

Physiological and Proteomic Responses of the Tetraploid Robinia pseudoacacia L. to High CO2 Levels

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Response of Morphological and Gas Exchange Parameters of Tetraploid R. pseudoacacia to High CO2
2.2. Response of Antioxidant System of Tetraploid R. pseudoacacia to High CO2
2.3. Changes in Chlorophyll Levels and Fluorescence Parameters
2.4. Effect of High CO2 on Leaf Respiration Parameters
2.5. Effect of High CO2 on Leaf Stomatal Movement
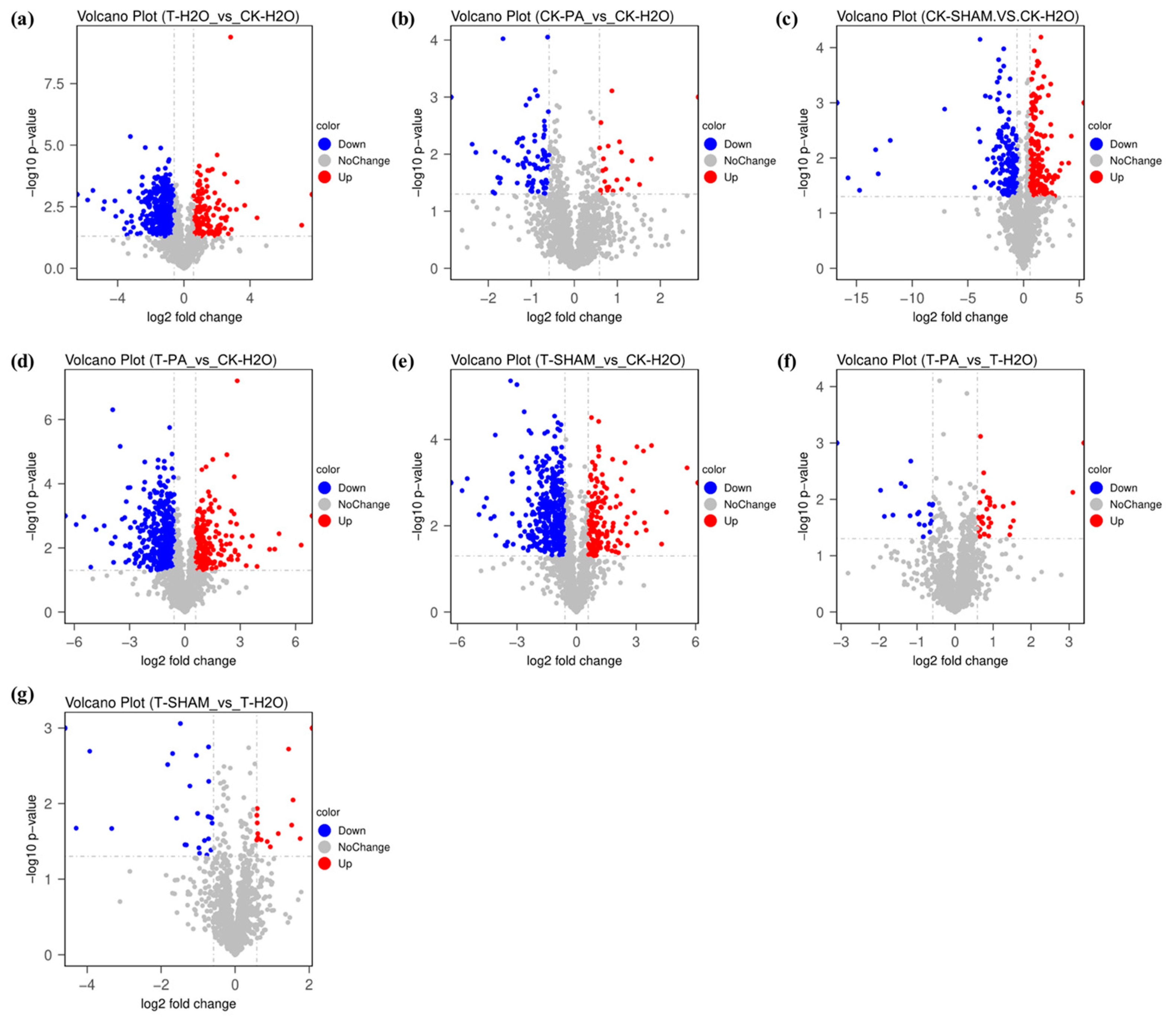
2.6. Proteomic Analysis of Tetraploid R. pseudoacacia Based on High CO2 Conditions
2.7. KOG Functional Annotation and GO Enrichment Analysis of Tetraploid R. pseudoacacia DAPs’ Response to High CO2
2.8. Pathway Analysis of DAPs in Tetraploid R. pseudoacacia under High CO2 Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatment
4.2. Determination of Relative Water Content and Relative Electrical Conductivity
4.3. Determination of Chlorophyll Content and Photosynthetic and Fluorescence Parameters
4.4. Determination of Leaf Respiration Parameters
4.5. Stomatal Movement
4.6. Fluorescent Probe Staining
4.7. Histochemical Staining
4.8. Determination of H2O2 and O2·− Content
4.9. Determination of Antioxidant Enzyme Activity
4.10. Quantitative Proteomics Analysis
4.11. Functional Annotation and Cluster Analysis
4.12. Data Submission
4.13. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Zhang, S.K.; Song, W.Q.; Tigabu, M.; Fu, M.; Xue, H.F.; Sun, A.R.; Zhao, M.H.; Cai, K.W.; Li, Y.; et al. Climate response of radial growth and early selection of Larix olgensis at four trials in northeast China. Dendrochronologia 2022, 73, 125955. [Google Scholar] [CrossRef]
- Cubasch, U.; Wuebbles, D.; Chen, D.; Facchini, M.C.; Frame, D.; Mahowald, N.; Winther, J.G. Introduction. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 95–123. [Google Scholar]
- Morin, F.; André, M.; Betsche, T. Growth Kinetics, Carbohydrate, and Leaf Phosphate Content of Clover (Trifolium subterraneum L.) after Transfer to a High CO2 Atmosphere or to High Light and Ambient Air 1. Plant Physiol. 1992, 99, 89–95. [Google Scholar] [CrossRef]
- Siriphanich, J.; Kader, A.A. Changes in Cytoplasmic and Vacuolar pH in Harvested Lettuce Tissue as Influenced by CO2. J. Am. Soc. Hortic. Sci. 1986, 111, 73–77. [Google Scholar] [CrossRef]
- Larrigaudiere, C.; Pintoó, E.; Lentheric, S.; Vendrell, M. Involvement of oxidative processes in the development of core browning in controlled-atmosphere stored pears. J. Hortic. Sci. Biotechnol. 2001, 76, 157–162. [Google Scholar] [CrossRef]
- Shuyun, Y.; Qingguo, Z.; Yuelin, J.; Fengwen, W.; Xiaofei, Z. Effect of high CO2 density on photosynthesis speed of Pinus massoniana. J. Anhui Agric. Univ. 2006, 33, 100–104. [Google Scholar]
- Jacotot, A.; Marchand, C.; Gensous, S.; Allenbach, M. Effects of elevated atmospheric CO2 and increased tidal flooding on leaf gas-exchange parameters of two common mangrove species: Avicennia marina and Rhizophora stylosa. Photosynth. Res. 2018, 138, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Hale, B.A.; Ormrod, D.P. Amelioration of Ozone-Induced Oxidative Damage in Wheat Plants Grown under High Carbon Dioxide (Role of Antioxidant Enzymes). Plant Physiol. 1995, 109, 421–432. [Google Scholar] [CrossRef]
- Maroco, J.P.; Edwards, G.E.; Ku, M.S. Photosynthetic acclimation of maize to growth under elevated levels of carbon dioxide. Planta 1999, 210, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Tomimatsu, H.; Sakata, T.; Fukayama, H.; Tang, Y. Short-term effects of high CO2 accelerate photosynthetic induction in Populus koreana × trichocarpa with always-open stomata regardless of phenotypic changes in high CO2 growth conditions. Tree Physiol. 2019, 39, 474–483. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, S.; Liu, Y.; Jia, X. Stomatal response of Pinus sylvestriformis to elevated CO2 concentrations during the four years of exposure. J. For. Res. 2005, 16, 15–18. [Google Scholar]
- Chen, J.; Hu, Y.; Yan, R.; Hu, H. Effect of high carbon dioxide injury on the physiological characteristics of fresh-cut garlic scapes. Sci. Hortic. 2019, 250, 359–365. [Google Scholar] [CrossRef]
- Niu, Q.; Huang, D.; Aierken, Y.; Chen, C. Effects of CO2 and Temperature on Canopy Photosynthesis of Muskmelon. Acta Hortic. Sin. 2006, 33, 272–277. [Google Scholar]
- Schulze, S.; Dubeaux, G.; Ceciliato, P.H.O.; Munemasa, S.; Nuhkat, M.; Yarmolinsky, D.; Aguilar, J.; Diaz, R.; Azoulay-Shemer, T.; Steinhorst, L.; et al. A role for calcium-dependent protein kinases in differential CO2- and ABA-controlled stomatal closing and low CO2-induced stomatal opening in Arabidopsis. New Phytol. 2020, 229, 2765–2779. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A. Role of Ca2+ and EGTA on Stomatal Movements in Commelina communis L. Plant Physiol. 1985, 79, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.; Ilan, N.; Grantz, D.A. Calcium Effects on Stomatal Movement in Commelina communis L. Plant Physiol. 1988, 87, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.A.R.; Mcainsh, M.R.; Mansfield, T.A.; Hetherington, A.M. Carbon dioxide induces increases in guard cell cytosolic free calcium. Plant J. 2010, 9, 297–304. [Google Scholar] [CrossRef]
- Haworth, M.; Killi, D.; Materassi, A.; Raschi, A. Coordination of stomatal physiological behavior and morphology with carbon dioxide determines stomatal control. Am. J. Bot. 2015, 102, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Ries, A.B. Carbonic anhydrases function as mediators of CO-induced stomatal movements and regulators of stomatal development in Arabidopsis thaliana. Diss. Theses-Gradworks 2009, 59, 253–257. [Google Scholar]
- He, J.; Zhang, R.X.; Peng, K.; Tagliavia, C.; Li, S.; Xue, S.; Liu, A.; Hu, H.; Zhang, J.; Hubbard, K.E.; et al. The BIG protein distinguishes the process of CO2-induced stomatal closure from the inhibition of stomatal opening by CO2. New Phytol. 2018, 218, 232–241. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Younts, T.L.B.; Muday, G.K. Flavonols control pollen tube growth and integrity by regulating ROS homeostasis during high-temperature stress. Proc. Natl. Acad. Sci. USA 2018, 115, E11188–E11197. [Google Scholar] [CrossRef]
- Sierla, M.; Waszczak, C.; Vahisalu, T.; Kangasjärvi, J. Reactive Oxygen Species in the Regulation of Stomatal Movements. Plant Physiol. 2016, 171, 1569–1580. [Google Scholar] [CrossRef] [PubMed]
- He, J.J.; Zhang, R.X.; Kim, D.S.; Sun, P.; Liu, H.G.; Liu, Z.M.; Hetherington, A.M.; Liang, Y.K. ROS of Distinct Sources and Salicylic Acid Separate Elevated CO2-Mediated Stomatal Movements in Arabidopsis. Front. Plant Sci. 2020, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Matsumoto, Y.; Etoh, T.; Iwai, S. Nitric oxide (NO)-dependent and NO-independent signaling pathways act in ABA-inhibition of stomatal opening. Plant Signal. Behav. 2008, 3, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S.; Lou, C. Involvement of Ca2+ in Stomatal Movements of Vicia faba L. Regulated by Nitric Oxide. J. Plant Physiol. Mol. Biol. 2003, 29, 342–346. [Google Scholar]
- Song, Y.; Miao, Y.; Song, C.-P. Behind the scenes: The roles of reactive oxygen species in guard cells. New Phytol. 2014, 201, 1121–1140. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Parihar, P.; Singh, S.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Reactive oxygen species signaling and stomatal movement: Current updates and future perspectives. Redox Biol. 2017, 11, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Waheeda, K.; Kitchel, H.; Wang, Q.; Chiu, P.L. Molecular mechanism of Rubisco activase: Dynamic assembly and Rubisco remodeling. Front. Mol. Biosci. 2023, 10, 1125922. [Google Scholar] [CrossRef]
- Loughran, T.F.; Boysen, L.; Bastos, A.; Hartung, K.; Havermann, F.; Li, H.; Nabel, J.; Obermeier, W.A.; Pongratz, J. Past and Future Climate Variability Uncertainties in the Global Carbon Budget Using the MPI Grand Ensemble. Glob. Biogeochem. Cycles 2021, 35, e2021GB007019. [Google Scholar] [CrossRef]
- Ikkonen, E.N.; Shibaeva, T.G.; Sherudilo, E.G.; Titov, A.F. Effect of Continuous Lighting on Mitochondrial Respiration in Solanacea Plants. Russ. J. Plant Physiol. 2022, 69, 114. [Google Scholar] [CrossRef]
- McDonald, A.E. Unique opportunities for future research on the alternative oxidase of plants. Plant Physiol. 2023, 191, 2084–2092. [Google Scholar] [CrossRef]
- Torrentino-Madamet, M.; Almeras, L.; Travaille, C.; Sinou, V.; Pophillat, M.; Belghazi, M.; Fourquet, P.; Jammes, Y.; Parzy, D. Proteomic analysis revealed alterations of the Plasmodium falciparum metabolism following salicylhydroxamic acid exposure. Res. Rep. Trop. Med. 2011, 2, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Mallo, N.; Lamas, J.; Leiro, J.M. Evidence of an Alternative Oxidase Pathway for Mitochondrial Respiration in the Scuticociliate Philasterides dicentrarchi. Protist 2013, 164, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Yoda, M.; Shimizu, H.; Sugano, S.; Mizuno, M.; Kino, K.; Usami, S. Contribution of cyanide-insensitive respiratory pathway, catalyzed by the alternative oxidase, to citric acid production in Aspergillus niger. Biosci. Biotechnol. Biochem. 2000, 64, 2034–2039. [Google Scholar] [CrossRef]
- Onda, Y.; Kato, Y.; Abe, Y.; Ito, T.; Ito-Inaba, Y.; Morohashi, M.; Ito, Y.; Ichikawa, M.; Matsukawa, K.; Otsuka, M.; et al. Pyruvate-sensitive AOX exists as a non-covalently associated dimer in the homeothermic spadix of the skunk cabbage, Symplocarpus renifolius. Febs Lett. 2007, 581, 5852–5858. [Google Scholar] [CrossRef]
- Umbach, A.L.; Wiskich, J.T.; Siedow, J.N. Regulation of alternative oxidase kinetics by pyruvate and intermolecular disulfide bond redox status in soybean seedling mitochondria. FEBS Lett. 1994, 348, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Carré, J.E.; Affourtit, C.; Moore, A.L. Interaction of purified alternative oxidase from thermogenic Arum maculatum with pyruvate. Febs Lett. 2011, 585, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.R.; Ma, W.T.; Xu, Y.N.; Wang, X.M.; Li, L.; Tcherkez, G.; Gong, X.Y. Short- and long-term responses of leaf day respiration to elevated atmospheric CO2. Plant Physiol. 2023, 191, 2204–2217. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Lorenc-Plucinska, G.; Reich, P.B. Acclimation of respiratory temperature responses in northern and southern populations of Pinus banksiana. New Phytol. 2009, 181, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, G.Q.; Sun, B.; Zhang, S.; Zhang, Y.Q.; Liao, Y.W.K.; Zhou, Y.H.; Xia, X.J.; Shi, K.; Yu, J.Q. Stimulated Leaf Dark Respiration in Tomato in an Elevated Carbon Dioxide Atmosphere. Sci. Rep. 2013, 3, 3433. [Google Scholar] [CrossRef]
- Zhang, G.; Li, Y.; Li, F.; Xu, Z.; Sun, Y. Effects of root age on biomass and leaf nutrition in tetraploid Robinia pseudoacacia. J. Beijing For. Univ. 2009, 31, 37–41. [Google Scholar]
- Tan, X.; Peng, Z.; Jia, Z.; Ma, L. Influence of air temperatures on photosynthetic light-response curves of Robinia pseudoacacia L. J. Beijing For. Univ. 2010, 32, 64–68. [Google Scholar]
- Meng, F.J.; Pang, H.Y.; Huang, F.L.; Liu, L.; Wang, Y.J. Tetraploid black locust Robinia pseudoacacia L. Increased salt tolerance by activation of the antioxidant system. Biotechnol. Biotechnol. Equip. 2012, 26, 3351–3358. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, X.; Shen, Y. Dynamic changes of anti-oxidation system in new cultvars of Robinia pseudoacacia under gradual drought stress of soil. J. Zhejiang For. Coll. 2005, 22, 166–169. [Google Scholar]
- Liu, J.; Zhang, Z.; Li, Y.; Han, J.; Si, H.; Mi, Y.; Wang, S.; Wei, X.; Yang, H.; Sun, Y.; et al. Effects of the vegetative propagation method on juvenility in Robinia pseudoacacia L. For. Res. 2022, 2, 17. [Google Scholar] [CrossRef]
- Xu, F.; Jiang, M.; Meng, F. Short-term effect of elevated CO2 concentration (0.5%) on mitochondria in diploid and tetraploid black locust Robinia pseudoacacia L. Ecol. Evol. 2017, 7, 4651–4660. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hornbacher, J.; Papenbrock, J.; Markakis, M.N.; Asard, H.; Beemster, G.T.S. Elevated CO2 mitigates the impact of drought stress by upregulating glucosinolate metabolism in Arabidopsis thaliana. Plant Cell Environ. 2023, 46, 812–830. [Google Scholar] [CrossRef] [PubMed]
- Maurer, R.; Tapia, M.E.; Shor, A.C. Exogenous Root Uptake of Glycine Betaine Mitigates Improved Tolerance to Salinity Stress in Avicenna germinans under Ambient and Elevated CO2 Conditions. FASEB J. 2020, 34, 1–1. [Google Scholar] [CrossRef]
- Cohen, I.; Lichston, J.E.; de Macêdo, C.E.C.; Rachmilevitch, S. Leaf coordination between petiole vascular development and water demand in response to elevated CO2 in tomato plants. Plant Direct 2022, 6, e371. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.P.; Melo, N.M.J.; Pereira, E.G.; Halfeld, A.D.; Gomes, I.N.; Prado, C. Responses of woody Cerrado species to rising atmospheric CO2 concentration and water stress: Gains and losses. Funct. Plant Biol. 2016, 43, 1183–1193. [Google Scholar] [CrossRef]
- Silvola, J.; Ahlholm, U. Photosynthesis in willows (Salix × dasyclados) grown at different CO2 concentrations and fertilization levels. Oecologia 1992, 91, 208–213. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B.; Naumburg, E.S.; Koch, G.W.; Kubiske, M.E.; Smith, S.D. Photosynthesis, carboxylation and leaf nitrogen responses of 16 species to elevated pCO2 across four free-air CO2 enrichment experiments in forest, grassland and desert. Glob. Chang. Biol. 2004, 10, 2121–2138. [Google Scholar] [CrossRef]
- Nowak, R.S.; Ellsworth, D.S.; Smith, S.D. Functional responses of plants to elevated atmospheric CO2—Do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol. 2004, 162, 253–280. [Google Scholar] [CrossRef]
- Li, C.T.; Trigani, K.; Zuñiga, C.; Eng, R.; Chen, E.; Zengler, K.; Betenbaugh, M.J. Examining the impact of carbon dioxide levels and modulation of resulting hydrogen peroxide in Chlorella vulgaris. Algal Res.-Biomass Biofuels Bioprod. 2021, 60, 102492. [Google Scholar] [CrossRef]
- Ezraty, B.; Chabalier, M.; Ducret, A.; Maisonneuve, E.; Dukan, S. CO2 exacerbates oxygen toxicity. Embo Rep. 2011, 12, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Gollan, P.J.; Tikkanen, M.; Aro, E.M. Photosynthetic light reactions: Integral to chloroplast retrograde signalling. Curr. Opin. Plant Biol. 2015, 27, 180–191. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Dinakar, C.; Vishwakarma, A.; Raghayendra, A.S.; Padmasree, K. Alternative Oxidase Pathway Optimizes Photosynthesis During Osmotic and Temperature Stress by Regulating Cellular ROS, Malate Valve and Antioxidative Systems. Front. Plant Sci. 2016, 7, 171347. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- del Río, L.A.; Sandalio, L.M.; Corpas, F.J.; Palma, J.M.; Barroso, J.B. Reactive oxygen species and reactive nitrogen species in peroxisomes.: Production, scavenging, and role in cell signaling. Plant Physiol. 2006, 141, 330–335. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Zhu, L.; Wu, J.; Li, M.; Fang, H.; Zhang, J.; Chen, Y.; Chen, J.; Cheng, T. Genome-wide discovery of CBL genes in Nitraria tangutorum Bobr. and functional analysis of NtCBL1-1 under drought and salt stress. For. Res. 2023, 3, 28. [Google Scholar] [CrossRef]
- Chen, Y.; Li, M.; Gong, L.; Song, Y. Effects of Exogenous Signal Substances on POD, CAT Activity and ROS Content in Cistanche deserticola Y. C. Ma Seeds During Germination and Haustorium Formation. Chin. Agric. Sci. Bull. 2014, 30, 128–132. [Google Scholar]
- Li, Y.; Zhou, Q.; Li, F.; Liu, X.; Luo, Y. Effects of tetrabromobisphenol A as an emerging pollutant on wheat (Triticum aestivum) at biochemical levels. Chemosphere 2008, 74, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Zhang, F. Effects of Tetrabromobisphenol a Stress on Growth and Physiological Characteristics of Soybean Seedling. Bull. Environ. Contam. Toxicol. 2017, 98, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Guo, Y.; Ma, Y.; Yang, M.; Fu, R.; Sun, Y. Elevated CO2 delayed yellowing by maintaining chlorophyll biosynthesis and inhibiting chlorophyll degradation and carotenoid accumulation of postharvest broccoli. Postharvest Biol. Technol. 2022, 194, 112089. [Google Scholar] [CrossRef]
- Steer, B.T.; Walker, D.A. Inhibition of Chlorophyll Synthesis by High Concentrations of Carbon Dioxide. Plant Physiol. 1965, 40, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Jin, Z.X.; Li, J.M. Interactive effects of copper stress and arbuscular mycorrhizal fungi on photosynthetic characteristics and chlorophyll fluorescence parameters of elsholtzia splendens. Pak. J. Bot. 2017, 49, 1531–1540. [Google Scholar]
- Laisk, A.; Oja, V.; Rasulov, B.; Eichelmann, H.; Sumberg, A. Quantum Yields and Rate Constants of Photochemical and Nonphotochemical Excitation Quenching (Experiment and Model). Plant Physiol. 1997, 115, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Dahal, K.; Vanlerberghe, G.C. Growth at Elevated CO2 Requires Acclimation of the Respiratory Chain to Support Photosynthesis. Plant Physiol. 2018, 178, 82–100. [Google Scholar] [CrossRef]
- Gandin, A.; Duffes, C.; Day, D.A.; Cousins, A.B. The Absence of Alternative Oxidase AOX1A Results in Altered Response of Photosynthetic Carbon Assimilation to Increasing CO2 in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 1627–1637. [Google Scholar] [CrossRef]
- Hedrich, R.; Neimanis, S.; Savchenko, G.; Felle, H.H.; Kaiser, W.M.; Heber, U. Changes in apoplastic pH and membrane potential in leaves in relation to stomatal responses to CO2, malate, abscisic acid or interruption of water supply. Planta 2001, 213, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 peculation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.J.; Wang, Y.J.; Wang, X.W.; Voronin, P.Y. Effect of doubled CO2 on morphology: Inhibition of stomata development in growing birch Betula platyphylla Suk. leaves. Russ. J. Plant Physiol. 2005, 52, 171–175. [Google Scholar] [CrossRef]
- Ma, X.N.; Bai, L. Elevated CO2 and Reactive Oxygen Species in Stomatal Closure. Plants 2021, 10, 410. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, L.; Khan, D.; Chen, Y.; Pu, X.; Wang, X.; Guan, M.; Rengel, Z.; Chen, Q. Nitric oxide acts downstream of reactive oxygen species in phytomelatonin receptor 1 (PMTR1)-mediated stomatal closure in Arabidopsis. J. Plant Physiol. 2023, 282, 153917. [Google Scholar] [CrossRef] [PubMed]
- Correa Rojo, A.; Heylen, D.; Aerts, J.; Thas, O.; Hooyberghs, J.; Ertaylan, G.; Valkenborg, D. Towards Building a Quantitative Proteomics Toolbox in Precision Medicine: A Mini-Review. Front. Physiol. 2021, 12, 723510. [Google Scholar] [CrossRef]
- Wang, L.; Wang, F.; Zhang, Y.; Ma, Y.; Guo, Y.; Zhang, X. Enhancing the ascorbate–glutathione cycle reduced fermentation by increasing NAD+ levels during broccoli head storage under controlled atmosphere. Postharvest Biol. Technol. 2020, 165, 111169. [Google Scholar] [CrossRef]
- Sicher, R.C.; Hatch, A.L.; Stumpf, D.K.; Jensen, R.G. Ribulose 1,5-Bisphosphate and Activation of the Carboxylase in the Chloroplast. Plant Physiol. 1981, 68, 252–255. [Google Scholar] [CrossRef]
- Fish, L.E.; Bogorad, L. Identification and analysis of the maize P700 chlorophyll a apoproteins PSI-A1 and PSI-A2 by high pressure liquid chromatography analysis and partial sequence determination. J. Biol. Chem. 1986, 261, 8134–8139. [Google Scholar] [CrossRef]
- Fernandez-Velasco, J.G.; Jamshidi, A.; Gong, X.S.; Zhou, J.; Ueng, R.Y. Photosynthetic electron transfer through the cytochrome b6f complex can bypass cytochrome f. J. Biol. Chem. 2001, 276, 30598–30607. [Google Scholar] [CrossRef]
- Ustynyuk, L.Y.; Tikhonov, A.N. Plastoquinol Oxidation: Rate-Limiting Stage in the Electron Transport Chain of Chloroplasts. Biochemistry 2022, 87, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- de Weerd, F.L.; van Stokkum, I.H.M.; van Amerongen, H.; Dekker, J.P.; van Grondelle, R. Pathways for energy transfer in the core light-harvesting complexes CP43 and CP47 of photosystem II. Biophys. J. 2002, 82, 1586–1597. [Google Scholar] [CrossRef]
- Bricker, T.M.; Frankel, L.K. The structure and function of CP47 and CP43 in Photosystem II. Photosynth. Res. 2002, 72, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Karapetyan, N.V. Organization and role of the long-wave chlorophylls in the photosystem I of the Cyanobacterium spirulina. Membr. Cell Biol. 1998, 12, 571–584. [Google Scholar] [PubMed]
- Ishikita, H.; Loll, B.; Biesiadka, J.; Saenger, W.; Knapp, E.W. Redox potentials of chlorophylls in the photosystem II reaction center. Biochemistry 2005, 44, 4118–4124. [Google Scholar] [CrossRef]
- Xu, Y.H.; Liu, R.; Yan, L.; Liu, Z.Q.; Jiang, S.C.; Shen, Y.Y.; Wang, X.F.; Zhang, D.P. Light-harvesting chlorophyll-binding proteins are required for stomatal response to abscisic acid in Arabidopsis. J. Exp. Bot. 2012, 63, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Brettel, K.; Leibl, W. Electron transfer in photosystem I. Biochim. Biophys. Acta 2001, 1507, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Sunde, R.A.; Hoekstra, W.G. Structure, Synthesis and Function of Glutathione Peroxidase. Nutr. Rev. 1980, 38, 265–273. [Google Scholar] [CrossRef]
- Babich, H.; Liebling, E.J.; Burger, R.F.; Zuckerbraun, H.L.; Schuck, A.G. Choice of DMEM, formulated with or without pyruvate, plays an important role in assessing the in vitro cytotoxicity of oxidants and prooxidant nutraceuticals. Vitr. Cell. Dev. Biol. -Anim. 2009, 45, 226–233. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Wang, G.; Guo, D.; Xing, K.; Zhang, S. Biochemical responses of the copepod Centropages tenuiremis to CO2-driven acidified seawater. Water Sci. Technol. 2012, 65, 30–37. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, S.W.; Du, Y.L.; Yin, K.D. Improved drought tolerance in soybean by protein elicitor AMEP412 induced ROS accumulation and scavenging. Biotechnol. Biotechnol. Equip. 2022, 36, 401–412. [Google Scholar] [CrossRef]
- Calatayud, A.; San Bautista, A.; Pascual, B.; Maroto, J.V.; Lopez-Galarza, S. Use of chlorophyll fluorescence imaging as diagnostic technique to predict compatibility in melon graft. Sci. Hortic. 2013, 149, 13–18. [Google Scholar] [CrossRef]
- Xu, L.; Hu, Y.; Jin, G.; Lei, P.; Sang, L.; Luo, Q.; Liu, Z.; Guan, F.; Meng, F.; Zhao, X. Physiological and Proteomic Responses to Drought in Leaves of Amygdalus mira (Koehne) Yü et Lu. Front. Plant Sci. 2021, 12, 620499. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.W.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhang, S.; Lei, P.; Guo, L.; Zhao, X.; Meng, F. Physiological and Proteomic Responses of the Tetraploid Robinia pseudoacacia L. to High CO2 Levels. Int. J. Mol. Sci. 2024, 25, 5262. https://doi.org/10.3390/ijms25105262
Li J, Zhang S, Lei P, Guo L, Zhao X, Meng F. Physiological and Proteomic Responses of the Tetraploid Robinia pseudoacacia L. to High CO2 Levels. International Journal of Molecular Sciences. 2024; 25(10):5262. https://doi.org/10.3390/ijms25105262
Chicago/Turabian StyleLi, Jianxin, Subin Zhang, Pei Lei, Liyong Guo, Xiyang Zhao, and Fanjuan Meng. 2024. "Physiological and Proteomic Responses of the Tetraploid Robinia pseudoacacia L. to High CO2 Levels" International Journal of Molecular Sciences 25, no. 10: 5262. https://doi.org/10.3390/ijms25105262
APA StyleLi, J., Zhang, S., Lei, P., Guo, L., Zhao, X., & Meng, F. (2024). Physiological and Proteomic Responses of the Tetraploid Robinia pseudoacacia L. to High CO2 Levels. International Journal of Molecular Sciences, 25(10), 5262. https://doi.org/10.3390/ijms25105262