


Identification of Photosynthesis Characteristics and Chlorophyll Metabolism in Leaves of Citrus Cultivar (Harumi) with Varying Degrees of Chlorosis

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
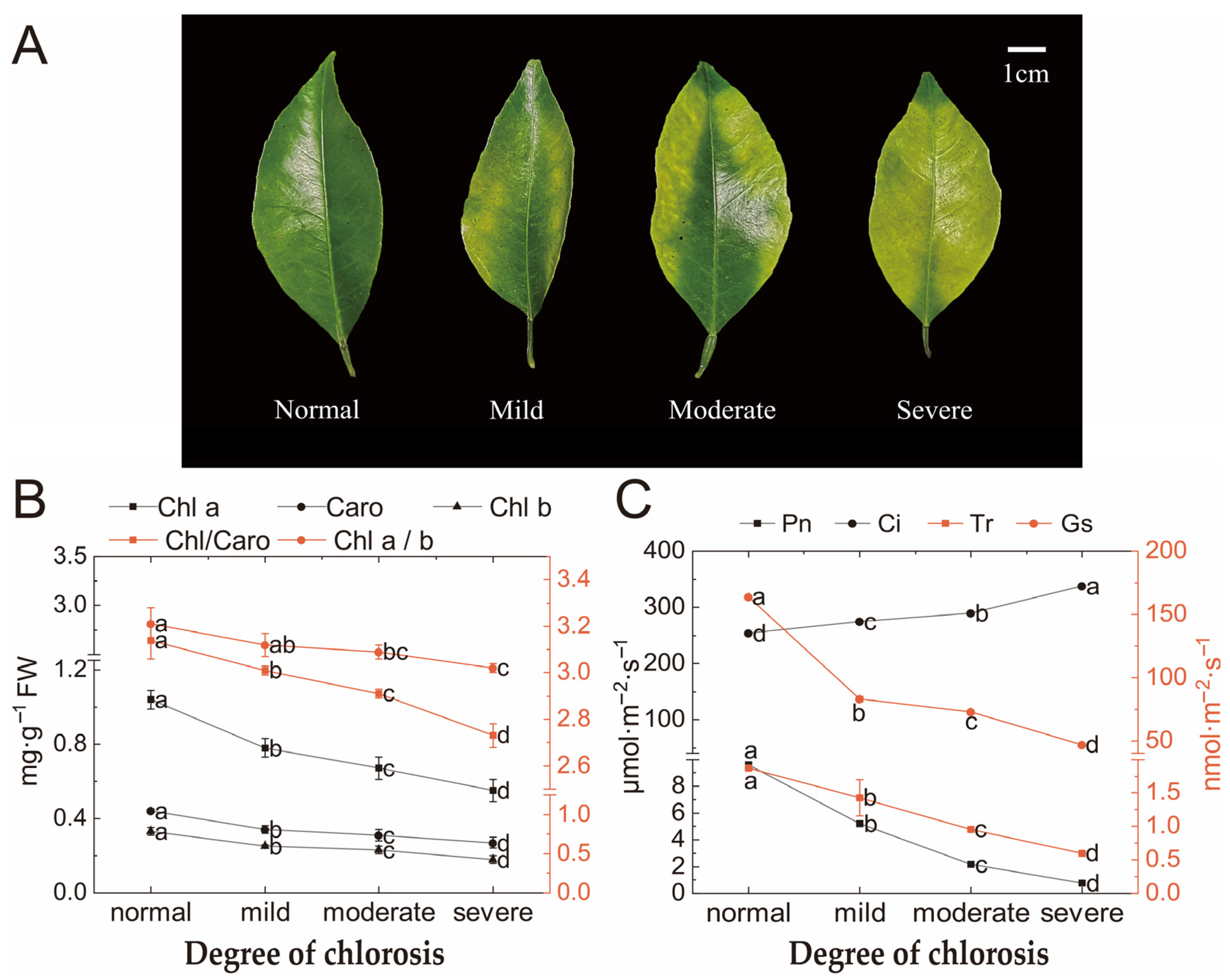
2.1. Photosynthetic Pigments and Photosynthetic Parameters
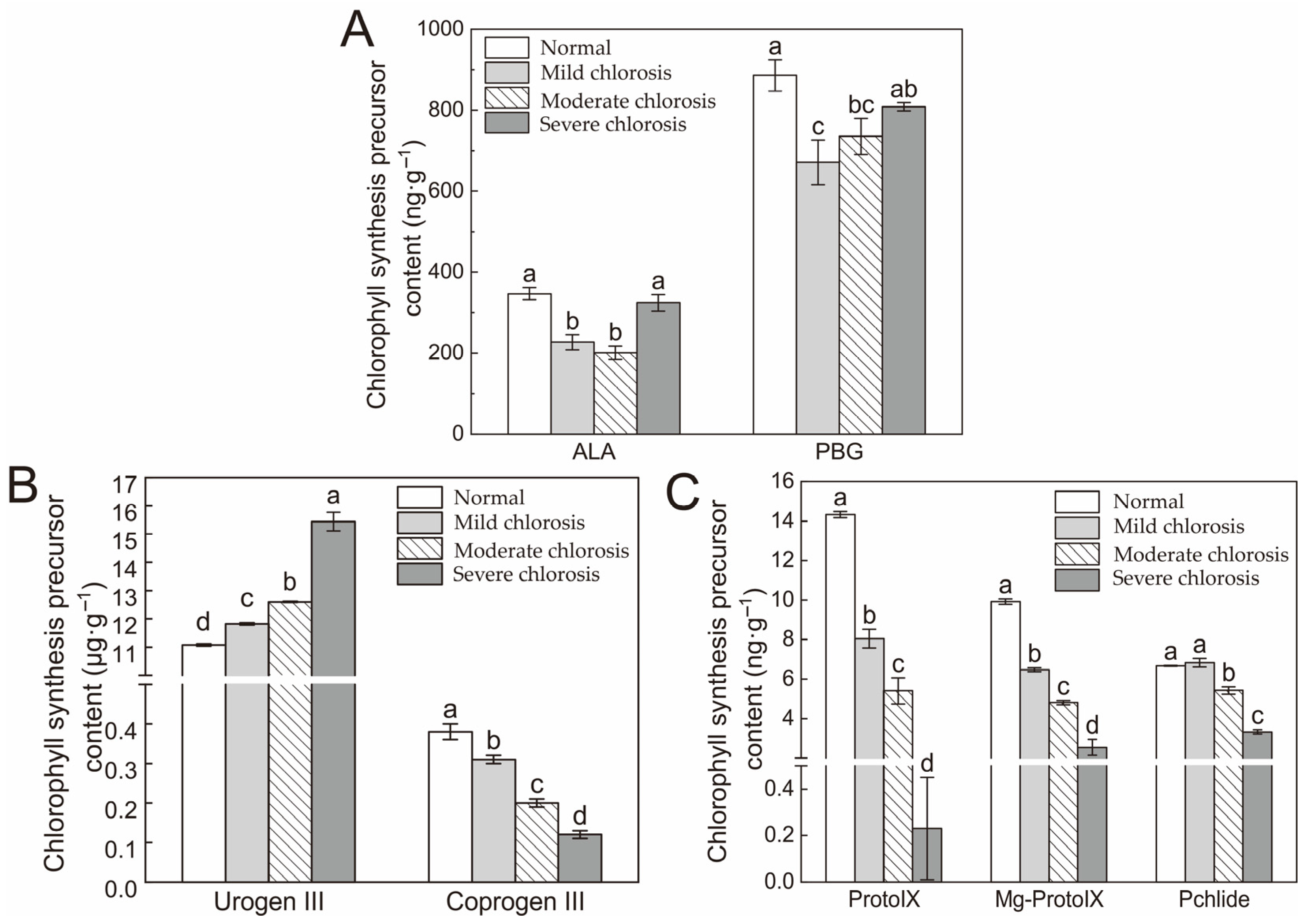
2.2. Chlorophyll Synthesis Precursors
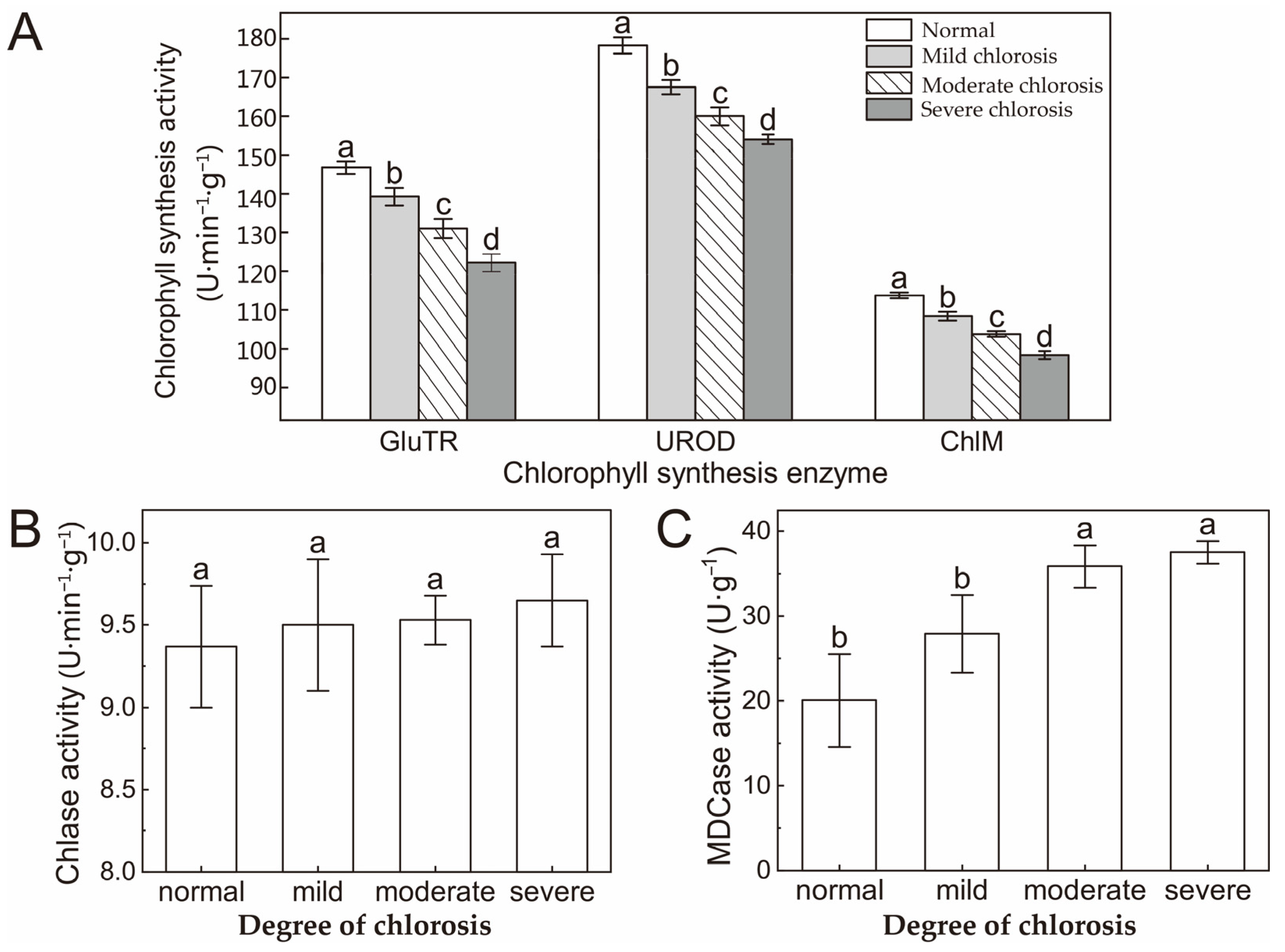
2.3. Activity of Chlorophyll Synthesis and Degradation Enzyme
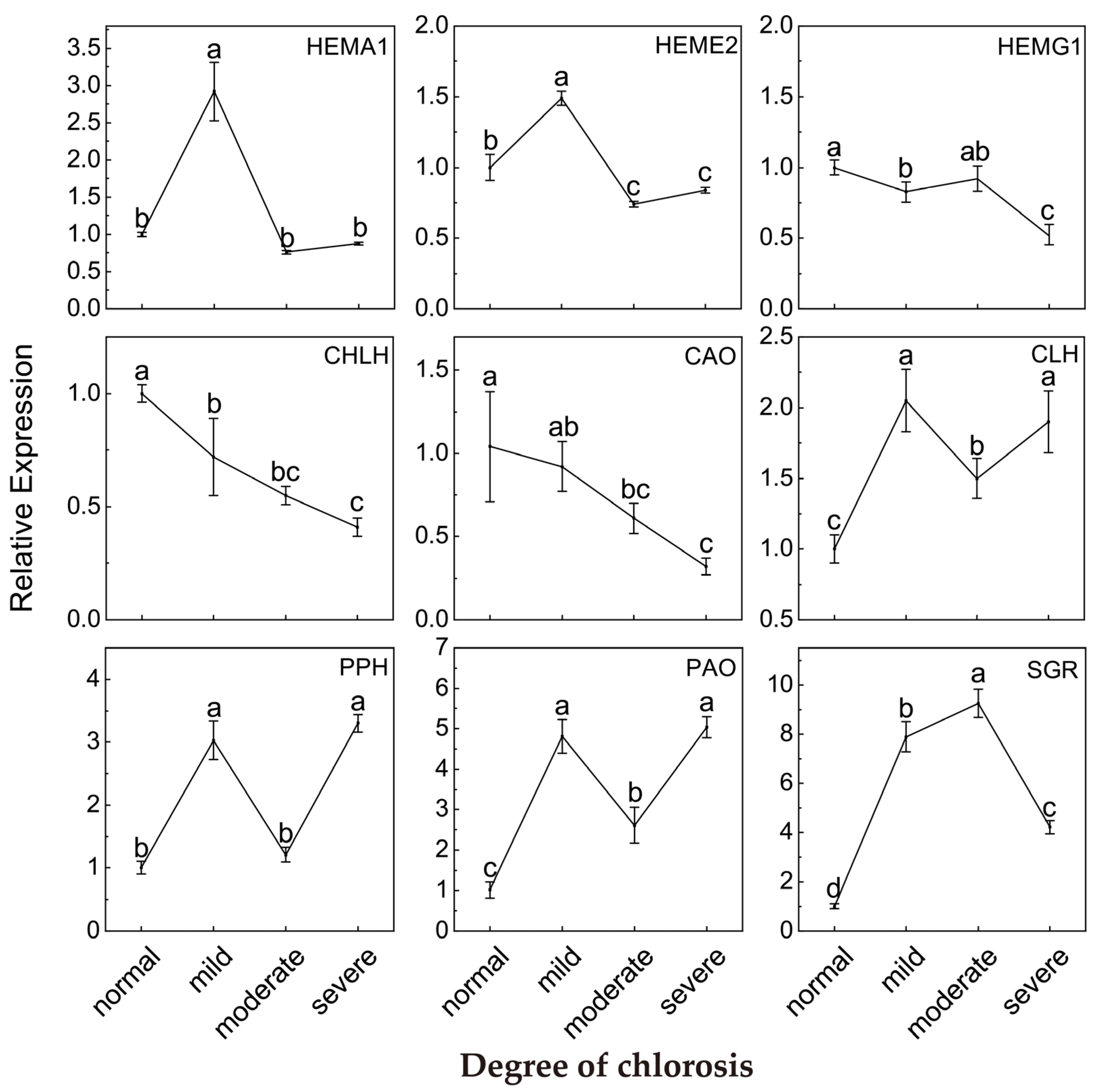
2.4. Relative Expression of Genes Involved in Chlorophyll Metabolism Pathway
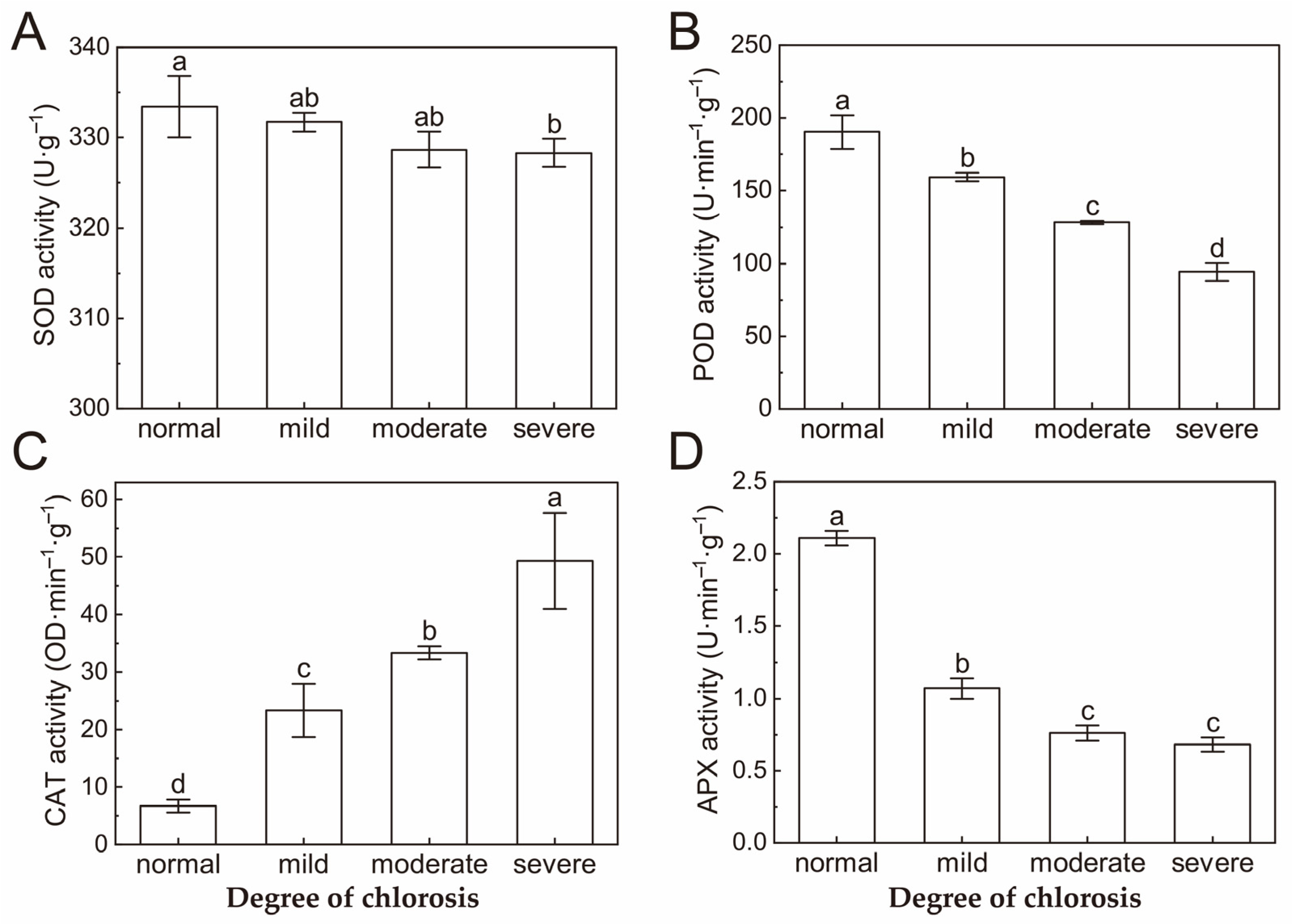
2.5. Antioxidant Enzyme Activity
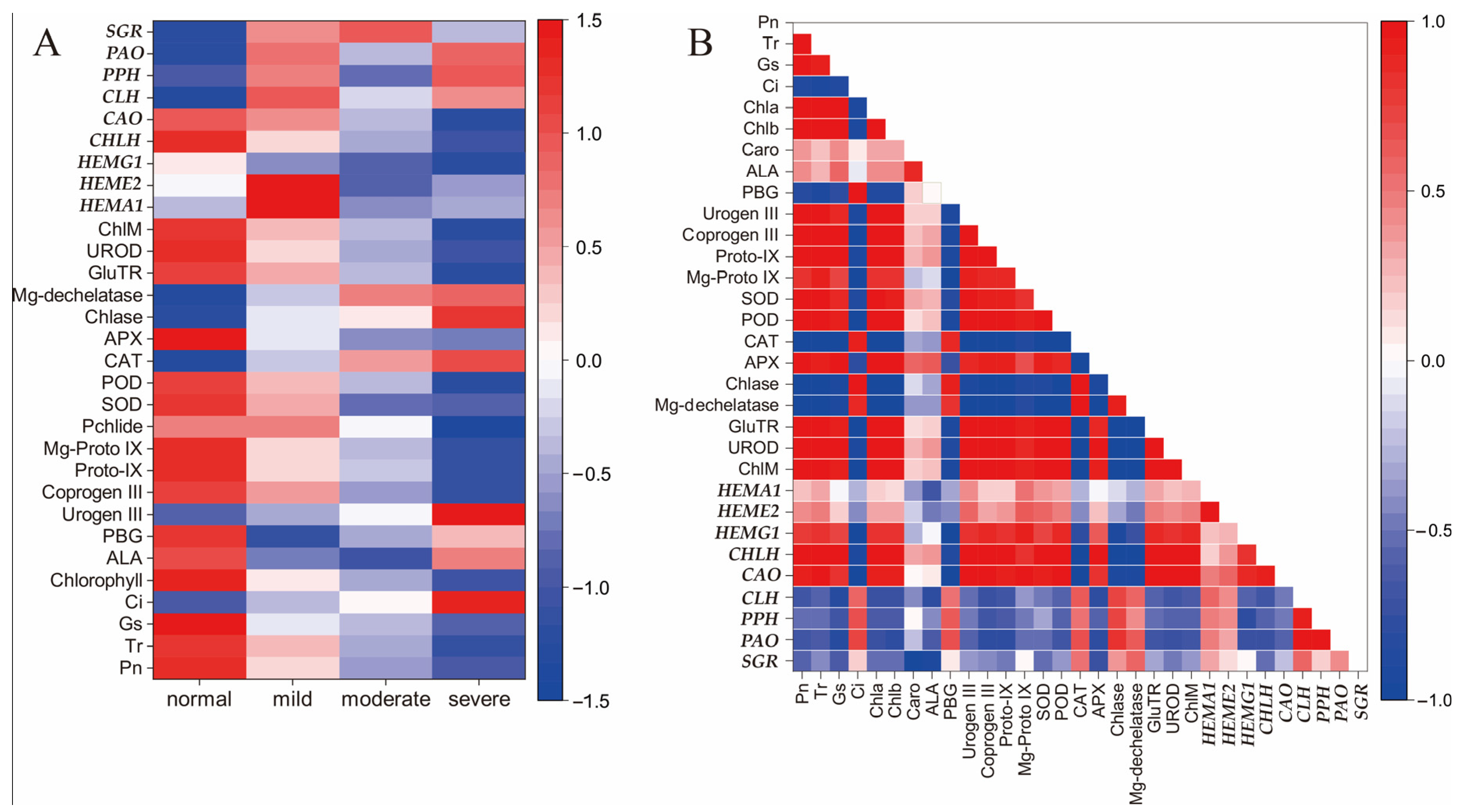
2.6. Correlation Analysis
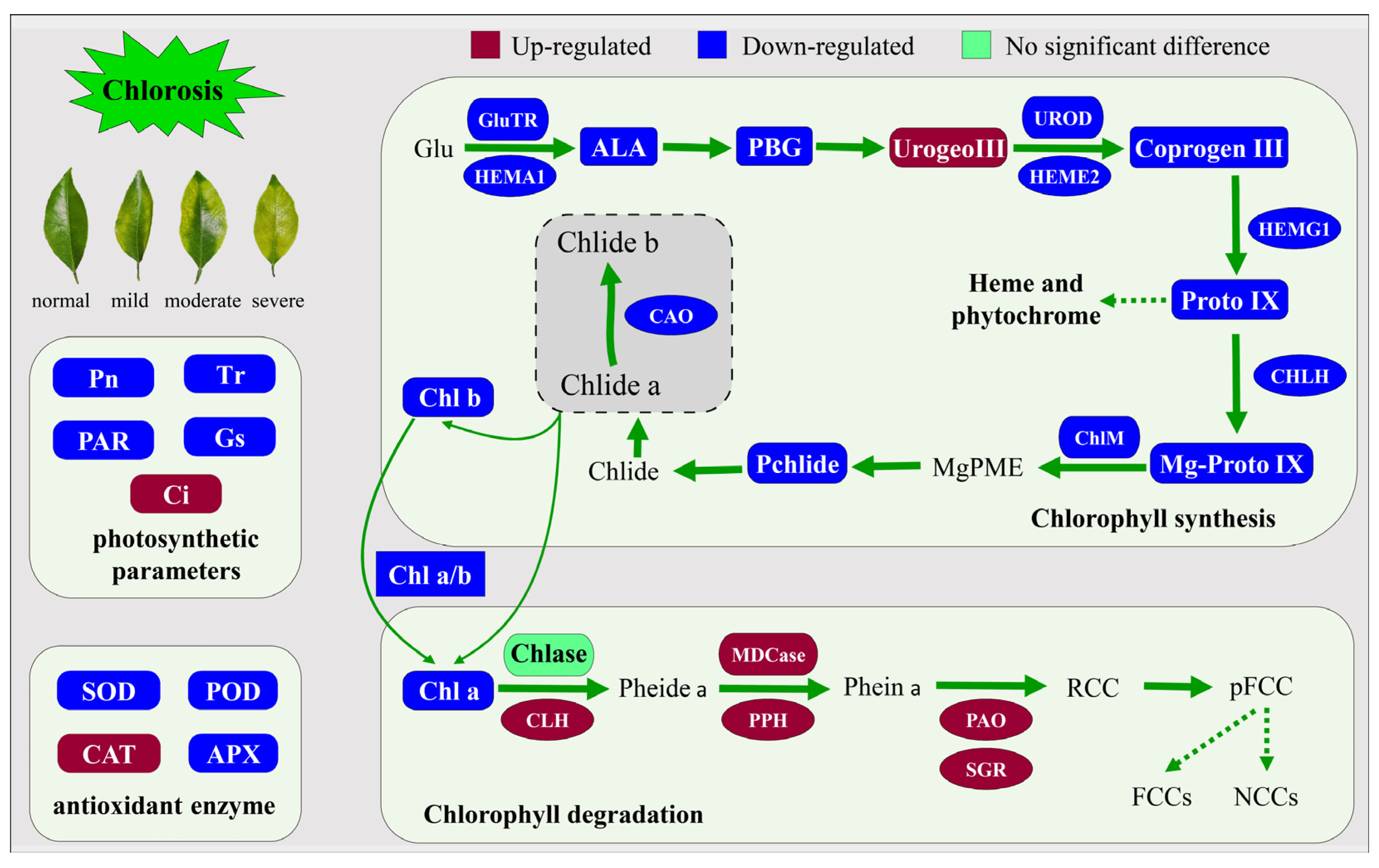
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatment
4.2. Determination of Photosynthetic Parameters
4.3. Determination of Photosynthetic Pigments
4.4. Determination of Chlorophyll Synthesis Precursor Substance Content
4.5. Determination of Antioxidant Enzyme Activity
4.6. Determination of Enzyme Activity Related to Chlorophyll Synthesis
4.7. Determination of the Chlase (Chlorophyllase) and Mg-Dechelatase Activity
4.7.1. Enzyme Extraction
4.7.2. Preparation of Substrates
4.7.3. Chlorophyllase Activity
4.7.4. Mg-Dechelatase Activity
4.7.5. Detection of Related Gene Expression in Chlorophyll Metabolism Pathway
4.8. Correlation and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naseer, M.A.U.R.; Mehdi, M.; Ashfaq, M.; Hassan, S.; Abid, M. Effect of Marketing Channel Choice on the Profitability of Citrus Farmers: Evidence Form Punjab-Pakistan. Pak. J. Agric. Sci. 2019, 56, 1003–1011. [Google Scholar]
- Sun, L.; Nasrullah; Ke, F.; Nie, Z.; Wang, P.; Xu, J. Citrus Genetic Engineering for Disease Resistance: Past, Present and Future. Int. J. Mol. Sci. 2019, 20, 5256. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhao, C.; Shi, H.; Liao, Y.; Xu, F.; Du, H.; Xiao, H.; Zheng, J. Nutrients and bioactives in citrus fruits: Different citrus varieties, fruit parts, and growth stages. Crit. Rev. Food Sci. Nutr. 2021, 63, 2018–2041. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef] [PubMed]
- Takishita, F.; Nishikawa, F.; Matsumoto, H.; Kato, M. Fruit Thinning and Physiological Disorders in Citrus Variety ‘Harumi’. Rev. Agric. Sci. 2021, 9, 20–31. [Google Scholar] [CrossRef]
- Liao, L.; Li, Y.; Bi, X.; Xiong, B.; Wang, X.; Deng, H.; Zhang, M.; Sun, G.; Jin, Z.; Huang, Z.; et al. Transcriptome analysis of Harumi tangor fruits: Insights into interstock-mediated fruit quality. Front. Plant Sci. 2022, 13, 995913. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, Å. The Plastid Development in Various Types of Chlorophyll Mutations. Hereditas 1942, 28, 483–492. [Google Scholar] [CrossRef]
- Shalygo, N.; Czarnecki, O.; Peter, E.; Grimm, B. Expression of chlorophyll synthase is also involved in feedback-contro l of chlorophyll biosynthesis. Plant Mol. Biol. 2009, 71, 425–436. [Google Scholar] [CrossRef]
- Qiu, K.; Li, Z.; Yang, Z.; Chen, J.; Wu, S.; Zhu, X.; Gao, S.; Gao, J.; Ren, G.; Kuai, B.; et al. EIN3 and ORE1 Accelerate Degreening during Ethylene-Mediated Leaf Sene scence by Directly Activating Chlorophyll Catabolic Genes in Arabidops is. PLoS Genet. 2015, 11, e1005399. [Google Scholar] [CrossRef]
- Sandhu, D.; Coleman, Z.; Atkinson, T.; Rai, K.M.; Mendu, V. Genetics and Physiology of the Nuclearly Inherited Yellow Foliar Mutan ts in Soybean. Front. Plant Sci. 2018, 9, 471. [Google Scholar] [CrossRef]
- Khan, I.; Zada, A.; Jia, T.; Hu, X. Effect of the Enhanced Production of Chlorophyll b on the Light Acclimation of Tomato. Int. J. Mol. Sci. 2023, 24, 3377. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Gu, T.; Khan, I.; Zada, A.; Jia, T. Research Progress in the Interconversion, Turnover and Degradation of Chlorophyll. Cells 2021, 10, 3134. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Kagawa, T.; Oikawa, K.; Suetsugu, N.; Miyao, M.; Wada, M. Chloroplast avoidance movement reduces photodamage in plants. Nature 2002, 420, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Rissler, H.M.; Durnford, D.G. Isolation of a novel carotenoid-rich protein in Cyanophora paradoxa that is immunologically related to the light-harvesting complexes of photosynthetic eukaryotes. Plant Cell Physiol. 2005, 46, 416–424. [Google Scholar] [CrossRef]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Et Biophys. Acta-Bioenerg. 2011, 1807, 968–976. [Google Scholar] [CrossRef]
- Hortensteiner, S. Update on the biochemistry of chlorophyll breakdown. Plant Mol. Biol. 2013, 82, 505–517. [Google Scholar] [CrossRef] [PubMed]
- op den Camp, R.G.; Przybyla, D.; Ochsenbein, C.; Laloi, C.; Kim, C.; Danon, A.; Wagner, D.; Hideg, E.; Gobel, C.; Feussner, I.; et al. Rapid induction of distinct stress responses after the release of singlet oxygen in Arabidopsis. Plant Cell 2003, 15, 2320–2332. [Google Scholar] [CrossRef]
- Mur, L.A.; Aubry, S.; Mondhe, M.; Kingston-Smith, A.; Gallagher, J.; Timms-Taravella, E.; James, C.; Papp, I.; Hortensteiner, S.; Thomas, H.; et al. Accumulation of chlorophyll catabolites photosensitizes the hypersensitive response elicited by Pseudomonas syringae in Arabidopsis. New Phytol. 2010, 188, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Hauenstein, M.; Hortensteiner, S.; Aubry, S. Side-chain modifications of phyllobilins may not be essential for chlorophyll degradation in Arabidopsis. Plant Direct 2022, 6, e441. [Google Scholar] [CrossRef]
- Yamatani, H.; Ito, T.; Nishimura, K.; Yamada, T.; Sakamoto, W.; Kusaba, M. Genetic analysis of chlorophyll synthesis and degradation regulated by BALANCE of CHLOROPHYLL METABOLISM. Plant Physiol. 2022, 189, 419–432. [Google Scholar] [CrossRef]
- Schumacher, I.; Menghini, D.; Ovinnikov, S.; Hauenstein, M.; Fankhauser, N.; Zipfel, C.; Hortensteiner, S.; Aubry, S. Evolution of chlorophyll degradation is associated with plant transition to land. Plant J. 2022, 109, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Meng, F.; Mo, F.; Qi, H.; Wang, P.; Chen, X.; Liu, J.; Ghanizadeh, H.; Zhang, H.; Wang, A. Slym1 control the color etiolation of leaves by facilitating the decomposition of chlorophyll in tomato. Plant Sci. 2022, 324, 111457. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morita, R.; Nishimura, M.; Yamaguchi, H.; Kusaba, M. Mendel’s green cotyledon gene encodes a positive regulator of the chlorophyll-degrading pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 14169–14174. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Moreau, C.; Chinoy, C.; Spanner, R.; Dalmais, M.; Le Signor, C.; Bendahmane, A.; Klenell, M.; Domoney, C. SGRL can regulate chlorophyll metabolism and contributes to normal plant growth and development in Pisum sativum L. Plant Mol. Biol. 2015, 89, 539–558. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Yu, Q.; Williams-Carrier, R.; Maliga, P.; Barkan, A. Engineered PPR proteins as inducible switches to activate the expression of chloroplast transgenes. Nat. Plants 2019, 5, 505–511. [Google Scholar] [CrossRef]
- Armarego-Marriott, T.; Sandoval-Ibanez, O.; Kowalewska, L. Beyond the darkness: Recent lessons from etiolation and de- etiolation studies. J. Exp. Bot. 2020, 71, 1215–1225. [Google Scholar] [CrossRef]
- Hirashima, M.; Tanaka, R.; Tanaka, A. Light-independent cell death induced by accumulation of pheophorbide a in Arabidopsis thaliana. Plant Cell Physiol. 2009, 50, 719–729. [Google Scholar] [CrossRef]
- Wuthrich, K.L.; Bovet, L.; Hunziker, P.E.; Donnison, I.S.; Hortensteiner, S. Molecular cloning, functional expression and characterisation of RCC reductase involved in chlorophyll catabolism. Plant J. 2000, 21, 189–198. [Google Scholar] [CrossRef]
- Hortensteiner, S.; Krautler, B. Chlorophyll breakdown in higher plants. Biochim. Biophys. Acta 2011, 1807, 977–988. [Google Scholar] [CrossRef]
- Yamasato, A.; Nagata, N.; Tanaka, R.; Tanaka, A. The N-terminal domain of chlorophyllide a oxygenase confers protein instability in response to chlorophyll B accumulation in Arabidopsis. Plant Cell 2005, 17, 1585–1597. [Google Scholar] [CrossRef]
- Collignon, L.N.; Normand, C.B. Photobiology: Principles, Applications, and Effects; Nova Science Publishers, Inc.: New York, NY, USA, 2011. [Google Scholar]
- Zeiger, E.; Field, C. Photocontrol of the Functional Coupling between Photosynthesis and Stomatal Conductance in the Intact Leaf: Blue Light and Par-Dependent Photosystems in Guard Cells. Plant Physiol. 1982, 70, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Hu, P.; Lin, M.; Ye, X.; Chen, L.; Huang, Z. Photosynthetic Characteristics and Chloroplast Ultrastructure Responses of Citrus Leaves to Copper Toxicity Induced by Bordeaux Mixture in Greenhouse. Int. J. Mol. Sci. 2022, 23, 9835. [Google Scholar] [CrossRef] [PubMed]
- Adams Iii, W.; Demmig-Adams, B.; Garab, G.; Govindjee. Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria. In Advances in Photosynthesis and Respiration, Including Bioenergy and Related Processes, 1st ed.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Harpaz-Saad, S.; Azoulay, T.; Arazi, T.; Ben-Yaakov, E.; Mett, A.; Shiboleth, Y.M.; Hörtensteiner, S.; Gidoni, D.; Gal-On, A.; Goldschmidt, E.E.; et al. Chlorophyllase is a rate-limiting enzyme in chlorophyll catabolism and is posttranslationally regulated. Plant Cell 2007, 19, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhang, M.; Bai, L.; Han, X.; Ge, Y.; Wang, W.; Li, J. Effects of 1-methylcyclopropene (1-MCP) on the expression of genes involved in the chlorophyll degradation pathway of apple fruit during storage. Food Chem. 2020, 308, 125707. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.Y.; Wu, Y.Y.; Li, J.; Chen, D.; Jiang, G.L.; Song, H.Y.; Yin, X.R.; Liu, X.F.; Li, M.Z.; Sun, S.X. Transcriptome analysis reveals the roles of chlorophyll a/b-binding proteins (CABs) and stay-green (SGR) in chlorophyll degradation during fruit development in kiwifruit. N. Z. J. Crop Hortic. Sci. 2021, 49, 106–126. [Google Scholar] [CrossRef]
- Duncan, J.; Bibby, T.; Tanaka, A.; Barber, J. Exploring the ability of chlorophyll b to bind to the CP43′ protein induced under iron deprivation in a mutant of Synechocystis PCC 6803 containing the cao gene. FEBS Lett. 2003, 541, 171–175. [Google Scholar] [CrossRef]
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Antioxidants and Antioxidant Enzymes in Higher Plants, 1st ed.; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Khan, M.I.R.; Khan, N.A. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress, 1st ed.; Springer: Singapore, 2017. [Google Scholar]
- Kyungwon, M.; Bing, L.; Sang-Ryong, L.; Rajeev, A. Supplemental calcium improves freezing tolerance of spinach (Spinacia oleracea L.) by mitigating membrane and photosynthetic damage, and bolstering anti-oxidant and cell-wall status. Sci. Hortic. 2021, 288, 110212. [Google Scholar]
- Xing, F.; Fu, X.-z.; Wang, N.-q.; Xi, J.-l.; Huang, Y.; Zhou, W.; Ling, L.-l.; Peng, L.-z. Physiological changes and expression characteristics of ZIP family genes under zinc deficiency in navel orange (Citrus sinensis). J. Integr. Agric. 2016, 15, 803–811. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Yu, X.; Hu, S.; He, C.; Zhou, J.; Qu, F.; Ai, Z.; Chen, Y.; Ni, D. Chlorophyll Metabolism in Postharvest Tea (Camellia sinensis L.) Leaves: Variations in Color Values, Chlorophyll Derivatives, and Gene Expression Levels under Different Withering Treatments. J. Agric. Food Chem. 2019, 67, 10624–10636. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Yao, X.; Zhang, Y.; Li, J.; Wang, X.; Xu, Z.; Chen, W. Characterization and fine mapping of thermo-sensitive chlorophyll deficit mutant1 in rice (Oryza sativa L.). Breed. Sci. 2015, 65, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 2022, 726, 109248. [Google Scholar] [CrossRef] [PubMed]
- Sosa Alderete, L.G.; Ronchi, H.; Monjes, N.M.; Agostini, E. Tobacco hairy root’s peroxidases are rhythmically controlled by phenol exposure. Enzym. Microb. Technol. 2021, 149, 109856. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Shen, Y.-b.; Zhang, Z.-x. Effects of mechanical damage and herbivore wounding on H2O2 metabolism and antioxidant enzyme activities in hybrid poplar leaves. J. For. Res. 2009, 20, 156–160. [Google Scholar] [CrossRef]
- Costa, M.L.; Civello, P.M.; Chaves, A.R.; Martínez, G.A. Effect of ethephon and 6-benzylaminopurine on chlorophyll degrading enzymes and a peroxidase-linked chlorophyll bleaching during post-harvest senescence of broccoli (Brassica oleracea L.) at 20 °C. Postharvest Biol. Technol. 2005, 35, 191–199. [Google Scholar] [CrossRef]
- Suzuki, T.; Kunieda, T.; Murai, F.; Morioka, S.; Shioi, Y. Mg-dechelation activity in radish cotyledons with artificial and native substrates, Mg-chlorophyllin a and chlorophyllide a. Plant Physiol. Biochem. 2005, 43, 459–464. [Google Scholar] [CrossRef]
- Arkus, K.A.; Cahoon, E.B.; Jez, J.M. Mechanistic analysis of wheat chlorophyllase. Arch. Biochem. Biophys. 2005, 438, 146–155. [Google Scholar] [CrossRef]
- Pillitteri, L.J.; Lovatt, C.J.; Walling, L.L. Isolation and Characterization of LEAFY and APETALA1 Homologues from Citrus sinensis L. Osbeck ‘Washington’. J. Am. Soc. Hortic. Sci. 2004, 129, 845–856. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Correlation Coefficients | Pn | Tr | Gs | Ci |
|---|---|---|---|---|
| Chl a | 0.9898 | 0.9681 | 0.9456 | 0.8462 |
| Chl b | 0.9806 | 0.9557 | 0.9563 | 0.8479 |
| Carotenoid | 0.9800 | 0.9258 | 0.9806 | 0.7800 |
| T-Chl | 0.9856 | 0.9586 | 0.9558 | 0.8402 |
| Gene Name | Accession Number | Length | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|---|---|
| HEMA1 | XM_006472322 | 136 | GTCTTCACCAGCACAGCATCTGA | CACAAGAGCCCACATTACGAGGAA |
| HEME2 | XM_006486147 | 62 | CTGTAGCGGAACCGAAAAATG | TCCTCGAACAGCTTTCAGCAA |
| HEMG1 | XM_006444561 | 162 | TTCTGTAGATGCTGCCGGTG | AATGTTTCCACCCCTTGGCT |
| CHLH | XM_006489925 | 89 | GTGGCGACCCTATCAGGAAC | TGCTGCTGTGGTGGGAATAG |
| CAO | XM_006472723 | 57 | TCGCATCCAATGCCCATAT | TTCTCGCATTTCCCATCTGTT |
| CLH | XM_006443932 | 62 | GAAACGAATCGAGGGATCCA | CTTCAGAAACGCCACCACAA |
| PPH | XM_006482877 | 61 | GATGCAGGTAGTTTCCCAAAAGA | GCAAGCCCGGAATTAAAACC |
| PAO | XM_006487933 | 60 | AAGCAAGAATTTGTCTCCACTACGA | TCTGATGCTGCAGGGTCTGA |
| SGR | XM_006477286 | 167 | CAAGGTCATCTCATCAAGGA | GATTCTACTCCGTTCTTACAAG |
| Actin | XM_006464503 | 195 | CATCCCTCAGCACCTTCC | CCAACCTTAGCACTTCTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, B.; Li, L.; Li, Q.; Mao, H.; Wang, L.; Bie, Y.; Zeng, X.; Liao, L.; Wang, X.; Deng, H.; et al. Identification of Photosynthesis Characteristics and Chlorophyll Metabolism in Leaves of Citrus Cultivar (Harumi) with Varying Degrees of Chlorosis. Int. J. Mol. Sci. 2023, 24, 8394. https://doi.org/10.3390/ijms24098394
Xiong B, Li L, Li Q, Mao H, Wang L, Bie Y, Zeng X, Liao L, Wang X, Deng H, et al. Identification of Photosynthesis Characteristics and Chlorophyll Metabolism in Leaves of Citrus Cultivar (Harumi) with Varying Degrees of Chlorosis. International Journal of Molecular Sciences. 2023; 24(9):8394. https://doi.org/10.3390/ijms24098394
Chicago/Turabian StyleXiong, Bo, Ling Li, Qin Li, Huiqiong Mao, Lixinyi Wang, Yuhui Bie, Xin Zeng, Ling Liao, Xun Wang, Honghong Deng, and et al. 2023. "Identification of Photosynthesis Characteristics and Chlorophyll Metabolism in Leaves of Citrus Cultivar (Harumi) with Varying Degrees of Chlorosis" International Journal of Molecular Sciences 24, no. 9: 8394. https://doi.org/10.3390/ijms24098394
APA StyleXiong, B., Li, L., Li, Q., Mao, H., Wang, L., Bie, Y., Zeng, X., Liao, L., Wang, X., Deng, H., Zhang, M., Sun, G., & Wang, Z. (2023). Identification of Photosynthesis Characteristics and Chlorophyll Metabolism in Leaves of Citrus Cultivar (Harumi) with Varying Degrees of Chlorosis. International Journal of Molecular Sciences, 24(9), 8394. https://doi.org/10.3390/ijms24098394