

Palmelloid Formation and Cell Aggregation Are Essential Mechanisms for High Light Tolerance in a Natural Strain of Chlamydomonas reinhardtii
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Two Natural Chlamydomonas Strains Displayed Different Growth Variations under High Light
2.2. Variation in Photosynthetic Efficiency and Pigment Levels
2.3. Responses to Oxidative Stress and Strategies to Mitigate Oxidative Stress
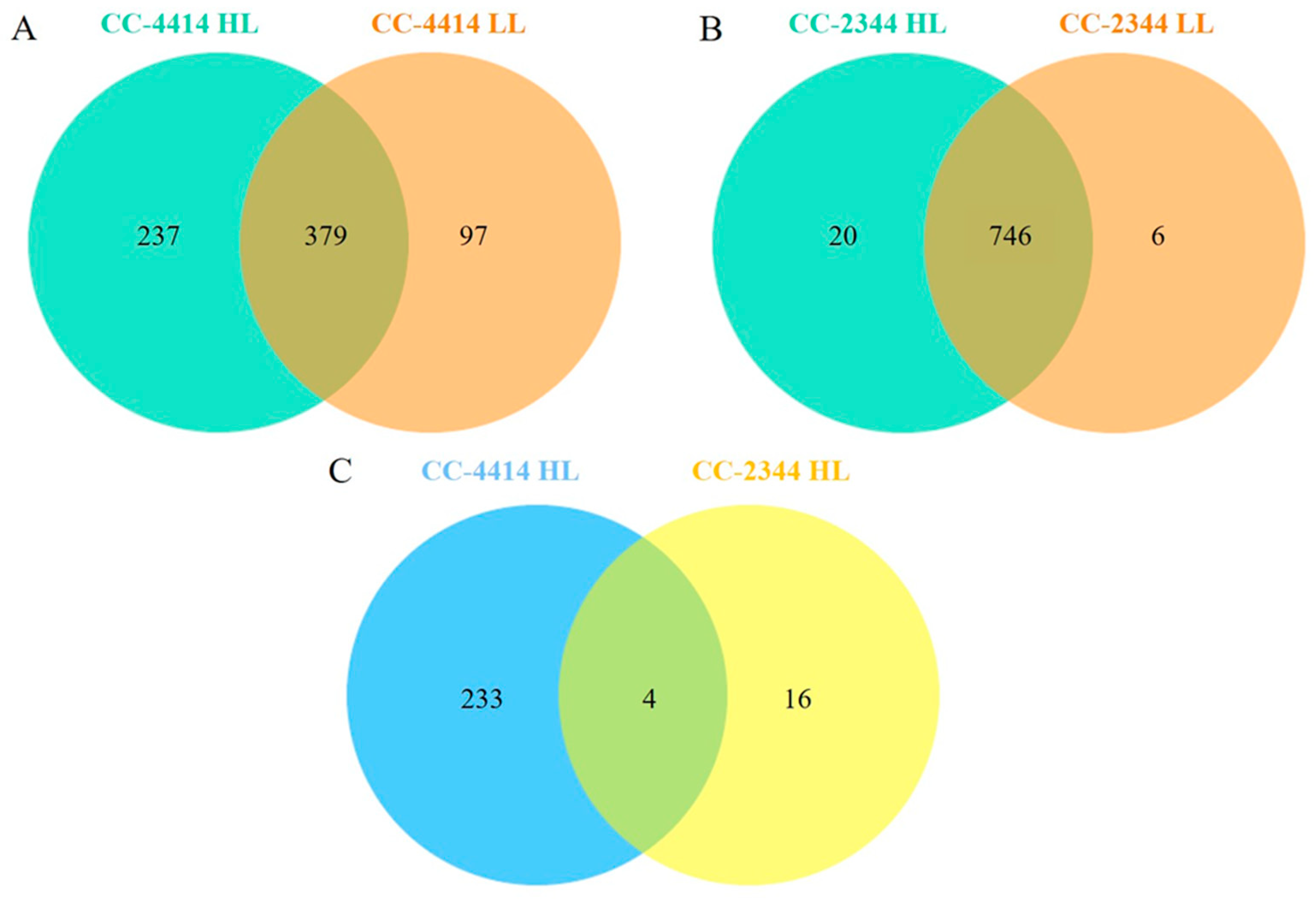
2.4. Proteome Analysis
2.5. Cell Morphology and Aggregation
3. Discussion
3.1. ROS Management
3.2. Protein Degradation by the Ubiquitin–Proteasome Pathway and Autophagy
3.3. Palmelloid Formation and Cell Clustering under Severe Stress
4. Materials and Methods
4.1. Chlamydomonas Reinhardtii Strains and Culture Conditions
4.2. Growth and Morphological Observation
4.3. Pigment Analysis
4.4. Lipid Peroxidation
4.5. Chlorophyll Fluorescence and NPQ Analysis
4.6. Superoxide Dismutase (SOD) Activity Assay
4.7. Proteome Analysis
4.7.1. Protein Extraction
4.7.2. Protein Digestion
4.7.3. Liquid Chromatography–Tandem Mass Spectrometry (LC/MS-MS)
4.7.4. Bioinformatics and Data Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niyogi, K.K. Photoprotection Revisited: Genetic and Molecular Approaches. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Tokutsu, R.; Li, A.; Takizawa, K.; Song, C.; Murata, K.; Yamasaki, T.; Liu, Z.; Minagawa, J.; Li, M. Structural basis of LhcbM5-mediated state transitions in green algae. Nat. Plants 2021, 7, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Nicol, L.; Nawrocki, W.J.; Croce, R. Disentangling the sites of non-photochemical quenching in vascular plants. Nat. Plants 2019, 5, 1177–1183. [Google Scholar] [CrossRef]
- Foyer, C.H.; Neukermans, J.; Queval, G.; Noctor, G.; Harbinson, J. Photosynthetic control of electron transport and the regulation of gene expression. J. Exp. Bot. 2012, 63, 1637–1661. [Google Scholar] [CrossRef]
- Blaby, I.K.; Glaesener, A.G.; Mettler, T.; Fitz-Gibbon, S.T.; Gallaher, S.D.; Liu, B.; Boyle, N.R.; Kropat, J.; Stitt, M.; Johnson, S.; et al. Systems-level analysis of nitrogen starvation-induced modifications of carbon metabolism in a Chlamydomonas reinhardtii starchless mutant. Plant Cell 2013, 25, 4305–4323. [Google Scholar] [CrossRef]
- Virtanen, O.; Khorobrykh, S.; Tyystjärvi, E. Acclimation of Chlamydomonas reinhardtii to extremely strong light. Photosynth. Res. 2021, 147, 91–106. [Google Scholar] [CrossRef]
- Saroussi, S.; Karns, D.A.J.; Thomas, D.C.; Bloszies, C.; Fiehn, O.; Posewitz, M.C.; Grossman, A.R. Alternative outlets for sustaining photosynthetic electron transport during dark-to-light transitions. Proc. Natl. Acad. Sci. USA 2019, 116, 11518–11527. [Google Scholar] [CrossRef]
- Renberg, L.; Johansson, A.I.; Shutova, T.; Stenlund, H.; Aksmann, A.; Raven, J.A.; Gardeström, P.; Moritz, T.; Samuelsson, G. A metabolomic approach to study major metabolite changes during acclimation to limiting CO2 in Chlamydomonas reinhardtii. Plant Physiol. 2010, 154, 187–196. [Google Scholar] [CrossRef]
- Schmollinger, S.; Mühlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing mechanisms in Chlamydomonas affect the transcriptome, the proteome, and photosynthetic metabolism. Plant Cell. 2014, 26, 1410–1435. [Google Scholar] [CrossRef]
- Meagher, E.; Rangsrikitphot, P.; Faridi, B.; Zamzam, G.; Durnford, D.G. Photoacclimation to high-light stress in Chlamydomonas reinhardtii during conditional senescence relies on generating pH-dependent, high-quenching centres. Plant Physiol. Biochem. 2021, 158, 136–145. [Google Scholar] [CrossRef]
- Morishita, J.; Tokutsu, R.; Minagawa, J.; Hisabori, T.; Wakabayashi, K. Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis. Plants 2021, 10, 1483. [Google Scholar] [CrossRef]
- Wang, F.; Qi, Y.; Malnoë, A.; Choquet, Y.; Wollman, F.; de Vitry, C. The High Light Response and Redox Control of Thylakoid FtsH Protease in Chlamydomonas reinhardtii. Mol. Plant 2016, 10, 99–114. [Google Scholar] [CrossRef]
- Nama, S.; Madireddi, S.K.; Yadav, R.M.; Subramanyam, R. Non-photochemical quenching-dependent acclimation and thylakoid organization of Chlamydomonas reinhardtii to high light stress. Photosynth. Res. 2019, 139, 387–400. [Google Scholar] [CrossRef]
- Roach, T.; Baur, T.; Stöggl, W.; Krieger-Liszkay, A. Chlamydomonas reinhardtii responding to high light: A role for 2-propenal (acrolein). Physiol. Plant 2017, 161, 75–87. [Google Scholar] [CrossRef]
- Shi, M.; Zhou, L.; Cui, M.; Ge, T.; Chen, F.; Zhao, L. Roles of Light-Harvesting Complex Stress-Related Proteins in the Stress Responses of Chlamydomonas. J. Plant Sci. 2022, 10, 1–11. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Glaesener, A.G.; Pellegrini, M.; Merchant, S.S. Chlamydomonas Genome Resource for Laboratory Strains Reveals a Mosaic of Sequence Variation, Identifies True Strain Histories, and Enables Strain-Specific Studies. Plant Cell 2015, 27, 2335–2352. [Google Scholar] [CrossRef]
- Schierenbeck, L.; Ries, D.; Rogge, K.; Grewe, S.; Weisshaar, B.; Kruse, O. Fast forward genetics to identify mutations causing a high light tolerant phenotype in Chlamydomonas reinhardtii by whole-genome-sequencing. BMC Genom. 2015, 16, 57. [Google Scholar] [CrossRef]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef]
- Flowers, J.M.; Hazzouri, K.M.; Pham, G.M.; Rosas, U.; Bahmani, T.; Khraiwesh, B.; Nelson, D.R.; Kenan, J.; Abdrabu, R.; Harris, E.H.; et al. Whole-Genome Resequencing Reveals Extensive Natural Variation in the Model Green Alga Chlamydomonas reinhardtii. Plant Cell 2015, 27, 2353–2369. [Google Scholar] [CrossRef]
- de Carpentier, F.; Lemaire, S.D.; Danon, A. When Unity Is Strength: The Strategies Used by Chlamydomonas to Survive Environmental Stresses. Cells 2019, 8, 1307. [Google Scholar] [CrossRef] [PubMed]
- Ballottari, M.; Dall’Osto, L.; Morosinotto, T.; Bassi, R. Contrasting behavior of higher plant photosystem I and II antenna systems during acclimation. J. Biol. Chem. 2007, 282, 8947–8958. [Google Scholar] [CrossRef] [PubMed]
- Kouřil, R.; Wientjes, E.; Bultema, J.B.; Croce, R.; Boekema, E.J. High-light vs. low-light: Effect of light acclimation on photosystem II composition and organization in Arabidopsis thaliana. Biochim. Biophys. Acta 2013, 1827, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Cleland, R.E.; Melis, A.; Neale, P.J. Mechanism of photoinhibition: Photochemical reaction center inactivation in system II of chloroplasts. Photosyn. Res. 1986, 9, 79–88. [Google Scholar] [CrossRef]
- Durnford, D.G.; Price, J.A.; McKim, S.M.; Sarchfield, M.L. Light-harvesting complex gene expression is controlled by both transcriptional and post-transcriptional mechanisms during photoacclimation in Chlamydomonas reinhardtii. Physiol. Plant. 2003, 118, 193–205. [Google Scholar] [CrossRef]
- Lüder, U.H.; Wiencke, C. Acclimation of photosynthesis and pigments during and after six months of darkness in Palmaria decipiens (Rhodophyta): A study to simulate antarctic winter sea ice cover. J. Phycol. 2002, 38, 904–913. [Google Scholar] [CrossRef]
- Cantrell, M.; Peers, G. A mutant of Chlamydomonas without LHCSR maintains high rates of photosynthesis, but has reduced cell division rates in sinusoidal light conditions. PLoS ONE 2017, 12, e0179395. [Google Scholar] [CrossRef]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef]
- Virtanen, O.; Valev, D.; Kruse, O.; Wobbe, L.; Tyystjärvi, E. Photoinhibition and continuous growth of the wild-type and a high-light tolerant strain of Chlamydomonas reinhardtii. Photosynthetica 2019, 57, 617–626. [Google Scholar] [CrossRef]
- de Carpentier, F.; Maes, A.; Marchand, C.H.; Chung, C.; Durand, C.; Crozet, P.; Lemaire, S.D.; Danon, A. How abiotic stress-induced socialization leads to the formation of massive aggregates in Chlamydomonas. Plant Physiol. 2022, 190, 1927–1940. [Google Scholar] [CrossRef]
- Avin-Wittenberg, T. Autophagy and its role in plant abiotic stress management. Plant Cell Environ. 2019, 42, 1045–1053. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Su, T.; Li, X.; Yang, M.; Shao, Q.; Zhao, Y.; Ma, C.; Wang, P. Autophagy: An Intracellular Degradation Pathway Regulating Plant Survival and Stress Response. Front. Plant Sci. 2020, 11, 164. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Tanaka, K.; Imamura, S. Microalgal Target of Rapamycin (TOR): A Central Regulatory Hub for Growth, Stress Response and Biomass Production. Plant Cell Physiol. 2020, 61, 675–684. [Google Scholar] [CrossRef]
- Goodenough, U.; Blaby, I.; Caserocd, D.; Gallaher, S.D.; Goodsona, C.; Johnsone, S.; Leef, J.; Merchantbd, S.S.; Pellegrini, M.; Rothg, R.; et al. The path to triacylglyceride obesity in the sta6 strain of Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 591–613. [Google Scholar] [CrossRef]
- Pérez-Martín, M.; Blaby-Haasb, C.E.; Pérez-Péreza, M.E.; Andrés-Garrido, A.; Blaby, I.K.; Merchantb, S.S.; Crespo, J.S. Activation of Autophagy by Metals in Chlamydomonas reinhardtii. Eukaryot. Cell 2015, 14, 964–973. [Google Scholar] [CrossRef]
- Tran, Q.G.; Cho, K.; Park, S.; Kim, U.; Lee, Y.; Kim, H. Impairment of starch biosynthesis results in elevated oxidative stress and autophagy activity in Chlamydomonas reinhardtii. Sci. Rep. 2019, 9, 9856. [Google Scholar] [CrossRef]
- Pérez-Pérez, M.E.; Lemaire, S.D.; Crespo, J.L. Reactive oxygen species and autophagy in plants and algae. Plant Physiol. 2012, 160, 156–164. [Google Scholar] [CrossRef]
- Smalle, J.; Vierstra, R.D. The ubiquitin 26S proteasome proteolytic pathway. Annu. Rev. Plant. Biol. 2004, 55, 555–590. [Google Scholar] [CrossRef]
- Budenholzer, L.; Cheng, C.; Li, Y.; Hochstrasser, M. Proteasome Structure and Assembly. J. Mol. Biol. 2017, 429, 3500–3524. [Google Scholar] [CrossRef]
- Walliwalagedara, C.; Keulen, H.; Willard, B.; Wei, R. Differential Proteome Analysis of Chlamydomonas reinhardtii Response to Arsenic Exposure. Am. J. Plant. Sci. 2012, 3, 764–772. [Google Scholar] [CrossRef]
- Heredia-Martínez, L.G.; Andrés-Garrido, A.; Martínez-Force, E.; Pérez-Pérez, M.E.; Crespo, J.L. Chloroplast Damage Induced by the Inhibition of Fatty Acid Synthesis Triggers Autophagy in Chlamydomonas. Plant Physiol. 2018, 178, 1112–1129. [Google Scholar] [CrossRef] [PubMed]
- Valledor, L.; Furuhashi, T.; Hanak, A.; Weckwerth, W. Systemic Cold Stress Adaptation of Chlamydomonas reinhardtii. Mol. Cell Proteom. 2013, 12, 2032–2047. [Google Scholar] [CrossRef] [PubMed]
- Jamers, A.; Ven, K.; Moens, L.; Robbens, J.; Potters, G.; Guisez, Y.; Blust, R.; Coen, W.D. Effect of copper exposure on gene expression profiles in Chlamydomonas reinhardtii based on microarray analysis. Aquat. Toxicol. 2006, 80, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K.J. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 2013, 79, 4774–4785. [Google Scholar] [CrossRef]
- Franzmann, T.; Alberti, S. Ubiquitin protein helps cells to recover from stress. Nature 2021, 597, 183–184. [Google Scholar] [CrossRef]
- Stone, S.L. Role of the Ubiquitin Proteasome System in Plant Response to Abiotic Stress. Int. Rev. Cell. Mol. Biol. 2019, 343, 65–110. [Google Scholar]
- Lurling, M.; Beekman, W. Palmelloids formation in Chlamydomonas reinhardtii: Defence against rotifer predators? Ann. Limnol. 2006, 42, 65–72. [Google Scholar] [CrossRef]
- Iwasa, K.; Mukakami, S. Palinelloid Foriiiation of Chlatnydomonas I. Palinelloid Incluction by Organic Acids. Physiol. Plant. 1968, 21, 1224–1233. [Google Scholar] [CrossRef]
- Iwasa, K.; Murakami, S. Paliiielloid Formation of Chlamydotnonas II. Mechanism of Palmelloid Formation by Organic Acid. Physiol. Plant. 1969, 22, 43–50. [Google Scholar] [CrossRef]
- Olsen, Y.; Knutsen, G.; Lien, T. Characteristics of Phosphorus Limitation in Chlamydomonas reinhardtii (chlorophyceae) and Its Palmelloids. J. Phycol. 1983, 19, 313–319. [Google Scholar] [CrossRef]
- Samadani, M.; Dewez, D. Cadmium accumulation and toxicity affect the extracytoplasmic polyphosphate level in Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 2018, 166, 200–206. [Google Scholar] [CrossRef]
- Takouridis, S.J.; Tribe, D.E.; Gras, S.L.; Martin, J. The selective breeding of the freshwater microalga Chlamydomonas reinhardtii for growth in salinity. Bioresour. Technol. 2015, 184, 18–22. [Google Scholar] [CrossRef]
- Khona, D.K.; Shirolikar, S.M.; Gawde, K.K.; Hom, E.; Deodhar, M.A.; D’Souza, J.S. Characterization of salt stress-induced palmelloids in the green alga, Chlamydomonas reinhardtii. Algal Res. 2016, 16, 434–448. [Google Scholar] [CrossRef]
- Neelam, S.; Subramanyam, R. Alteration of photochemistry and protein degradation of photosystem II from Chlamydomonas reinhardtii under high salt grown cells. J. Photochem. Photobiol. B 2013, 124, 63–70. [Google Scholar] [CrossRef]
- Visviki, I.; Santikul, D. The pH tolerance of Chlamydomonas applanata (Volvocales, Chlorophyta). Arch. Environ. Contam. Toxicol. 2000, 38, 147–151. [Google Scholar] [CrossRef]
- Nakamura, K.; Bray, D.F.; Wagenaar, E.B. Ultrastructure of Chlamydomonas eugametos Palmelloids Induced by Chloroplatinic Acid Treatment. J. Bacteriol. 1975, 121, 338–343. [Google Scholar] [CrossRef]
- Nakamura, K.; Sakon, M.; Hatanaka, M.K. Chemical Factors Affecting Palmelloid-Forming Activity of Chloroplatinic Acid on Chlamydontonas eugametos. Physiol. Plant. 1976, 36, 293–296. [Google Scholar] [CrossRef]
- Karsten, U.; Karsten, U.; Lembcke, S.; Schumann, R. The effects of ultraviolet radiation on photosynthetic performance, growth and sunscreen compounds in aeroterrestrial biofilm algae isolated from building facades. Planta 2007, 225, 991–1000. [Google Scholar] [CrossRef]
- Karsten, U.; Holzinger, A. Green algae in alpine biological soil crust communities: Acclimation strategies against ultraviolet radiation and dehydration. Biodivers. Conserv. 2014, 23, 1845–1858. [Google Scholar] [CrossRef]
- Li, Y.; Jones, L.; McQueen-Mason, S. Expansins and cell growth. Curr. Opin. Plant. Biol. 2003, 6, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Sorimachi, H. Calpains: An elaborate proteolytic system. Biochim. Biophys. Acta 2012, 1824, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Zheng, J.J. PDZ domains and their binding partners: Structure, specificity, and modification. Cell Commun. Signal. 2010, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Szyszka-Mroz, B.; Ivanov, A.G.; Trick, C.G.; Hüner, N.P. Palmelloid formation in the Antarctic psychrophile, Chlamydomonas priscuii, is photoprotective. Front. Plant Sci. 2022, 13, 911035. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, S.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Melis, A.; Spangfort, M.; Andersson, B. Light-absorption and electron transport balance between photosystem II and Photosystem I in spinach chloroplasts. Photochem. Photobiol. 1987, 45, 129–136. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. BioTechniques 2003, 34, 374–378. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder1, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, 362–368. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniprotID | Function/Component | Protein Name | Peptide | p-Value | Fold Change |
|---|---|---|---|---|---|
| A0A7S0YFV0 | Proteolysis/cell membrane | Calpain catalytic domain-containing protein | LNPKIAR | 0.00 | 20.58 |
| A0A7S3QLH0 | PDZ domain-containing protein | VLLEALQK | 0.05 | 15.66 | |
| A0A835WE54 | SUMO transferase activity | SAP domain-containing protein | LLELFEDSYHLVSNGSVPRDMWR | 0.02 | 20.05 |
| A0A0S2LQ22 | Photosynthesis/chloroplast thylakoid membrane | Cytochrome b6-f complex subunit 6 | VVKLM | 0.01 | 19.37 |
| A0A250X4U9 | ATP binding | Helicase ATP-binding domain-containing protein | STGGGGGGGGKGGR | 0.01 | 19.05 |
| A0A250X4T6 | Protein phosphorylation/cell membrane | Protein kinase domain-containing protein | EGSGMIK | 0.01 | 19.06 |
| A0A835WI51 | Nucleus | Helicase C-terminal domain-containing protein | AAARDAAAAAR | 0.01 | 17.79 |
| A0A835SUR5 | CRC domain-containing protein | AGAKGGGAAAASGAPGGGR | 0.01 | 17.07 | |
| A0A150G407 | TFIIS N-terminal domain-containing protein | VLGGM | 0.00 | 14.96 | |
| A0A6A0AIR0 | Plant-type cell wall organization | Expansin-like EG45 domain-containing protein | GIQPG | 0.01 | 17.35 |
| A0A6A0AK22 | Ubiquitin-protein transferase activity | RBR-type E3 ubiquitin transferase | GYVNNAIANGPASLDLRCPTPK | 0.00 | 17.25 |
| A0A172WYN8 | mRNA processing/nucleus | Mac1 | MMHSLHARVETAR | 0.01 | 16.96 |
| A0A2J8AGW4 | E3 ubiquitin-protein ligase | GPVAAPR | 0.00 | 16.94 | |
| A0A835SR56 | Alternative oxidase activity | Ubiquinol oxidase | TMKACQDETVGQDIISR | 0.01 | 16.93 |
| Q6PLP9 | Protein deubiquitination | Ubiquitin carboxyl-terminal hydrolase | FNLMALVGNRADIYSSR | 0.00 | 16.90 |
| A0A835XUN6 | DNA replication/nucleus | Sld5 domain-containing protein | RGMTTFSLPEIYHER | 0.00 | 16.87 |
| A0A250XGZ4 | RNA binding | mRNA cap-binding protein | DRYTV | 0.01 | 16.80 |
| A0A150G279 | Protein dephosphorylation | Protein tyrosine phosphatase | FVFDSK | 0.00 | 16.26 |
| A0A7S3VKA6 | Metalloendopeptidase activity/nucleus | SprT-like domain-containing protein | KPQPADCRGR | 0.01 | 16.23 |
| A0A150GXF6 | Guanyl-nucleotide exchange factor activity/cytosol | SEC7 domain-containing protein | MSAEQVYHPAAEIVAIALR | 0.00 | 15.32 |
| A0A2K3CUA0 | mRNA 5′-splice site recognition | Pre-mRNA-processing factor 39 | KEEAAAEGAEGAEVK | 0.00 | 14.64 |
| A0A7S0VD46 | Protein folding/unfolded protein binding | Prefoldin subunit 2 | KPASQGVLV | 0.05 | 13.29 |
| UniprotID | Function/Component | Protein Name | Peptide | p-Value | Fold Change | |
|---|---|---|---|---|---|---|
| CC-4414 | CC-2344 | |||||
| A0A7S0YJX0 | Unknown | Uncharacterized protein | PGFGGGER | 0.02 | 18.82 | 17.34 |
| A0A2J7ZZC2 | Adenylate kinase activity | Adenylate kinase | EGMTVVTR | 0.01 | 16.60 | 19.50 |
| A0A2K3DSM2 | Potassium:proton antiporter activity/plasma membrane | Na_H_Exchanger domain-containing protein | GVGGMGVGMGGGAAGTGPGGGGGGGGR | 0.00 | 15.63 | 17.68 |
| A8JFC7 | Unknown | Copper homeostasis protein cutC homolog | VELCAALIEGGITPSAGMIR | 0.01 | 15.61 | 19.46 |
| UniprotID | Function/Component | Protein Name | Peptide | p-Value | Fold Change |
|---|---|---|---|---|---|
| A0A835XQZ1 | Membrane | Basic proline-rich protein-like | ASLGVLLSSGAGGGGGGKGAGAK | 0.01 | 26.81 |
| A0A835SW25 | ATP binding/membrane | Protein kinase domain-containing protein | SSSNR | 0.01 | 20.93 |
| A0A2K3CS73 | Unknown | DUF4604 domain-containing protein | GGLRTAQGGGGAGK | 0.04 | 20.31 |
| A0A2J8A5D8 | Nucleic acid binding | CCHC-type domain-containing protein | VLLPPSK | 0.01 | 19.93 |
| D8U4G9 | Unknown | ANK_REP_REGION domain-containing protein | VPGLAGALKR | 0.01 | 19.75 |
| A0A250X3Y5 | Unknown | DUF4378 domain-containing protein | MSGGG | 0.00 | 19.72 |
| A0A835Y1G5 | Unknown | Protein kinase domain-containing protein | AATLPRAAAPNAAAK | 0.00 | 19.14 |
| A0A250XQV9 | Unknown | Uncharacterized protein | SSTATR | 0.01 | 18.16 |
| D8UA57 | Unknown | DUF4485 domain-containing protein | DAIGSQRGAK | 0.01 | 18.03 |
| A0A835WLJ1 | Sphingomyelin phosphodiesterase activity | ANK_REP_REGION domain-containing protein | AAGPGAGAGADVAR | 0.00 | 17.95 |
| A0A2K3DJS2 | Unknown | CobW C-terminal domain-containing protein | APAPAPGPASGPAPAPGPASGSAGMAAMCAEERR | 0.00 | 17.77 |
| Q8WKX7 | Chloroplast/ endonuclease activity | LAGLIDADG_2 domain-containing protein | MIIKSGEK | 0.01 | 17.50 |
| A0A1W5IVS0 | Unknown | RlsD | GSHDGVGR | 0.00 | 17.30 |
| A0A835T9S6 | Phosphorus-oxygen lyase activity | Guanylate cyclase domain-containing protein | RTGAPVQQHR | 0.01 | 16.89 |
| A0A2K3CW68 | Guanylate cyclase activity/plasma membrane | Guanylate cyclase domain-containing protein | TRIQPVLR | 0.01 | 16.20 |
| A0A2K3CPC2 | Unknown | GSCFA domain-containing protein | DPGLPSECVGR | 0.04 | 15.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suwannachuen, N.; Leetanasaksakul, K.; Roytrakul, S.; Phaonakrop, N.; Thaisakun, S.; Roongsattham, P.; Jantasuriyarat, C.; Sanevas, N.; Sirikhachornkit, A. Palmelloid Formation and Cell Aggregation Are Essential Mechanisms for High Light Tolerance in a Natural Strain of Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2023, 24, 8374. https://doi.org/10.3390/ijms24098374
Suwannachuen N, Leetanasaksakul K, Roytrakul S, Phaonakrop N, Thaisakun S, Roongsattham P, Jantasuriyarat C, Sanevas N, Sirikhachornkit A. Palmelloid Formation and Cell Aggregation Are Essential Mechanisms for High Light Tolerance in a Natural Strain of Chlamydomonas reinhardtii. International Journal of Molecular Sciences. 2023; 24(9):8374. https://doi.org/10.3390/ijms24098374
Chicago/Turabian StyleSuwannachuen, Nittaya, Kantinan Leetanasaksakul, Sittiruk Roytrakul, Narumon Phaonakrop, Siriwan Thaisakun, Peerapat Roongsattham, Chatchawan Jantasuriyarat, Nuttha Sanevas, and Anchalee Sirikhachornkit. 2023. "Palmelloid Formation and Cell Aggregation Are Essential Mechanisms for High Light Tolerance in a Natural Strain of Chlamydomonas reinhardtii" International Journal of Molecular Sciences 24, no. 9: 8374. https://doi.org/10.3390/ijms24098374
APA StyleSuwannachuen, N., Leetanasaksakul, K., Roytrakul, S., Phaonakrop, N., Thaisakun, S., Roongsattham, P., Jantasuriyarat, C., Sanevas, N., & Sirikhachornkit, A. (2023). Palmelloid Formation and Cell Aggregation Are Essential Mechanisms for High Light Tolerance in a Natural Strain of Chlamydomonas reinhardtii. International Journal of Molecular Sciences, 24(9), 8374. https://doi.org/10.3390/ijms24098374