

Transgenerational Paternal Inheritance of TaCKX GFMs Expression Patterns Indicate a Way to Select Wheat Lines with Better Parameters for Yield-Related Traits


Abstract
:1. Introduction
2. Results
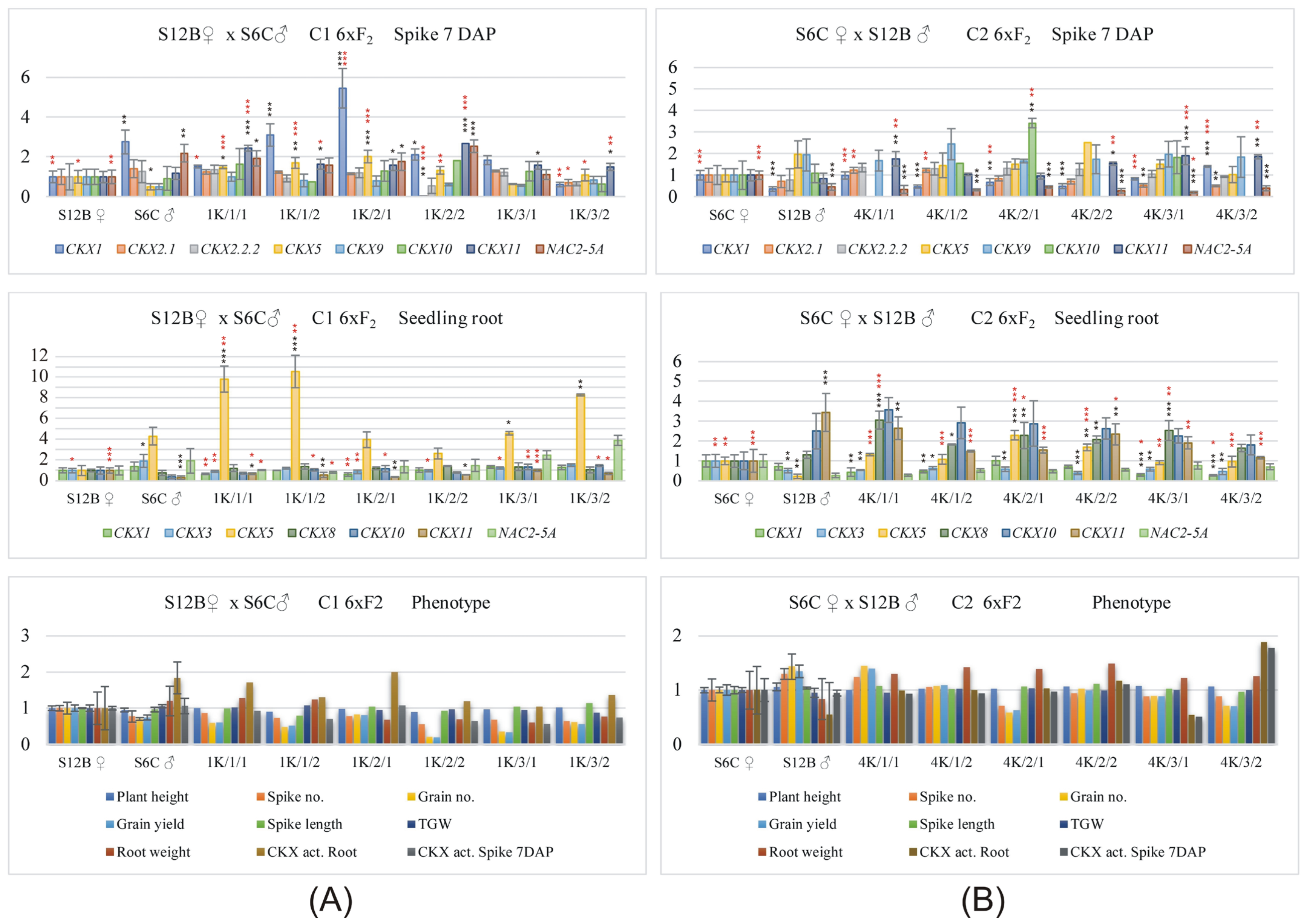
2.1. Reciprocal Crosses Indicate That the Expression Patterns of Most of the TaCKX GFM and Yield-Related Traits Are Mainly Inherited from the Male Parent
2.2. Cooperating and Opposite-Functioning Genes
2.3. The Correlation Coefficients between TaCKX GFMs and NAC2 Expression, CKX Activity, and Yield-Related Traits Were Significant for Both Parents or the Maternal or Paternal Parent Separately
2.4. Correlations between TaCKX GFM and NAC2 Expression and Yield-Related Traits in the Group of Maternal Plants, and F2 and Paternal Plants, and F2 of Reciprocal Crosses
2.4.1. Seed Number
2.4.2. Spike Number
2.4.3. TGW
2.4.4. Root Mass
2.4.5. Semi-Empty Spikes
3. Discussion
3.1. The Expression Patterns of Most TaCKX GFMs and TaNAC2-5A Are Mainly Inherited from the Paternal Parent
3.2. Cooperation of TaCKX GFMs and TaNAC2-5A in the Determination of Yield-Related Traits
3.3. Regulation of Yield-Related Traits by TaCKX GFMs and TaNAC2-5A in the F2 Generation
4. Materials and Methods
4.1. Plant Material
4.2. Cross-Breeding
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative RT-qPCR
4.5. Analysis of CKX Activity
4.6. Measurement of Yield-Related Traits
4.7. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Yoo, M.J.; Liu, X.X.; Pires, J.C.; Soltis, P.S.; Soltis, D.E. Nonadditive Gene Expression in Polyploids. Annu. Rev. Genet. 2014, 48, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Leach, L.J.; Belfield, E.J.; Jiang, C.F.; Brown, C.; Mithani, A.; Harberd, N.P. Patterns of homoeologous gene expression shown by RNA sequencing in hexaploid bread wheat. BMC Genom. 2014, 15, 276. [Google Scholar] [CrossRef] [PubMed]
- Jameson, P.E.; Song, J.C. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef]
- Jablonski, B.; Bajguz, A.; Bocian, J.; Orczyk, W.; Nadolska-Orczyk, A. Genotype-Dependent Effect of Silencing of TaCKX1 and TaCKX2 on Phytohormone Crosstalk and Yield-Related Traits in Wheat. Int. J. Mol. Sci. 2021, 22, 1494. [Google Scholar] [CrossRef]
- Jablonski, B.; Ogonowska, H.; Szala, K.; Bajguz, A.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of TaCKX1 Mediates Expression of Other TaCKX Genes to Increase Yield Parameters in Wheat. Int. J. Mol. Sci. 2020, 21, 4908. [Google Scholar] [CrossRef]
- Jablonski, B.; Szala, K.; Przyborowski, M.; Bajguz, A.; Chmur, M.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. TaCKX2.2 Genes Coordinate Expression of Other TaCKX Family Members, Regulate Phytohormone Content and Yield-Related Traits of Wheat. Int. J. Mol. Sci. 2021, 22, 4142. [Google Scholar] [CrossRef]
- Szala, K.; Ogonowska, H.; Lugowska, B.; Zmijewska, B.; Wyszynska, R.; Dmochowska-Boguta, M.; Orczyk, W.; Nadolska-Orczyk, A. Different sets of TaCKX genes affect yield-related traits in wheat plants grown in a controlled environment and in field conditions. BMC Plant Biol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Perry, L.; Kisiala, A.; Olechowski, H.; Emery, R.J.N. Cytokinin activity during early kernel development corresponds positively with yield potential and later stage ABA accumulation in field-grown wheat (Triticum aestivum L.). Planta 2020, 252, 1–16. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, J.Q.; Song, J.C.; Jameson, P.E. Cytokinin dehydrogenase: A genetic target for yield improvement in wheat. Plant Biotechnol. J. 2020, 18, 614–630. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, J.; Song, J.C.; Jameson, P.E. Cytokinin glucosyl transferases, key regulators of cytokinin homeostasis, have potential value for wheat improvement. Plant Biotechnol. J. 2021, 19, 878–896. [Google Scholar] [CrossRef]
- Chen, L.; Jameson, G.B.; Guo, Y.C.; Song, J.C.; Jameson, P.E. The LONELY GUY gene family: From mosses to wheat, the key to the formation of active cytokinins in plants. Plant Biotechnol. J. 2022, 20, 625–645. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Lai, N.; Kisiala, A.B.; Emery, R.J.N. Isopentenyltransferases as master regulators of crop performance: Their function, manipulation, and genetic potential for stress adaptation and yield improvement. Plant Biotechnol. J. 2021, 19, 1297–1313. [Google Scholar] [CrossRef]
- Wang, N.; Chen, J.; Gao, Y.; Zhou, Y.; Chen, M.; Xu, Z.; Fang, Z.; Ma, Y. Genomic analysis of isopentenyltransferase genes and functional characterization of TaIPT8 indicates positive effects of cytokinins on drought tolerance in wheat. Crop. J. 2023, 11, 46–56. [Google Scholar] [CrossRef]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmulling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Will cytokinins underpin the second ‘Green Revolution’? J. Exp. Bot. 2020, 71, 6872–6875. [Google Scholar] [CrossRef]
- Schwarz, I.; Scheirlinck, M.T.; Otto, E.; Bartrina, I.; Schmidt, R.C.; Schmulling, T. Cytokinin regulates the activity of the inflorescence meristem and components of seed yield in oilseed rape. J. Exp. Bot. 2020, 71, 7146–7159. [Google Scholar] [CrossRef]
- Chang, C.; Lu, J.; Zhang, H.P.; Ma, C.X.; Sun, G.L. Copy Number Variation of Cytokinin Oxidase Gene Tackx4 Associated with Grain Weight and Chlorophyll Content of Flag Leaf in Common Wheat. PLoS ONE 2015, 10, e0145970. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, Y.L.; Gao, L.F.; Zhao, G.Y.; Zhou, R.H.; Zhang, B.S.; Jia, J.Z. TaCKX6-D1, the ortholog of rice OsCKX2, is associated with grain weight in hexaploid wheat. New Phytol. 2012, 195, 74–584. [Google Scholar] [CrossRef]
- Lu, J.; Chang, C.; Zhang, H.P.; Wang, S.X.; Sun, G.; Xiao, S.H.; Ma, C.X. Identification of a Novel Allele of TaCKX6a02 Associated with Grain Size, Filling Rate and Weight of Common Wheat. PLoS ONE 2015, 10, e0144765. [Google Scholar] [CrossRef] [PubMed]
- Ramireddy, E.; Hosseini, S.A.; Eggert, K.; Gillandt, S.; Gnad, H.; von Wiren, N.; Schmulling, T. Root Engineering in Barley: Increasing Cytokinin Degradation Produces a Larger Root System, Mineral Enrichment in the Shoot and Improved Drought Tolerance. Plant Physiol. 2018, 177, 1078–1095. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Nehnevajova, E.; Kollmer, I.; Novak, O.; Strnad, M.; Kramer, U.; Schmulling, T. Root-Specific Reduction of Cytokinin Causes Enhanced Root Growth, Drought Tolerance, and Leaf Mineral Enrichment in Arabidopsis and Tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Bocian, J.; Hameed, A.; Orczyk, W.; Nadolska-Orczyk, A. Cis-Regulation by NACs: A Promising Frontier in Wheat Crop Improvement. Int. J. Mol. Sci. 2022, 23, 5431. [Google Scholar] [CrossRef]
- He, X.; Qu, B.Y.; Li, W.J.; Zhao, X.Q.; Teng, W.; Ma, W.Y.; Ren, Y.Z.; Li, B.; Li, Z.S.; Tong, Y.P. The Nitrate-Inducible NAC Transcription Factor TaNAC2-5A Controls Nitrate Response and Increases Wheat Yield. Plant Physiol. 2015, 169, 991–2005. [Google Scholar] [CrossRef]
- Zhao, D.; Derkx, A.P.; Liu, D.C.; Buchner, P.; Hawkesford, M.J. Overexpression of a NAC transcription factor delays leaf senescence and increases grain nitrogen concentration in wheat. Plant Biol. 2015, 17, 904–913. [Google Scholar] [CrossRef]
- Gimenez, E.; Benavente, E.; Pascual, L.; Garcia-Sampedro, A.; Lopez-Fernandez, M.; Vazquez, J.F.; Giraldo, P. An F-2 Barley Population as a Tool for Teaching Mendelian Genetics. Plants 2021, 10, 694. [Google Scholar] [CrossRef]
- Nadolska-Orczyk, A.; Orczyk, W.; Przetakiewicz, A. Agrobacterium-mediated transformation of cereals—From technique development to its application. Acta Physiol. Plant. 2000, 22, 77–88. [Google Scholar] [CrossRef]
- Howells, R.M.; Craze, M.; Bowden, S.; Wallington, E.J. Efficient generation of stable, heritable gene edits in wheat using CRISPR/Cas9. BMC Plant Biol. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Luo, M.; Li, H.Y.; Chakraborty, S.; Morbitzer, R.; Rinaldo, A.; Upadhyaya, N.; Bhatt, D.; Louis, S.; Richardson, T.; Lahaye, T.; et al. Efficient TALEN-mediated gene editing in wheat. Plant Biotechnol. J. 2019, 17, 2026–2028. [Google Scholar] [CrossRef]
- Hudzieczek, V.; Hobza, R.; Capal, P.; Safar, J.; Dolezel, J. If Mendel Was Using CRISPR: Genome Editing Meets Non-Mendelian Inheritance. Adv. Funct. Mater. 2022, 32, 2202585. [Google Scholar] [CrossRef]
- Cortes, A.J.; This, D.; Chavarro, C.; Madrinan, S.; Blair, M.W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortes, A.J.; This, D. Identification of an ERECTA gene and its drought adaptation associations with wild and cultivated common bean. Plant Sci. 2016, 242, 250–259. [Google Scholar] [CrossRef]
- Zemlyanskaya, E.V.; Dolgikh, V.A.; Levitsky, V.G.; Mironova, V. Transcriptional regulation in plants: Using omics data to crack the cis-regulatory code. Curr. Opin. Plant Biol. 2021, 63, 102058. [Google Scholar] [CrossRef]
- Fagny, M.; Austerlitz, F. Polygenic Adaptation: Integrating Population Genetics and Gene Regulatory Networks. Trends Genet. 2021, 37, 631–638. [Google Scholar] [CrossRef]
- Batista, R.A.; Kohler, C. Genomic imprinting in plants-revisiting existing models. Gene Dev. 2020, 34, 24–36. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Zilberman, D. Evolution and function of genomic imprinting in plants. Gene Dev. 2015, 29, 2517–2531. [Google Scholar] [CrossRef]
- Waters, A.J.; Bilinski, P.; Eichten, S.R.; Vaughn, M.W.; Ross-Ibarra, J.; Gehring, M.; Springer, N.M. Comprehensive analysis of imprinted genes in maize reveals allelic variation for imprinting and limited conservation with other species. Proc. Natl. Acad. Sci. USA 2013, 110, 19639–19644. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Hsieh, P.H.; Ruan, D.L.; Nishimura, T.; Sharma, M.K.; Sharma, R.; Ye, X.Y.; Nguyen, N.D.; Nijjar, S.; Ronald, P.C.; et al. Divergence among rice cultivars reveals roles for transposition and epimutation in ongoing evolution of genomic imprinting. Proc. Natl. Acad. Sci. USA 2021, 118, e2104445118. [Google Scholar] [CrossRef]
- Robert, H.S.; Park, C.; Gutierrez, C.L.; Wojcikowska, B.; Pencik, A.; Novak, O.; Chen, J.Y.; Grunewald, W.; Dresselhaus, T.; Friml, J.; et al. Maternal auxin supply contributes to early embryo patterning in Arabidopsis. Nat. Plants 2018, 4, 548–553. [Google Scholar] [CrossRef]
- Zhao, P.; Zhou, X.M.; Shen, K.; Liu, Z.Z.; Cheng, T.H.; Liu, D.N.; Cheng, Y.B.; Peng, X.B.; Sun, M.X. Two-Step Maternal-to-Zygotic Transition with Two-Phase Parental Genome Contributions. Dev. Cell 2019, 49, 882. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.R.; Evans, M.M.S. Maternal regulation of seed growth and patterning in flowering plants. Curr. Top Dev. Biol. 2020, 140, 257–282. [Google Scholar] [PubMed]
- Olsen, O.A. The Modular Control of Cereal Endosperm Development. Trends Plant Sci. 2020, 25, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Pan, M.; Zhiguo, E.; Zhou, Y.; Niu, B.; Chen, C. The maternally expressed polycomb group gene OsEMF2a is essential for endosperm cellularization and imprinting in rice. Plant Commun. 2021, 2, 100092. [Google Scholar] [CrossRef]
- Jia, D.; Chen, L.G.; Yin, G.; Yang, X.; Gao, Z.; Guo, Y.; Sun, Y.; Tang, W. Brassinosteroids regulate outer ovule integument growth in part via the control of inner no outer by brassinozole-resistant family transcription factors. J. Integr. Plant Biol. 2020, 62, 1093. [Google Scholar] [CrossRef]
- Lafon-Placette, C.; Hatorangan, M.R.; Steige, K.A.; Cornille, A.; Lascoux, M.; Slotte, T.; Kohler, C. Paternally expressed imprinted genes associate with hybridization barriers in Capsella. Nat. Plants 2018, 4, 352. [Google Scholar] [CrossRef]
- Dai, D.W.; Mudunkothge, J.S.; Galli, M.; Char, S.N.A.; Davenport, R.; Zhou, X.J.; Gustin, J.L.; Spielbauer, G.; Zhang, J.Y.; Barbazuk, W.B.; et al. Paternal imprinting of dosage-effect defective1 contributes to seed weight xenia in maize. Nat. Commun. 2022, 13, 1–11. [Google Scholar] [CrossRef]
- Long, J.; Walker, J.; She, W.; Aldridge, B.; Gao, H.; Deans, S.; Vickers, M.; Feng, X. Nurse cell--derived small RNAs define paternal epigenetic inheritance in Arabidopsis. Science 2021, 373, eabh0556. [Google Scholar] [CrossRef]
- Gehring, M. Epigenetic dynamics during flowering plant reproduction: Evidence for reprogramming? New Phytol. 2019, 224, 91–96. [Google Scholar] [CrossRef]
- Ono, A.; Kinoshita, T. Epigenetics and plant reproduction: Multiple steps for responsibly handling succession. Curr. Opin. Plant Biol. 2021, 61, 102032. [Google Scholar] [CrossRef]
- Kirkbride, R.C.; Lu, J.; Zhang, C.; Mosher, R.A.; Baulcombe, D.C.; Chen, Z.J. Maternal small RNAs mediate spatial-temporal regulation of gene expression, imprinting, and seed development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 2761–2766. [Google Scholar] [CrossRef]
- Tuteja, R.; McKeown, P.C.; Ryan, P.; Morgan, C.C.; Donoghue, M.T.A.; Downing, T.; O’Connell, M.J.; Spillane, C. Paternally Expressed Imprinted Genes under Positive Darwinian Selection in Arabidopsis thaliana. Mol. Biol. Evol. 2019, 36, 1239–1253. [Google Scholar] [CrossRef]
- Moreno-Romero, J.; Jiang, H.; Santos-Gonzalez, J.; Kohler, C. Parental epigenetic asymmetry of PRC2-mediated histone modifications in the Arabidopsis endosperm. EMBO J. 2016, 35, 1298–1311. [Google Scholar] [CrossRef]
- Quadrana, L.; Colot, V. Plant Transgenerational Epigenetics. Annu. Rev. Genet. 2016, 50, 467–491. [Google Scholar] [CrossRef]
- Zalewski, W.; Galuszka, P.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of the HvCKX1 gene decreases the cytokinin oxidase/dehydrogenase level in barley and leads to higher plant productivity. J. Exp. Bot. 2010, 61, 1839–1851. [Google Scholar] [CrossRef]
- Gasparis, S.; Przyborowski, M.; Kala, M.; Nadolska-Orczyk, A. Knockout of the HvCKX1 or HvCKX3 Gene in Barley (Hordeum vulgare L.) by RNA-Guided Cas9 Nuclease Affects the Regulation of Cytokinin Metabolism and Root Morphology. Cells 2019, 8, 782. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.Y.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Ogonowska, H.; Barchacka, K.; Gasparis, S.; Jablonski, B.; Orczyk, W.; Dmochowska-Boguta, M.; Nadolska-Orczyk, A. Specificity of expression of TaCKX family genes in developing plants of wheat and their co-operation within and among organs. PLoS ONE 2019, 14, e0214239. [Google Scholar] [CrossRef]
- Li, Y.L.; Song, G.Q.; Gao, J.; Zhang, S.J.; Zhang, R.Z.; Li, W.; Chen, M.L.; Liu, M.; Xia, X.C.; Risacher, T.; et al. Enhancement of grain number per spike by RNA interference of cytokinin oxidase 2 gene in bread wheat. Hereditas 2018, 155, 1–8. [Google Scholar] [CrossRef]
- Zhang, W.; Peng, K.X.; Cui, F.B.; Wang, D.L.; Zhao, J.Z.; Zhang, Y.J.; Yu, N.N.; Wang, Y.Y.; Zeng, D.L.; Wang, Y.H.; et al. Cytokinin oxidase/dehydrogenase OsCKX11 coordinates source and sink relationship in rice by simultaneous regulation of leaf senescence and grain number. Plant Biotechnol. J. 2020, 19, 335–350. [Google Scholar] [CrossRef]
- Li, W.J.; He, X.; Chen, Y.; Jing, Y.F.; Shen, C.C.; Yang, J.B.; Teng, W.; Zhao, X.Q.; Hu, W.J.; Hu, M.Y.; et al. A wheat transcription factor positively sets seed vigour by regulating the grain nitrate signal. New Phytol. 2020, 225, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.J.; He, J.M.; Liu, L.N.; Deng, Q.M.; Yao, X.F.; Liu, C.M.; Qiao, Y.L.; Li, P.; Ming, F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol. J. 2020, 18, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Frebort, I.; Sebela, M.; Galuszka, P.; Werner, T.; Schmulling, T.; Pec, P. Cytokinin oxidase/cytokinin dehydrogenase assay: Optimized procedures and applications. Anal. Biochem. 2002, 306, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M.; Williams, W.L. New, Rapid, Sensitive Method for Protein Determination. Fed. Proc. 1976, 35, 274. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| M | P | F2 | |
|---|---|---|---|
| C1 = S12B × S6C | |||
| CKX expression 7 DAP | CKX1↓ | CKX1↑ | CKX1, 11↑↑ |
 | NAC2↓ | NAC2↑ | CKX5, NAC2↑ |
| CKX5, 9↑ | CKX5, 9↓ | ||
| CKX expression root | CKX5, NAC2↓↓ | CKX5↑↑ | CKX5↑↑↑, NAC2↑ |
 | CKX3↓ | CKX3↑, NAC2↑ | |
| CKX11, 10↑ | CKX11, 10↓ | CKX11↓ | |
| yield-related traits | yield↑ | yield↓ | yield↓↓ |
| CKX act. spike= | CKX act. spike= | CKX act. spike↓↓ | |
| root=↓ | root=↑ | root=↓ | |
| CKX act. root↓↓ | CKX act. root↑↑ | CKX act. root↑↑ | |
| C2 = S6C × S12B | |||
| CKX expression 7 DAP | CKX5, 9↓ | CKX5, 9↑ | CKX9, 10, 11, 5↑ |
| CKX1, 2.1, NAC2↑ | CKX1, 2.1, NAC2↓ | CKX1, NAC2↓ |
| CKX expression root | CKX10, 11↓↓ | CKX10, 11↑↑ | CKX5, 8, 10, 11↑↑ |
| CKX1, 3, 5, NAC2↑ | CKX1, 3, 5, NAC2↓ | CKX1, 3, NAC2↓ |
| yield-related traits | yield↓ | yield↑ | yield=↓ |
| CKX act. spike= | CKX act. spike= | CKX act. Spike= | |
| root↑ | root=↓ | root↑↑ | |
| CKX act. root↑ | CKX act. root↓ | CKX act. root= | |
| C3 = D16 × KOH7 | |||
| CKX expression 7 DAP | CKX11↓↓ | CKX11↑↑ | CKX5, 9↑↑ |
| CKX2.1, 2.2.2↓ | CKX2.1, 2.2.2↑ | |
| CKX10↑ | CKX10↓ | ||
| CKX expression root | NAC2↓↓ | NAC2↑↑ | CKX3↓ |
| CKX1, 5, 10, 11↓↓ | CKX1, 5, 10, 11↑↑ | CKX1, 8, 11, NAC2↑↑ |
| yield-related traits | yield↑ | yield↓ | yield↓ |
| CKX act. spike=↑ | CKX act. spike=↓ | CKX act. spike↓↓ | |
| root=↑ | root=↓ | root= | |
| semi-empty spikes↑↑↑ | |||
| C4 = KOH7 × D16 | |||
| CKX expression 7 DAP | CKX9, 10↓ | CKX9, 10↑ | CKX5, 9↑↑ |
| CKX11↑↑ | CKX11↓↓ | CKX11, NAC2↓↓ |
| CKX2.1, 2.2.2↑ | CKX2.1, 2.2.2↓ | CKX2.1, 2.2.2↓ | |
| CKX expression root | CKX3, 8↓ | CKX3, 8↑ | CKX8↑↑ |
| CKX1, 5, 10, 11↑ | CKX1, 5, 10, 11↓ | CKX3,10↓ |
| NAC2↑↑ | NAC2↓↓ | ||
| yield-related traits | yield↓ | yield↑ | yield↑ |
| CKX act. spike=↓ | CKX act. spike=↑ | CKX act. spike= | |
| root↓ | root↑ | root=↓ | |
| C5 = D19 × D16 | |||
| CKX expression 7 DAP | CKX2.2.2↑ | CKX2.2.2↓ | CKX2.2.2↓ |
| CKX10↓ | CKX10↑ | CKX9↑ |
| CKX expression root | CKX 5, 8, NAC2↑ | CKX5, 8, NAC2↓↓ | CKX3, 5, 8, 11↓ |
| CKX1, 3, 10, 11↑ | CKX1, 3, 10, 11↓ | (CKX10, NAC2↑) |
| yield-related traits | yield↑ | yield↓ | yield↓↓ |
| CKX act. spike=↓ | CKX act. spike=↑ | CKX act. spike=↑ | |
| root= | root= | root↓ | |
| semi-empty spikes↓ | semi-empty spikes↑ | semi-empty spikes↑↑↑ | |
| C6 = D16 × D19 | |||
| CKX expression 7 DAP | CKX2.2.2, 5, 9↓ | CKX2.2.2, 5, 9↑ | |
| CKX10↑ | CKX10↓ | CKX1, 10, 11, NAC2↓ |
| CKX expression root | CKX5, 8, NAC2↓↓ | CKX5, 8, NAC2↑↑ | CKX1, NAC2↑↑ |
| CKX1, 3, 10, 11↓ | CKX1, 3,10, 11↑ | CKX8, 10↑ |
| yield-related traits | yield↓ | yield↑↑ | yield↑ |
| CKX act. spike=↑ | CKX act. spike=↓ | CKX act. spike= | |
| root= | root= | root↓ | |
| C7 = D19 × KOH7 | |||
| CKX expression 7 DAP | CKX2.1, 11, NAC2↓ | CKX2.1, 11, NAC2↑ | CKX10↑ |
| CKX9↑ | CKX9↓ | CKX2.2.2, 9↓ |
| NAC2↓ | |||
| CKX expression root | NAC2↓↓ | NAC2↑↑ | |
| CKX1, 10, 11↓ | CKX1, 10, 11↑ | CKX11↑ |
| CKX3, 5, 8↑ | CKX3, 5, 8↓ | CKX3, 5, 8, 10↓ | |
| yield-related traits | yield↑ | yield↓ | yield↓ |
| CKX act. spike= | CKX act. spike= | CKX act. spike↑ | |
| root=↑ | root=↓ | root= | |
| semi-empty spikes↑ | semi-empty spikes↑↑ | ||
| C8 = KOH7 × D19 | |||
| CKX expression 7 DAP | CKX5, 9, 10↓ | CKX5, 9, 10↑ | CKX9, 10↑ |
| CKX11↑ | CKX11↓ | CKX11, NAC2↓ |
| CKX2.1, NAC2=↑ | CKX2.1, NAC2=↓ | CKX2.1, 2.2.2↓ | |
| CKX expression root | CKX8↓↓ | CKX8↑↑ | CKX3, 8↑↑ |
| CKX3, 5↓ | CKX3, 5↑ | CKX1, 10, NAC2↓ |
| NAC2↑↑ | NAC2↓↓ | ||
| yield-related traits | yield↓ | yield↑↑ | yield↑↑ |
| CKX act. spike= | CKX act. spike= | CKX act. spike↑ | |
| root=↓ | root=↑ | root= |
| Cross | P Spike P Expr. +/P expr. − | P Root P Expr. +/P Expr. − | F2 Spike/Root (Yield, Root Mass in F2) |
|---|---|---|---|
| C1 | 1, NAC2/5, 9 | 3, 5, NAC2/10, 11 | NAC2+/NAC2+ y−, r = −, As−− |
| C2 | 5, 9/1, NAC2 | 10, 11/3, 5, NAC2 | NAC2−/NAC2− y = −, r+, As = |
| C3 | 2.1, 11, 2.2.2/10 | NAC2, 1, 5, 10, 11/3 | 10−, 11+/10+,11+, NAC2+ y−, r=, As−, s-e+++ |
| C4 | 9, 10/2.1, 11, 2.2.2 | 3, 8/1, 5, 10, 11, NAC2 | 10+, 11−/10−, 11−, NAC2− y+, r = −, As= |
| C5 | 2.2.2/10 | ?/3, 5, 8, NAC2, 1, 10, 11 | 10−/10−, NAC2− y−, r−, As = +, s-e+++ |
| C6 | 2.2.2, 5, 9/10 | 3, 5, 8, NAC2, 1, 10, 11/none | 5+, 10−/ 5+, 10+, NAC2+ y+, r−, As= |
| C7 | 2.1, 11, NAC2/9 | 1, 10, 11, NAC2/3, 5, 8 | 11+, NAC2+/11+, NAC2+ y−, r=, As+, s-e+++ |
| C8 | 5, 9, 10/2.1, 11, NAC2 | 3, 5, 8/NAC2 | 5+, NAC2−/5+, NAC2− y++, r=, As+ |
| 7 DAP Spike | Seedling Root | Yield-Related Traits | F2 Phenotype | ||
|---|---|---|---|---|---|
| Seed Number | |||||
| C1 S12B × S6C | M + F2 | CKX1−, CKX5− | CKX11+ | spike number+ | yield−−, CKX act. −−, root=−, CKX act. root++ |
| P + F2 | nc | CKX11+ | spike number+ | ||
| C2 S6C × S12B | M + F2 | nc | nc | CKX act. root−, spike number++ | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | CKX3+ | CKX act. root−, spike number+ | ||
| C3 D16 × KOH7 | M + F2 | CKX2.1+,CKX2.2.2+ | CKX3+, CKX8+,CKX11− | plant height+, spike number+ | yield−, CKX act. −−, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1+, CKX2.2.2+ | CKX1−, CKX3++, CKX8+, CKX11− | plant height+, spike number+ | ||
| C4 KOH7 × D16 | M + F2 | nc | nc | plant height+, spike number+ | yield+, CKX act.=, root= |
| P + F2 | nc | nc | plant height+, spike number+ | ||
| C5 D19 × D16 | M + F2 | CKX1− | CKX3+, CKX5+, NAC2+ | plant height+, spike number+ | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX3+, NAC2+ | plant height+ spike number+ | ||
| C6 D16 × D19 | M + F2 | nc | nc | spike number+ | yield+, CKX act.=, root=− |
| P + F2 | nc | nc | spike number+ | ||
| C7 D19 × KOH7 | M + F2 | nc | CKX1− | spike number++ | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX1− | spike number++ | ||
| C8 KOH7 × D19 | M + F2 | nc | CKX1− | empty spikes−, spike number+ | yield++, CKX act.=+, root= |
| P + F2 | CKX2.1+ | CKX1− | empty spikes−, spike number+ | ||
| Seed yield | |||||
| C1 S12B × S6C | M + F2 | CKX11−, NAC2− | CKX11+ | spike number+, seed number++ | yield−, CKX act. −−, root=−, CKX act. root++ |
| P + F2 | nc | CKX11+ | spike number+, seed number++ | ||
| C2 S6C × S12B | M + F2 | nc | CKX10+ | spike number+, seed number++ | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | CKX10+, CKX3+ | spike number+, seed number++ | ||
| C3 D16 × KOH7 | M + F2 | CKX2.1+, CKX2.2.2+ | CKX3+, CKX8+ | plant height+, spike number+, seed number++ | yield−, CKX act. −−, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1+, CKX2.2.2+ | CKX3+, CKX8+ | plant height+, spike number+, seed number++ | ||
| C4 KOH7 × D16 | M + F2 | nc | nc | plant height+, spike number+, seed number++ | yield+, CKX act.=, root= |
| P + F2 | nc | nc | plant height+, spike number+, seed number++ | ||
| C5 D19 × D16 | M + F2 | CKX1− | CKX3+, CKX5+, NAC2+ | plant height+, spike number+, seed number++ | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX3+, NAC2+ | plant height+, spike number+, seed number++ | ||
| C6 D16 × D19 | M + F2 | CKX2.1+ | nc | plant height+, spike number+, seed number++ | yield+, CKX act.=, root=− |
| P + F2 | CKX1− | nc | plant height+, spike number+, seed number++ | ||
| C7 D19 × KOH7 | M + F2 | nc | nc | plant height+, spike number++, seed number++ | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | nc | plant height+, spike number++, seed number++ | ||
| C8 KOH7 × D19 | M + F2 | nc | CKX1− | spike number+, seed number++ | yield++, CKX act.=+, root= |
| P + F2 | nc | CKX1− | spike number+, seed number++ | ||
| Spike number | |||||
| C1 S12B × S6C | M + F2 | CKX2.1+, CKX2.2.2+, CKX11− | CKX11+ | plant height+ | yield−, CKX act.−−, root=−, CKX act. root++ |
| P + F2 | CKX2.1+, CKX2.2.2+ | nc | CKX act.+, plant height+ | ||
| C2 S6C × S12B | M + F2 | CKX2.2.2+ | nc | nc | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | NAC2− | nc | ||
| C3 D16 × KOH7 | M + F2 | nc | CKX8+ | nc | yield−, CKX act. −−, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX8+ | nc | ||
| C4 KOH7 × D16 | M + F2 | nc | CKX5+ | nc | yield+, CKX act.=, root= |
| P + F2 | nc | CKX5+ | nc | ||
| C5 D19 × D16 | M + F2 | nc | CKX1−, CKX5+, NAC2+ | nc | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX1−, CKX5+, NAC2+ | nc | ||
| C6 D16 × D19 | M + F2 | nc | nc | nc | yield+, CKX act.=, root=− |
| P + F2 | CKX2.1+ | nc | nc | ||
| C7 D19 × KOH7 | M + F2 | nc | CKX1− | nc | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX1−, CKX8− | nc | ||
| C8 KOH7 × D19 | M + F2 | nc | CKX1−, CKX5−, CKX8− | nc | yield++, CKX act.=+, root= |
| P + F2 | nc | CKX1−, CKX5− | nc | ||
| TGW | |||||
| C1 S12B × S6C | M + F2 | CKX2.1+, CKX10+, NAC2+ | nc | nc | yield−, CKX act. −−, root=−, CKX act. root++ |
| P + F2 | CKX2.2.2+, NA2C+ | nc | nc | ||
| C2 S6C × S12B | M + F2 | nc | nc | CKX act. root++, seed number− | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | nc | CKX act. root++, seed number− | ||
| C3 D16 × KOH7 | M + F2 | nc | nc | plant height+, yield+, semi-empty−, seed number+ | yield−, CKX act. −−, root=−, semi-empty spikes++ |
| P + F2 | nc | nc | plant height+, yield+ | ||
| C4 KOH7 × D16 | M + F2 | CKX2.2.2− | CKX10+, NAC2+ | CKX act.− | yield+, CKX act.=, root= |
| P + F2 | CKX2.2.2− | CKX10+,NAC2+ | CKX act.− | ||
| C5 D19 × D16 | M + F2 | nc | NAC2+ | yield+, seed number+ | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | NAC2+ | yield+ | ||
| C6 D16 × D19 | M + F2 | nc | nc | plant height+ | yield+, CKX act.=, root=− |
| P + F2 | nc | nc | plant height+ | ||
| C7 D19 × KOH7 | M + F2 | CKX2.1−, CKX2.2.2−, CKX9− | nc | spike length− | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1−,CKX2.2.2−, CKX5+ | nc | nc | ||
| C8 KOH7 × D19 | M + F2 | CKX2.1−, CKX2.2.2− | nc | nc | yield++, CKX act.=+, root= |
| P + F2 | CKX2.1−, CKX2.2.2−!, CKX10− | nc | nc | ||
| Root mass | |||||
| C1 S12B × S6C | M + F2 | CKX5+,CKX11+, NAC2+ | CKX1−, CKX3−, CKX10−, CKX11−, | seed number− | yield−, CKX act. −−, root=−, CKX act. root++ |
| P + F2 | nc | NAC2−, CKX1−, CKX10−, CKX11− | seed number− | ||
| C2 S6C × S12B | M + F2 | NAC2− | nc | nc | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | nc | nc | ||
| C3 D16 × KOH7 | M + F2 | CKX1+, CKX11+ | CKX5+ | nc | yield−, CKX act.−−, root=−, semi-empty spikes++ |
| P + F2 | CKX1+, CKX11+, NAC2+ | CKX5+, CKX8− | nc | ||
| C4 KOH7 × D16 | M + F2 | nc | CKX3+, CKX8+, NAC2− | CKX act.+ | yield+, CKX act.=, root= |
| P + F2 | CKX2.1+, CKX2.2.2+, CKX10+ | CKX3+, CKX8+, NAC2− | nc | ||
| C5 D19 × D16 | M + F2 | NAC2− | CKX1−, CKX11+ | nc | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1+ | nc | nc | ||
| C6 D16 × D19 | M + F2 | CKX2.1+, CKX2.2.2+ | nc | yield+ | yield+, CKX act.=, root=− |
| P + F2 | CKX2.2.2+ | nc | plant height+ | ||
| C7 D19 × KOH7 | M + F2 | nc | CKX1− | nc | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | NAC2− | CKX1− | spike length+ | ||
| C8 KOH7 × D19 | M + F2 | CKX10− | CKX3− | nc | yield++, CKX act.=+, root= |
| P + F2 | nc | CKX3− | yield+ | ||
| Plant height | |||||
| C1 S12B × S6C | M + F2 | nc | nc | nc | yield−, CKX act. −−, root=−, CKX act.root++ |
| P + F2 | nc | nc | nc | ||
| C2 S6C × S12B | M + F2 | CKX11+ | CKX1− | nc | yield−, CKX act.=, root+, CKX act.root= |
| P + F2 | nc | CKX5−, CKX11+ | nc | ||
| C3 D16 × KOH7 | M + F2 | nc | CKX11− | nc | yield−, CKX act.−−, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX11− | nc | ||
| C4 KOH7 × D16 | M + F2 | nc | nc | nc | yield+, CKX act.=, root= |
| P + F2 | CKX2.2.2− | NAC2+ | nc | ||
| C5 D19 × D16 | M + F2 | CKX1− | CKX1−, NAC2+ | nc | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | CKX1−, NAC2+ | nc | ||
| C6 D16 × D19 | M + F2 | CKX1−, CKX5− | CKX5− | nc | yield+, CKX act.=, root=− |
| P + F2 | CKX5− | CKX5− | nc | ||
| C7 D19 × KOH7 | M + F2 | nc | nc | nc | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | nc | nc | ||
| C8 KOH7 × D19 | M + F2 | CKX2.2.2−, CKX9− | CKX11+ | nc | yield++, CKX act.=+, root= |
| P + F2 | CKX9− | nc | nc | ||
| Spike length | |||||
| C1 S12B × S6C | M + F2 | NAC2− | nc | semi-empty spikes−, seed number+, yield+ | yield−, CKX act.−−, root=−, CKX act. root++ |
| P + F2 | NAC2− | nc | spike number+, seed number+, yield+ | ||
| C2 S6C × S12B | M + F2 | CKX9+, CKX11+ | nc | plant height+ | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | nc | nc | nc | ||
| C3 D16 × KOH7 | M + F2 | CKX2.1+ | CKX3+, CKX11− | plant height+, seed number+, yield+ | yield−, CKX act.−−, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1+, CKX2.2.2+ | CKX1−, CKX3++, CKX11− | plant height+, seed number+, yield+ | ||
| C4 KOH7 × D16 | M + F2 | CKX1+, CKX10+ | nc | plant height+, seed number+, yield+ | yield+, CKX act.=, root= |
| P + F2 | nc | nc | seed number+, yield+ | ||
| C5 D19 × D16 | M + F2 | CKX5−, CKX11+, NAC2− | CKX11+ | nc | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | nc | seed number+, yield+ | ||
| C6 D16 × D19 | M + F2 | nc | nc | spike number+, seed number+, yield+ | yield+, CKX act.=, root=− |
| P + F2 | nc | nc | seed number+, yield+ | ||
| C7 D19 × KOH7 | M + F2 | CKX1+, CKX9+ | nc | seed number+ | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | CKX9+, NAC2− | nc | nc | ||
| C8 KOH7 × D19 | M + F2 | nc | nc | nc | yield++, CKX act.=+, root= |
| P + F2 | CKX2.1+ | nc | seed number+ | ||
| Semi-empty spikes | |||||
| C1 S12B × S6C | M + F2 | CKX5+ | CKX5−, CKX10− | nc | yield−, CKX act. −−, root=−, CKX act. root++ |
| P + F2 | CKX5+!, CKX9+, CKX11+ | nc | nc | ||
| C2 S6C × S12B | M + F2 | CKX9+ | nc | nc | yield−, CKX act.=, root+, CKX act. root= |
| P + F2 | CKX9+ | CKX8− | empty spikes+ | ||
| C3 D16 × KOH7 | M + F2 | CKX9+ | nc | nc | yield−, CKX act.−−, root=−, semi-empty spikes++ |
| P + F2 | CKX11− | nc | nc | ||
| C4 KOH7 × D16 | M + F2 | nc | nc | spike number+ | yield+, CKX act.=, root= |
| P + F2 | nc | nc | spike number+ | ||
| C5 D19 × D16 | M + F2 | nc | nc | nc | yield−−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | CKX2.1− | nc | nc | ||
| C6 D16 × D19 | M + F2 | CKX5+ | CKX5+ | spike number+ | yield+, CKX act.=, root=− |
| P + F2 | CKX5+ | nc | seed number+ | ||
| C7 D19 × KOH7 | M + F2 | CKX10+ | nc | nc | yield−, CKX act.+, root=−, semi-empty spikes++ |
| P + F2 | nc | nc | nc | ||
| C8 KOH7 × D19 | M + F2 | nc | CKX5−, CKX8−,NAC2− | nc | yield++, CKX act.=+, root= |
| P + F2 | CKX10+ | CKX5−, NAC2− | nc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szala, K.; Dmochowska-Boguta, M.; Bocian, J.; Orczyk, W.; Nadolska-Orczyk, A. Transgenerational Paternal Inheritance of TaCKX GFMs Expression Patterns Indicate a Way to Select Wheat Lines with Better Parameters for Yield-Related Traits. Int. J. Mol. Sci. 2023, 24, 8196. https://doi.org/10.3390/ijms24098196
Szala K, Dmochowska-Boguta M, Bocian J, Orczyk W, Nadolska-Orczyk A. Transgenerational Paternal Inheritance of TaCKX GFMs Expression Patterns Indicate a Way to Select Wheat Lines with Better Parameters for Yield-Related Traits. International Journal of Molecular Sciences. 2023; 24(9):8196. https://doi.org/10.3390/ijms24098196
Chicago/Turabian StyleSzala, Karolina, Marta Dmochowska-Boguta, Joanna Bocian, Waclaw Orczyk, and Anna Nadolska-Orczyk. 2023. "Transgenerational Paternal Inheritance of TaCKX GFMs Expression Patterns Indicate a Way to Select Wheat Lines with Better Parameters for Yield-Related Traits" International Journal of Molecular Sciences 24, no. 9: 8196. https://doi.org/10.3390/ijms24098196
APA StyleSzala, K., Dmochowska-Boguta, M., Bocian, J., Orczyk, W., & Nadolska-Orczyk, A. (2023). Transgenerational Paternal Inheritance of TaCKX GFMs Expression Patterns Indicate a Way to Select Wheat Lines with Better Parameters for Yield-Related Traits. International Journal of Molecular Sciences, 24(9), 8196. https://doi.org/10.3390/ijms24098196