
Solid Lipid Nanoparticles Delivering a DNA Vaccine Encoding Helicobacter pylori Urease A Subunit: Immune Analyses before and after a Mouse Model of Infection

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Synthesis of Lipoplex-A Vaccine and Control Vaccine
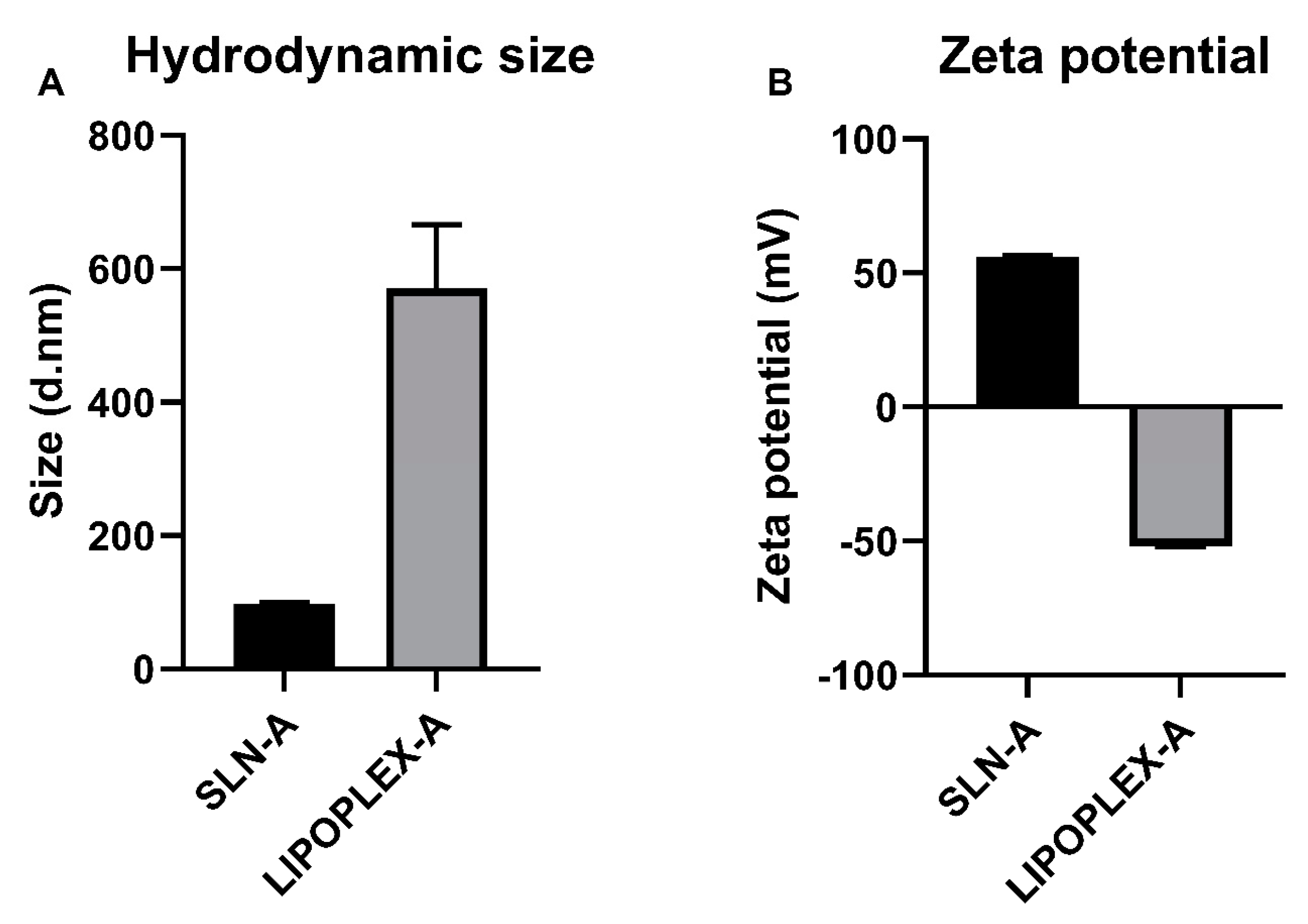
2.2. TEM Characterisation of SLN-A and Lipoplex-A for Vaccine Preparation
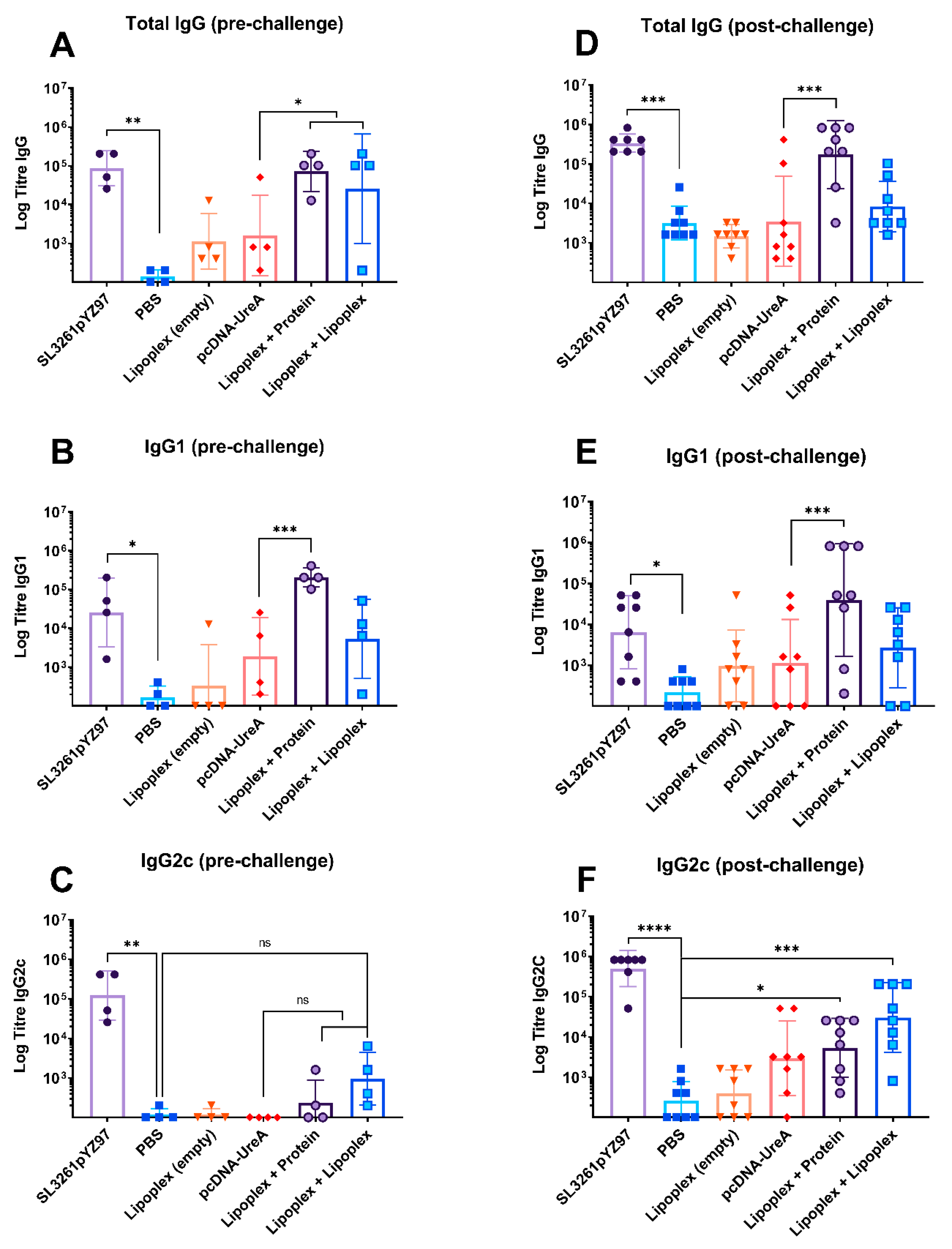
2.3. Humoral Responses to Vaccination and Challenge
2.3.1. Pre-Challenge Antigen-Specific IgG, IgG1, and IgG2c Antibody Levels
2.3.2. Endpoint Antigen-Specific IgG, IgG1, and IgG2c Antibody Levels
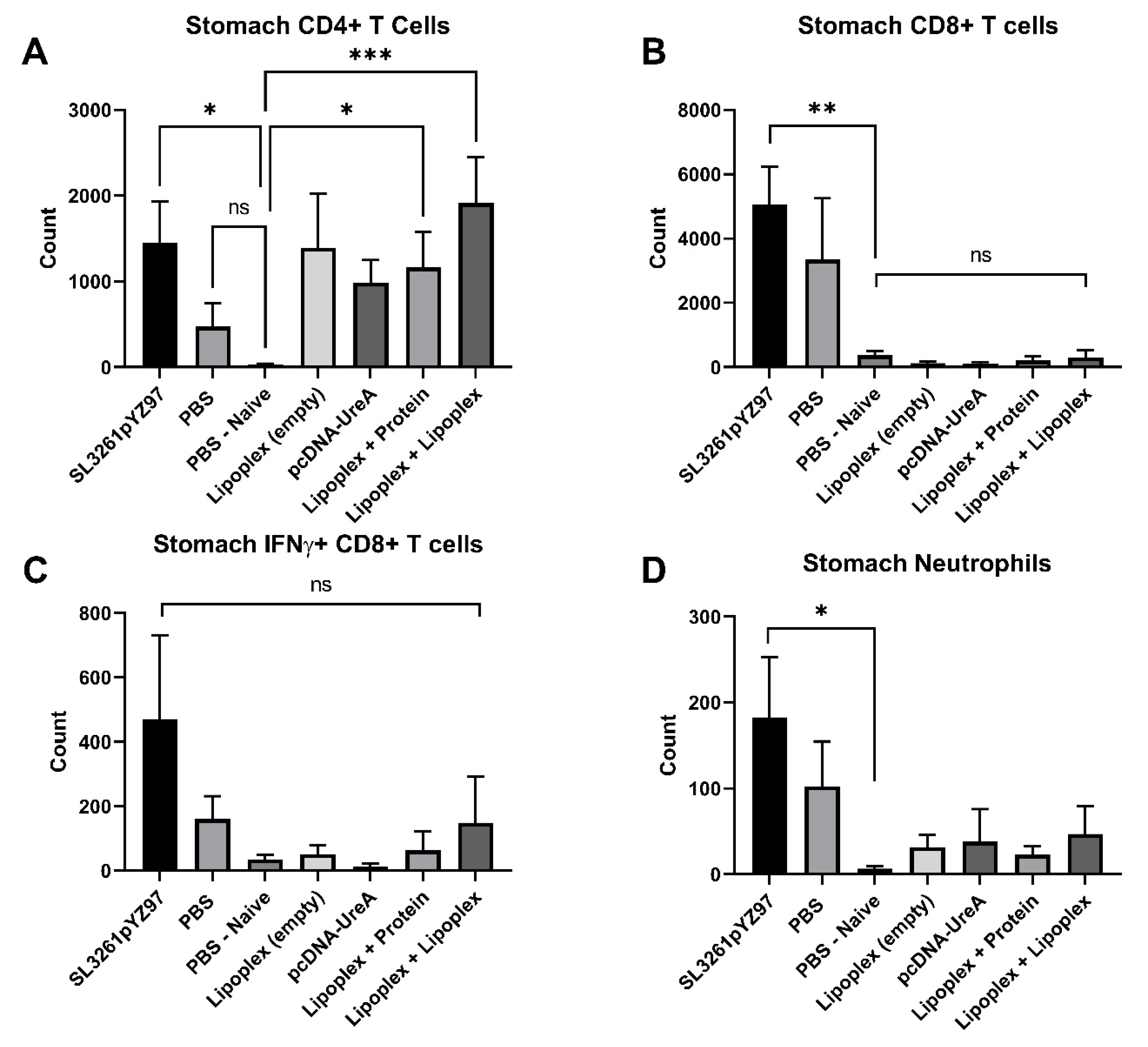
2.4. Analysis of Gastric Infiltrates
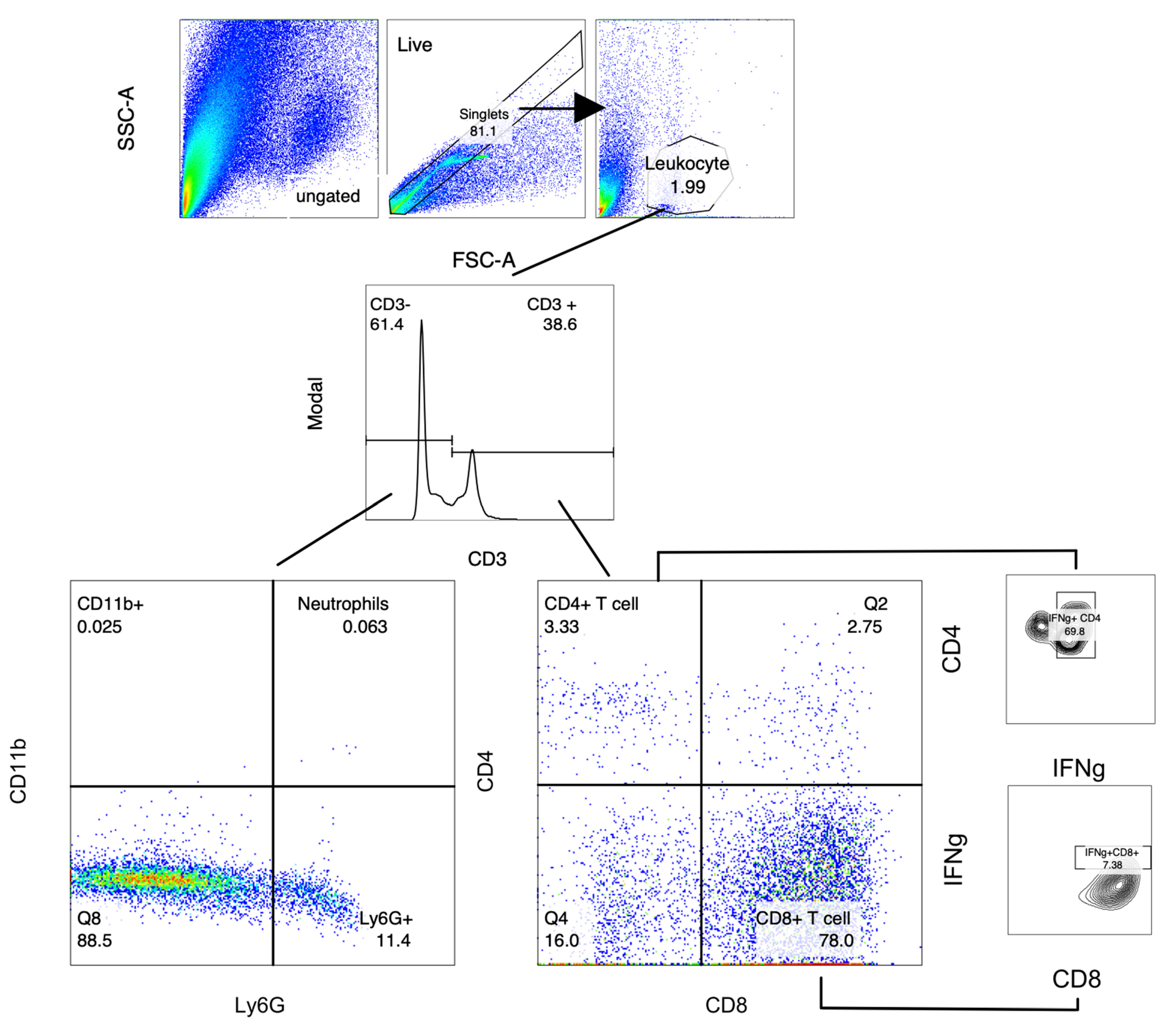
2.5. Analysis of Immune Cell Populations from Mouse Stomach Tissue
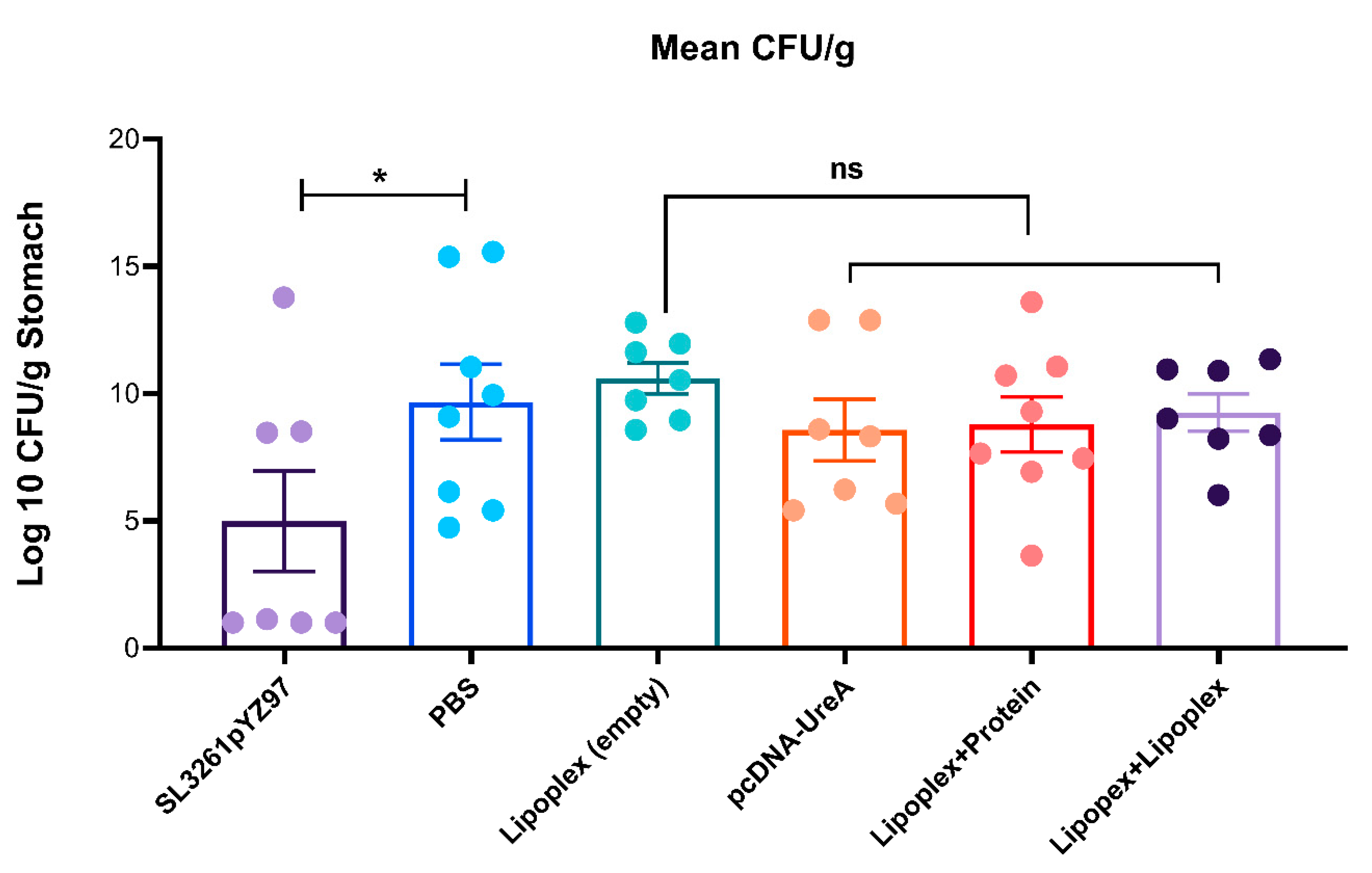
2.6. Determination of H. pylori Burdens from Mouse Stomachs
3. Discussion
4. Materials and Methods
4.1. Formulation of Vaccines and Characterisation by Dynamic Light Scattering (DLS) and TEM
4.2. Immunisation of Mice
4.3. Challenge with H. pylori
4.4. Blood and Tissue Collection
4.5. Antibody Determination by ELISA Assay
4.6. Immune Cell Population Quantification by FACS
4.7. DNA Extraction from Mouse Stomach Homogenates and H. pylori Burden Analysis via Quantitative PCR
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- IARC. Schistosomes, liver flukes and Helicobacter pylori. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Lyon, 7–14 June 1994. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 61, 1–241. [Google Scholar]
- Michetti, P.; Kreiss, C.; Kotloff, K.L.; Porta, N.; Blanco, J.L.; Bachmann, D.; Herranz, M.; Saldinger, P.F.; Corthesy-Theulaz, I.; Losonsky, G.; et al. Oral immunization with urease and Escherichia coli heat-labile enterotoxin is safe and immunogenic in Helicobacter pylori-infected adults. Gastroenterology 1999, 116, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Sweeney, E.G.; Sigal, M.; Zhang, H.C.; Remington, S.J.; Cantrell, M.A.; Kuo, C.J.; Guillemin, K.; Amieva, M.R. Chemodetection and Destruction of Host Urea Allows Helicobacter pylori to Locate the Epithelium. Cell Host Microbe 2015, 18, 147–156. [Google Scholar] [CrossRef]
- Guo, L.; Yang, H.; Tang, F.; Yin, R.; Liu, H.; Gong, X.; Wei, J.; Zhang, Y.; Xu, G.; Liu, K. Oral Immunization with a Multivalent Epitope-Based Vaccine, Based on NAP, Urease, HSP60, and HpaA, Provides Therapeutic Effect on H. pylori Infection in Mongolian gerbils. Front. Cell. Infect. Microbiol. 2017, 7, 349. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wu, Y.; Xu, Z.; Ma, R.; Ding, Y.; Bai, X.; Rong, Q.; Zhang, Y.; Li, B.; Ji, X. Mechanistic Insight into the Interaction Between Helicobacter pylori Urease Subunit Alpha and Its Molecular Chaperone Hsp60. Front. Microbiol. 2019, 10, 153. [Google Scholar] [CrossRef]
- Lucas, B.; Bumann, D.; Walduck, A.; Koesling, J.; Develioglu, L.; Meyer, T.F.; Aebischer, T. Adoptive transfer of CD4+ T cells specific for subunit A of Helicobacter pylori urease reduces H. pylori stomach colonization in mice in the absence of interleukin-4 (IL-4)/IL-13 receptor signaling. Infect. Immun. 2001, 69, 1714–1721. [Google Scholar] [CrossRef]
- Katsande, P.M.; Nguyen, V.D.; Nguyen, T.L.P.; Nguyen, T.K.C.; Mills, G.; Bailey, D.M.D.; Christie, G.; Hong, H.A.; Cutting, S.M. Prophylactic immunization to Helicobacter pylori infection using spore vectored vaccines. Helicobacter 2023, 28, e12997. [Google Scholar] [CrossRef]
- Lehours, P.; Ferrero, R.L. Review: Helicobacter: Inflammation, immunology, and vaccines. Helicobacter 2019, 24 (Suppl. S1), e12644. [Google Scholar] [CrossRef]
- Milani, M.; Sharifi, Y.; Rahmati-Yamchi, M.; Somi, M.H.; Akbarzadeh, A. Immunology and vaccines and nanovaccines for Helicobacter pylori infection. Expert Rev. Vaccines 2015, 14, 833–840. [Google Scholar] [CrossRef]
- Stubljar, D.; Jukic, T.; Ihan, A. How far are we from vaccination against Helicobacter pylori infection? Expert Rev. Vaccines 2018, 17, 935–945. [Google Scholar] [CrossRef]
- Tang, D.; DeVit, M.; Johnston, S.A. Genetic immunization is a simple method for eliciting an immune response. Nature 1992, 356, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical applications of DNA vaccines: Current progress. Clin. Infect. Dis. 2011, 53, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Dauphin, G.; Zientara, S. West Nile virus: Recent trends in diagnosis and vaccine development. Vaccine 2007, 25, 5563–5576. [Google Scholar] [CrossRef]
- Atherton, M.J.; Morris, J.S.; McDermott, M.R.; Lichty, B.D. Cancer immunology and canine malignant melanoma: A comparative review. Vet. Immunol. Immunopathol. 2016, 169, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Khobragade, A.; Bhate, S.; Ramaiah, V.; Deshpande, S.; Giri, K.; Phophle, H.; Supe, P.; Godara, I.; Revanna, R.; Nagarkar, R.; et al. Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): The interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet 2022, 399, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Chozhavel Rajanathan, T.M.; Chandra, H.; Pericherla, H.P.R.; Kumar, S.; Choonia, H.S.; Bajpai, M.; Singh, A.K.; Sinha, A.; Saini, G.; et al. Immunogenic potential of DNA vaccine candidate, ZyCoV-D against SARS-CoV-2 in animal models. Vaccine 2021, 39, 4108–4116. [Google Scholar] [CrossRef]
- Li, L.; Petrovsky, N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev. Vaccines 2016, 15, 313–329. [Google Scholar] [CrossRef]
- Barry, M.E.; Pinto-Gonzalez, D.; Orson, F.M.; McKenzie, G.J.; Petry, G.R.; Barry, M.A. Role of endogenous endonucleases and tissue site in transfection and CpG-mediated immune activation after naked DNA injection. Hum. Gene Ther. 1999, 10, 2461–2480. [Google Scholar] [CrossRef]
- Penumarthi, A.; Basak, P.; Smooker, P.; Shukla, R. Hitching a Ride: Enhancing Nucleic Acid Delivery into Target Cells through Nanoparticles. In Nanoscience in Medicine; Daima, H.K., Pn, N., Ranjan, S., Dasgupta, N., Lichtfouse, E., Eds.; Springer International Publishing: Cham, Switzerland, 2020; Volume 1, pp. 373–457. [Google Scholar]
- Dhadwar, S.S.; Kiernan, J.; Wen, J.; Hortelano, G. Repeated oral administration of chitosan/DNA nanoparticles delivers functional FVIII with the absence of antibodies in hemophilia A mice. J. Thromb. Haemost. 2010, 8, 2743–2750. [Google Scholar] [CrossRef]
- Zupančič, E.; Curato, C.; Paisana, M.; Rodrigues, C.; Porat, Z.; Viana, A.S.; Afonso, C.A.M.; Pinto, J.; Gaspar, R.; Moreira, J.N.; et al. Rational design of nanoparticles towards targeting antigen-presenting cells and improved T cell priming. J. Control. Release 2017, 258, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.J.; Lee, Y.T.; Lee, Y.; Kim, K.H.; Kang, S.M. Distinct Effects of Monophosphoryl Lipid A, Oligodeoxynucleotide CpG, and Combination Adjuvants on Modulating Innate and Adaptive Immune Responses to Influenza Vaccination. Immune Netw. 2017, 17, 326–342. [Google Scholar] [CrossRef] [PubMed]
- Gregg, K.A.; Harberts, E.; Gardner, F.M.; Pelletier, M.R.; Cayatte, C.; Yu, L.; McCarthy, M.P.; Marshall, J.D.; Ernst, R.K. Rationally Designed TLR4 Ligands for Vaccine Adjuvant Discovery. mBio 2017, 8, e00492-17. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.E.; Skakic, I.; Dekiwadia, C.; Shukla, R.; Taki, A.C.; Walduck, A.; Smooker, P.M. Solid Lipid Nanoparticle Carrier Platform Containing Synthetic TLR4 Agonist Mediates Non-Viral DNA Vaccine Delivery. Vaccines 2020, 8, 551. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.E.; Skakic, I.; Smooker, P.M. Design and Preparation of Solid Lipid Nanoparticle (SLN)-Mediated DNA Vaccines. Methods Mol. Biol. 2022, 2412, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Skakic, I.; Francis, J.E.; Dekiwadia, C.; Aibinu, I.; Huq, M.; Taki, A.C.; Walduck, A.; Smooker, P.M. An Evaluation of Urease A Subunit Nanocapsules as a Vaccine in a Mouse Model of Helicobacter pylori Infection. Vaccines 2023, 11, 1652. [Google Scholar] [CrossRef]
- Gomez-Duarte, O.; Lucas, B.; Yan, Z.; Panthel, K.; Haas, R.; Meyer, T.F. Protection of mice against gastric colonization by Helicobacter pylori by single oral dose immunization with attenuated Salmonella typhimurium producing urease subunits A and B. Vaccine 1998, 16, 460–471. [Google Scholar] [CrossRef]
- Becher, D.; Deutscher, M.E.; Simpfendorfer, K.R.; Wijburg, O.L.; Pederson, J.S.; Lew, A.M.; Strugnell, R.A.; Walduck, A.K. Local recall responses in the stomach involving reduced regulation and expanded help mediate vaccine-induced protection against Helicobacter pylori in mice. Eur. J. Immunol. 2010, 40, 2778–2790. [Google Scholar] [CrossRef]
- Aebischer, T.; Bumann, D.; Epple, H.J.; Metzger, W.; Schneider, T.; Cherepnev, G.; Walduck, A.K.; Kunkel, D.; Moos, V.; Loddenkemper, C.; et al. Correlation of T cell response and bacterial clearance in human volunteers challenged with Helicobacter pylori revealed by randomised controlled vaccination with Ty21a-based Salmonella vaccines. Gut 2008, 57, 1065–1072. [Google Scholar] [CrossRef]
- Ferrero, R.L.; Thiberge, J.-M.; Huerre, M.; Labigne, A.S. Immune Responses of Specific-Pathogen-Free Mice to Chronic Helicobacter pylori (Strain SS1) Infection. Infect. Immun. 1998, 66, 1349–1355. [Google Scholar] [CrossRef]
- Arshad, U.; Sarkar, S.; Alipour Talesh, G.; Sutton, P. A lack of role for antibodies in regulating Helicobacter pylori colonization and associated gastritis. Helicobacter 2020, 25, e12681. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Wilken, H.; Faller, G.; Lohoff, M. Systemic Th1 immunization of mice against Helicobacter pylori infection with CpG oligodeoxynucleotides as adjuvants does not protect from infection but enhances gastritis. Infect. Immun. 2004, 72, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Bazin-Lee, H.; Evans, J.T.; Casella, C.R.; Mitchell, T.C. MPL Adjuvant Contains Competitive Antagonists of Human TLR4. Front. Immunol. 2020, 11, 577823. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, J.T.; Myers, K.R. Monophosphoryl Lipid A as an Adjuvant. In Vaccine Design: The Subunit and Adjuvant Approach; Powell, M.F., Newman, M.J., Eds.; Springer: Boston, MA, USA, 1995; pp. 495–524. [Google Scholar]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef]
- Diepen, M.T.v.; Chapman, R.; Douglass, N.; Galant, S.; Moore, P.L.; Margolin, E.; Ximba, P.; Morris, L.; Rybicki, E.P.; Williamson, A.-L.; et al. Prime-Boost Immunizations with DNA, Modified Vaccinia Virus Ankara, and Protein-Based Vaccines Elicit Robust HIV-1 Tier 2 Neutralizing Antibodies against the CAP256 Superinfecting Virus. J. Virol. 2019, 93, e02155-18. [Google Scholar] [CrossRef] [PubMed]
- Da’Dara, A.A.; Skelly, P.J.; Walker, C.M.; Harn, D.A. A DNA-prime/protein-boost vaccination regimen enhances Th2 immune responses but not protection following Schistosoma mansoni infection. Parasite Immunol. 2003, 25, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Gil, A.; Shen, S.; Coley, S.; Gibson, L.; Diamond, D.J.; Wang, S.; Lu, S. DNA vaccine prime followed by boost with live attenuated virus significantly improves antigen-specific T cell responses against human cytomegalovirus. Hum. Vaccines Immunother. 2014, 9, 2120–2132. [Google Scholar] [CrossRef]
- Taylor, N.S.; Fox, J.G. Animal models of Helicobacter-induced disease: Methods to successfully infect the mouse. Methods Mol. Biol. 2012, 921, 131–142. [Google Scholar] [CrossRef]
- Zhang, S.; Moss, S.F. Rodent models of Helicobacter infection, inflammation, and disease. Methods Mol. Biol. 2012, 921, 89–98. [Google Scholar] [CrossRef]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate Immune Response in Th1- and Th2-Dominant Mouse Strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Benga, L.; Grunwald, T.; Müller, U.; Schrödl, W.; Baums, C.G. Comparative analysis of humoral immune responses and pathologies of BALB/c and C57BL/6 wildtype mice experimentally infected with a highly virulent Rodentibacter pneumotropicus (Pasteurella pneumotropica) strain. BMC Microbiol. 2018, 18, 45. [Google Scholar] [CrossRef]
- Panthel, K.; Faller, G.; Haas, R. Colonization of C57BL/6J and BALB/c wild-type and knockout mice with Helicobacter pylori: Effect of vaccination and implications for innate and acquired immunity. Infect. Immun. 2003, 71, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Gary, E.N.; Weiner, D.B. DNA vaccines: Prime time is now. Curr. Opin. Immunol. 2020, 65, 21–27. [Google Scholar] [CrossRef]
- Hobernik, D.; Bros, M. DNA Vaccines-How Far From Clinical Use? Int. J. Mol. Sci. 2018, 19, 3605. [Google Scholar] [CrossRef] [PubMed]
- Jorritsma, S.H.T.; Gowans, E.J.; Grubor-Bauk, B.; Wijesundara, D.K. Delivery methods to increase cellular uptake and immunogenicity of DNA vaccines. Vaccine 2016, 34, 5488–5494. [Google Scholar] [CrossRef] [PubMed]
- Jazayeri, M.; Soleimanjahi, H.; Fotouhi, F.; Pakravan, N. Comparison of intramuscular and footpad subcutaneous immunization with DNA vaccine encoding HSV-gD2 in mice. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 453–461. [Google Scholar] [CrossRef]
- Ledwith, B.J.; Manam, S.; Troilo, P.J.; Barnum, A.B.; Pauley, C.J.; Griffiths II, T.G.; Harper, L.B.; Beare, C.M.; Bagdon, W.J.; Nichols, W.W. Plasmid DNA vaccines: Investigation of integration into host cellular DNA following intramuscular injection in mice. Intervirology 2000, 43, 258–272. [Google Scholar] [CrossRef]
- Ottsjö, L.S.; Flach, C.F.; Clements, J.; Holmgren, J.; Raghavan, S. A double mutant heat-labile toxin from Escherichia coli, LT(R192G/L211A), is an effective mucosal adjuvant for vaccination against Helicobacter pylori infection. Infect. Immun. 2013, 81, 1532–1540. [Google Scholar] [CrossRef]
- Garhart, C.A.; Heinzel, F.P.; Czinn, S.J.; Nedrud, J.G. Vaccine-induced reduction of Helicobacter pylori colonization in mice is interleukin-12 dependent but gamma interferon and inducible nitric oxide synthase independent. Infect. Immun. 2003, 71, 910–921. [Google Scholar] [CrossRef]
- Hatzifoti, C.; Bajaj-Elliott, M.; Dorrell, N.; Anyim, M.; Prentice, M.B.; Nye, K.E.; Wren, B.; Morrow, W.J.W. A plasmid immunization construct encoding urease B of Helicobacter pylori induces an antigen-specific antibody response and upregulates the expression of β-defensins and IL-10 in the stomachs of immunized mice. Vaccine 2004, 22, 2651–2659. [Google Scholar] [CrossRef] [PubMed]
- Longet, S.; Abautret-Daly, A.; Davitt, C.J.H.; McEntee, C.P.; Aversa, V.; Rosa, M.; Coulter, I.S.; Holmgren, J.; Raghavan, S.; Lavelle, E.C. An oral alpha-galactosylceramide adjuvanted Helicobacter pylori vaccine induces protective IL-1R- and IL-17R-dependent Th1 responses. npj Vaccines 2019, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Shirzad, H.; Elahi, S.; Azadegan-Dehkordi, F.; Rahimian, G.; Shafigh, M.; Rashidii, R.; Sarafnejad, A.; Rafieian-Kopaei, M.; Faridani, R.; et al. Downregulated regulatory T cell function is associated with increased peptic ulcer in Helicobacter pylori-infection. Microb. Pathog. 2017, 110, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Penumarthi, A.; Parashar, D.; Abraham, A.N.; Dekiwadia, C.; Macreadie, I.; Shukla, R.; Smooker, P.M. Solid lipid nanoparticles mediate non-viral delivery of plasmid DNA to dendritic cells. J. Nanopart. Res. 2017, 19, 210. [Google Scholar] [CrossRef]
- Gorrell, R.J.; Wijburg, O.L.C.; Pedersen, J.S.; Walduck, A.K.; Kwok, T.; Strugnell, R.A.; Robins-Browne, R.M. Contribution of secretory antibodies to intestinal mucosal immunity against Helicobacter pylori. Infect. Immun. 2013, 81, 3880–3893. [Google Scholar] [CrossRef]
- Tan, M.P.; Kaparakis, M.; Galic, M.; Pedersen, J.; Pearse, M.; Wijburg, O.L.C.; Janssen, P.H.; Strugnell, R.A. Chronic Helicobacter pylori infection does not significantly alter the microbiota of the murine stomach. Appl. Environ. Microbiol. 2007, 73, 1010–1013. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vaccine | Boost | Dose per Vaccination | Challenged |
|---|---|---|---|---|
| A | S. typhimurium SL3261pYZ97 | - | 1 × 107 CFU | ✓ |
| B | PBS-sham | PBS | 100 µL PBS | ✓ |
| C | PBS-sham | PBS | 100 µL PBS | ✗ |
| D | Lipoplex-A (empty plasmid vector) | Lipoplex-A (empty plasmid vector) | 50 µg DNA + 100 µg SLN-A | ✓ |
| E | DNA only (pcDNA-UreA) | DNA only (pcDNA-UreA) | 50 µg DNA | ✓ |
| F | Lipoplex-A | UreA protein | 50 µg DNA + 100 µg SLN-A, 50 µg UreA protein (boost) | ✓ |
| G | Lipoplex-A | Lipoplex-A | 50 µg DNA + 100 µg SLN-A | ✓ |
| FORWARD | 5′-CTTAACCATAGAACTGCATTTGAAACTAC-3′ |
| REVERSE | 5′-GGTCGCCTTCGCAATGAGTA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francis, J.E.; Skakic, I.; Majumdar, D.; Taki, A.C.; Shukla, R.; Walduck, A.; Smooker, P.M. Solid Lipid Nanoparticles Delivering a DNA Vaccine Encoding Helicobacter pylori Urease A Subunit: Immune Analyses before and after a Mouse Model of Infection. Int. J. Mol. Sci. 2024, 25, 1076. https://doi.org/10.3390/ijms25021076
Francis JE, Skakic I, Majumdar D, Taki AC, Shukla R, Walduck A, Smooker PM. Solid Lipid Nanoparticles Delivering a DNA Vaccine Encoding Helicobacter pylori Urease A Subunit: Immune Analyses before and after a Mouse Model of Infection. International Journal of Molecular Sciences. 2024; 25(2):1076. https://doi.org/10.3390/ijms25021076
Chicago/Turabian StyleFrancis, Jasmine E., Ivana Skakic, Debolina Majumdar, Aya C. Taki, Ravi Shukla, Anna Walduck, and Peter M. Smooker. 2024. "Solid Lipid Nanoparticles Delivering a DNA Vaccine Encoding Helicobacter pylori Urease A Subunit: Immune Analyses before and after a Mouse Model of Infection" International Journal of Molecular Sciences 25, no. 2: 1076. https://doi.org/10.3390/ijms25021076
APA StyleFrancis, J. E., Skakic, I., Majumdar, D., Taki, A. C., Shukla, R., Walduck, A., & Smooker, P. M. (2024). Solid Lipid Nanoparticles Delivering a DNA Vaccine Encoding Helicobacter pylori Urease A Subunit: Immune Analyses before and after a Mouse Model of Infection. International Journal of Molecular Sciences, 25(2), 1076. https://doi.org/10.3390/ijms25021076