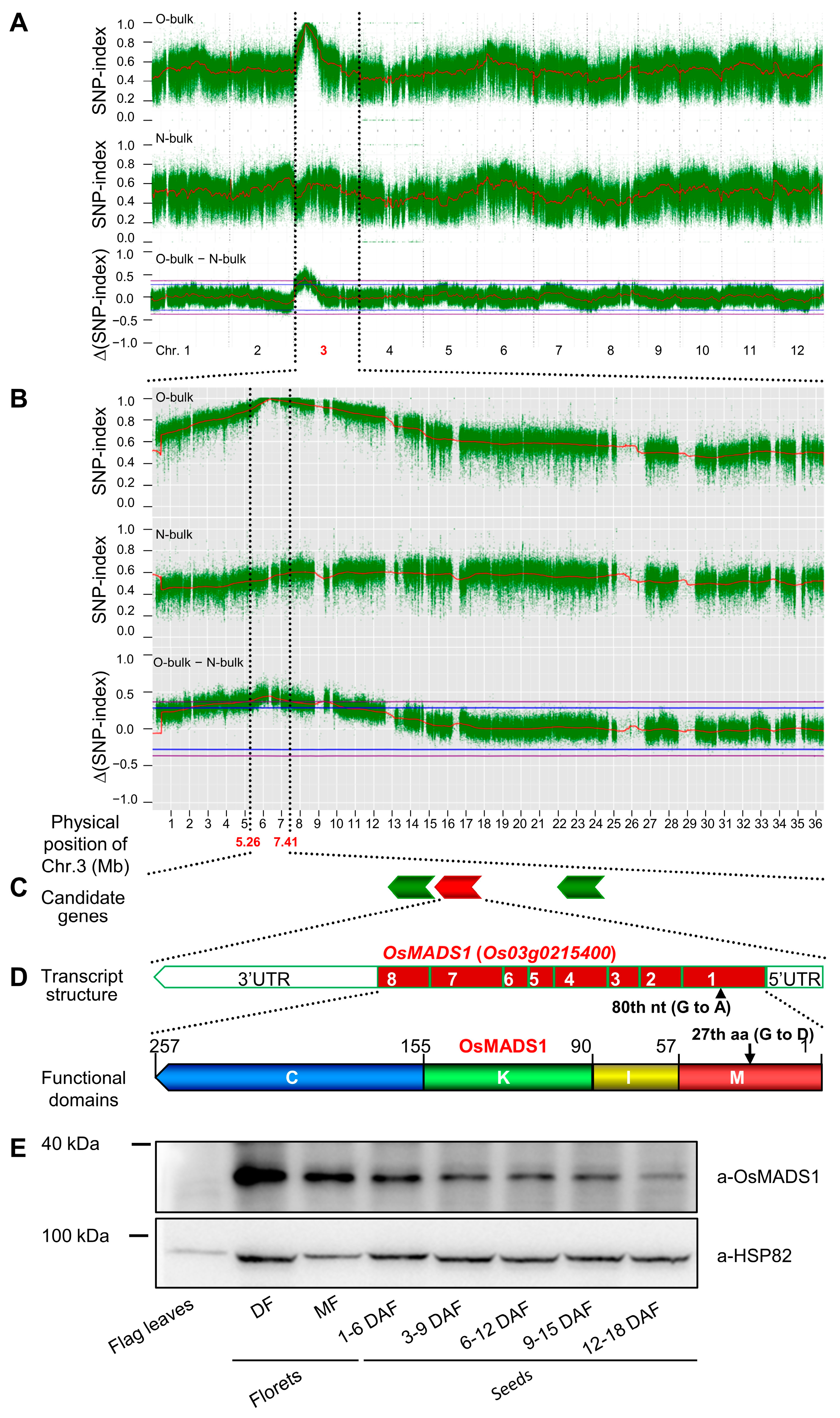
4.7. Mapping-by-Sequencing the Causal Gene of Grain Quality Phenotype in Olr
For mapping and cloning the causal gene through the mapping-by-sequencing method, we initially constructed an F
2 segregating population by a cross between Olr (maternal plant) and Nip (paternal plant) [
10]. Then, the maternal bulk DNA (OM-bulk) and paternal bulk DNA (NP-bulk) of the parental plants were prepared by extracting and mixing equal amounts (500 ng total DNA) of DNA samples from nine individual plants of Olr and Nip. In addition, DNA samples were extracted from 152 individual plants showing Olr grain shape (O-bulk), and another 152 individual plants showing normal grain shape (N-bulk) in the F
2 segregating population; and the two groups of DNA samples were mixed with the equal amount (500 ng total DNA) of the 152 DNA samples in each group, and finally to be served as the O-bulk and N-bulk, respectively. Then, the DNA samples frozen by drikold were shipped to Shanghai OE Biotech Inc. (Shanghai 201100, China) for DNA library construction, high throughput sequencing, and initial data analysis.
Pooled libraries with insert sizes of around 350–500 bp were prepared and sequenced on a HiSeq X™ Ten system (Illumina, San Diego, CA, USA) by using 150-base paired-end sequencing. In addition, the short reads obtained from the four samples were aligned to the reference genome of the
japonica cv. Nip (IRGSP-1.0) using BWA software (version 0.7.17, Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Cambridge, UK) [
34]. Alignment files were converted to SAM/BAM files using SAMtools and applied to the SNP-calling.
Unique SNP sites were filtered (ratio ≥ 0.8) in both maternal bulk (OM-bulk) and paternal bulk (NP-bulk) by comparing the two bulks. The intersections among unique SNP sites from the two maternal & paternal bulks and SNP sites of the progeny bulks (O-bulk and N-bulk) were taken. Then the SNP index of the screened SNP sites was calculated. SNP-index = Number of the reads with the SNP derived from the maternal plant, Olr/(Number of the reads with the SNP derived from the maternal plant, Olr + Number of the reads with the SNP derived from the paternal plant, Nip). If SNP-index = 0, it indicates that all the reads of the SNP were derived from the paternal plant, Nip; but if SNP-index = 1, it indicates that all the reads of the SNP were derived from the maternal plant, Olr.
Sliding window analysis was subsequently applied to calculate the SNP-index of O-bulk and N-bulk and to calculate the ∆(SNP-index) between O-bulk and N-bulk (O-bulk—N-bulk) based on a 1 Mb interval with a 10 kb sliding window. The SNP-index plots (O-bulk and N-bulk), ∆(SNP-index) plot (O-bulk—N-bulk), as well as the corresponding linkage maps, were simultaneously drawn and constructed. For the ∆(SNP-index) plot, the 99% (p < 0.01) and 95% (p < 0.05) statistical confidence intervals were set under the null hypothesis of no causal gene, or QTL was identified within the confidence intervals.
Location and further screening of the candidate causal SNPs and corresponding candidate genes were based on the criteria that SNP-index in paternal bulk (PN-bulk) is 0.00, SNP-index in maternal bulk (MO-bulk) is no less than 0.85, SNP-index in O-bulk is 1.00, SNP-index in N-bulk ranges from 0.30 to 0.40, the corresponding Δ(SNP-index) between O-bulk and N-bulk ranges from 0.60 to 0.70, the SNP is located within the gene which either caused missense mutation or stop codon of the encoding protein. The screened candidate causal SNPs, positions, corresponding candidate genes, location of mutations within genes, effects on amino acid changes, and predicted gene functions were annotated using SnpEff software (version 5.1,
https://pcingola.github.io/SnpEff/, accessed on 27 April 2023).
4.8. Immunoblot Analysis
The proteins were extracted from the flag leaves, florets, and seeds of rice using the Lysis Buffer [50 mM Tris-HCl, pH 7.5; 150 mM NaCl; 1 mM EDTA; 10% glycerol; 2 mM Na3VO4; 25 mM glycerophosphate; 10 mM NaF; 0.05–0.1% Tween20; 1 × Protease Inhibitor Cocktail (MedChemExpress, Shanghai, China); 1 mM PMSF]. The obtained protein lysates were immunoblotted with the OsMADS1 antibody (Catalog No.: A20328; ABclonal Technology Co., Ltd., Wuhan, China), and the abundance of HSP82 protein detected by the HSP82 antibody (Beijing Protein Innovation Co., Ltd., Beijing, China) was used as an internal control.
4.9. Three-Dimensional Protein Structure Prediction
The 3D protein structure prediction of wild-type OsMADS1 and mutated OsMADS1
Olr proteins was performed using the PyMOL molecular visualization program (
https://pymol.org/2/, accessed on 27 April 2023).
4.10. Protein Subcellular Localization
The full-length CDS (coding sequence) of wild-type
OsMADS1 and mutated
OsMADS1Olr was amplified from the Nipponbare and Oat-like rice cDNA using the PCR primers listed in
Table S8. The verified CDS fragments of
OsMADS1 and
OsMADS1Olr were cloned in the frame in front of the eGFP CDS in the pJIT163-P
35S::
eGFP vector to construct the pJIT163-P
35S::
OsMADS1-eGFP and pJIT163-P
35S::
OsMADS1Olr-eGFP transient overexpression vector driven by the 35S promoter. P
35S::
OsMADS1-eGFP or pJIT163-P
35S::
OsMADS1Olr-eGFP vector was transferred into and transiently expressed in rice protoplasts using the methods as described previously [
35,
36]. The eGFP fluorescent signals were detected using a confocal laser-scanning microscope (Leica TCS SP8, Wetzlar, Germany).
4.11. qRT-PCR Analysis
To verify the Illumina comprehensive transcriptomic data, the same batch of samples for preparation of the Illumina sequencing libraries was used to analyze the expression profiles of storage protein-related genes in Nip and Olr by qRT-PCR analysis. The qRT-PCR analysis, including cDNA synthesis, was performed following the method described by Li et al. (2020) [
10]. The
OsActin gene in rice was used as an endogenous control. The relative gene expression levels were calculated by using the 2
−ΔΔCT algorithm. The primers used in the qRT-PCR analysis are listed in
Table S9.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}