
Programmed Death-Ligand (PD-L1), Epidermal Growth Factor (EGF), Relaxin, and Matrix Metalloproteinase-3 (MMP3): Potential Biomarkers of Malignancy in Canine Mammary Neoplasia


 , and
, and
Abstract
:1. Introduction
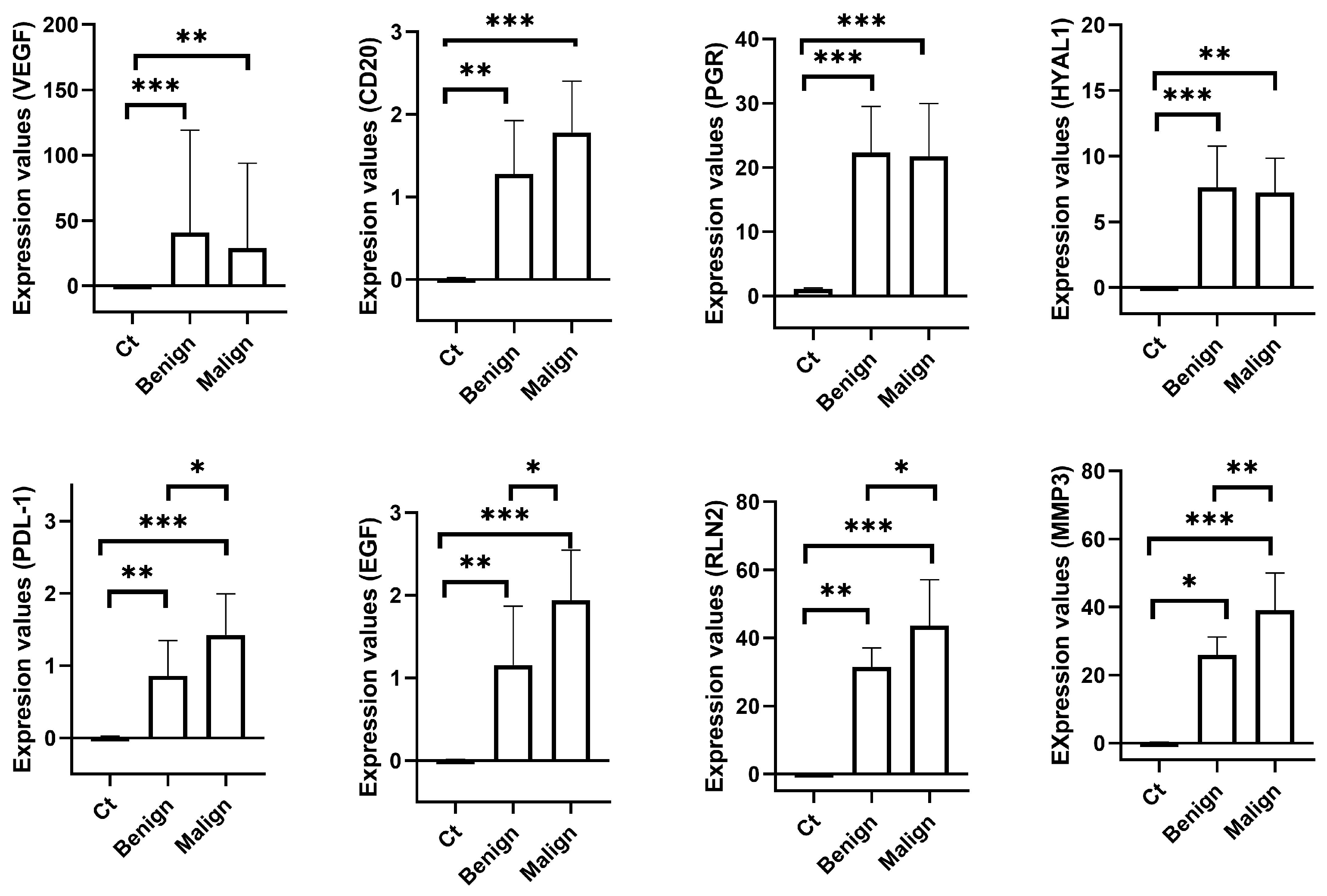
2. Results
2.1. Tumours Typification
2.2. Estimation of Gene Efficiency
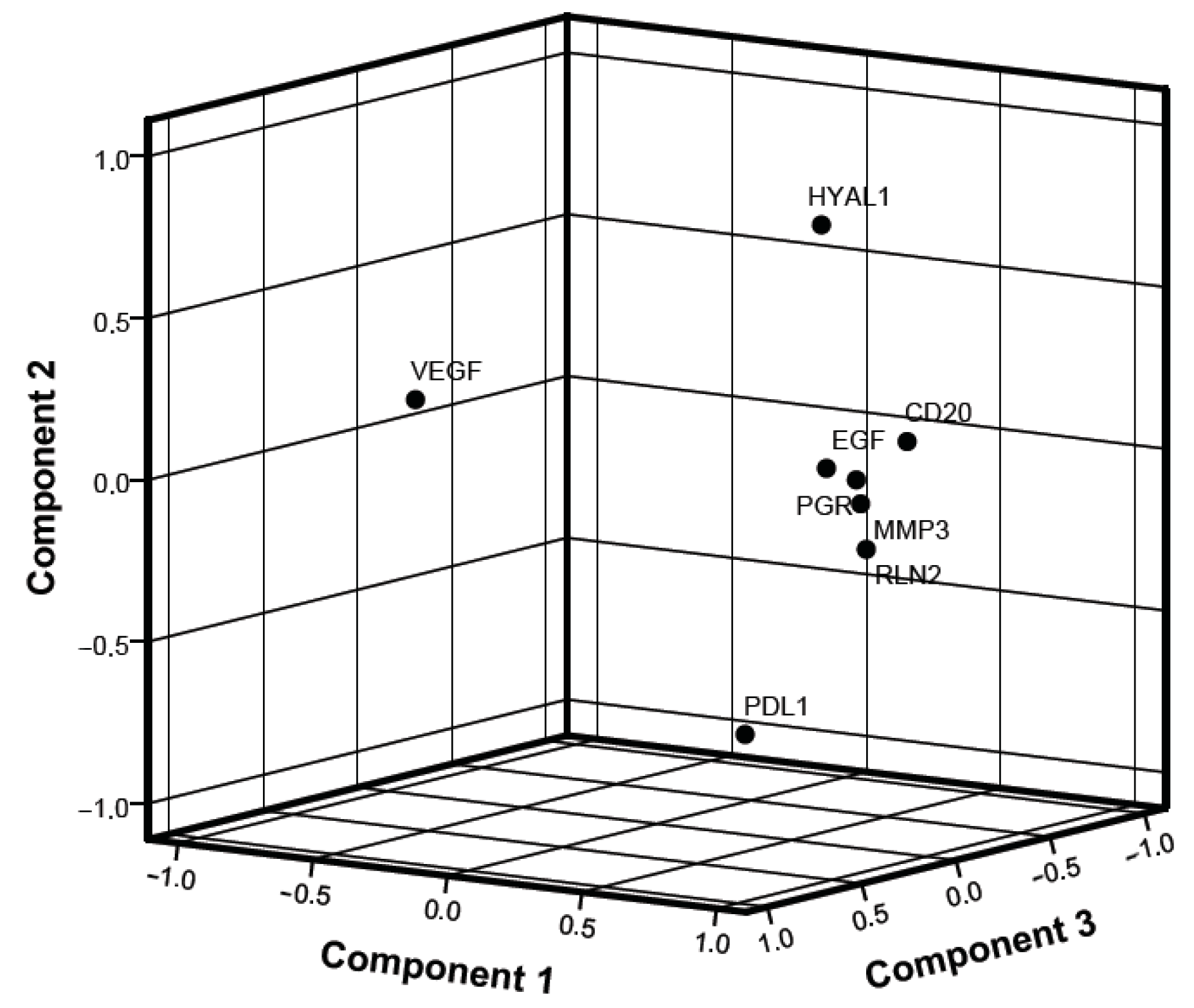
2.3. Correlation between Genes
3. Discussion
4. Materials and Methods
4.1. Animals and Sampling
4.2. Extraction of RNA and Synthesis of Complementary DNA
4.3. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.4. Standardization Strategy
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene Name | Category | Accession Numbers | Forward Primer | Reverse Primer |
|---|---|---|---|---|
| GADPH | Housekeeping | NM_001003142 | TGTCCCCACCCCCAATGTATC | CTCCGATGCCTGCTTCACTACCTT |
| HPRT | Housekeeping | AY_283372 | AGCTTGCTGGTGAAAAGGAC | TTATAGTCAAGGGCATATCC |
| RPL8 | Housekeeping | XM_532360.2 | CCATGAATCCTGTGGAGC | GTAGAGGGTTTGCCGATG |
| RPS19 | Housekeeping | XM_533657.3 | CCTTCCTCAAAAAGTCTGGG | GTTCTCATCGTAGGGAGCAAG |
| HYAL1 | Malignity | XM_005632508.4 | CACAGGGAAGGGACAGATGT | TGTTCTCCCAGCTTACCCAG |
| MMP3 | Malignity | NM_001002967.1 | CCTAGCGCTCTGATGTACCC | GGACTGGATGCCATTCACAT |
| RLN2 | Malignity | NM_001003132.1 | TGCTAGGTGTCTGGCTGCTACTAA | CGGACATAATCACGACCACATG |
| VEGFα | Malignity | NM_001003175.2 | GTGCCCACTGAGGAGTTCAAC | CCCTATGTGCTGGCCTTGAT |
| PGR | Hormonal | NM_001003074.1 | CGAGTCATTACCTCAGAAGATTTGTTT | CTTCCATTGCCCTTTTAAAGAAGA |
| CD20 | Malignity | AB210085.1 | AACTTTGCAGAACCTCCCCAG | AAGCTGTGAACACTAACGCCT |
| EGF | Malignity | NM_001003094.3 | GTTGAAAGTTCGAGCCCCCT | CAGTTCCAGTTCTCCGGGTC |
| PD-L1 | Malignity | AB898678.1 | CCGCCAGCAGGTCACTT | TCCATTGTCACATTGCCACC |
References
- Morris, J.S. Genomic and proteomic profiling for cancer diagnosis in dogs. Vet. J. 2016, 215, 101–109. [Google Scholar] [PubMed]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumors. Nature 2000, 406, 747–752. [Google Scholar] [PubMed]
- Graham, J.C.; O’Keefe, D.A.; Gelberg, H.B. Immunohistochemical assay for detecting estrogen receptors in canine mammary tumours. Am. J. Vet. Res. 1999, 60, 627–630. [Google Scholar] [PubMed]
- Gama, A.; Alves, A.; Schmitt, F. Identification of molecular phenotypes in canine mammary carcinomas with clinical implications: Application of the human classification. Virchows Arch. 2008, 453, 123–132. [Google Scholar]
- Ding, S.; Chen, S.; Shen, K. Single-cell RNA sequencing in breast cancer: Understanding tumor heterogeneity and paving roads to individualized therapy. Cancer Commun. 2020, 40, 329–344. [Google Scholar]
- González Castro, L.N.; Tirosh, I.; Suvà, M.L. Decoding cancer biology one cell at a time. Cancer Discov. 2021, 11, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, L.; Zhang, Y.; Li, Z.; Siemers, N.; Zhang, Z. Insights gained from single-cell analysis of immune cells in the tumor microenvironment. Annu. Rev. Immunol. 2021, 39, 583–609. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Kim, D.Y.; Henry, C.J.; Bryan, J.N.; Robinson, K.L.; Eaton, A.M. Programmed death ligand 1 is expressed in canine B cell lymphoma and downregulated by MEK inhibitors. Vet. Comp. Oncol. 2017, 15, 1527–1536. [Google Scholar] [CrossRef]
- Sakalauskaite, S.; Salteniene, V.; Nikitina, D.; Ugenskiene, R.; Riskeviciene, V.; Karveliene, B.; Juodziukyniene, N. VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 genes expression and the association with tumor clinicopathological parameters and dog breed assessment. Vet. Sci. 2021, 8, 212. [Google Scholar] [CrossRef]
- Pawlowski, K.M.; Maciejewski, H.; Majchrzad, K.; Dolka, I.; Mol, J.A.; Motyl, T.; Król, M. Fiver markers useful for the distinction of canine mammary malignancy. BMC Vet. Res. 2013, 9, 138. [Google Scholar] [CrossRef]
- Peña, L.; Gama, A.; Goldsmithy, M.H.; Abadie, J.; Benazzi, C.; Castagnaro, M.; Díez, L.; Gärtner, F.; Hellmén, E.; Kiupel, M.; et al. Canine mammary tumors: A review and consensus of standard guideline on epithelial and myoepithelial phenotype makers, HER2, and hormone receptor assessment using immunohistochemistry. Vet. Pathol. 2014, 51, 127–145. [Google Scholar] [CrossRef]
- Selvarajah, G.T.; Bonestro, F.A.S.; Timmermans Sprang, E.P.M.; Kirpensteijn, J.; Mol, J.A. Reference gene validation for gene expression normalization in canine osteosarcoma: A geNorm algorithm approach. Bmc Vet. Res. 2017, 13, 354. [Google Scholar]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumours. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [PubMed]
- Gavalas, N.G.; Liontos, M.; Trachana, S.P.; Bragatuni, T.; Arapinis, C.; Liacos, C.; Dimopoulus, M.A.; Bamias, A. Angiogenesis-related pathways in the pathogenesis of ovarian cancer. Int. J. Mol. Sci. 2013, 12, 15885–15909. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Bosça, A.B.; Susman, S.; Marginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar] [PubMed]
- Romon, R.; Adriaenssens, R.; Lagadec, C.; Germain, E.; Hondermarck, H.; Le Bourhis, X. Nerve growth factor promotes breast cancer angiogenesis by activating multiple pathways. Mol. Cancer 2010, 9, 157. [Google Scholar] [PubMed]
- Greenberg, S.; Rugo, H.S. Triple-negative breast cancer: Role of antiangiogenic agents. Cancer J. 2010, 16, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Taneja, P.; Maglic, D.; Kai, F.; Zhu, S.; Kendig, R.D.; Fry, E.A.; Inoeu, K. Classical and novel prognostic markers for breast cancer and their clinical significance. Clin. Med. Insights Oncol. 2010, 4, 15–34. [Google Scholar]
- Zhang, J.; Lu, A.; Li, L.; Yue, J.; Lu, Y. p16 modulates VEGF expression via its interaction with HIF-1alpha in breast cancer cells. Cancer Investig. 2010, 28, 588–597. [Google Scholar]
- Koukourakis, M.I.; Limberis, V.; Tentes, I.; Kontomanolis, E.; Kortsaris, A.; Sivridis, E.; Giatromanolaki, A. Serum VEGF levels and tissue activation of VEGFR2/KDR receptors in patients with breast and gynecologic cancer. Cytokine 2011, 53, 370–375. [Google Scholar]
- Qiu, C.W.; Lin, D.G.; Wang, J.P.; Li, C.Y.; Deng, G.Z. Expression and significance of PTEN and VEGF in canine mammary gland tumours. Vet. Res. Commun. 2008, 32, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Al-Dissi, A.; Haines, D.M.; Singh, B.; Kidney, B.A. Immunohistochemical expression of vascular endothelial growth factor and vascular endothelial growth factor receptor-2 in canine simple mammary gland adenocarcinomas. Can. Vet. J. 2010, 51, 1109–1114. [Google Scholar]
- Millanta, F.; Caneschi, V.; Ressel, L.; Citi, S.; Poli, A. Expression of vascular endothelial growth factor in canine inflammatory and non-inflammatory mammary carcinoma. J. Comp. Pathol. 2010, 142, 26–42. [Google Scholar]
- Moschetta, M.G.; Maschio, L.B.; Jardim-Perassi, B.V.; Gelaleti, G.B.; Lopes, J.R.; Leonel, C.; Conçalves Ndo, N.; Ferreira, L.C.; Martins, G.R.; Borin, T.F.; et al. Prognostic value of vascular endothelial growth factor and hypoxia-inducible factor 1α in canine malignant mammary tumours. Oncol. Rep. 2015, 33, 2345–2353. [Google Scholar] [CrossRef]
- Rismanchi, S.; Muhammadnejad, S.; Amanpour, S.; Muhammadnejad, A. First pathological study of canine primary breast lymphoma and the description of its clinicopathological characteristics as an animal model for human primary breast lymphoma. Biomed. Rep. 2015, 3, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Galadima, M.; Kotova, I.; Schmidt, R.; Pastor, J.; Schröder, C.; Rodríguez-Gil, J.E.; Rivera del Alamo, M.M. Canine mammary neoplasia induces variations in the peripheral blood levels of CD20, CD45RA, and CD99. Int. J. Mol. Sci. 2023, 24, 9222. [Google Scholar] [CrossRef]
- Schmidt, M.; Böhm, D.; von Törne, C.; Steiner, E.; Puhl, A.; Pilch, H.; Lehr, H.A.; Hengstler, J.G.; Kölbl, H.; Gehrmann, M. The humoral immune system has a key prognostic impact in node-negative breast cancer. Cancer Res. 2008, 68, 5405–5413. [Google Scholar] [CrossRef] [PubMed]
- Olkhanud, P.B.; Damdinsuren, B.; Bodogai, M.; Gress, R.E.; Sen, R.; Wejksza, K.; Malchinkhuu, E.; Wersto, R.P.; Biragyn, A. Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4+ T Cells to T-regulatory cells. Cancer Res. 2011, 71, 3505–3515. [Google Scholar]
- Schnellhardt, S.; Erber, R.; Büttner-Herold, M.; Rosahl, M.C.; Ott, O.J.; Strnad, V.; Beckman, M.W.; King, L.; Hartmann, A.; Fietkau, R.; et al. Tumour-infiltrating inflammatory cells in early breast cancer: An underrated prognostic and predictive factor? Int. J. Mol. Sci. 2020, 21, 8238. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Worley, D.R.; Goldschmidt, M.H. Mammary gland tumors in dogs. In Withrow & MacEwen’s Small Animal Clinical Oncology, 5th ed.; Withrow, S.J., Vail, D.M., Eds.; Saunders Elsevier: St. Louis, MO, USA, 2013; pp. 538–556. [Google Scholar]
- Martin de las Mulas, J.; Millán, Y.; Dios, R. A prospective analysis of immunohistochemically determined estrogen receptor Alpha and progesterone receptor expression and host tumor factors as predictors of disease-free period in mammary tumors of the dog. Vet. Pathol. 2005, 42, 200–212. [Google Scholar]
- Sartorius, C.A.; Melvill, M.Y.; Hovland, A.R.; Tung, L.; Takimoto, G.S.; Horwitz, K.B. A third transactivation function (AF3) of human progesterone receptors located in the unique N-terminal segment of the B-isoform. Mol. Endocrinol. 1994, 8, 1347–1360. [Google Scholar] [PubMed]
- Geraldes, M.; Gärtner, F.; Schmitt, F. Immunohistochemical study of hormonal receptors and cell proliferation in normal canine mammary glands and spontaneous mammary tumours. Vet. Rec. 2000, 146, 403–406. [Google Scholar] [PubMed]
- Guil-Luna, S.; Stenvang, J.; Brünner, N.; Sánchez-Céspedes, R.; Millán, Y.; Gómez-Laguna, J.; Martín de las Mulas, J. Progesterone receptor isoform analysis by quantitative real-time polymerase chain reaction in formalin-fixed, paraffin-embeddes canine mammary dysplasias and tumors. Vet. Pathol. 2013, 51, 895–902. [Google Scholar] [CrossRef]
- Guil-Luna, S.; Stenvang, J.; Brünner, N.; De Andrés, F.J.; Rollón, E.; Domingo, V.; Sánchez-Céspedes, R.; Millán, Y.; Martín de las Mulas, J. Progesterone receptor isoform A my regulate the effects of neoadjuvant aglepristone in canine mammary carcinoma. BMC Vet. Res. 2014, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Csoka, T.; Stern, R. The hyaluronidases: A chemical, biological and clinical overview. TIGG 1996, 8, 419–434. [Google Scholar] [CrossRef]
- Varallo, G.R.; Jardim-Perassi, B.V.; Alexandre, P.A.; Fukumasu, H.; Zuccari, D.A.P.C. Global gene expression profile in canine mammary carcinomas. Vet. J. 2019, 254, 105393. [Google Scholar] [PubMed]
- West, D.C.; Hampson, I.N.; Arnold, F.; Fumar, S. Angiogenesis induced by degradation products of hyaluronic acid. Science 1985, 228, 1324–1326. [Google Scholar] [CrossRef]
- Lokeshwar, V.B.; Obek, C.; Soloway, M.S.; Block, N.L. Tumor-associated hyaluronic acid: A new sensitive and specific urine marker for bladder cancer. Canc. Res. 1997, 57, 773–777. [Google Scholar]
- Chao, K.L.; Muthukumar, L.; Herzberg, O. Structure of human hyaluronidase-1, a hyaluronan hydrolyzing enzyme involved in tumor growth and angiogenesis. Biochemistry 2007, 46, 6911–6920. [Google Scholar] [CrossRef]
- Golshani, R.; Hautmann, S.H.; Estrella, V.; Cohen, B.L.; Kyle, C.C.; Manoharan, M.; Jorda, M.; Soloway, M.S.; Lokeshwar, V.B. HAS1 expression in bladder cancer and its relation to urinary HA test. Int. J. Canc. 2007, 120, 1712–1720. [Google Scholar] [CrossRef]
- Bharadwaj, A.G.; Kovar, J.L.; Loghman, E.; Elowsky, C.; Oakley, G.G.; Simpson, M.A. Spontaneous metastasis of prostate cancer is promoted by excess hyaluronan synthesis and processing. Am. J. Pathol. 2009, 174, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Kuppusamy, H.; Pilarski, L.M. Aberrant splice variants of HAS1 (Hyaluronan synthase 1) multimerize with and modulate normally spliced HAS1 protein: A potential mechanism promoting human cancer. J. Biol. Chem. 2009, 284, 18840–18850. [Google Scholar] [CrossRef]
- Adamia, S.; Pilarski, P.M.; Belch, A.R.; Pilarski, L.M. Aberrant splicing, hyaluronan synthases and intracellular hyaluronan as drivers of oncogenesis and potential drug targets. Curr. Canc. Targets 2013, 13, 347–361. [Google Scholar]
- Sheppard, K.A.; Fitz, L.J.; Lee, J.M.; Benander, C.; George, J.A.; Wooters, J.; Qiu, Y.H.; Jussif, J.M.; Carter, L.L.; Wood, C.R.; et al. PD-1 inhibits T-cell receptor induced phosphorylation of the zap70/cd3zeta signalosome an downstream signaling to pkctheta. FEBS Lett. 2004, 574, 37–41. [Google Scholar] [CrossRef]
- Mahoney, K.M.; Rennert, P.D.; Freeman, G.J. Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 2015, 14, 561–584. [Google Scholar] [PubMed]
- Sun, A.; Fourcade, J.; Pagliano, O.; Chauvin, J.M.; Sander, C.; Kirkwood, J.M. IL10 and PD-1 cooperate to limit the activity of tumor-specific CD8+ T cells. Cancer Res. 2015, 75, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. J. Exp. Med. 2008, 8, 467–477. [Google Scholar] [CrossRef]
- Beckers, R.K.; Selinger, C.I.; Vilain, R.; Madore, J.; Wilmott, J.S.; Harvey, K.; Holliday, A.; Cooper, C.L.; Robbins, E.; Gillet, D.; et al. Programmed death ligand 1 expression in triple-negative breast cancer is associated with tumour-infiltrating lymphocytes and improved outcome. Histopathology 2016, 69, 25–34. [Google Scholar]
- Qin, T.; Zeng, Y.D.; Qin, G.; Xu, F.; Lu, J.B.; Fang, W.F.; Xue, C.; Zhan, J.H.; Zhang, X.K.; Zheng, Q.F.; et al. High PDL-1 expression was associated with poor prognosis in 870 Chinese patients with breast cancer. Oncotarget 2015, 6, 33972–33981. [Google Scholar] [CrossRef]
- Constantinou, C.; Papadopoulos, S.; Karyda, E.; Alexopoulos, A.; Agnanti, N.; Batistatou, A.; Harisi, H. Expression and clinical significance of claudin-7, PDL-1, PTEN, c-Kit, c-Met, c-Myc, ALK, CK5/6, CK17, p53, EFGR, Ki67, p63 in triple-negative breast cancer—A single centre prospective observational study. In Vivo 2018, 32, 303–311. [Google Scholar]
- Shosu, K.; Sakurai, M.; Inoue, K.; Nakagawa, T.; Sakai, H.; Morimoto, M.; Okuda, M.; Noguchi, S.; Mizuno, T. Programmed cell death ligand 1 expression in canine cancer. In Vivo 2016, 30, 195–204. [Google Scholar] [PubMed]
- Ariyarathna, H.; Thomson, N.A.; Aberdein, D.; Perrott, M.R.; Munday, J.S. Increased programmed death ligand (PD-L1) and cytotoxic T-lymphocyte antigen-4 (CTLA-4) expression is associated with metastasis and poor prognosis in malignant canine mammary gland tumours. Vet. Immunol. Immunopath. 2020, 230, 110142. [Google Scholar] [CrossRef] [PubMed]
- Lanuti, M.; Liu, G.; Goodwin, J.M.; Zhai, R.; Fuchs, B.C.; Asomaning, K.; Su, L.; Nishioka, N.S.; Tanabe, K.K.; Chistiani, D.C. A functional epidermal growth factor (eGF) polymorphism, EGF serum levels, and esophageal adenocarcinoma risk and outcome. Clin. Cancer Res. 2008, 14, 3216–3222. [Google Scholar] [CrossRef] [PubMed]
- Ciardiello, F.; Tortora, G. A novel approach in the treatment of cancer: Targeting the epidermal growth factor receptor. Clin. Cancer Res. 2001, 7, 2958–2970. [Google Scholar]
- Bublil, E.M.; Yarden, Y. The EGF receptor family: Spearheading a merger of signaling and therapeutics. Curr. Opin. Cell Biol. 2007, 19, 124–134. [Google Scholar] [CrossRef]
- Henson, E.S.; Gibson, S.B. Surviving cell death through epidermal growth factor (EGF) signal transduction pathways: Implications for cancer therapy. Cell. Signal. 2006, 18, 2089–2097. [Google Scholar] [CrossRef]
- Okamoto, I.; Roka, F.; Krögler, J.; Endler, G.; Kaufmann, S.; Tockner, S.; Marsik, C.; Jilma, B.; Mannhalter, C.; Wagner, O.; et al. The EGF A61G polymorphism is associated with disease-free period and survival in malignant melanoma. J. Investig. Dermatol. 2006, 126, 2242–2246. [Google Scholar]
- Kovar, F.M.; Thallinger, C.; Marsik, C.L.; Perkmann, T.; Puhalla, H.; Haslacher, H.; Wrba, F.; Wagner, O.F.; Gruenberger, T.; Endler, G. The EGF 61A/G polymorphism—A predictive marker for recurrence of liver metastases from colorectal cancer. Wien. Klin. Wochenscher. 2009, 121, 638–643. [Google Scholar]
- Araujo, A.P.; Costa, B.M.; Pinto-Correia, A.L.; Grafoso, M.; Ferreira, P.; Dinis-Ribeiro, M.; Costa, S.; Reix, R.M.; Medeiros, R. Association between EGF + 61ª/G polymorphism and gastric cancer in Caucasians. World. J. Gastroenterol. 2011, 17, 488–492. [Google Scholar]
- Kennedy, K.C.; Qurollo, B.A.; Rose, B.J.; Thamm, D.H. Epidermal growth factor enhances the malignant phenotype in canine mammary carcinoma cell lines. Vet. Comp. Oncol. 2011, 9, 196–206. [Google Scholar]
- Klopfleisch, R.; Lenze, D.; Hummel, M.; Gruber, A.D. The metastatic cascade is reflected in the transcriptome of metastatic canine mammary carcinomas. Vet. J. 2011, 190, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Queiroga, F.L.; Pérez-Alenza, D.; Silvan, G.; Peña, L.; Illera, J.C. Positive correlation of steroid hormones and EGF in canine mammary cancer. J. Ster. Biochem. Mol. Biol. 2009, 115, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Roche, P.J.; Gunnersen, J.M.; Hammond, V.E.; Tregear, G.W.; Wintour, E.M.; Beck, F. Mice without a functional relaxin gene are unable to deliver milk to their puppies. Endocrinology 1999, 140, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Samuel, C.S.; Tregear, G.W.; Beck, F.; Wintour, E.M. Collagen studies in late pregnant relaxin null mice. Biol. Reprod. 2000, 63, 697–703. [Google Scholar] [PubMed]
- Binder, C.; Hagemann, T.; Husen, B.; Schulz, B.; Einspanier, A. Relaxin enhances in-vitro invasiveness of breast cancer cell lines by up-regulation of matrix metalloproteases. Mol. Hum. Reprod. 2002, 8, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Binder, C.; Simon, A.; Binder, L.; Hagemann, T.; Schulz, M.; Emons, G.; Trumper, L.; Einspanier, A. Elevated concentrations of serum relaxin are associated with metastatic disease in breast cancer patients. Breast Cancer Res. Treat. 2004, 87, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Lamp, O.; Honscha, K.U.; Jakob, J.; Lamp, J.; Schweizer, S.; Reischauer, A.; Gottschalk, J.; Hahn, A.; Ebert, M.; Rothemund, S.; et al. Investigation of the local expression of the relaxin systemin canine mammary tumours. Reprod. Dom. Anim. 2009, 44 (Suppl. S2), 224–229. [Google Scholar]
- Lamp, O.; Honscha, K.U.; Schweizer, S.; Heckmann, A.; Blaschzik, S.; Einspanier, A. The metastatic potential of canine mammary tumours can be assessed by mRNA expression analysis of connective tissue modulators. Vet. Comp. Onc. 2013, 11, 70–85. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar]
- Rudolph-Owen, L.A.; Matrisian, L.M. Matrix metalloproteinases in remodeling of the normal and the neoplastic mammary gland. J. Mammary Gland Biol. Neoplasia 1998, 3, 177–189. [Google Scholar]
- Wiseman, B.S.; Sternlicht, M.D.; Lund, R.L.; Alexander, C.M.; Mott, J.; Bissell, P.S.; Itohara, S.; Werb, Z. Site-specific inductive and inhibitory activities of MMP-2 and MMP-3 orchestrate mammary gland branching morphogenesis. J. Cell. Biol. 2003, 162, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Hadler-Olsen, E.; Winberg, J.O.; Uhlin-Hansen, L. Matrix metalloproteinases in cancer: Their value as diagnostic and prognostic markers and therapeutic targets. Tumour Biol. 2013, 34, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Lochter, A.; Galosy, S.; Muschler, J.; Freedman, N.; Werb, Z.; Bissell, M.J. Matrix metalloproteinase stromelysin-1 triggers a cascade of molecular alterations that leads to stable epithelial-to-mesenchymal conversion and a premalignant phenotype in mammary epithelial cells. J. Cell Biol. 1997, 139, 1861–1872. [Google Scholar] [PubMed]
- Sternlicht, M.D.; Lochter, A.; Sympson, C.J.; Huey, B.; Rougier, J.P.; Gray, J.W.; Pinkel, D.; Bissell, M.J.; Werb, Z. The stromal proteinase MMP3/stromelysin-1 promotes mammary carcinogenesis. Cell 1999, 98, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; Maguire, T.M.; Hill, A.; McDermott, E.; O’Higgins, N. Metalloproteinases: A role in breast carcinogenesis invasion and metastasis. Breast Cancer Res. 2000, 2, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Torres-Mekia, G.; Stern, M.; Lundgreen, A.; Hines, L.; Giuliano, A.; Baumgartner, K.; Herrick, J.; Wolff, R.K. Matrix metalloproteinase genes are associated with breast cancer risk and survival: The Breast Cancer Health Disparities Study. PLoS ONE 2013, 8, e63165. [Google Scholar] [CrossRef]
- Mehner, C.; Miller, E.; Nassar, A.; Bamlet, W.R.; Radisky, E.S.; Radisky, D.C. Tumor cell expression of MMP3 as a prognostic factor for poor survival in pancreatic, pulmonary, and mammary carcinoma. Genes Cancer 2015, 6, 480–489. [Google Scholar]
- AbdRaboh, N.R.; Bayoumi, F.A. Gene polymorphism of matrix metalloproteinases 3 and 9 in breast cancer. Gene Rep. 2016, 5, 151–156. [Google Scholar] [CrossRef]
- Pandey, M.; Kumar, B.V.S.; Singh, S.; Verma, R. Development of recombinant matric metalloproteinase-3 based sandwich ELISA for sero-diagnosis of canine mammary carcinomas. J. Immunoass. Immunochem. 2016, 38, 34–44. [Google Scholar]
- Goel, H.L.; Mercurio, A.M. VEGF targets the tumour cell. Nat. Rev. Cancer 2013, 13, 871–882. [Google Scholar]
- Callahan, M.K.; Postow, M.A.; Wolchok, J.D. Targeting T cell co-receptors for cancer therapy. Immunity 2016, 44, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune checkpoint blockade: A common denominator approach to cancer therapy. Cancer Cell 2015, 27, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Aresu, L.; Ferraresso, S.; Marconato, L.; Cascione, L.; Napoli, S.; Gaudio, E.; Kwee, I.; Tarantelli, C.; Testa, A.; Maniaci, C.; et al. New molecular and therapeutic insights into canine diffuse large B cell lymphoma elucidates the role of the dog as a model for human disease. Haematologica 2019, 104, e256–e259. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, N.; Konnai, S.; Okagawa, T.; Nishimori, A.; Ikebuchi, R.; Izumi, Y.; Takagi, S.; Kagawa, Y.; Nakajima, C.; Suzuki, Y. Immunohistochemical analysis of PD-L1 expression in canine malignant cancers and PD-1 expression on lymphocytes in canine oral melanoma. PLoS ONE 2016, 11, e0157176. [Google Scholar] [CrossRef]
- Hudziak, R.M.; Lewis, G.D.; Winget, M.; Fendly, B.M.; Shepard, H.M.; Ullrich, A. p185HER2 monoclonal antibody has antiproliferative effects in vitro and sensitizes human breast tumor cells to tumor necrosis factor. Mol. Cell. Biol. 1989, 9, 1165–1172. [Google Scholar]
- Jacobs, A.T.; Martinez Castaneda-Cruz, D.; Rose, M.M.; Connelly, L. Targeted therapy for breast cancer: An overview of drug classes and outcomes. Biochem. Pharmacol. 2022, 204, 115209. [Google Scholar]
- Guil-Luna, S.; Sánchez-Céspedes, R.; Millán, Y.; De Andrés, F.J.; Rollón, E.; Domingo, V.; Guscetti, F.; Martín de las Mulas, J. Aglepristone decreases proliferation in progesterone receptor-positive canine mammary carcinomas. J. Vet. Intern. Mec. 2011, 25, 518–523. [Google Scholar] [CrossRef]
- Guil-Luna, S.; Millán, Y.; De Andrés, J.; Rollón, R.; Domingo, V.; García-Macías, J.; Sánchez-Céspedes, R.; Martín de las Mulas, J. Prognostic impact of neoadjuvant aglepristone treatment in clinicopathological parameters of progesterone receptor-positive canine mammary carcinomas. Vet. Comp. Oncol. 2017, 15, 391–399. [Google Scholar]
- Unemori, E.N.; Pickford, L.B.; Salles, A.L.; Piercy, C.E.; Grove, B.H.; Erikson, M.E.; Amento, E.P. Relaxin induces an extracellular matrix-degrading phenotype in human lung fibroblasts in vitro and inhibits lung fibrosis in a murine model in vivo. J. Clin. Investig. 1996, 98, 2739–2745. [Google Scholar] [CrossRef]
- Qin, X.; Chua, P.K.; Ohira, R.H.; Bryant-Greenwood, G.D. An autocrine/paracrine role of human decidual relaxin. II. Stromelysin-1 (MMP-3) and tissue inhibitor of matrix metalloproteinase-1 (TIMP-1). Biol. Reprod. 1997, 56, 812–820. [Google Scholar] [CrossRef]
- Kapila, S.; Xie, Y. Targeted induction of collagenase and stromelysin by relaxin in unprimed and beta-estradiol-primed diarthrodial joint fibrocartilaginous cells but not in synoviocytes. Lab. Investig. 1998, 78, 925–938. [Google Scholar] [PubMed]
- Lenhart, J.A.; Ryan, P.L.; Ohleth, K.; Palmer, S.S.; Bagnell, C.A. Relaxin increases secretion of matrix metalloproteinase-2 and matrix metalloproteinase-9 during uterine and cervical growth and remodelling in the pig. Endocrinology 2001, 142, 3941–3949. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, Y.S.; Kim, H.; Huang, J.H.; Yoon, A.R.; Yun, C.O. Relaxin expression from tumor-targeting adenoviruses and its intratumoral spread, apoptosis induction, and efficacy. J. Natl. Cancer Inst. 2006, 98, 1482–1493. [Google Scholar] [PubMed]
- Ganesh, S.; Gonzalez Edick, M.; Idamakanti, N.; Abramova, M.; Vanroey, M.; Robinson, M.; Yun, C.O.; Jooss, K. Relaxin-expressing, fiber chimeric oncolytic adenovirus prolongs survival of tumor-bearing mice. Cancer Res. 2007, 67, 4399–4407. [Google Scholar] [CrossRef]
- Li, Z.Y.; Liu, Y.; Tuve, S.; Xun, Y.; Fan, X.; Min, L.; Feng, Q.; Kiviat, N.; Kiem, H.P.; Disis, M.L.; et al. Toward a stem cell gene therapy for breast cancer. Blood 2009, 113, 5423–5433. [Google Scholar] [CrossRef]
- Tan, J.X.; Wang, X.Y.; Li, H.Y.; Su, X.L.; Wang, L.; Ran, L.; Zheng, K.; Ren, G.S. HYAL1 overexpression is correlated with the malignant behavior of human breast cancer. Int. J. Cancer 2011, 128, 1303–1315. [Google Scholar]
- Tan, J.X.; Wang, X.Y.; Su, X.L.; Li, H.Y.; Shi, Y.; Wang, L.; Ren, G.S. Upregulation of HYAL1 expression in breast cancer promoted tumor cell proliferation, migration, invasion and angiogenesis. PLoS ONE 2011, 7, e0022836. [Google Scholar] [CrossRef]


| Malignant (n = 21) | Benign (n = 38) | ||
|---|---|---|---|
| Typification | n | Typification | n |
| Complex adenocarcinoma | 2 | Adenoma | 36 |
| Complex carcinoma | 8 | Hemangioma | 1 |
| Solid carcinoma | 1 | ||
| Anaplastic carcinoma | 2 | ||
| Adenosquamous carcinoma | 1 | ||
| Tubullo-papillary carcinoma | 6 | ||
| Intraductal-papillary carcinoma | 1 | ||
| Gene | Slope | R2 | E* | E (%) |
|---|---|---|---|---|
| GADPH | −3.25 | 0.99 | 2.03 | 103 |
| HPRT | −3.11 | 0.99 | 2.05 | 105 |
| RPL8 | −3.27 | 0.94 | 2.02 | 102 |
| RPS19 | −3.25 | 0.99 | 2.03 | 103 |
| HYAL1 | −3.16 | 0.99 | 2.07 | 107 |
| MMP3 | −3.21 | 0.99 | 2.05 | 105 |
| RLN2 | −3.29 | 0.99 | 2.01 | 101 |
| VEGFA | −3.20 | 0.94 | 2.05 | 105 |
| PGR | −3.25 | 0.99 | 2.03 | 103 |
| CD20 | −3.29 | 0.98 | 2.02 | 102 |
| PD-L1 | −2.87 | 0.98 | 2.23 | 123 |
| EGF | −3.05 | 0.96 | 2.13 | 113 |
| VEGF-α | CD20 | PGR | HYAL1 | PD-L1 | EGF | RLN2 | MMP3 | |
|---|---|---|---|---|---|---|---|---|
| VEGFα | …. | |||||||
| CD20 | - | …. | ||||||
| PGR | - | 0.558 ** | …. | |||||
| HYAL1 | - | - | - | …. | ||||
| PD-L1 | - | - | - | - | …. | |||
| EGF | - | 0.790 *** | 0.502 ** | 0.418 * | - | …. | ||
| RLN2 | −0.467 ** | 0.546 ** | 0.435 * | - | - | 0.718 *** | …. | |
| MMP3 | −0.546 ** | - | 0.464 * | - | - | 0.500 ** | 0.507 ** | …. |
| VEGF-α | CD20 | PGR | HYAL1 | PD-L1 | EGF | RLN2 | MMP3 | |
|---|---|---|---|---|---|---|---|---|
| VEGFα | …. | |||||||
| CD20 | - | …. | ||||||
| PGR | - | 0.850 *** | …. | |||||
| HYAL1 | - | - | - | …. | ||||
| PD-L1 | - | - | - | −0.725 ** | …. | |||
| EGF | - | - | 0.702 *** | - | −0.566 * | …. | ||
| RLN2 | - | 0.725 ** | 0.754 *** | - | - | 0.547 * | …. | |
| MMP3 | - | 0.654 ** | 0.718 ** | - | - | 0.662 * | 0.718 ** | …. |
| Component | |||
|---|---|---|---|
| 1 | 2 | 3 | |
| VEGFα | −0.292 | 0.295 | 0.863 |
| CD20 | 0.838 | 0.136 | −0.132 |
| PGR | 0.809 | 0.047 | 0.096 |
| HYAL1 | 0.334 | 0.716 | −0.397 |
| PD-L1 | 0.296 | −0.812 | −0.046 |
| EGF | 0.825 | 0.111 | 0.278 |
| RLN2 | 0.884 | −0.152 | 0.151 |
| MMP3 | −0.292 | 0.295 | 0.863 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galadima, M.; Teles, M.; Pastor, J.; Hernández-Losa, J.; Rodríguez-Gil, J.E.; Rivera del Alamo, M.M. Programmed Death-Ligand (PD-L1), Epidermal Growth Factor (EGF), Relaxin, and Matrix Metalloproteinase-3 (MMP3): Potential Biomarkers of Malignancy in Canine Mammary Neoplasia. Int. J. Mol. Sci. 2024, 25, 1170. https://doi.org/10.3390/ijms25021170
Galadima M, Teles M, Pastor J, Hernández-Losa J, Rodríguez-Gil JE, Rivera del Alamo MM. Programmed Death-Ligand (PD-L1), Epidermal Growth Factor (EGF), Relaxin, and Matrix Metalloproteinase-3 (MMP3): Potential Biomarkers of Malignancy in Canine Mammary Neoplasia. International Journal of Molecular Sciences. 2024; 25(2):1170. https://doi.org/10.3390/ijms25021170
Chicago/Turabian StyleGaladima, Makchit, Mariana Teles, Josep Pastor, Javier Hernández-Losa, Joan Enric Rodríguez-Gil, and Maria Montserrat Rivera del Alamo. 2024. "Programmed Death-Ligand (PD-L1), Epidermal Growth Factor (EGF), Relaxin, and Matrix Metalloproteinase-3 (MMP3): Potential Biomarkers of Malignancy in Canine Mammary Neoplasia" International Journal of Molecular Sciences 25, no. 2: 1170. https://doi.org/10.3390/ijms25021170
APA StyleGaladima, M., Teles, M., Pastor, J., Hernández-Losa, J., Rodríguez-Gil, J. E., & Rivera del Alamo, M. M. (2024). Programmed Death-Ligand (PD-L1), Epidermal Growth Factor (EGF), Relaxin, and Matrix Metalloproteinase-3 (MMP3): Potential Biomarkers of Malignancy in Canine Mammary Neoplasia. International Journal of Molecular Sciences, 25(2), 1170. https://doi.org/10.3390/ijms25021170