

Editing Metabolism, Sex, and Microbiome: How Can We Help Poplar Resist Pathogens?

 ,
,  ,
,
Abstract
:
1. Introduction
2. A Brief History of the Genus Populus and a List of Poplar Pathogens

3. The Immunity of Poplars
3.1. A Brief Overview of the Major Plant Receptors and Signaling Pathways
3.2. Role of Primary and Secondary Metabolisms in Poplar Disease Resistance
3.3. PRs and Related Defensive Peptides and Proteins in Poplars
3.4. Transcription Factors Regulating the Immune Response
3.5. Contribution of microRNAs to the Regulation of Defense Responses
3.6. Intra-Population Differences in Resilience and Related Molecular Mechanisms
4. Environmental Factors Protecting Poplar from Infections: Endophytes, Phytophages and Chemical Elements
4.1. Endophytes
4.2. Phytophages
4.3. Elemental Defense Hypothesis
5. The Sex of the Tree as a Factor in Determining the Effectiveness of Plant Protection
6. Development of Sustainable Genetically Modified Poplars—Prospects and Challenges
6.1. Methods for Creating Genetically Modified Poplars and the Potential of CRISPR/Cas-Based Genomic Editing
- To influence the activity of transcription factors, mainly from the MYB and WRKY families, the overexpression of MYB factors that activate the biosynthesis of secondary metabolites can be used to increase the concentration of flavonoids in the plant. This can be achieved by Agrobacterium-mediated transformation. The knockout of MYBs that inhibit the biosynthesis of secondary metabolites can be accomplished using CRISPR/Cas. The overexpression of WRKYs from different families may increase disease resistance in poplar, and, although there is evidence that such overexpression in Arabidopsis may increase susceptibility to some pathogens due to SA-JA antagonism, this may not be an issue for poplar because SA and JA probably have a positive feedback in poplar.
- MicroRNA editing. It can be carried out through overexpression or STTM-mediated knockdown. Transgenic constructs can be introduced using agrobacteria. This field is just developing now, and there are only a few related works.
- MLO gene knockout. There are four candidate genes whose knockout can provide poplar resistance to powdery mildew: PtMLO17, 18, 19 and 24. Agrobacterium must be used for delivery, and the editing itself is performed using the CRISPR/Cas system.
- The knockout of genes from the SWEET, LOB, etc. families discussed in this chapter or already carried out on other plants. But to do this, it is necessary to find exactly those homologues in the poplar genome whose knockout will provide it with resistance to certain diseases. The protocol will be similar to that used for MLOs: transformation using agrobacteria and CRISPR/Cas-mediated knockout.
6.2. Possible Approaches to Engineering the White Poplar Microbiome
6.3. Problems and Perspectives Related to the Application of Genome Editing Techniques to Increase Poplar Resistance to Pathogens
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.J.; Wang, X.R.; Zeng, Q.Y. De novo assembly of white poplar genome and genetic diversity of white poplar population in Irtysh River basin in China. Sci. China Life Sci. 2019, 62, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Fuertes, A.; Oliveira, N.; Cañellas, I.; Sixto, H.; Rodríguez-Soalleiro, R. An Economic Overview of Populus Spp. in Short Rotation Coppice Systems under Mediterranean Conditions: An Assessment Tool for Decision-Making. Renew. Sustain. Energy Rev. 2021, 151, 111577. [Google Scholar] [CrossRef]
- Palmer, M.A.; Bernhardt, E.S.; Chornesky, E.A.; Collins, S.L.; Dobson, A.P.; Duke, C.S.; Gold, B.D.; Jacobson, R.B.; Kingsland, S.E.; Kranz, R.H.; et al. Ecological Science and Sustainability for the 21st Century. Front. Ecol. Environ. 2005, 3, 4–11. [Google Scholar] [CrossRef]
- Wang, Z.; MacFarlane, D.W. Evaluating the Biomass Production of Coppiced Willow and Poplar Clones in Michigan, USA, over Multiple Rotations and Different Growing Conditions. Biomass Bioenergy 2012, 46, 380–388. [Google Scholar] [CrossRef]
- Quiroga, A.; Marzocchi, V.; Rintoul, I. Influence of Wood Treatments on Mechanical Properties of Wood–Cement Composites and of Populus Euroamericana Wood Fibers. Compos. Part B Eng. 2016, 84, 25–32. [Google Scholar] [CrossRef]
- Rostampour Haftkhani, A.; Hematabadi, H. Effect of Layer Arrangement on Bending Strength of Cross-Laminated Timber (CLT) Manufactured from Poplar (Populus deltoides L.). Buildings 2022, 12, 608. [Google Scholar] [CrossRef]
- Wang, H.; Xue, Y.; Chen, Y.; Li, R.; Wei, J. Lignin Modification Improves the Biofuel Production Potential in Transgenic Populus Tomentosa. Ind. Crops Prod. 2012, 37, 170–177. [Google Scholar] [CrossRef]
- Biswal, A.K.; Hao, Z.; Pattathil, S.; Yang, X.; Winkeler, K.; Collins, C.; Mohanty, S.S.; Richardson, E.A.; Gelineo-Albersheim, I.; Hunt, K.; et al. Downregulation of GAUT12 in Populus Deltoides by RNA Silencing Results in Reduced Recalcitrance, Increased Growth and Reduced Xylan and Pectin in a Woody Biofuel Feedstock. Biotechnol. Biofuels 2015, 8, 41. [Google Scholar] [CrossRef]
- Macaya-Sanz, D.; Chen, J.; Kalluri, U.C.; Muchero, W.; Tschaplinski, T.J.; Gunter, L.E.; Simon, S.J.; Biswal, A.K.; Bryan, A.C.; Payyavula, R.; et al. Agronomic Performance of Populus Deltoides Trees Engineered for Biofuel Production. Biotechnol. Biofuels 2017, 10, 253. [Google Scholar] [CrossRef]
- Porth, I.; El-Kassaby, Y.A. Using Populus as a Lignocellulosic Feedstock for Bioethanol. Biotechnol. J. 2015, 10, 510–524. [Google Scholar] [CrossRef]
- Liberloo, M.; Calfapietra, C.; Lukac, M.; Godbold, D.; Luo, Z.; Polle, A.; Hoosbeek, M.R.; Kull, O.; Marek, M.; Raines, C.; et al. Woody Biomass Production during the Second Rotation of a Bio-energy Populus Plantation Increases in a Future High CO2 World. Glob. Chang. Biol. 2006, 12, 1094–1106. [Google Scholar] [CrossRef]
- Gao, S.; Chen, J.; Tang, Y.; Xie, J.; Zhang, R.; Tang, J.; Zhang, X. Ecosystem Carbon (CO2 and CH4) Fluxes of a Populus Dettoides Plantation in Subtropical China during and Post Clear-Cutting. For. Ecol. Manag. 2015, 357, 206–219. [Google Scholar] [CrossRef]
- Berlizov, A.N.; Blum, O.B.; Filby, R.H.; Malyuk, I.A.; Tryshyn, V.V. Testing Applicability of Black Poplar (Populus nigra L.) Bark to Heavy Metal Air Pollution Monitoring in Urban and Industrial Regions. Sci. Total Environ. 2007, 372, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Muftakhova, S.I.; Blonskaya, L.N.; Sabirzyanov, I.G.; Konashova, S.I.; Timeryanov, A. Age Dynamics of Growth and Development of Populus Pyramidalis in City Planting. Int. J. Environ. Stud. 2021, 78, 77–86. [Google Scholar] [CrossRef]
- Tőzsér, D.; Horváth, R.; Simon, E.; Magura, T. Heavy Metal Uptake by Plant Parts of Populus Species: A Meta-Analysis. Environ Sci. Pollut. Res. 2023, 30, 69416–69430. [Google Scholar] [CrossRef]
- Di Lonardo, S.; Capuana, M.; Arnetoli, M.; Gabbrielli, R.; Gonnelli, C. Exploring the Metal Phytoremediation Potential of Three Populus Alba L. Clones Using an In Vitro Screening. Environ. Sci. Pollut. Res. 2011, 18, 82–90. [Google Scholar] [CrossRef]
- Rafati, M.; Khorasani, N.; Moattar, F.; Shirvany, A.; Moraghebi, F.; Hosseinzadeh, S. Phytoremediation Potential of Populus Alba and Morus Alba for Cadmium, Chromuim and Nickel Absorption from Polluted Soil. Int. J. Environ. Res. 2011, 5, 961–970. [Google Scholar] [CrossRef]
- Guleria, I.; Kumari, A.; Lacaille-Dubois, M.-A.; Nishant; Kumar, V.; Saini, A.K.; Dhatwalia, J.; Lal, S. A Review on the Genus Populus: A Potential Source of Biologically Active Compounds. Phytochem. Rev. 2022, 21, 987–1046. [Google Scholar] [CrossRef]
- Tebbi, S.O.; Debbache-Benaida, N. Phytochemistry, Chemical Composition and Therapeutic Uses of Populus Nigra L. Aerial Parts from 1991–2021 Onwards: An Overview. Sustain. Chem. Pharm. 2022, 30, 100880. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The Genome of Black Cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef]
- Taylor, G. Populus: Arabidopsis for Forestry. Do We Need a Model Tree? Ann. Bot. 2002, 90, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.; Jansson, S.; Strauss, S.H.; Tuskan, G.A. Why and How Populus Became a “Model Tree”. In Genetics and Genomics of Populus; Jansson, S., Bhalerao, R., Groover, A., Eds.; Springer New York: New York, NY, USA, 2010; pp. 3–14. ISBN 978-1-4419-1540-5. [Google Scholar]
- Douglas, C.J. Populus as a Model Tree. In Comparative and Evolutionary Genomics of Angiosperm Trees; Groover, A., Cronk, Q., Eds.; Plant Genetics and Genomics: Crops and Models; Springer International Publishing: Cham, Switzerland, 2017; Volume 21, pp. 61–84. ISBN 978-3-319-49327-5. [Google Scholar]
- Zeng, Y.; Song, H.; Xia, L.; Yang, L.; Zhang, S. The Responses of Poplars to Fungal Pathogens: A Review of the Defensive Pathway. Front. Plant Sci. 2023, 14, 1107583. [Google Scholar] [CrossRef] [PubMed]
- Newcombe, G. A Review of Exapted Resistance to Diseases of Populus. Eur. J. For. Pathol. 1998, 28, 209–216. [Google Scholar] [CrossRef]
- Rossetti, M.F.; Stoker, C.; Ramos, J.G. Agrochemicals and Neurogenesis. Mol. Cell. Endocrinol. 2020, 510, 110820. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, J.; Gustavsson, M.; Dekker, S.C.; Van Wezel, A.P. Towards ‘One Substance—One Assessment’: An Analysis of EU Chemical Registration and Aquatic Risk Assessment Frameworks. J. Environ. Manag. 2021, 280, 111692. [Google Scholar] [CrossRef]
- Sundhar, S.; Shakila, R.J.; Jeyasekaran, G.; Aanand, S.; Shalini, R.; Arisekar, U.; Surya, T.; Malini, N.A.H.; Boda, S. Risk Assessment of Organochlorine Pesticides in Seaweeds along the Gulf of Mannar, Southeast India. Mar. Pollut. Bull. 2020, 161, 111709. [Google Scholar] [CrossRef]
- Zhang, L.; Xi, Z.; Wang, M.; Guo, X.; Ma, T. Plastome Phylogeny and Lineage Diversification of Salicaceae with Focus on Poplars and Willows. Ecol. Evol. 2018, 8, 7817–7823. [Google Scholar] [CrossRef]
- Zhang, Z.-S.; Zeng, Q.-Y.; Liu, Y.-J. Frequent Ploidy Changes in Salicaceae Indicates Widespread Sharing of the Salicoid Whole Genome Duplication by the Relatives of Populus L. and Salix L. BMC Plant Biol. 2021, 21, 535. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, L.; Zhang, Z.; Li, M.; Wang, D.; Zhang, X.; Xi, Z.; Keefover-Ring, K.; Smart, L.B.; DiFazio, S.P.; et al. Phylogenomics of the Genus Populus Reveals Extensive Interspecific Gene Flow and Balancing Selection. New Phytol. 2020, 225, 1370–1382. [Google Scholar] [CrossRef]
- Rösch, P.H.; Rösch, P. Chinese Wood Sculptures of the 11th to 13th Centuries: Images of Water-Moon Guanyin in Northern Chinese Temples and Western Collections; Ibidem-Verl: Stuttgart, Germany, 2007; ISBN 978-3-89821-662-3. [Google Scholar]
- Destination/Romania: Iasi—The County of Centuries-Old Trees. 4 September 2014. Available online: https://www.agerpres.ro/engleza-destinatie-romania/2014/09/04/destination-romania-iasi-the-county-of-centuries-old-trees-13-57-22 (accessed on 15 November 2023).
- Pourtet, J. Unasylva . 1951, 5. Available online: https://www.fao.org/3/x5359e/x5359e02.htm#the%20poplar%20%20%20its%20place%20in%20the%20world (accessed on 15 November 2023).
- Section 4—Poplar (Populus L.). In Safety Assessment of Transgenic Organisms, Volume 2; Harmonisation of Regulatory Oversight in Biotechnology; OECD: Paris, France, 2006; pp. 100–135. ISBN 978-92-64-09539-7.
- Zhang, B.; Zhu, W.; Diao, S.; Wu, X.; Lu, J.; Ding, C.; Su, X. The Poplar Pangenome Provides Insights into the Evolutionary History of the Genus. Commun. Biol. 2019, 2, 215. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Petrýdesová, J.; Bacigálová, K.; Sulo, P. The Reassignment of Three ‘Lost’ Taphrina Species (Taphrina Bullata, Taphrina Insititiae and Taphrina Rhizophora) Supported by the Divergence of Nuclear and Mitochondrial DNA. Int. J. Syst. Evol. Microbiol. 2013, 63, 3091–3098. [Google Scholar] [CrossRef] [PubMed]
- Spiers, A.G. Comparative Studies of Host Specificity and Symptoms Exhibited by Poplars Infected with Marssonina Brunnea, Marssonina Castagnei and Marssonina Populi. Eur. J. For. Pathol. 1984, 14, 202–218. [Google Scholar] [CrossRef]
- Newcombe, G. First Report of Pestalotiopsis Populi-Nigrae on Poplar in North America. Plant Dis. 2000, 84, 595. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Li, L.; Li, D.; Zhuge, Q. Characterization, Expression Profiling, and Functional Analysis of a Populus Trichocarpa Defensin Gene and Its Potential as an Anti-Agrobacterium Rooting Medium Additive. Sci. Rep. 2019, 9, 15359. [Google Scholar] [CrossRef] [PubMed]
- Chudinova, E.M.; Elansky, S.N. First Report of Septotinia Populiperda on Potato Tubers in Russia. J. Plant Pathol. 2021, 103, 665. [Google Scholar] [CrossRef]
- Bakhshi, M.; Arzanlou, M.; Zare, R.; Groenewald, J.Z.; Crous, P.W. New Species of Septoria Associated with Leaf Spot Diseases in Iran. Mycologia 2019, 111, 1056–1071. [Google Scholar] [CrossRef]
- Newcombe, G. Native Venturia Inopina Sp. Nov., Specific to Populus Trichocarpa and Its Hybrids. Mycol. Res. 2003, 107, 108–116. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria Parasitica, the Causal Agent of Chestnut Blight: Invasion History, Population Biology and Disease Control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef]
- Zhang, H.; Kong, N.; Ji, S.; Liu, B.; Tian, Z.; Qi, J.; Liu, Z. First Report of Leaf Blight Caused by Nigrospora oryzae on Poplar in China. Plant Dis. 2022, 106(3), 1063. [Google Scholar] [CrossRef]
- Lutter, R.; Drenkhan, R.; Tullus, A.; Jürimaa, K.; Tullus, T.; Tullus, H. First record of Entoleuca mammata in hybrid aspen plantations in hemiboreal Estonia and stand–environmental factors affecting its prevalence. Eur. J. For. Res. 2019, 138, 263–274. [Google Scholar] [CrossRef]
- Winton, L.M.; Adams, G.C.; Ruess, R.W. Determining the Novel Pathogen Neodothiora Populina as the Causal Agent of the Aspen Running Canker Disease in Alaska. Can. J. Plant Pathol. 2022, 44, 103–114. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, W.; Sun, N.; Wang, X.; Feng, Y.; Zhang, X. Identification and Functional Verification of Differences in Phenolic Compounds Between Resistant and Susceptible Populus Species. Phytopathology 2020, 110, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Unsicker, S.B.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Accumulation of Catechin and Proanthocyanidins in Black Poplar Stems After Infection by Plectosphaerella Populi: Hormonal Regulation, Biosynthesis and Antifungal Activity. Front. Plant Sci. 2019, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Ruess, R.W.; Winton, L.M.; Adams, G.C. Widespread Mortality of Trembling Aspen (Populus tremuloides) throughout Interior Alaskan Boreal Forests Resulting from a Novel Canker Disease. PLoS ONE 2021, 16, e0250078. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cheng, Q. Heterologous Expression of Arabidopsis Pattern Recognition Receptor RLP23 Increases Broad-spectrum Resistance in Poplar to Fungal Pathogens. Mol. Plant Pathol. 2023, 24, 80–86. [Google Scholar] [CrossRef]
- Dawwam, G.E.; Sehim, A.E. Promising Biological Agents Represented in Bacillus Velezensis 33RB and Aspergillus Niger 46SF Endophytic Isolates for Controlling Populus Tomentosa Wilt and Anthracnose Diseases. Egypt. J. Biol. Pest Control 2022, 32, 144. [Google Scholar] [CrossRef]
- Wang, L.; Ran, L.; Hou, Y.; Tian, Q.; Li, C.; Liu, R.; Fan, D.; Luo, K. The Transcription Factor MYB115 Contributes to the Regulation of Proanthocyanidin Biosynthesis and Enhances Fungal Resistance in Poplar. New Phytol. 2017, 215, 351–367. [Google Scholar] [CrossRef]
- Su, T.; Zhou, B.; Cao, D.; Pan, Y.; Hu, M.; Zhang, M.; Wei, H.; Han, M. Transcriptomic Profiling of Populus Roots Challenged with Fusarium Reveals Differential Responsive Patterns of Invertase and Invertase Inhibitor-Like Families within Carbohydrate Metabolism. J. Fungi 2021, 7, 89. [Google Scholar] [CrossRef]
- Newcombe, G.; Fraser, S.J.; Ridout, M.; Busby, P.E. Leaf Endophytes of Populus Trichocarpa Act as Pathogens of Neighboring Plant Species. Front. Microbiol. 2020, 11, 573056. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.L.; Ziegler, D.J.; Giesbrecht, S.; McLoughlin, A.; Wan, J.; Khan, D.; Hoi, V.; Whyard, S.; Belmonte, M.F. Control of White Mold (Sclerotinia sclerotiorum) through Plant-Mediated RNA Interference. Sci. Rep. 2023, 13, 6477. [Google Scholar] [CrossRef]
- Vialle, A.; Frey, P.; Hambleton, S.; Bernier, L.; Hamelin, R.C. Poplar Rust Systematics and Refinement of Melampsora Species Delineation. Fungal Divers. 2011, 50, 227–248. [Google Scholar] [CrossRef]
- Hu, Q.; Wang, H.; Ng, T.B. Isolation and Purification of Polysaccharides with Anti-Tumor Activity from Pholiota adiposa (Batsch) P. Kumm. (Higher Basidiomycetes). Int. J. Med. Mushr. 2012, 14, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hou, R.; Yan, J.; Xu, K.; Wu, X.; Lin, W.; Zheng, M.; Fu, J. Purification and Characterization of Inonotus Hispidus Exopolysaccharide and Its Protective Effect on Acute Alcoholic Liver Injury in Mice. Int. J. Biol. Macromol. 2019, 129, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Striganavičiūtė, G.; Žiauka, J.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Impact of Plant-Associated Bacteria on the In Vitro Growth and Pathogenic Resistance against Phellinus Tremulae of Different Aspen (Populus) Genotypes. Microorganisms 2021, 9, 1901. [Google Scholar] [CrossRef]
- Zhou, L.-W.; Vlasák, J.; Qin, W.-M.; Dai, Y.-C. Global Diversity and Phylogeny of the Phellinus Igniarius Complex (Hymenochaetales, Basidiomycota) with the Description of Five New Species. Mycologia 2016, 108, 192–204. [Google Scholar] [CrossRef]
- Vasaitis, R.; Menkis, A.; Lim, Y.W.; Seok, S.; Tomsovsky, M.; Jankovsky, L.; Lygis, V.; Slippers, B.; Stenlid, J. Genetic Variation and Relationships in Laetiporus Sulphureus s. Lat., as Determined by ITS rDNA Sequences and in Vitro Growth Rate. Mycol. Res. 2009, 113, 326–336. [Google Scholar] [CrossRef]
- Minter, D.W.; Soliman, G.S. Laetiporus sulphureus. [Descriptions of Fungi and Bacteria]. Descr. Fungi Bact. 2022, 231, 2297. [Google Scholar] [CrossRef]
- Schubert, M.; Fink, S.; Schwarze, F.W.M.R. Evaluation of Trichoderma Spp. as a Biocontrol Agent against Wood Decay Fungi in Urban Trees. Biol. Control 2008, 45, 111–123. [Google Scholar] [CrossRef]
- Badalyan, S.; Zhuykova, E.; Mukhin, V. The Phylogenetic Analysis of Armenian Collections of Medicinal Tinder Polypore Fomes Fomentarius (Agaricomycetes, Polyporaceae). Ital. J. Mycol. 2022, 51, 23–33. [Google Scholar] [CrossRef]
- Szczepkowski, A.; Kowalczuk, W. Current Conservation Status of the Fungus Spongipellis Spumeus in Poland Revised Based on New Data. Pol. J. Ecol. 2020, 68, 1. [Google Scholar] [CrossRef]
- Doty, S.L.; Joubert, P.M.; Firrincieli, A.; Sher, A.W.; Tournay, R.; Kill, C.; Parikh, S.S.; Okubara, P. Potential Biocontrol Activities of Populus Endophytes against Several Plant Pathogens Using Different Inhibitory Mechanisms. Pathogens 2022, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.-H.; Ryvarden, L.; et al. A Revised Family-Level Classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Matheny, P.B. The Relative Ages of Ectomycorrhizal Mushrooms and Their Plant Hosts Estimated Using Bayesian Relaxed Molecular Clock Analyses. BMC Biol. 2009, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Ekanayaka, A. Preliminary Classification of Leotiomycetes. Mycosphere 2019, 10, 310–489. [Google Scholar] [CrossRef]
- Baral, H.-O.; Rönsch, P.; Richter, U.; Urban, A.; Kruse, J.; Bemmann, M.; Kummer, V.; Valencia, F.J.; Huth, W. Schroeteria decaisneana, S. poeltii, and Ciboria ploettneriana (Sclerotiniaceae, Helotiales, Ascomycota), Three Parasites on Veronica Seeds: First Report of Teleomorphs in Schroeteria. Mycol. Prog. 2022, 21, 359–407. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of Lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef]
- Ismail, S.I.; Batzer, J.C.; Harrington, T.C.; Crous, P.W.; Lavrov, D.V.; Li, H.; Gleason, M.L. Ancestral State Reconstruction Infers Phytopathogenic Origins of Sooty Blotch and Flyspeck Fungi on Apple. Mycologia 2016, 108, 292–302. [Google Scholar] [CrossRef]
- Lackus, N.D.; Morawetz, J.; Xu, H.; Gershenzon, J.; Dickschat, J.S.; Köllner, T.G. The Sesquiterpene Synthase PtTPS5 Produces (1S,5S,7R,10R)-Guaia-4(15)-En-11-Ol and (1S,7R,10R)-Guaia-4-En-11-Ol in Oomycete-Infected Poplar Roots. Molecules 2021, 26, 555. [Google Scholar] [CrossRef]
- Ichinose, Y.; Taguchi, F.; Mukaihara, T. Pathogenicity and Virulence Factors of Pseudomonas Syringae. J. Gen. Plant Pathol. 2013, 79, 285–296. [Google Scholar] [CrossRef]
- Goychuk, A.; Kulbanska, I.; Shvets, M.; Pasichnyk, L.; Patyka, V.; Kalinichenko, A.; Degtyareva, L. Bacterial Diseases of Bioenergy Woody Plants in Ukraine. Sustainability 2023, 15, 4189. [Google Scholar] [CrossRef]
- Nesme, X. Differential Host-Pathogen Interactions Among Clones of Poplar and Strains of Xanthomonas populi Pv. populi. Phytopathology 1994, 84, 101. [Google Scholar] [CrossRef]
- Li, A.; He, W. Molecular Aspects of an Emerging Poplar Canker Caused by Lonsdalea populi. Front. Microbiol. 2019, 10, 2496. [Google Scholar] [CrossRef] [PubMed]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Marković, M.; Orlović, S. Bacterial Canker Disease on Populus × Euramericana Caused by Lonsdalea populi in Serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- Tóth, T.; Lakatos, T.; Koltay, A. Lonsdalea Quercina Subsp. Populi Subsp. Nov., Isolated from Bark Canker of Poplar Trees. Int. J. Syst. Evol. Microbiol. 2013, 63, 2309–2313. [Google Scholar] [CrossRef] [PubMed]
- Abelleira, A.; Moura, L.; Aguín, O.; Salinero, C. First Report of Lonsdalea populi Causing Bark Canker Disease on Poplar in Portugal. Plant Dis. 2019, 103, 2121. [Google Scholar] [CrossRef]
- Li, Y.; He, W.; Ren, F.; Guo, L.; Chang, J.; Cleenwerck, I.; Ma, Y.; Wang, H. A Canker Disease of Populus × Euramericana in China Caused by Lonsdalea quercina Subsp. Populi. Plant Dis. 2014, 98, 368–378. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, W.; Wang, M.; Cheng, Q. Cloning and Characterization of the PtVIP1 Gene in Populus. J. For. Res. 2019, 30, 2259–2266. [Google Scholar] [CrossRef]
- Naylor, M.; Reeves, J.; Cooper, J.I.; Edwards, M.-L.; Wang, H. Construction and Properties of a Gene-Silencing Vector Based on Poplar Mosaic Virus (Genus Carlavirus). J. Virol. Methods 2005, 124, 27–36. [Google Scholar] [CrossRef]
- Smith, C.M.; Campbell, M.M. Populus Genotypes Differ in Infection by, and Systemic Spread of, Poplar mosaic virus. Plant Pathol. 2004, 53, 780–787. [Google Scholar] [CrossRef]
- Von Bargen, S.; Al Kubrusli, R.; Gaskin, T.; Fürl, S.; Hüttner, F.; Blystad, D.; Karlin, D.G.; Jalkanen, R.; Büttner, C. Characterisation of a Novel Emaravirus Identified in Mosaic-diseased Eurasian Aspen (Populus tremula). Ann. Appl. Biol. 2020, 176, 210–222. [Google Scholar] [CrossRef]
- Hibben, C.R.; Bozarth, R.F.; Reese, J. Identification of Tobacco Necrosis Virus in Deteriorating Clones of Aspen. For. Sci. 1979, 25, 557–567. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef]
- Martin, R.R. Isolation of a Potyvirus from Declining Clones of Populus. Phytopathology 1982, 72, 1158. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, W.; Shen, W.; Li, M.; Fu, Y.; Li, Z.; Li, J.; Liu, H.; Su, X.; Zhang, B.; et al. Integrated Transcriptome and microRNA Sequencing Analyses Reveal Gene Responses in Poplar Leaves Infected by the Novel Pathogen Bean Common Mosaic Virus (BCMV). Front. Plant Sci. 2023, 14, 1163232. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, R.; Hamza, M.; Rehman, A.U.; Kamal, H.; Tahir, M.N.; Mansoor, S.; Scheffler, B.E.; Briddon, R.W.; Amin, I. Asymptomatic Populus Alba: A Tree Serving as a Reservoir of Begomoviruses and Associated Satellites. Australas. Plant Pathol. 2022, 51, 577–586. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Hwang, S.-Y.; Osier, T.L. Phytochemical Variation in Quaking Aspen: Effects on Gypsy Moth Susceptibility to Nuclear Polyhedrosis Virus. J. Chem. Ecol. 1999, 25, 1331–1341. [Google Scholar] [CrossRef]
- Lo, C.-C.; Bonner, C.A.; Xie, G.; D’Souza, M.; Jensen, R.A. Cohesion Group Approach for Evolutionary Analysis of Aspartokinase, an Enzyme That Feeds a Branched Network of Many Biochemical Pathways. Microbiol. Mol. Biol. Rev. 2009, 73, 594–651. [Google Scholar] [CrossRef]
- Wolf, Y.I.; Silas, S.; Wang, Y.; Wu, S.; Bocek, M.; Kazlauskas, D.; Krupovic, M.; Fire, A.; Dolja, V.V.; Koonin, E.V. Doubling of the Known Set of RNA Viruses by Metagenomic Analysis of an Aquatic Virome. Nat. Microbiol. 2020, 5, 1262–1270. [Google Scholar] [CrossRef]
- Zhao, J.P.; Su, X.H. Patterns of Molecular Evolution and Predicted Function in Thaumatin-like Proteins of Populus trichocarpa. Planta 2010, 232, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Li, Y.; Chen, Y.; Zhang, X.; Guan, H.; Yin, T. Melampsora Larici-Populina, the Main Rust Pathogen, Causes Loss in Biomass Production of Black Cottonwood Plantations in the South of China. Phytoparasitica 2013, 41, 337–344. [Google Scholar] [CrossRef]
- Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced Breeding for Biotic Stress Resistance in Poplar. Plants 2022, 11, 2032. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Petre, B.; Hacquard, S.; Duplessis, S.; Rouhier, N. Genome Analysis of Poplar LRR-RLP Gene Clusters Reveals RISP, a Defense-Related Gene Coding a Candidate Endogenous Peptide Elicitor. Front. Plant Sci. 2014, 5, 111. [Google Scholar] [CrossRef] [PubMed]
- Cope, K.R.; Prates, E.T.; Miller, J.I.; Demerdash, O.N.A.; Shah, M.; Kainer, D.; Cliff, A.; Sullivan, K.A.; Cashman, M.; Lane, M.; et al. Exploring the Role of Plant Lysin Motif Receptor-like Kinases in Regulating Plant-Microbe Interactions in the Bioenergy Crop Populus. Comput. Struct. Biotechnol. J. 2023, 21, 1122–1139. [Google Scholar] [CrossRef]
- Muhr, M. Characterization of Populus x Canescens LysM Receptor-Like Kinases CERK1-1 and CERK1-2 and Their Role in Chitin Signaling. Ph.D. Thesis, Georg-August-University Göttingen, Göttingen, Germany, 2022. [Google Scholar]
- Zhao, Y.; Zheng, X.; Tabima, J.F.; Sondreli, K.; Zhu, S.; Hundley, H.; Bauer, D.; Barry, K.; Zhang, Y.; Schmutz, J.; et al. Secreted Effector Proteins of the Poplar Leaf Spot and Stem Canker Pathogen Sphaerulina musiva Manipulate Plant Immunity and Contribute to Virulence in Diverse Ways. Mol. Plant-Microbe Interact. 2023, 36, 779–795. [Google Scholar] [CrossRef]
- Germain, H.; Séguin, A. Innate Immunity: Has Poplar Made Its BED? New Phytol. 2011, 189, 678–687. [Google Scholar] [CrossRef]
- Marchal, C.; Zhang, J.; Zhang, P.; Fenwick, P.; Steuernagel, B.; Adamski, N.M.; Boyd, L.; McIntosh, R.; Wulff, B.B.H.; Berry, S.; et al. BED-Domain-Containing Immune Receptors Confer Diverse Resistance Spectra to Yellow Rust. Nat. Plants 2018, 4, 662–668. [Google Scholar] [CrossRef]
- Plett, J.M.; Daguerre, Y.; Wittulsky, S.; Vayssières, A.; Deveau, A.; Melton, S.J.; Kohler, A.; Morrell-Falvey, J.L.; Brun, A.; Veneault-Fourrey, C.; et al. Effector MiSSP7 of the Mutualistic Fungus Laccaria Bicolor Stabilizes the Populus JAZ6 Protein and Represses Jasmonic Acid (JA) Responsive Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 8299–8304. [Google Scholar] [CrossRef]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D. Salicylic Acid Signaling in Disease Resistance. Plant Sci. 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Chen, Y.-H.; Ortega, M.A.; Tsai, C.-J. The Diversity of Salicylic Acid Biosynthesis and Defense Signaling in Plants: Knowledge Gaps and Future Opportunities. Curr. Opin. Plant Biol. 2023, 72, 102349. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, G.; Cao, C.; Liu, P. Metabolism, Signaling, and Transport of Jasmonates. Plant Commun. 2021, 2, 100231. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate Action in Plant Growth and Development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef]
- Dubois, M.; Van Den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughan-Hirsch, J.; Van De Poel, B. The Regulation of Ethylene Biosynthesis: A Complex Multilevel Control Circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene Signaling in Plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Yang, F.; Chai, S.; Wang, L.; De Dios, V.R.; Tan, W.; Yao, Y. Ethylene Activates Poplar Defense against Dothiorella gregaria Sacc. by Regulating Reactive Oxygen Species Accumulation. Physiol. Plant. 2022, 174, e13726. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Tsai, C.; Unsicker, S.B.; Xue, L.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Salicylic Acid Activates Poplar Defense against the Biotrophic Rust Fungus Melampsora Larici-populina via Increased Biosynthesis of Catechin and Proanthocyanidins. New Phytol. 2019, 221, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Schmidt, A.; Reichelt, M.; Tsai, C.; Gershenzon, J. Lack of Antagonism between Salicylic Acid and Jasmonate Signalling Pathways in Poplar. New Phytol. 2022, 235, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.A. Systemic Acquired Resistance and Salicylic Acid: Past, Present, and Future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef] [PubMed]
- Balint-Kurti, P. The Plant Hypersensitive Response: Concepts, Control and Consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef]
- Dalio, R.J.D.; Paschoal, D.; Arena, G.D.; Magalhães, D.M.; Oliveira, T.S.; Merfa, M.V.; Maximo, H.J.; Machado, M.A. Hypersensitive Response: From NLR Pathogen Recognition to Cell Death Response. Ann. Appl. Biol. 2021, 178, 268–280. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, M.; Zhou, H.; Zhou, X.; Wang, Y. Metabolite Profiles of Populus in Response to Pathogen Stress. Biochem. Biophys. Res. Commun. 2015, 465, 421–426. [Google Scholar] [CrossRef]
- Su, T.; Han, M.; Min, J.; Zhou, H.; Zhang, Q.; Zhao, J.; Fang, Y. Functional Characterization of Invertase Inhibitors PtC/VIF1 and 2 Revealed Their Involvements in the Defense Response to Fungal Pathogen in Populus trichocarpa. Front. Plant Sci. 2020, 10, 1654. [Google Scholar] [CrossRef]
- Siemens, J.; González, M.; Wolf, S.; Hofmann, C.; Greiner, S.; Du, Y.; Rausch, T.; Roitsch, T.; Ludwig-Müller, J. Extracellular Invertase Is Involved in the Regulation of Clubroot Disease in Arabidopsis thaliana. Mol. Plant Pathol. 2011, 12, 247–262. [Google Scholar] [CrossRef]
- Tauzin, A.S.; Giardina, T. Sucrose and Invertases, a Part of the Plant Defense Response to the Biotic Stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Veillet, F.; Gaillard, C.; Coutos-Thévenot, P.; La Camera, S. Targeting the AtCWIN1 Gene to Explore the Role of Invertases in Sucrose Transport in Roots and during Botrytis Cinerea Infection. Front. Plant Sci. 2016, 7, 1899. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Han, M.; Min, J.; Chen, P.; Mao, Y.; Huang, Q.; Tong, Q.; Liu, Q.; Fang, Y. Genome-Wide Survey of Invertase Encoding Genes and Functional Characterization of an Extracellular Fungal Pathogen-Responsive Invertase in Glycine Max. Int. J. Mol. Sci. 2018, 19, 2395. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Liu, J.; Lin, X.; Hu, S.; Yang, Y.; Li, D.; Chen, L.; Huai, B.; Huang, L.; Voegele, R.T.; et al. A Unique Invertase Is Important for Sugar Absorption of an Obligate Biotrophic Pathogen during Infection. New Phytol. 2017, 215, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xu, X.; Xiong, Y.; Wei, H.; Yao, K.; Huang, T.; Long, Y.; Su, T. Genome-Wide Survey and Expression Analyses of Hexokinase Family in Poplar (Populus trichocarpa). Plants 2022, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, W.; Zhang, Y.; Xing, J.; Li, J.; Feng, J.; Su, X.; Zhao, J. Fungal Canker Pathogens Trigger Carbon Starvation by Inhibiting Carbon Metabolism in Poplar Stems. Sci. Rep. 2019, 9, 10111. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Li, P.; Zhang, Y.; Li, J.; Liu, Y.; Lachenbruch, B.; Su, X.; Zhao, J. Fungal Pathogens of Canker Disease Trigger Canopy Dieback in Poplar Saplings by Inducing Functional Failure of the Phloem and Cambium and Carbon Starvation in the Xylem. Physiol. Mol. Plant Pathol. 2020, 112, 101523. [Google Scholar] [CrossRef]
- Han, M.; Xu, X.; Li, X.; Xu, M.; Hu, M.; Xiong, Y.; Feng, J.; Wu, H.; Zhu, H.; Su, T. New Insight into Aspartate Metabolic Pathways in Populus: Linking the Root Responsive Isoenzymes with Amino Acid Biosynthesis during Incompatible Interactions of Fusarium Solani. Int. J. Mol. Sci. 2022, 23, 6368. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of Secondary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef]
- Movahedi, A.; Almasi Zadeh Yaghuti, A.; Wei, H.; Rutland, P.; Sun, W.; Mousavi, M.; Li, D.; Zhuge, Q. Plant Secondary Metabolites with an Overview of Populus. Int. J. Mol. Sci. 2021, 22, 6890. [Google Scholar] [CrossRef]
- Weng, J.; Chapple, C. The Origin and Evolution of Lignin Biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.-G.; Muchero, W. Regulation of Lignin Biosynthesis and Its Role in Growth-Defense Tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Escamilla-Trevino, L.; Song, L.; Rao, X.; Serrani-Yarce, J.C.; Palacios, M.D.; Engle, N.; Choudhury, F.K.; Tschaplinski, T.J.; Venables, B.J.; et al. 4-Coumarate 3-Hydroxylase in the Lignin Biosynthesis Pathway Is a Cytosolic Ascorbate Peroxidase. Nat. Commun. 2019, 10, 1994. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Barros, J. Lignin Biosynthesis: Old Roads Revisited and New Roads Explored. Open Biol. 2019, 9, 190215. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Kamimura, N.; Tokue, Y.; Nakata, M.T.; Yamamoto, M.; Hu, S.; Masai, E.; Mitsuda, N.; Kajita, S. Identification of Enzymatic Genes with the Potential to Reduce Biomass Recalcitrance through Lignin Manipulation in Arabidopsis. Biotechnol. Biofuels 2020, 13, 97. [Google Scholar] [CrossRef] [PubMed]
- Bagniewska-Zadworna, A.; Barakat, A.; Łakomy, P.; Smoliński, D.J.; Zadworny, M. Lignin and Lignans in Plant Defence: Insight from Expression Profiling of Cinnamyl Alcohol Dehydrogenase Genes during Development and Following Fungal Infection in Populus. Plant Sci. 2014, 229, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Gunter, L.E.; Ranjan, P.; Singan, V.R.; Engle, N.; et al. Genome-wide association studies and Expression-based Quantitative Trait Loci Analyses Reveal Roles of HCT 2 in Caffeoylquinic Acid Biosynthesis and Its Regulation by Defense-responsive Transcription Factors in Populus. New Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef]
- Yang, W.; Fortunati, E.; Gao, D.; Balestra, G.M.; Giovanale, G.; He, X.; Torre, L.; Kenny, J.M.; Puglia, D. Valorization of Acid Isolated High Yield Lignin Nanoparticles as Innovative Antioxidant/Antimicrobial Organic Materials. ACS Sustain. Chem. Eng. 2018, 6, 3502–3514. [Google Scholar] [CrossRef]
- Chen, M.; Li, Y.; Liu, H.; Zhang, D.; Shi, Q.-S.; Zhong, X.-Q.; Guo, Y.; Xie, X.-B. High Value Valorization of Lignin as Environmental Benign Antimicrobial. Mater. Today Bio 2023, 18, 100520. [Google Scholar] [CrossRef]
- Li, L.; Sun, W.; Wang, P.; Li, H.; Rehman, S.; Li, D.; Zhuge, Q. Characterization, Expression, and Functional Analysis of the Pathogenesis-Related Gene PtDIR11 in Transgenic Poplar. Int. J. Biol. Macromol. 2022, 210, 182–195. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Zhou, P.; Yang, X.; Tan, W.; Sun, W.; Zhuge, Q. Cloning and Expression Analysis of PeDIR19 Gene in Poplar. Plant Gene Trait 2023, 14, 1–10. [Google Scholar] [CrossRef]
- Liang, J.; Huang, X.; Ma, G. Antimicrobial Activities and Mechanisms of Extract and Components of Herbs in East Asia. RSC Adv. 2022, 12, 29197–29213. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, W.; Dong, H.; Liu, Z.; Ma, J.; Zhang, X. Salicylic Acid in Populus Tomentosa Is a Remote Signalling Molecule Induced by Botryosphaeria Dothidea Infection. Sci. Rep. 2018, 8, 14059. [Google Scholar] [CrossRef] [PubMed]
- Palma Ferreira, S. Populus euphratica: An Incompatible Host for Biotrophic Pathogens? Mol. Plant Pathol. 2016, 17, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, X.; Meng, Z.; Liu, Y. Efficacy of Topical 30% Salicylic Acid in Combination with Minocycline in Moderate to Severe Acne. Pak. J. Pharm. Sci. 2023, 36, 607–611. [Google Scholar] [PubMed]
- Cooper, B. The Detriment of Salicylic Acid to the Pseudomonas Savastanoi Pv. Phaseolicola Proteome. Mol. Plant-Microbe Interact. 2022, 35, 814–824. [Google Scholar] [CrossRef]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal Activity of Salicylic Acid against Penicillium Expansum and Its Possible Mechanisms of Action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef]
- Li, L.; Zhu, T.; Song, Y.; Feng, L.; Kear, P.J.; Riseh, R.S.; Sitohy, M.; Datla, R.; Ren, M. Salicylic Acid Fights against Fusarium Wilt by Inhibiting Target of Rapamycin Signaling Pathway in Fusarium oxysporum. J. Adv. Res. 2022, 39, 1–13. [Google Scholar] [CrossRef]
- Salgado Salomón, M.E.; Dresch, P.; Horak, E.; Galleguillos, F.; Barroetaveña, C.; Peintner, U. The Enigmatic Cortinarius magellanicus Complex Occurring in Nothofagaceae Forests of the Southern Hemisphere. Fungal Biol. 2018, 122, 1077–1097. [Google Scholar] [CrossRef]
- Molla, K.A. CRISPR-Cas9 Helps Solve a Piece of the Puzzle of the Biosynthesis of Salicinoids and Suggests a Role in the Growth-Defense Trade-off in Poplar. Plant Cell 2022, 34, 2819–2820. [Google Scholar] [CrossRef]
- Saint-Vincent, P.M.; Ridout, M.; Engle, N.L.; Lawrence, T.J.; Yeary, M.L.; Tschaplinski, T.J.; Newcombe, G.; Pelletier, D.A. Isolation, Characterization, and Pathogenicity of Two Pseudomonas syringae Pathovars from Populus trichocarpa Seeds. Microorganisms 2020, 8, 1137. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Gauthier, C.; Legault, J.; Lavoie, S.; Mshvildadze, V.; Pichette, A. Structure Elucidation of Anti-Methicillin Resistant Staphylococcus Aureus (MRSA) Flavonoids from Balsam Poplar Buds. Bioorg. Med. Chem. 2016, 24, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Nur, F.A.; Ma, J.; Wang, J.; Cao, C. Effects of Poplar Secondary Metabolites on Performance and Detoxification Enzyme Activity of Lymantria Dispar. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108587. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, M.; Lam, P.-Y.; Dini-Andreote, F.; Dai, L.; Wei, Z. Multifaceted Roles of Flavonoids Mediating Plant-Microbe Interactions. Microbiome 2022, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Yan, J.; Fu, Z.; Yang, T.; Ziemah, J.; Ullrich, M.S.; Kuhnert, N.; Zeng, H. Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi. J. Fungi 2022, 9, 52. [Google Scholar] [CrossRef]
- Naoumkina, M.A.; Zhao, Q.; Gallego-Giraldo, L.; Dai, X.; Zhao, P.X.; Dixon, R.A. Genome-wide Analysis of Phenylpropanoid Defence Pathways. Mol. Plant Pathol. 2010, 11, 829–846. [Google Scholar] [CrossRef]
- St-Pierre, A.; Blondeau, D.; Lajeunesse, A.; Bley, J.; Bourdeau, N.; Desgagné-Penix, I. Phytochemical Screening of Quaking Aspen (Populus tremuloides) Extracts by UPLC-QTOF-MS and Evaluation of Their Antimicrobial Activity. Molecules 2018, 23, 1739. [Google Scholar] [CrossRef]
- Abou Baker, D.H. An Ethnopharmacological Review on the Therapeutical Properties of Flavonoids and Their Mechanisms of Actions: A Comprehensive Review Based on up to Date Knowledge. Toxicol. Rep. 2022, 9, 445–469. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Wu, S.-C.; Yang, Z.-Q.; Liu, F.; Peng, W.-J.; Qu, S.-Q.; Li, Q.; Song, X.-B.; Zhu, K.; Shen, J.-Z. Antibacterial Effect and Mode of Action of Flavonoids From Licorice Against Methicillin-Resistant Staphylococcus Aureus. Front. Microbiol. 2019, 10, 2489. [Google Scholar] [CrossRef] [PubMed]
- Biharee, A.; Sharma, A.; Kumar, A.; Jaitak, V. Antimicrobial Flavonoids as a Potential Substitute for Overcoming Antimicrobial Resistance. Fitoterapia 2020, 146, 104720. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Unsicker, S.B.; Fellenberg, C.; Constabel, C.P.; Schmidt, A.; Gershenzon, J.; Hammerbacher, A. Flavan-3-Ols Are an Effective Chemical Defense against Rust Infection. Plant Physiol. 2017, 175, 1560–1578. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.; Muchero, W.; Weber, R.A.; Barros, J.; Chen, J.-G.; Tschaplinski, T.J.; Pu, Y.; Ragauskas, A.J. Cell Wall Response of Field Grown Populus to Septoria Infection. Front. Plant Sci. 2023, 14, 1089011. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wang, L.; Han, Z.; Jiang, Y.; Zhao, L.; Liu, H.; Yang, L.; Luo, K. Molecular Cloning and Characterization of PtrLAR3, a Gene Encoding Leucoanthocyanidin Reductase from Populus trichocarpa, and Its Constitutive Expression Enhances Fungal Resistance in Transgenic Plants. J. Exp. Bot. 2012, 63, 2513–2524. [Google Scholar] [CrossRef] [PubMed]
- Sels, J.; Mathys, J.; De Coninck, B.M.A.; Cammue, B.P.A.; De Bolle, M.F.C. Plant Pathogenesis-Related (PR) Proteins: A Focus on PR Peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Wang, P.; Li, D.; Yin, T.; Zhuge, Q. Characterization, Expression Profiling, and Functional Analysis of PtDef, a Defensin-Encoding Gene from Populus Trichocarpa. Front. Microbiol. 2020, 11, 106. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Liu, G.; Zhu, S.; Chen, Y.; Yu, C.; Zhong, F.; Zhang, J. Characteristics, Expression Profile, and Function of Non-Specific Lipid Transfer Proteins of Populus Trichocarpa. Int. J. Biol. Macromol. 2022, 202, 468–481. [Google Scholar] [CrossRef]
- Jia, Z.; Gou, J.; Sun, Y.; Yuan, L.; Tang, Q.; Yang, X.; Pei, Y.; Luo, K. Enhanced Resistance to Fungal Pathogens in Transgenic Populus Tomentosa Carr. by Overexpression of an nsLTP-like Antimicrobial Protein Gene from Motherwort (Leonurus japonicus). Tree Physiol. 2010, 30, 1599–1605. [Google Scholar] [CrossRef]
- Rinaldi, C.; Kohler, A.; Frey, P.; Duchaussoy, F.; Ningre, N.; Couloux, A.; Wincker, P.; Le Thiec, D.; Fluch, S.; Martin, F.; et al. Transcript Profiling of Poplar Leaves upon Infection with Compatible and Incompatible Strains of the Foliar Rust Melampsora Larici-populina. Plant Physiol. 2007, 144, 347–366. [Google Scholar] [CrossRef]
- Petre, B.; Hecker, A.; Germain, H.; Tsan, P.; Sklenar, J.; Pelletier, G.; Séguin, A.; Duplessis, S.; Rouhier, N. The Poplar Rust-Induced Secreted Protein (RISP) Inhibits the Growth of the Leaf Rust Pathogen Melampsora Larici-populina and Triggers Cell Culture Alkalinisation. Front. Plant Sci. 2016, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Yevtushenko, D.P.; Misra, S. Enhancing Disease Resistance in Poplar through Modification of Its Natural Defense Pathway. Plant Mol. Biol. 2019, 100, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Song, J.; Huang, R.; Huang, M.; Xu, L. Cloning and Expression Analysis of Chitinase Genes from Populus Canadensis. Russ. J. Plant Physiol. 2013, 60, 396–403. [Google Scholar] [CrossRef]
- Jia, Z.; Sun, Y.; Yuan, L.; Tian, Q.; Luo, K. The Chitinase Gene (Bbchit1) from Beauveria Bassiana Enhances Resistance to Cytospora Chrysosperma in Populus tomentosa Carr. Biotechnol. Lett. 2010, 32, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, H.; Jia, Z.; Fang, Q.; Luo, K. Combined Expression of Antimicrobial Genes (Bbchit1 and LJAMP2) in Transgenic Poplar Enhances Resistance to Fungal Pathogens. Tree Physiol. 2012, 32, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhou, Y.; Movahedi, A.; Wei, H.; Zhuge, Q. Thaumatin-like Protein(Pe-TLP)Acts as a Positive Factor in Transgenic Poplars Enhanced Resistance to Spots Disease. Physiol. Mol. Plant Pathol. 2020, 112, 101512. [Google Scholar] [CrossRef]
- Diao, J.; Gu, W.; Jiang, Z.; Wang, J.; Zou, H.; Zong, C.; Ma, L. Comprehensive Analysis of Universal Stress Protein Family Genes and Their Expression in Fusarium oxysporum Response of Populus davidiana × P. alba Var. pyramidalis Louche Based on the Transcriptome. Int. J. Mol. Sci. 2023, 24, 5405. [Google Scholar] [CrossRef]
- Chi, Y.H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, S.D.; Bae, S.B.; Paeng, S.K.; Chae, H.B.; et al. The Physiological Functions of Universal Stress Proteins and Their Molecular Mechanism to Protect Plants from Environmental Stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef]
- Ji, S.D.; Wang, Z.Y.; Fan, H.J.; Zhang, R.S.; Yu, Z.Y.; Wang, J.J.; Liu, Z.H. Heterologous Expression of the Hsp24 from Trichoderma Asperellum Improves Antifungal Ability of Populus Transformant Pdpap-Hsp24 s to Cytospora Chrysosperma and Alternaria Alternate. J. Plant Res. 2016, 129, 921–933. [Google Scholar] [CrossRef]
- Cai, G.; Zhang, Y.; Huang, L.; Wang, N. Uncovering the Role of PdePrx12 Peroxidase in Enhancing Disease Resistance in Poplar Trees. J. Fungi 2023, 9, 410. [Google Scholar] [CrossRef]
- Liao, W.; Ji, L.; Wang, J.; Chen, Z.; Ye, M.; Ma, H.; An, X. Identification of Glutathione S-Transferase Genes Responding to Pathogen Infestation in Populus tomentosa. Funct. Integr. Genom. 2014, 14, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Duan, Y.; Yin, J.; Ye, S.; Zhu, J.; Zhang, F.; Lu, W.; Fan, D.; Luo, K. Genome-Wide Identification and Characterization of the Populus WRKY Transcription Factor Family and Analysis of Their Expression in Response to Biotic and Abiotic Stresses. J. Exp. Bot. 2014, 65, 6629–6644. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY Transcription Factors and Plant Defense Responses: Latest Discoveries and Future Prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, L.; Zhu, Y.; Li, Y.; Yan, H.; Xiang, Y. Comparative Genomic Analysis of the WRKY III Gene Family in Populus, Grape, Arabidopsis and Rice. Biol. Direct 2015, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Guo, L.; Ma, X.; Zhao, X.; Jiao, B.; Li, C.; Luo, K. The WRKY Transcription Factors PtrWRKY18 and PtrWRKY35 Promote Melampsora Resistance in Populus. Tree Physiol. 2017, 37, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Bai, X.-D.; Chen, K.; Gu, C.-R.; Yu, Q.-B.; Jiang, J.; Liu, G.-F. Role of PsnWRKY70 in Regulatory Network Response to Infection with Alternaria alternata (Fr.) Keissl in Populus. Int. J. Mol. Sci. 2022, 23, 7537. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Jiang, Y.; Guo, L.; Ling, Z.; Ye, S.; Duan, Y.; Li, C.; Luo, K. Isolation and Characterization of a Subgroup IIa WRKY Transcription Factor PtrWRKY40 from Populus trichocarpa. Tree Physiol. 2015, 35, 1129–1139. [Google Scholar] [CrossRef]
- Ye, S.; Jiang, Y.; Duan, Y.; Karim, A.; Fan, D.; Yang, L.; Zhao, X.; Yin, J.; Luo, K.; Li, C. Constitutive Expression of the Poplar WRKY Transcription Factor PtoWRKY60 Enhances Resistance to Dothiorella gregaria Sacc. in Transgenic Plants. Tree Physiol. 2014, 34, 1118–1129. [Google Scholar] [CrossRef]
- Duan, Y.; Jiang, Y.; Ye, S.; Karim, A.; Ling, Z.; He, Y.; Yang, S.; Luo, K. PtrWRKY73, a Salicylic Acid-Inducible Poplar WRKY Transcription Factor, Is Involved in Disease Resistance in Arabidopsis Thaliana. Plant Cell Rep. 2015, 34, 831–841. [Google Scholar] [CrossRef]
- Levée, V.; Major, I.; Levasseur, C.; Tremblay, L.; MacKay, J.; Séguin, A. Expression Profiling and Functional Analysis of Populus WRKY23 Reveals a Regulatory Role in Defense. New Phytol. 2009, 184, 48–70. [Google Scholar] [CrossRef]
- Qi, M.; Wu, R.; Song, Z.; Dong, B.; Chen, T.; Wang, M.; Cao, H.; Du, T.; Wang, S.; Li, N.; et al. Sorbitol Reduces Sensitivity to Alternaria by Promoting Ceramide Kinases (CERK) Expression through Transcription Factor Pswrky25 in Populus (Populus simonii Carr.). Genes 2022, 13, 405. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Lu, B.; Dong, Y.; Li, Y.; Yang, M. Cloning and Functional Identification of PeWRKY41 from Populus × Euramericana. Ind. Crops Prod. 2022, 175, 114279. [Google Scholar] [CrossRef]
- Shen, Z.; Yao, J.; Sun, J.; Chang, L.; Wang, S.; Ding, M.; Qian, Z.; Zhang, H.; Zhao, N.; Sa, G.; et al. Populus Euphratica HSF Binds the Promoter of WRKY1 to Enhance Salt Tolerance. Plant Sci. 2015, 235, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Salih, H.; Htet, N.N.W.; Azeem, F.; Zhan, R. Genomic Analysis of WD40 Protein Family in the Mango Reveals a TTG1 Protein Enhances Root Growth and Abiotic Tolerance in Arabidopsis. Sci. Rep. 2021, 11, 2266. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Ma, D.; Constabel, C.P. The MYB182 Protein Down-Regulates Proanthocyanidin and Anthocyanin Biosynthesis in Poplar by Repressing Both Structural and Regulatory Flavonoid Genes. Plant Physiol. 2015, 167, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Reichelt, M.; Yoshida, K.; Gershenzon, J.; Constabel, C.P. Two R2R3- MYB Proteins Are Broad Repressors of Flavonoid and Phenylpropanoid Metabolism in Poplar. Plant J. 2018, 96, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Li, C.; Ma, X.; Luo, K. PtrMYB57 Contributes to the Negative Regulation of Anthocyanin and Proanthocyanidin Biosynthesis in Poplar. Plant Cell Rep. 2017, 36, 1263–1276. [Google Scholar] [CrossRef]
- Mellway, R.D.; Tran, L.T.; Prouse, M.B.; Campbell, M.M.; Constabel, C.P. The Wound-, Pathogen-, and Ultraviolet B-Responsive MYB134 Gene Encodes an R2R3 MYB Transcription Factor That Regulates Proanthocyanidin Synthesis in Poplar. Plant Physiol. 2009, 150, 924–941. [Google Scholar] [CrossRef]
- Bai, Q.; Duan, B.; Ma, J.; Fen, Y.; Sun, S.; Long, Q.; Lv, J.; Wan, D. Coexpression of PalbHLH1 and PalMYB90 Genes from Populus Alba Enhances Pathogen Resistance in Poplar by Increasing the Flavonoid Content. Front. Plant Sci. 2020, 10, 1772. [Google Scholar] [CrossRef]
- Cho, J.-S.; Nguyen, V.P.; Jeon, H.-W.; Kim, M.-H.; Eom, S.H.; Lim, Y.J.; Kim, W.-C.; Park, E.-J.; Choi, Y.-I.; Ko, J.-H. Overexpression of PtrMYB119, a R2R3-MYB Transcription Factor from Populus trichocarpa, Promotes Anthocyanin Production in Hybrid Poplar. Tree Physiol. 2016, 36, 1162–1176. [Google Scholar] [CrossRef]
- Kim, M.-H.; Cho, J.-S.; Bae, E.-K.; Choi, Y.-I.; Eom, S.H.; Lim, Y.J.; Lee, H.; Park, E.-J.; Ko, J.-H. PtrMYB120 Functions as a Positive Regulator of Both Anthocyanin and Lignin Biosynthetic Pathway in a Hybrid Poplar. Tree Physiol. 2021, 41, 2409–2423. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, W.; Ran, L.; Dou, L.; Yao, S.; Hu, J.; Fan, D.; Li, C.; Luo, K. R2R3- MYB Transcription Factor MYB 6 Promotes Anthocyanin and Proanthocyanidin Biosynthesis but Inhibits Secondary Cell Wall Formation in Populus tomentosa. Plant J. 2019, 99, 733–751. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Yu, C.; Wang, C.; Jin, Y.; Zhang, H. MYB Transcription Factor PdMYB118 Directly Interacts with bHLH Transcription Factor PdTT8 to Regulate Wound-Induced Anthocyanin Biosynthesis in Poplar. BMC Plant Biol. 2020, 20, 173. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Tang, H.; Reichelt, M.; Piirtola, E.-M.; Salminen, J.-P.; Gershenzon, J.; Constabel, C.P. Poplar MYB117 Promotes Anthocyanin Synthesis and Enhances Flavonoid B-Ring Hydroxylation by up-Regulating the Flavonoid 3′,5′-Hydroxylase Gene. J. Exp. Bot. 2021, 72, 3864–3880. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Kong, L.; Yang, X.; Jiao, B.; Hu, J.; Zhang, Z.; Xu, C.; Luo, K. PtoMYB142, a Poplar R2R3-MYB Transcription Factor, Contributes to Drought Tolerance by Regulating Wax Biosynthesis. Tree Physiol. 2022, 42, 2133–2147. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Shen, K.; Newcombe, G.; Fan, J.; Chen, Q. Leaf Cuticle Can Contribute to Non-Host Resistance to Poplar Leaf Rust. Forests 2019, 10, 870. [Google Scholar] [CrossRef]
- Wei, M.; Xu, X.; Li, C. Identification and Expression of CAMTA Genes in Populus trichocarpa under Biotic and Abiotic Stress. Sci. Rep. 2017, 7, 17910. [Google Scholar] [CrossRef]
- Meraj, T.A.; Fu, J.; Raza, M.A.; Zhu, C.; Shen, Q.; Xu, D.; Wang, Q. Transcriptional Factors Regulate Plant Stress Responses Through Mediating Secondary Metabolism. Genes 2020, 11, 346. [Google Scholar] [CrossRef]
- Chen, S.; Wu, J.; Zhang, Y.; Zhao, Y.; Xu, W.; Li, Y.; Xie, J. Genome-Wide Analysis of Coding and Non-Coding RNA Reveals a Conserved miR164–NAC–mRNA Regulatory Pathway for Disease Defense in Populus. Front. Genet. 2021, 12, 668940. [Google Scholar] [CrossRef]
- Pan, J.; Wang, H.; Cheng, Q. Cloning and Functional Analysis of PtATAF11 Transcription Factor Gene of Populus trichocarpa. Tree Genet. Mol. Breed. 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Z.; Li, D.; Wang, F.; Zhang, R.; Wang, J. Molecular Characterization of the ERF Family in Susceptible Poplar Infected by Virulent Melampsora Larici-populina. Physiol. Mol. Plant Pathol. 2019, 108, 101437. [Google Scholar] [CrossRef]
- Diao, J.; Li, M.; Zhang, P.; Zong, C.; Ma, W.; Ma, L. Overexpression of the PdpapERF109 Gene Enhances Resistance of Populus davidiana × P. alba Var. Pyramidalis to Fusarium Oxysporum Infection. J. For. Res. 2022, 33, 1925–1937. [Google Scholar] [CrossRef]
- Ward, D. Agrobacterium VirE2 Gets the VIP1 Treatment in Plant Nuclear Import. Trends Plant Sci. 2002, 7, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lang, J.; Genot, B.; Bigeard, J.; Colcombet, J. MPK3 and MPK6 Control Salicylic Acid Signaling by Up-Regulating NLR Receptors during Pattern- and Effector-Triggered Immunity. J. Exp. Bot. 2022, 73, 2190–2205. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, D.; Tian, C. Mitogen-Activated Protein Kinase Cascade CgSte50-Ste11-Ste7-Mk1 Regulates Infection-Related Morphogenesis in the Poplar Anthracnose Fungus Colletotrichum gloeosporioides. Microbiol. Res. 2021, 248, 126748. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, H.-G.; Liu, M.; Wang, H.-L.; Yang, Q.; Yan, D.-H.; Zhang, Y.; Li, Z.; Feng, C.-H.; Niu, M.; et al. PeTGA1 Enhances Disease Resistance against Colletotrichum Gloeosporioides through Directly Regulating PeSARD1 in Poplar. Int. J. Biol. Macromol. 2022, 214, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lie, J.; Cui, K.; Yang, J.; Baloch, A.M.; Liu, S.; Zhang, Y.; Baloch, A.W.; Zhang, R. Expression Profile of PdpapHB12 Gene in Response to Stress for Populus Davidana × P. Alba Var. Pyramidlis. Pak. J. Bot. 2024, 56, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Q.; Wang, H.; Zhang, H.; Xu, X.; Li, C.; Yang, C. Comprehensive Analysis of Trihelix Genes and Their Expression under Biotic and Abiotic Stresses in Populus trichocarpa. Sci. Rep. 2016, 6, 36274. [Google Scholar] [CrossRef]
- Xiao, D.; Zhou, K.; Yang, X.; Yang, Y.; Ma, Y.; Wang, Y. Crosstalk of DNA Methylation Triggered by Pathogen in Poplars with Different Resistances. Front. Microbiol. 2021, 12, 750089. [Google Scholar] [CrossRef]
- Kong, X.; Yang, M.; Le, B.H.; He, W.; Hou, Y. The Master Role of siRNAs in Plant Immunity. Mol. Plant Pathol. 2022, 23, 1565–1574. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, S.; Zhou, Y.; Bai, J.; Huang, G.; Liu, X.; Zhang, Y.; Tang, D.; Lu, D. Transcriptional Regulation of the Immune Receptor FLS2 Controls the Ontogeny of Plant Innate Immunity. Plant Cell 2018, 30, 2779–2794. [Google Scholar] [CrossRef] [PubMed]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D.G. A Plant miRNA Contributes to Antibacterial Resistance by Repressing Auxin Signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.-L.; Zhao, J.-H.; Wang, S.; Jin, Y.; Chen, Z.-Q.; Fang, Y.-Y.; Hua, C.-L.; Ding, S.-W.; Guo, H.-S. Cotton Plants Export microRNAs to Inhibit Virulence Gene Expression in a Fungal Pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Y.; Wafula, E.K.; Honaas, L.A.; Ralph, P.E.; Jones, S.; Clarke, C.R.; Liu, S.; Su, C.; Zhang, H.; et al. Horizontal Gene Transfer Is More Frequent with Increased Heterotrophy and Contributes to Parasite Adaptation. Proc. Natl. Acad. Sci. USA 2016, 113, E7010–E7019. [Google Scholar] [CrossRef] [PubMed]
- Boccara, M.; Sarazin, A.; Thiébeauld, O.; Jay, F.; Voinnet, O.; Navarro, L.; Colot, V. The Arabidopsis miR472-RDR6 Silencing Pathway Modulates PAMP- and Effector-Triggered Immunity through the Post-Transcriptional Control of Disease Resistance Genes. PLoS Pathog. 2014, 10, e1003883. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Liang, C.; Wang, S.; Hou, Y.; Gao, L.; Liu, L.; He, W.; Ma, W.; Mo, B.; Chen, X. The Disease Resistance Protein SNC1 Represses the Biogenesis of microRNAs and Phased siRNAs. Nat. Commun. 2018, 9, 5080. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ding, Z.; Wu, K.; Yang, L.; Li, Y.; Yang, Z.; Shi, S.; Liu, X.; Zhao, S.; Yang, Z.; et al. Suppression of Jasmonic Acid-Mediated Defense by Viral-Inducible MicroRNA319 Facilitates Virus Infection in Rice. Mol. Plant 2016, 9, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, Z.; Wang, Y.; Zheng, L.; Ye, R.; Ji, Y.; Zhao, S.; Ji, S.; Liu, R.; Xu, L.; et al. Viral-Inducible Argonaute18 Confers Broad-Spectrum Virus Resistance in Rice by Sequestering a Host microRNA. eLife 2015, 4, e05733. [Google Scholar] [CrossRef]
- Zhao, J.-P.; Jiang, X.-L.; Zhang, B.-Y.; Su, X.-H. Involvement of microRNA-Mediated Gene Expression Regulation in the Pathological Development of Stem Canker Disease in Populus trichocarpa. PLoS ONE 2012, 7, e44968. [Google Scholar] [CrossRef]
- Gou, J.-Y.; Felippes, F.F.; Liu, C.-J.; Weigel, D.; Wang, J.-W. Negative Regulation of Anthocyanin Biosynthesis in Arabidopsis by a miR156-Targeted SPL Transcription Factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef]
- Chen, L.; Ren, Y.; Zhang, Y.; Xu, J.; Zhang, Z.; Wang, Y. Genome-Wide Profiling of Novel and Conserved Populus microRNAs Involved in Pathogen Stress Response by Deep Sequencing. Planta 2012, 235, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Gupta, O.P.; Sharma, P.; Gupta, R.K.; Sharma, I. Current Status on Role of miRNAs during Plant–Fungus Interaction. Physiol. Mol. Plant Pathol. 2014, 85, 1–7. [Google Scholar] [CrossRef]
- Liu, S.; Li, X.; Zhao, Y.; Xie, J. Transposon-Associated Small RNAs Involved in Plant Defense in Poplar. Res. Sq. 2021. in review. [Google Scholar] [CrossRef]
- Su, Y.; Li, H.-G.; Wang, Y.; Li, S.; Wang, H.-L.; Yu, L.; He, F.; Yang, Y.; Feng, C.-H.; Shuai, P.; et al. Poplar miR472a Targeting NBS-LRRs Is Involved in Effective Defence against the Necrotrophic Fungus Cytospora Chrysosperma. J. Exp. Bot. 2018, 69, 5519–5530. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, M.; Zhou, X.; Cao, Z. Identification of Novel miRNAs and Their Target Genes from Populus Szechuanica Infected with Melampsora Larici-populina. Mol. Biol. Rep. 2019, 46, 3083–3092. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cao, Z. Genome-Wide Expression Profiling of microRNAs in Poplar upon Infection with the Foliar Rust Fungus Melampsora Larici-populina. BMC Genom. 2015, 16, 696. [Google Scholar] [CrossRef]
- Li, D.; Wang, F.; Wang, C.; Zou, L.; Wang, Z.; Chen, Q.; Niu, C.; Zhang, R.; Ling, Y.; Wang, B. MicroRNA-Mediated Susceptible Poplar Gene Expression Regulation Associated with the Infection of Virulent Melampsora Larici-populina. BMC Genom. 2016, 17, 59. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Yin, Y.; Wang, J.; Wang, Y. Identification and Validation of miRNA Reference Genes in Poplar under Pathogen Stress. Mol. Biol. Rep. 2021, 48, 3357–3366. [Google Scholar] [CrossRef]
- Xu, Q.; Chang, F.; Huo, X.; Zhou, K.; Wang, Y. Prediction and Analysis of miRNA Targets in Poplar in Response to the Infection of Lonsdalea quercina Subsp populi. Tree Genet. Mol. Breed. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Tian, W.; Ge, Y.; Liu, X.; Dou, G.; Ma, Y. Identification and Characterization of Populus microRNAs in Response to Plant Growth-Promoting Endophytic Streptomyces Sp. SSD49. World J. Microbiol. Biotechnol. 2019, 35, 97. [Google Scholar] [CrossRef]
- Okabe, S. Do microRNA156a/168a Regulate Mycorrhizal Formation in Populus tomentosa? In Proceedings of the Japanese Forest Society Congress 134th Annual JFS Meeting, Tokyo, Japan, 25–27 March 2023. [Google Scholar]
- Wang, N.; Cao, P.; Xia, W.; Fang, L.; Yu, H. Identification and Characterization of Long Non-Coding RNAs in Response to Early Infection by Melampsora Larici-Populina Using Genome-Wide High-Throughput RNA Sequencing. Tree Genet. Genomes 2017, 13, 34. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, Y.; Ren, Y.; Wang, S.; Yang, M. Conjoint Analysis of Genome-Wide lncRNA and mRNA Expression during the Salicylic Acid Response in Populus × Euramericana. Plants 2023, 12, 1377. [Google Scholar] [CrossRef] [PubMed]
- Filiz, E.; Vatansever, R. Genome-Wide Identification of Mildew Resistance Locus O (MLO) Genes in Tree Model Poplar (Populus Trichocarpa): Powdery Mildew Management in Woody Plants. Eur. J. Plant Pathol. 2018, 152, 95–109. [Google Scholar] [CrossRef]
- Kohler, A.; Rinaldi, C.; Duplessis, S.; Baucher, M.; Geelen, D.; Duchaussoy, F.; Meyers, B.C.; Boerjan, W.; Martin, F. Genome-Wide Identification of NBS Resistance Genes in Populus trichocarpa. Plant Mol. Biol. 2008, 66, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Bresson, A.; Jorge, V.; Dowkiw, A.; Guerin, V.; Bourgait, I.; Tuskan, G.A.; Schmutz, J.; Chalhoub, B.; Bastien, C.; Faivre Rampant, P. Qualitative and Quantitative Resistances to Leaf Rust Finely Mapped within Two Nucleotide-binding Site Leucine-rich Repeat (NBS-LRR)-rich Genomic Regions of Chromosome 19 in Poplar. New Phytol. 2011, 192, 151–163. [Google Scholar] [CrossRef]
- Simon, S.J.; Tschaplinski, T.J.; LeBoldus, J.M.; Keefover-Ring, K.; Azeem, M.; Chen, J.; Macaya-Sanz, D.; MacDonald, W.L.; Muchero, W.; DiFazio, S.P. Host Plant Genetic Control of Associated Fungal and Insect Species in a Populus Hybrid Cross. Ecol. Evol. 2020, 10, 5119–5134. [Google Scholar] [CrossRef] [PubMed]
- La Mantia, J.; Klápště, J.; El-Kassaby, Y.A.; Azam, S.; Guy, R.D.; Douglas, C.J.; Mansfield, S.D.; Hamelin, R. Association Analysis Identifies Melampsora ×columbiana Poplar Leaf Rust Resistance SNPs. PLoS ONE 2013, 8, e78423. [Google Scholar] [CrossRef]
- Wei, S.; Wu, H.; Li, X.; Chen, Y.; Yang, Y.; Dai, M.; Yin, T. Identification of Genes Underlying the Resistance to Melampsora larici-Populina in an R. Gene Supercluster of the Populus Deltoides Genome. Plant Dis. 2020, 104, 1133–1143. [Google Scholar] [CrossRef]
- Muchero, W.; Sondreli, K.L.; Chen, J.-G.; Urbanowicz, B.R.; Zhang, J.; Singan, V.; Yang, Y.; Brueggeman, R.S.; Franco-Coronado, J.; Abraham, N.; et al. Association Mapping, Transcriptomics, and Transient Expression Identify Candidate Genes Mediating Plant–Pathogen Interactions in a Tree. Proc. Natl. Acad. Sci. USA 2018, 115, 11573–11578. [Google Scholar] [CrossRef]
- Louet, C.; Saubin, M.; Andrieux, A.; Persoons, A.; Gorse, M.; Pétrowski, J.; Fabre, B.; De Mita, S.; Duplessis, S.; Frey, P.; et al. A Point Mutation and Large Deletion at the Candidate Avirulence Locus AvrMlp7 in the Poplar Rust Fungus Correlate with Poplar RMlp7 Resistance Breakdown. Mol. Ecol. 2023, 32, 2472–2483. [Google Scholar] [CrossRef]
- Ren, F.; Yan, D.-H.; Wu, G.; Sun, X.; Song, X.; Li, R. Distinctive Gene Expression Profiles and Effectors Consistent With Host Specificity in Two Formae Speciales of Marssonina Brunnea. Front. Microbiol. 2020, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Lenz, R.R.; Shrestha, H.K.; Carrell, A.A.; Labbé, J.; Hettich, R.L.; Abraham, P.E.; LeBoldus, J.M. Proteomics Reveals Pathways Linked to Septoria Canker Resistance and Susceptibility in Populus trichocarpa. Front. Anal. Sci. 2022, 2, 1020111. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F. Integrating Transcriptome and Coexpression Network Analyses to Characterize Salicylic Acid- and Jasmonic Acid-Related Genes in Tolerant Poplars Infected with Rust. Int. J. Mol. Sci. 2021, 22, 5001. [Google Scholar] [CrossRef] [PubMed]
- La Mantia, J.; Unda, F.; Douglas, C.J.; Mansfield, S.D.; Hamelin, R. Overexpression of AtGolS3 and CsRFS in Poplar Enhances ROS Tolerance and Represses Defense Response to Leaf Rust Disease. Tree Physiol. 2018, 38, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ma, H.; Yue, Y.; Zhou, T.; Zhu, Z.; Wang, C. Integrated Transcriptomic and Transgenic Analyses Reveal Potential Mechanisms of Poplar Resistance to Alternaria Alternata Infection. BMC Plant Biol. 2022, 22, 413. [Google Scholar] [CrossRef] [PubMed]
- Lenz, R.R.; Louie, K.B.; Søndreli, K.L.; Galanie, S.S.; Chen, J.-G.; Muchero, W.; Bowen, B.P.; Northen, T.R.; LeBoldus, J.M. Metabolomic Patterns of Septoria Canker Resistant and Susceptible Populus trichocarpa Genotypes 24 Hours Postinoculation. Phytopathology 2021, 111, 2052–2066. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.; Yu, R.; Zhang, L.; Yang, Y.; Xiao, D.; Li, A.; Wang, Y. Integrated Transcriptomic and Metabolomic Profiles Reveal Adaptive Responses of Three Poplar Varieties against the Bacterial Pathogen Lonsdalea populi. Plant Cell Environ. 2023, 46, 306–321. [Google Scholar] [CrossRef]
- Prunier, J.; Giguère, I.; Ryan, N.; Guy, R.; Soolanayakanahally, R.; Isabel, N.; MacKay, J.; Porth, I. Gene Copy Number Variations Involved in Balsam Poplar (Populus balsamifera L.) Adaptive Variations. Mol. Ecol. 2019, 28, 1476–1490. [Google Scholar] [CrossRef]
- Hacquard, S.; Schadt, C.W. Towards a Holistic Understanding of the Beneficial Interactions across the Populus Microbiome. New Phytol. 2015, 205, 1424–1430. [Google Scholar] [CrossRef]
- Cregger, M.A.; Carper, D.L.; Christel, S.; Doktycz, M.J.; Labbé, J.; Michener, J.K.; Dove, N.C.; Johnston, E.R.; Moore, J.A.M.; Vélez, J.M.; et al. Plant–Microbe Interactions: From Genes to Ecosystems Using Populus as a Model System. Phytobiomes J. 2021, 5, 29–38. [Google Scholar] [CrossRef]
- Gladysh, N.S.; Bogdanova, A.S.; Kovalev, M.A.; Krasnov, G.S.; Volodin, V.V.; Shuvalova, A.I.; Ivanov, N.V.; Popchenko, M.I.; Samoilova, A.D.; Polyakova, A.N.; et al. Culturable Bacterial Endophytes of Wild White Poplar (Populus alba L.) Roots: A First Insight into Their Plant Growth-Stimulating and Bioaugmentation Potential. Biology 2023, 12, 1519. [Google Scholar] [CrossRef] [PubMed]
- Dove, N.C.; Veach, A.M.; Muchero, W.; Wahl, T.; Stegen, J.C.; Schadt, C.W.; Cregger, M.A. Assembly of the Populus Microbiome Is Temporally Dynamic and Determined by Selective and Stochastic Factors. mSphere 2021, 6, e01316-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Brettell, L.E.; Qiu, Z.; Singh, B.K. Microbiome-Mediated Stress Resistance in Plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Syed Ab Rahman, S.F.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging Microbial Biocontrol Strategies for Plant Pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Sachin, N.; Tsang, A.; Shaanker, R.U.; Dayanandan, S. Genome Sequence Resource of Bacillus velezensis EB14, a Native Endophytic Bacterial Strain with Biocontrol Potential Against the Poplar Stem Canker Causative Pathogen, Sphaerulina musiva. Phytopathology 2021, 111, 890–892. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Palys, S.; Di Falco, M.; Tsang, A.; Périnet, P.; Ramanan, U.S.; Dayanandan, S. Isolation and Characterization of Bacillus Velezensis EB14, an Endophytic Bacterial Strain Antagonistic to Poplar Stem Canker Pathogen Sphaerulina musiva and Its Interactions with the Endophytic Fungal Microbiome in Poplar. PhytoFrontiersTM 2021, 1, 229–238. [Google Scholar] [CrossRef]
- Martínez-Arias, C.; Macaya-Sanz, D.; Witzell, J.; Martín, J.A. Enhancement of Populus Alba Tolerance to Venturia Tremulae upon Inoculation with Endophytes Showing in Vitro Biocontrol Potential. Eur. J. Plant Pathol. 2019, 153, 1031–1042. [Google Scholar] [CrossRef]
- Tyagi, K.; Kumar, P.; Pandey, A.; Ginwal, H.S.; Barthwal, S.; Nautiyal, R.; Meena, R.K. First Record of Cladosporium Oxysporum as a Potential Novel Fungal Hyperparasite of Melampsora Medusae f. Sp. Deltoidae and Screening of Populus Deltoides Clones against Leaf Rust. 3 Biotech 2023, 13, 213. [Google Scholar] [CrossRef]
- Zhang, P.; Hao, H.; Wang, L.; Liu, Z.; Ma, L. Endophytes Bacillus Amyloliquefaciens AW3 (CGMCC1.16683) Improves the Growth of Populus Davidiana × Populus Bolleana (PdPap) and Induces Its Resistance to Wilt Disease by Fusarium Oxysporum Fox68 (CFCC86068). Eur. J. Plant Pathol. 2022, 162, 1–17. [Google Scholar] [CrossRef]
- Dong, F.; Wang, Y.; Tang, M. Study on the Molecular Mechanism of Laccaria Bicolor Helping Populus trichocarpa to Resist the Infection of Botryosphaeria Dothidea. J. Appl. Microbiol. 2022, 132, 2220–2233. [Google Scholar] [CrossRef]
- Raghavendra, A.K.H.; Newcombe, G. The Contribution of Foliar Endophytes to Quantitative Resistance to Melampsora Rust. New Phytol. 2013, 197, 909–918. [Google Scholar] [CrossRef]
- Fan, J.; Wang, S.; Ding, C.; Ma, C.; Chen, X.; Wang, J.; Yang, M.; Su, X. Correlation Analysis of the Bacterial Community and Wood Properties of Populus × Euramericana Cv. “74/76” Wet Heartwood. Front. Microbiol. 2022, 13, 868078. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Ma, Y.; Li, X.; Wu, J.; Martin, F.; Zhang, D. Multifeature Analysis of Age-related Microbiome Structures Reveals Defense Mechanisms of Populus Tomentosa Trees. New Phytol. 2023, 238, 1636–1650. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Meng, F.; Ge, W.; Ren, Y.; Bao, H.; Tian, C. Effects of Colletotrichum gloeosporioides and Poplar Secondary Metabolites on the Composition of Poplar Phyllosphere Microbial Communities. Microbiol. Spectr. 2023, 11, e04603-22. [Google Scholar] [CrossRef] [PubMed]
- Szuba, A. Ectomycorrhiza of populus. For. Ecol. Manag. 2015, 347, 156–169. [Google Scholar] [CrossRef]
- Timm, C.M.; Carter, K.R.; Carrell, A.A.; Jun, S.R.; Jawdy, S.S.; Vélez, J.M.; Gunter, L.E.; Yang, Z.; Nookaew, I.; Engle, N.L.; et al. Abiotic Stresses Shift Belowground Populus-Associated Bacteria Toward a Core Stress Microbiome. mSystems 2018, 3, e00070-17. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Peay, K.G.; Newcombe, G. Common Foliar Fungi of Populus Trichocarpa Modify Melampsora Rust Disease Severity. New Phytol. 2016, 209, 1681–1692. [Google Scholar] [CrossRef]
- Ridout, M.; Newcombe, G. The Frequency of Modification of Dothistroma Pine Needle Blight Severity by Fungi within the Native Range. For. Ecol. Manag. 2015, 337, 153–160. [Google Scholar] [CrossRef]
- Kwaśna, H.; Szewczyk, W.; Baranowska, M.; Gallas, E.; Wiśniewska, M.; Behnke-Borowczyk, J. Mycobiota Associated with the Vascular Wilt of Poplar. Plants 2021, 10, 892. [Google Scholar] [CrossRef]
- Eberl, F.; Fernandez De Bobadilla, M.; Reichelt, M.; Hammerbacher, A.; Gershenzon, J.; Unsicker, S.B. Herbivory Meets Fungivory: Insect Herbivores Feed on Plant Pathogenic Fungi for Their Own Benefit. Ecol. Lett. 2020, 23, 1073–1084. [Google Scholar] [CrossRef]
- Busby, P.E.; Lamit, L.J.; Keith, A.R.; Newcombe, G.; Gehring, C.A.; Whitham, T.G.; Dirzo, R. Genetics-based Interactions among Plants, Pathogens, and Herbivores Define Arthropod Community Structure. Ecology 2015, 96, 1974–1984. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.B.; Pawlus, A.D.; Brinkman, D.; Gardner, G.; Hegeman, A.D.; Spivak, M.; Cohen, J.D. 3-Acyl Dihydroflavonols from Poplar Resins Collected by Honey Bees Are Active against the Bee Pathogens Paenibacillus Larvae and Ascosphaera Apis. Phytochemistry 2017, 138, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Chen, J.; Zhou, S.; Yu, W.; Chen, G.; Chen, L.; Wang, X.; Shi, H.; Han, S.; Zhang, F. Testing the Elemental Defense Hypothesis with a Woody Plant Species: Cadmium Accumulation Protects Populus Yunnanensis from Leaf Herbivory and Pathogen Infection. Chemosphere 2020, 247, 125851. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zou, L.; Zhi, J.; Niu, Y.; Yan, S.; Yan, R.; Lin, T. Increased Soil Cadmium Concentrations Enhanced Poplar Defence against a Leaf Pathogenic Fungus Infection. J. Phytopathol. 2023, 171, 401–408. [Google Scholar] [CrossRef]
- Chen, J.; Qin, S.; Tang, J.; Chen, G.; Xie, J.; Chen, L.; Han, S.; Wang, X.; Zhu, T.; Liu, Y.; et al. Exogenous Nitrogen Enhances Poplar Resistance to Leaf Herbivory and Pathogen Infection after Exposure to Soil Cadmium Stress. Ecotoxicol. Environ. Saf. 2021, 208, 111688. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lu, Q.; Zheng, Z.; Li, S.; Li, S.; Liu, Y.; Zhu, T.; Chen, L.; Yang, C.; Han, S. Soil Cadmium Stress Affects the Phyllosphere Microbiome and Associated Pathogen Resistance Differently in Male and Female Poplars. J. Exp. Bot. 2023, 74, 2188–2202. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Tang, J.; He, F.; Chen, G.; Shi, Y.; Wang, X.; Han, S.; Li, S.; Zhu, T.; Chen, L. Sexual Differences in Above- and Belowground Herbivore Resistance between Male and Female Poplars as Affected by Soil Cadmium Stress. Sci. Total Environ. 2022, 803, 150081. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, A.; Yan, S. Effects of Zn Exposure on Populus Simonii Seedling Growth and Its Resistance to Leaf Rust. Forests 2023, 14, 783. [Google Scholar] [CrossRef]
- Müller, N.A.; Kersten, B.; Leite Montalvão, A.P.; Mähler, N.; Bernhardsson, C.; Bräutigam, K.; Carracedo Lorenzo, Z.; Hoenicka, H.; Kumar, V.; Mader, M.; et al. A Single Gene Underlies the Dynamic Evolution of Poplar Sex Determination. Nat. Plants 2020, 6, 630–637. [Google Scholar] [CrossRef]
- Cronk, Q.; Müller, N.A. Default Sex and Single Gene Sex Determination in Dioecious Plants. Front. Plant Sci. 2020, 11, 1162. [Google Scholar] [CrossRef]
- Melnikova, N.V.; Borkhert, E.V.; Snezhkina, A.V.; Kudryavtseva, A.V.; Dmitriev, A.A. Sex-Specific Response to Stress in Populus. Front. Plant Sci. 2017, 8, 1827. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Li, Z.; Wu, F.; Zhen, L. Sex-Specific Photosynthetic Capacity and Na+ Homeostasis in Populus Euphratica Exposed to NaCl Stress and AMF Inoculation. Front. Plant Sci. 2022, 13, 1066954. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Lin, X.; Wang, Y.; Yang, Q.; Liu, M. Nitrogen Level Induces Sex-Specific Cadmium Phloem Remobilization and Cell Wall Segregation in Populus Cathayana. Sci. Total Environ. 2023, 890, 164184. [Google Scholar] [CrossRef] [PubMed]
- Randriamanana, T.R.; Nybakken, L.; Lavola, A.; Aphalo, P.J.; Nissinen, K.; Julkunen-Tiitto, R. Sex-Related Differences in Growth and Carbon Allocation to Defence in Populus Tremula as Explained by Current Plant Defence Theories. Tree Physiol. 2014, 34, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, X.; Zhao, Y.; Korpelainen, H.; Li, C. Sex-Specific Nitrogen Allocation Tradeoffs in the Leaves of Populus Cathayana Cuttings under Salt and Drought Stress. Plant Physiol. Biochem. 2022, 172, 101–110. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, Y.; Liu, X.; Korpelainen, H.; Li, C. Ammonium and Nitrate Affect Sexually Different Responses to Salt Stress in Populus Cathayana. Physiol. Plant. 2022, 174, e13626. [Google Scholar] [CrossRef] [PubMed]
- Kasper, K.; Abreu, I.N.; Feussner, K.; Zienkiewicz, K.; Herrfurth, C.; Ischebeck, T.; Janz, D.; Majcherczyk, A.; Schmitt, K.; Valerius, O.; et al. Multi-omics Analysis of Xylem Sap Uncovers Dynamic Modulation of Poplar Defenses by Ammonium and Nitrate. Plant J. 2022, 111, 282–303. [Google Scholar] [CrossRef]
- He, F.; Wu, Z.; Zhao, Z.; Chen, G.; Wang, X.; Cui, X.; Zhu, T.; Chen, L.; Yang, P.; Bi, L.; et al. Drought Stress Drives Sex-Specific Differences in Plant Resistance against Herbivores between Male and Female Poplars through Changes in Transcriptional and Metabolic Profiles. Sci. Total Environ. 2022, 845, 157171. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Liu, J.; Korpelainen, H.; Li, C. Plant Sex Affects Plant-Microbiome Assemblies of Dioecious Populus Cathayana Trees under Different Soil Nitrogen Conditions. Microbiome 2022, 10, 191. [Google Scholar] [CrossRef]
- Liu, L.; Lu, L.; Li, H.; Meng, Z.; Dong, T.; Peng, C.; Xu, X. Divergence of Phyllosphere Microbial Communities Between Females and Males of the Dioecious Populus Cathayana. Mol. Plant-Microbe Interact. 2021, 34, 351–361. [Google Scholar] [CrossRef]
- Lin, T.; Tang, J.; Li, S.; Li, S.; Han, S.; Liu, Y.; Yang, C.; Chen, G.; Chen, L.; Zhu, T. Drought Stress-mediated Differences in Phyllosphere Microbiome and Associated Pathogen Resistance between Male and Female Poplars. Plant J. 2023, 115, 1100–1113. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; He, Y.; Korpelainen, H.; Niinemets, Ü.; Li, C. Allelochemicals and Soil Microorganisms Jointly Mediate Sex-specific Belowground Interactions in Dioecious Populus Cathayana. New Phytol. 2023, 240, 1519–1533. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Wang, X.; Li, D.; Zhuge, Q. Overexpression of PtDefensin enhances resistance to Septotis populiperda in transgenic poplar. Plant Sci. 2020, 292, 110379. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Yuan, G.; Lu, H.; Liu, Y.; Zhang, J.; Tuskan, G.A.; Muchero, W.; Chen, J.-G.; Yang, X. CRISPR/Cas9-Based Gene Activation and Base Editing in Populus. Hortic. Res. 2023, 10, uhad085. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Morrell-Falvey, J.L.; Labbé, J.L.; Muchero, W.; Kalluri, U.C.; Tuskan, G.A.; Chen, J.-G. Highly Efficient Isolation of Populus Mesophyll Protoplasts and Its Application in Transient Expression Assays. PLoS ONE 2012, 7, e44908. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Kuroda, H.; Hayashi, T.; Sakai, F. Efficient Plantlet Regeneration from Protoplasts Isolated from Suspension Cultures of Poplar (Populus alba L.). Plant Cell Rep. 1998, 17, 201–205. [Google Scholar] [CrossRef]
- Rahman, S.U.; Khan, M.O.; Ullah, R.; Ahmad, F.; Raza, G. Agrobacterium-Mediated Transformation for the Development of Transgenic Crops; Present and Future Prospects. Mol. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; He, Y.; Li, S.; Yan, L.; Li, Y.; Zhu, Z.; Xia, L. Plant Base Editing and Prime Editing: The Current Status and Future Perspectives. J. Integr. Plant Biol. 2023, 65, 444–467. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in Agriculture and Plant Biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Illa-Berenguer, E.; LaFayette, P.R.; Parrott, W.A. Editing Efficiencies with Cas9 Orthologs, Cas12a Endonucleases, and Temperature in Rice. Front. Genome Ed. 2023, 5, 1074641. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, G.; Zhao, Y.; Zhang, R.; Tang, X.; Li, L.; Jia, X.; Guo, Y.; Wu, Y.; Han, Y.; et al. An Efficient CRISPR–Cas12a Promoter Editing System for Crop Improvement. Nat. Plants 2023, 9, 588–604. [Google Scholar] [CrossRef] [PubMed]
- Sulis, D.B.; Jiang, X.; Yang, C.; Marques, B.M.; Matthews, M.L.; Miller, Z.; Lan, K.; Cofre-Vega, C.; Liu, B.; Sun, R.; et al. Multiplex CRISPR Editing of Wood for Sustainable Fiber Production. Science 2023, 381, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Wienert, B.; Cromer, M.K. CRISPR Nuclease Off-Target Activity and Mitigation Strategies. Front. Genome Ed. 2022, 4, 1050507. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Geng, Y.; Yao, J.; Fu, C.; Lu, M.; Wang, C.; Du, J. Efficient Genome Editing in Populus Using CRISPR/Cas12a. Front. Plant Sci. 2020, 11, 593938. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, V.M.G.; Brambilla, V.; Rogowsky, P.; Marocco, A.; Lanubile, A. The Enhancement of Plant Disease Resistance Using CRISPR/Cas9 Technology. Front. Plant Sci. 2018, 9, 1245. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Das, P.; Panda, D.; Xie, K.; Baig, M.J.; Molla, K.A. A Detailed Landscape of CRISPR-Cas-Mediated Plant Disease and Pest Management. Plant Sci. 2022, 323, 111376. [Google Scholar] [CrossRef]
- Khan, Z.A.; Kumar, R.; Dasgupta, I. CRISPR/Cas-Mediated Resistance against Viruses in Plants. Int. J. Mol. Sci. 2022, 23, 2303. [Google Scholar] [CrossRef]
- Li, X.; Sandgrind, S.; Moss, O.; Guan, R.; Ivarson, E.; Wang, E.S.; Kanagarajan, S.; Zhu, L.-H. Efficient Protoplast Regeneration Protocol and CRISPR/Cas9-Mediated Editing of Glucosinolate Transporter (GTR) Genes in Rapeseed (Brassica napus L.). Front. Plant Sci. 2021, 12, 680859. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous Editing of Three Homoeoalleles in Hexaploid Bread Wheat Confers Heritable Resistance to Powdery Mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid Generation of a Transgene-Free Powdery Mildew Resistant Tomato by Genome Deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef] [PubMed]
- Brauer, E.K.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Schernthaner, J.; Subramaniam, R.; Ouellet, T. Genome Editing of a Deoxynivalenol-Induced Transcription Factor Confers Resistance to Fusarium Graminearum in Wheat. Mol. Plant-Microbe Interact. 2020, 33, 553–560. [Google Scholar] [CrossRef]
- Bernhardsson, C.; Ingvarsson, P.K. Molecular Population Genetics of Elicitor-Induced Resistance Genes in European Aspen (Populus tremula L., Salicaceae). PLoS ONE 2011, 6, e24867. [Google Scholar] [CrossRef] [PubMed]
- Pathi, K.M.; Rink, P.; Budhagatapalli, N.; Betz, R.; Saado, I.; Hiekel, S.; Becker, M.; Djamei, A.; Kumlehn, J. Engineering Smut Resistance in Maize by Site-Directed Mutagenesis of LIPOXYGENASE 3. Front. Plant Sci. 2020, 11, 543895. [Google Scholar] [CrossRef]
- Gautam, T.; Dutta, M.; Jaiswal, V.; Zinta, G.; Gahlaut, V.; Kumar, S. Emerging Roles of SWEET Sugar Transporters in Plant Development and Abiotic Stress Responses. Cells 2022, 11, 1303. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene Targeting by the TAL Effector PthXo2 Reveals Cryptic Resistance Gene for Bacterial Blight of Rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.-S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-Spectrum Resistance to Bacterial Blight in Rice Using Genome Editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Zhang, J.; Song, C.; Li, Y.; Li, J.; Lu, M. Expression and Localization of SWEETs in Populus and the Effect of SWEET7 Overexpression in Secondary Growth. Tree Physiol. 2021, 41, 882–899. [Google Scholar] [CrossRef]
- Jia, H.; Orbovic, V.; Jones, J.B.; Wang, N. Modification of the PthA4 Effector Binding Elements in Type I Cs LOB 1 Promoter Using Cas9/Sg RNA to Produce Transgenic Duncan Grapefruit Alleviating XccΔpthA4:dCs LOB 1.3 Infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering Canker-resistant Plants through CRISPR/Cas9-targeted Editing of the Susceptibility Gene Cs LOB 1 Promoter in Citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome Editing of the Disease Susceptibility Gene Cs LOB 1 in Citrus Confers Resistance to Citrus Canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, Y.S.; Regan, S.; Busov, V. Members of the LATERAL ORGAN BOUNDARIES DOMAIN Transcription Factor Family Are Involved in the Regulation of Secondary Growth in Populus. Plant Cell 2010, 22, 3662–3677. [Google Scholar] [CrossRef] [PubMed]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA Virus Interference via CRISPR/Cas13a System in Plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z.; Shi, M.; Holmes, E.C. Using Metagenomics to Characterize an Expanding Virosphere. Cell 2018, 172, 1168–1172. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of Broad Virus Resistance in Non-transgenic Cucumber Using CRISPR/Cas9 Technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef]
- Pyott, D.E.; Sheehan, E.; Molnar, A. Engineering of CRISPR/Cas9-mediated Potyvirus Resistance in Transgene-free Arabidopsis Plants. Mol. Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.; Chadha-Mohanty, P. Novel Alleles of Rice eIF4G Generated by CRISPR/Cas9-targeted Mutagenesis Confer Resistance to Rice Tungro spherical virus. Plant Biotechnol. J. 2018, 16, 1918–1927. [Google Scholar] [CrossRef]
- Patrick, R.M.; Browning, K.S. The eIF4F and eIFiso4F Complexes of Plants: An Evolutionary Perspective. Comp. Funct. Genom. 2012, 2012, 287814. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Doty, S.L.; Sher, A.W.; Fleck, N.D.; Khorasani, M.; Bumgarner, R.E.; Khan, Z.; Ko, A.W.K.; Kim, S.-H.; DeLuca, T.H. Variable Nitrogen Fixation in Wild Populus. PLoS ONE 2016, 11, e0155979. [Google Scholar] [CrossRef]
- Sui, J.; Yu, Q.; Yang, K.; Yang, J.; Li, C.; Liu, X. Effects of Bacillus Subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar. Biology 2022, 11, 791. [Google Scholar] [CrossRef]
- Sui, J.; Yang, J.; Li, C.; Zhang, L.; Hua, X. Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar. Microorganisms 2023, 11, 486. [Google Scholar] [CrossRef] [PubMed]
- Timm, C.M.; Pelletier, D.A.; Jawdy, S.S.; Gunter, L.E.; Henning, J.A.; Engle, N.; Aufrecht, J.; Gee, E.; Nookaew, I.; Yang, Z.; et al. Two Poplar-Associated Bacterial Isolates Induce Additive Favorable Responses in a Constructed Plant-Microbiome System. Front. Plant Sci. 2016, 7, 497. [Google Scholar] [CrossRef] [PubMed]
- Ciadamidaro, L.; Pfendler, S.; Girardclos, O.; Zappelini, C.; Binet, P.; Bert, V.; Khasa, D.; Blaudez, D.; Chalot, M. Mycorrhizal Inoculation Effects on Growth and the Mycobiome of Poplar on Two Phytomanaged Sites after 7-Year-Short Rotation Coppicing. Front. Plant Sci. 2022, 13, 993301. [Google Scholar] [CrossRef] [PubMed]
- Labbé, J.L.; Weston, D.J.; Dunkirk, N.; Pelletier, D.A.; Tuskan, G.A. Newly Identified Helper Bacteria Stimulate Ectomycorrhizal Formation in Populus. Front. Plant Sci. 2014, 5, 579. [Google Scholar] [CrossRef]
- Kandel, S.L.; Firrincieli, A.; Joubert, P.M.; Okubara, P.A.; Leston, N.D.; McGeorge, K.M.; Mugnozza, G.S.; Harfouche, A.; Kim, S.-H.; Doty, S.L. An In Vitro Study of Bio-Control and Plant Growth Promotion Potential of Salicaceae Endophytes. Front. Microbiol. 2017, 8, 386. [Google Scholar] [CrossRef]
- Pfabel, C.; Eckhardt, K.-U.; Baum, C.; Struck, C.; Frey, P.; Weih, M. Impact of Ectomycorrhizal Colonization and Rust Infection on the Secondary Metabolism of Poplar (Populus trichocarpa × Deltoides). Tree Physiol. 2012, 32, 1357–1364. [Google Scholar] [CrossRef]
- Plett, J.M.; Khachane, A.; Ouassou, M.; Sundberg, B.; Kohler, A.; Martin, F. Ethylene and Jasmonic Acid Act as Negative Modulators during Mutualistic Symbiosis between L Accaria bicolor and P. opulus Roots. New Phytol. 2014, 202, 270–286. [Google Scholar] [CrossRef]
- Basso, V.; Kohler, A.; Miyauchi, S.; Singan, V.; Guinet, F.; Šimura, J.; Novák, O.; Barry, K.W.; Amirebrahimi, M.; Block, J.; et al. An Ectomycorrhizal Fungus Alters Sensitivity to Jasmonate, Salicylate, Gibberellin, and Ethylene in Host Roots. Plant Cell Environ. 2020, 43, 1047–1068. [Google Scholar] [CrossRef]
- Jiang, D.; Lin, R.; Tan, M.; Yan, J.; Yan, S. The Mycorrhizal-Induced Growth Promotion and Insect Resistance Reduction in Populus Alba × P. Berolinensis Seedlings: A Multi-Omics Study. Tree Physiol. 2022, 42, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Aleti, G.; Naidu, R.; Puschenreiter, M.; Mahmood, Q.; Rahman, M.M.; Wang, F.; Shaheen, S.; Syed, J.H.; Reichenauer, T.G. Microbe and Plant Assisted-Remediation of Organic Xenobiotics and Its Enhancement by Genetically Modified Organisms and Recombinant Technology: A Review. Sci. Total Environ. 2018, 628–629, 1582–1599. [Google Scholar] [CrossRef] [PubMed]
- Mahaffee, W.F.; Kloepper, J.W. Bacterial communities of the rhizosphere and endorhiza associated with field-grown cucumber plants inoculated with a plant growth-promoting rhizobacterium or its genetically modified derivative. Can. J. Microbiol. 1997, 43, 344–353. [Google Scholar] [CrossRef] [PubMed]
- De Cárcer, D.A.; Martín, M.; Mackova, M.; Macek, T.; Karlson, U.; Rivilla, R. The Introduction of Genetically Modified Microorganisms Designed for Rhizoremediation Induces Changes on Native Bacteria in the Rhizosphere but Not in the Surrounding Soil. ISME J. 2007, 1, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pan, L.; Ren, M.; Li, J.; Guan, X.; Tao, J. Comparison of Genetically Modified Insect-Resistant Maize and Non-Transgenic Maize Revealed Changes in Soil Metabolomes but Not in Rhizosphere Bacterial Community. GM Crops Food 2022, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.C.F.; Dini-Andreote, F.; Hannula, S.E.; Andreote, F.D.; Pereira E Silva, M.D.C.; Salles, J.F.; De Boer, W.; Van Veen, J.; Van Elsas, J.D. Different Selective Effects on Rhizosphere Bacteria Exerted by Genetically Modified versus Conventional Potato Lines. PLoS ONE 2013, 8, e67948. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lv, G.B.; Chen, K.; Yu, Q.; Niu, B.; Jiang, J.; Liu, G. Impact of PaGLK Transgenic Poplar on Microbial Community and Soil Enzyme Activity in Rhizosphere Soil. Front. Bioeng. Biotechnol. 2022, 10, 965209. [Google Scholar] [CrossRef]
- Hily, J.; Demanèche, S.; Poulicard, N.; Tannières, M.; Djennane, S.; Beuve, M.; Vigne, E.; Demangeat, G.; Komar, V.; Gertz, C.; et al. Metagenomic-based Impact Study of Transgenic Grapevine Rootstock on Its Associated Virome and Soil Bacteriome. Plant Biotechnol. J. 2018, 16, 208–220. [Google Scholar] [CrossRef]
- Krause, S.M.B.; Näther, A.; Ortiz Cortes, V.; Mullins, E.; Kessel, G.J.T.; Lotz, L.A.P.; Tebbe, C.C. No Tangible Effects of Field-Grown Cisgenic Potatoes on Soil Microbial Communities. Front. Bioeng. Biotechnol. 2020, 8, 603145. [Google Scholar] [CrossRef]
- Li, P.; Ye, S.; Chen, J.; Wang, L.; Li, Y.; Ge, L.; Wu, G.; Song, L.; Wang, C.; Sun, Y.; et al. Combined Metagenomic and Metabolomic Analyses Reveal That Bt Rice Planting Alters Soil C-N Metabolism. ISME Commun. 2023, 3, 4. [Google Scholar] [CrossRef]
- Tsatsakis, A.M.; Nawaz, M.A.; Kouretas, D.; Balias, G.; Savolainen, K.; Tutelyan, V.A.; Golokhvast, K.S.; Lee, J.D.; Yang, S.H.; Chung, G. Environmental Impacts of Genetically Modified Plants: A Review. Environ. Res. 2017, 156, 818–833. [Google Scholar] [CrossRef] [PubMed]
- Lottmann, J.; O’Callaghan, M.; Baird, D.; Walter, C. Bacterial and Fungal Communities in the Rhizosphere of Field-Grown Genetically Modified Pine Trees (Pinus radiata D.). Environ. Biosaf. Res. 2010, 9, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, L.; Lohaus, G.; Sirrenberg, A.; Karlovsky, P.; Bastien, C.; Pilate, G.; Polle, A. Ectomycorrhizal Colonization and Diversity in Relation to Tree Biomass and Nutrition in a Plantation of Transgenic Poplars with Modified Lignin Biosynthesis. PLoS ONE 2013, 8, e59207. [Google Scholar] [CrossRef] [PubMed]
- Beckers, B.; Op De Beeck, M.; Weyens, N.; Van Acker, R.; Van Montagu, M.; Boerjan, W.; Vangronsveld, J. Lignin Engineering in Field-Grown Poplar Trees Affects the Endosphere Bacterial Microbiome. Proc. Natl. Acad. Sci. USA 2016, 113, 2312–2317. [Google Scholar] [CrossRef] [PubMed]
- Stefani, F.O.P.; Moncalvo, J.-M.; Séguin, A.; Bérubé, J.A.; Hamelin, R.C. Impact of an 8-Year-Old Transgenic Poplar Plantation on the Ectomycorrhizal Fungal Community. Appl. Environ. Microbiol. 2009, 75, 7527–7536. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, L.; Thürmer, A.; Meinicke, P.; Buée, M.; Morin, E.; Martin, F.; Pilate, G.; Daniel, R.; Polle, A.; Reich, M. Fungal Soil Communities in a Young Transgenic Poplar Plantation Form a Rich Reservoir for Fungal Root Communities. Ecol. Evol. 2012, 2, 1935–1948. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.; Ding, C.; Zhang, B.; Huang, Q.; Huang, R.; Su, X. Endophytic Communities of Transgenic Poplar Were Determined by the Environment and Niche Rather Than by Transgenic Events. Front. Microbiol. 2019, 10, 588. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Ge, L.; Hu, C.; Wu, G.; Sun, Y.; Song, L.; Wu, X.; Pan, A.; Xu, Q.; et al. Environmental Behaviors of Bacillus Thuringiensis (Bt) Insecticidal Proteins and Their Effects on Microbial Ecology. Plants 2022, 11, 1212. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, J.; Chen, X.; Lv, J.; Jia, H.; Zhao, S.; Lu, M. An Empirical Assessment of Transgene Flow from a Bt Transgenic Poplar Plantation. PLoS ONE 2017, 12, e0170201. [Google Scholar] [CrossRef]
- Zuo, L.; Yang, R.; Zhen, Z.; Liu, J.; Huang, L.; Yang, M. A 5-Year Field Study Showed No Apparent Effect of the Bt Transgenic 741 Poplar on the Arthropod Community and Soil Bacterial Diversity. Sci. Rep. 2018, 8, 1956. [Google Scholar] [CrossRef]
- Wang, P.; Wei, H.; Sun, W.; Li, L.; Zhou, P.; Li, D.; Qiang, Z. Effects of Bt-Cry1Ah1 Transgenic Poplar on Target and Non-Target Pests and Their Parasitic Natural Enemy in Field and Laboratory Trials. Forests 2020, 11, 1255. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
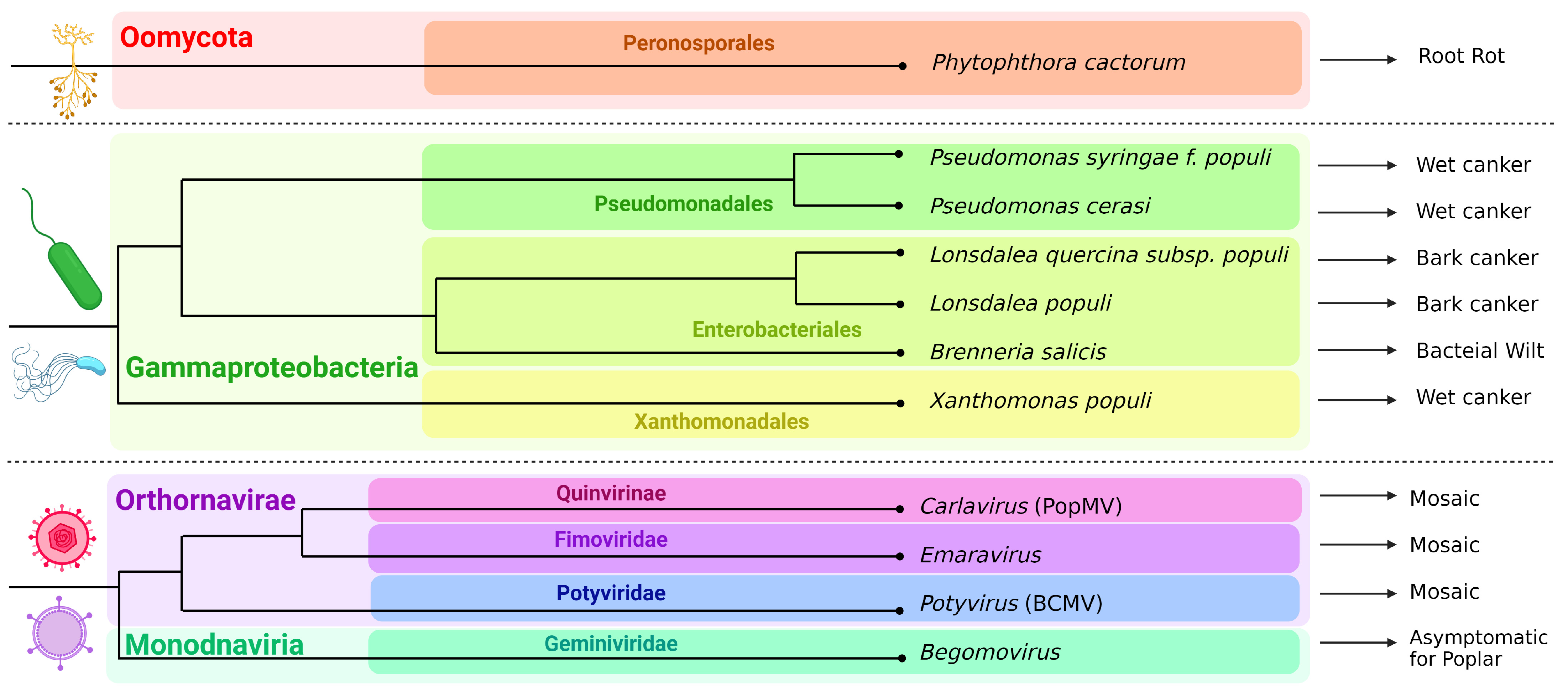{kind=link}
{kind=link}
{kind=link}
{kind=link}
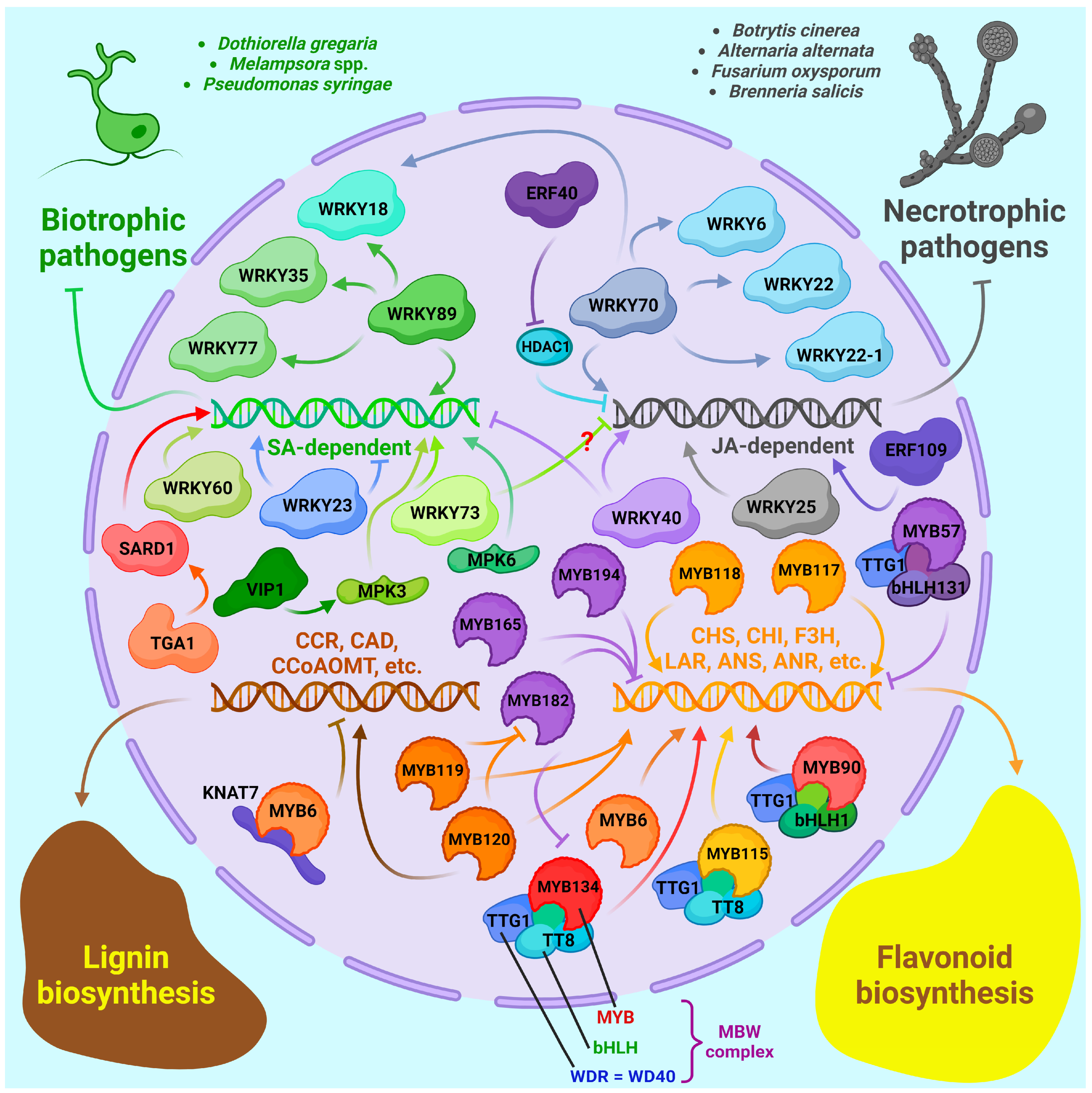{kind=link}
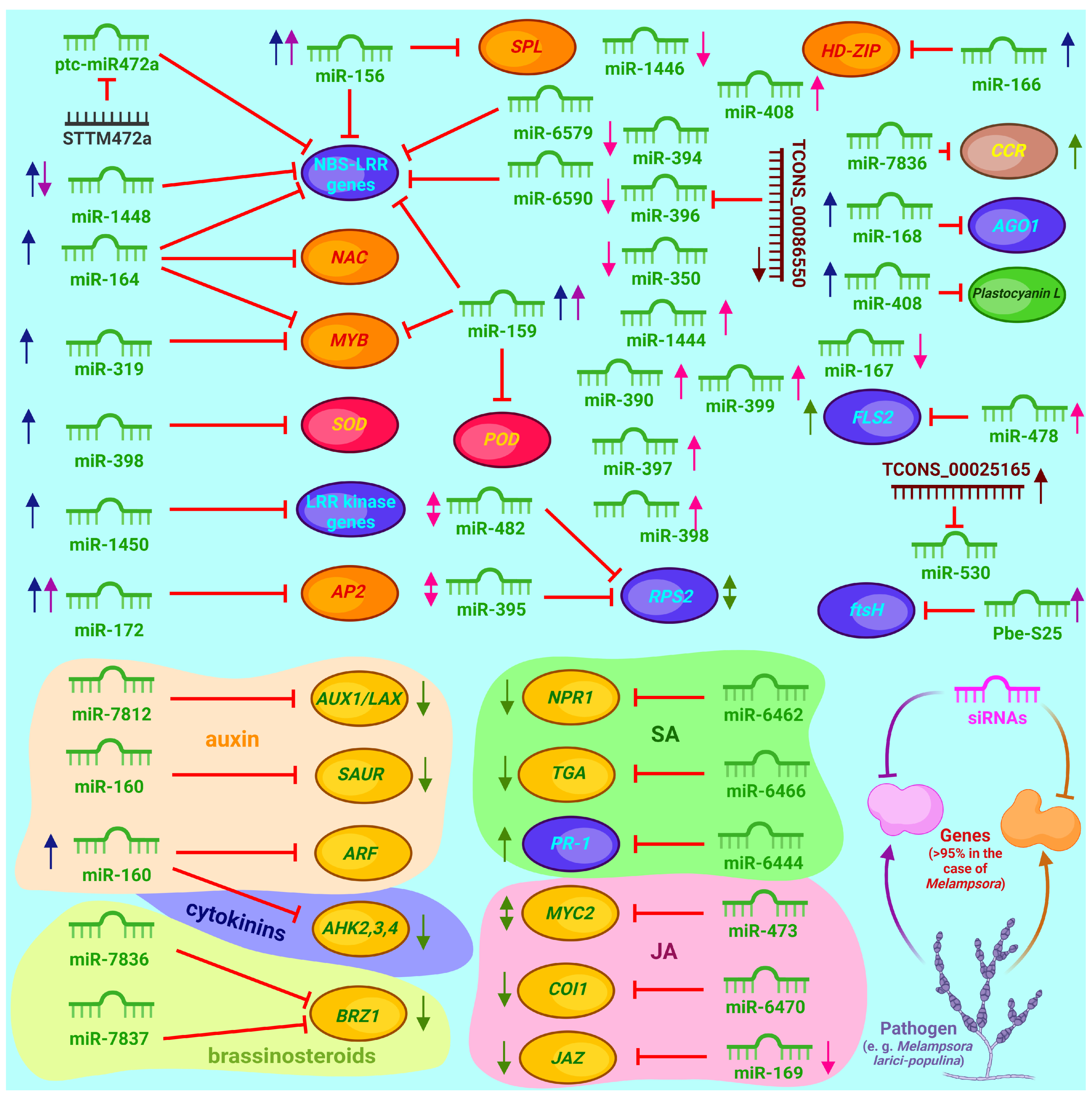{kind=link}
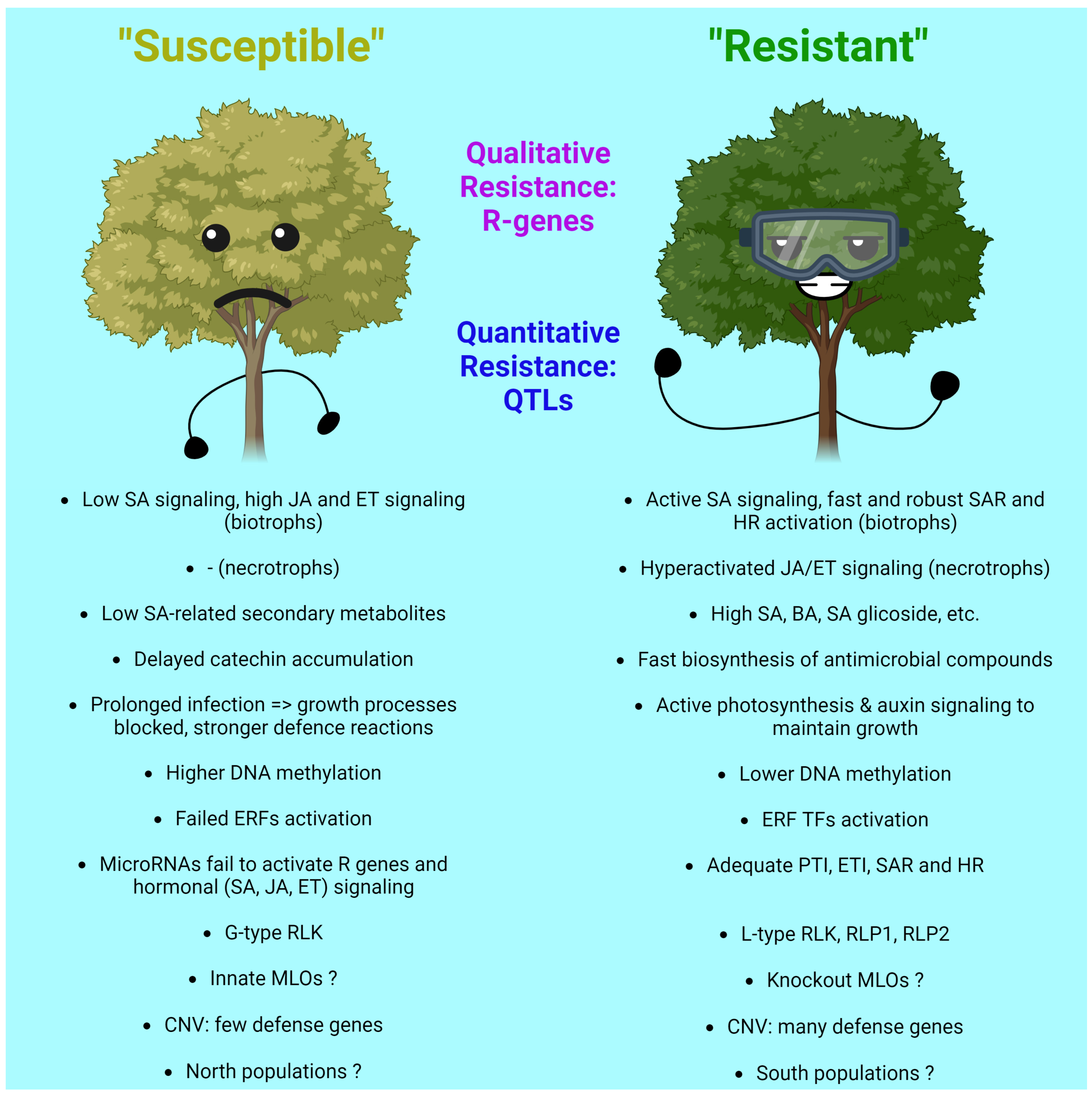{kind=link}
{kind=link}
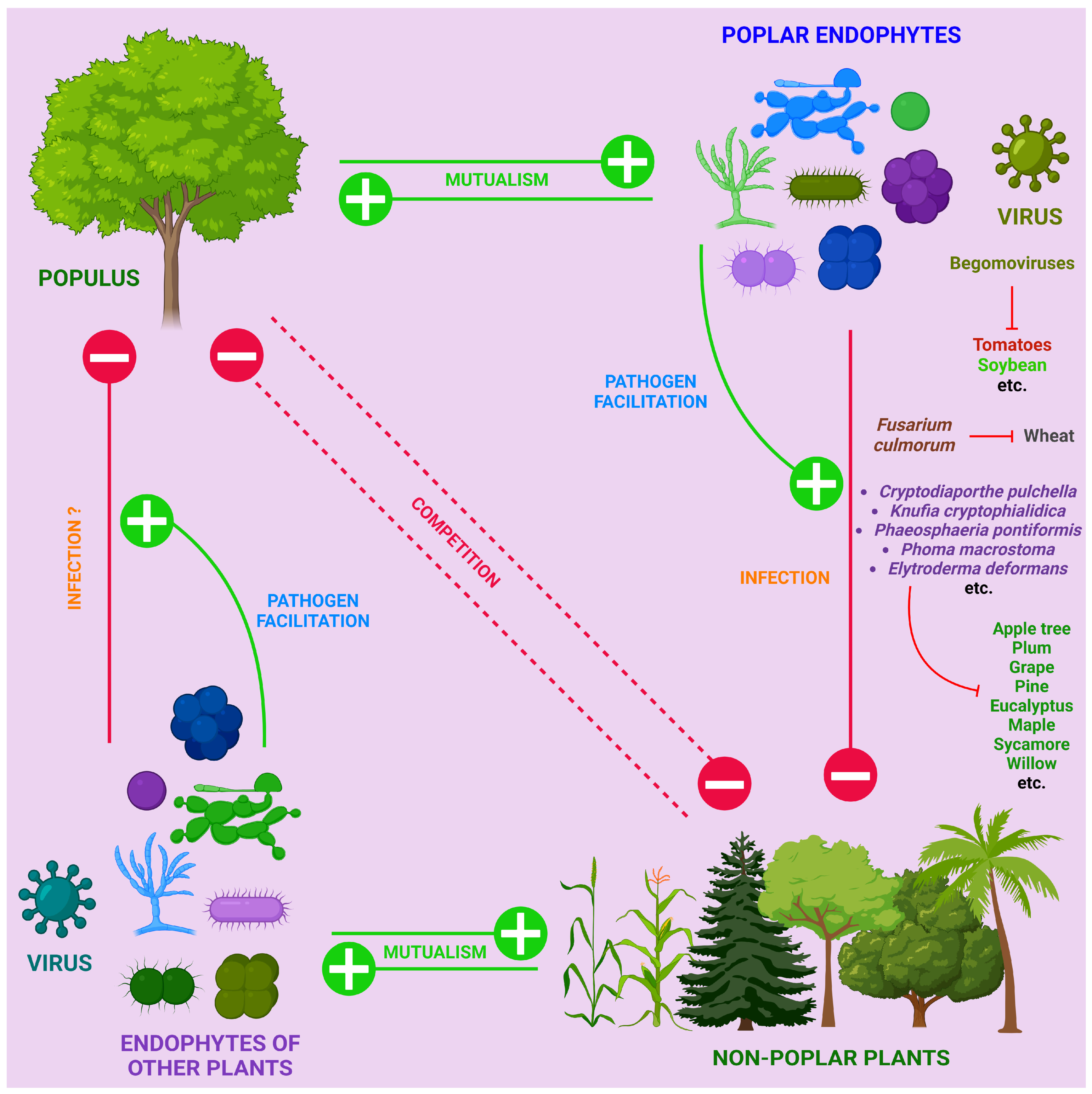{kind=link}
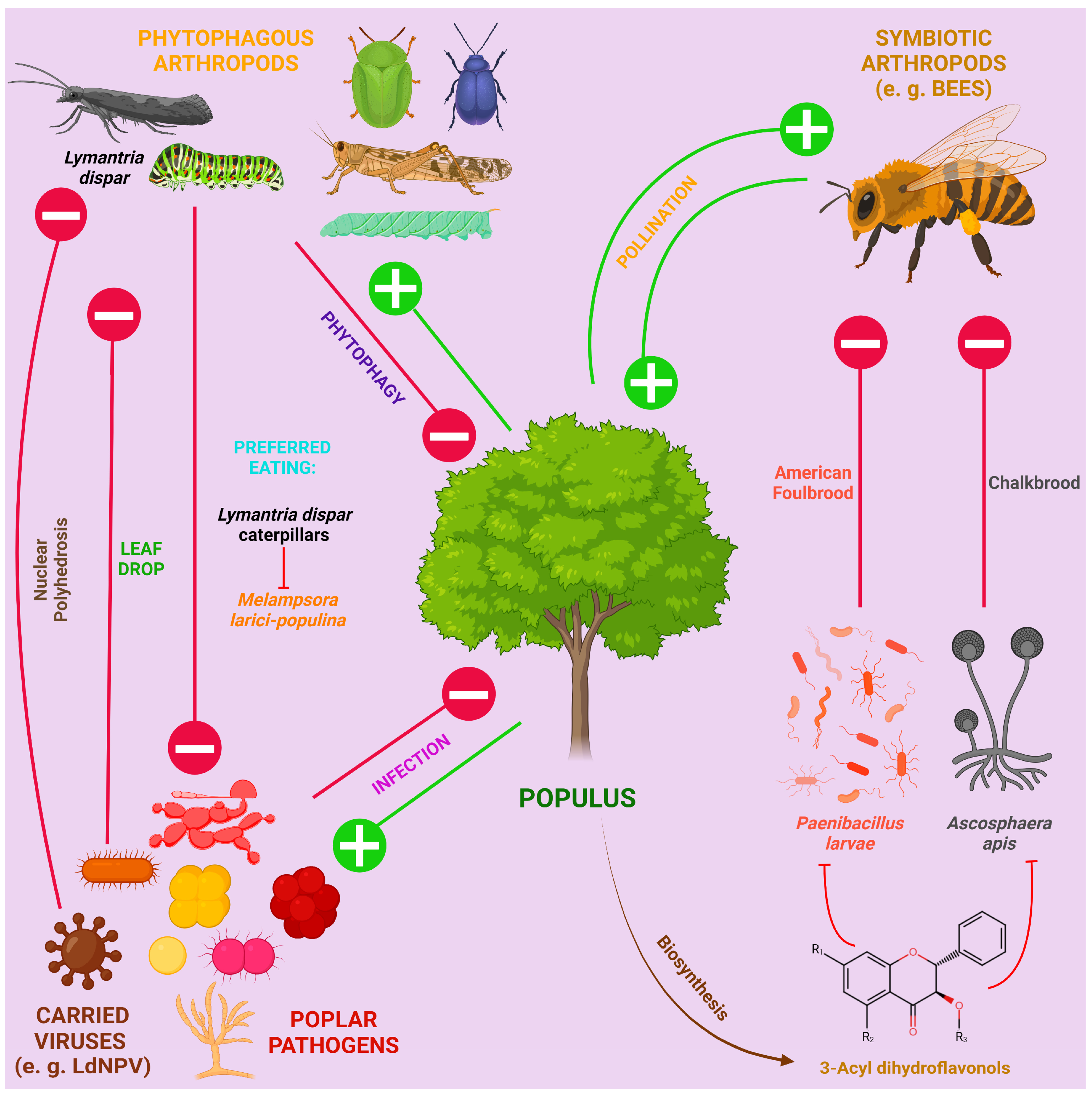{kind=link}
{kind=link}
| Article № | Poplar Species | Pathogen | Gene | Transgene | Method of Genetic Engineering | Change in Expression | Change in Resistance | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1 | P. davidiana× P. bolleana | M.brunnea | AtRLP23 | YES | A | ↑ | ↑ | Recognition of additional PAMP, which cannot be detected by poplar receptors—NLP24 | [53] |
| E. australis | ↑ | ||||||||
| 2 | P. tomentosa | D. gregaria | PtoACO7 | NO | ↑ | ↑ | Increased ET biosynthesis, levels and signaling | [117] | |
| 3 | P. trichocarpa | S. populiperda | PtDIR11 | NO | ↑ | ↑ | Increased content of lignans and flavonoids, activation of JA and ET pathways | [147] | |
| 4 | P. tomentosa | M. brunnea | PtrLAR3 | NO * | A | ↑ | ↑ | Increased proanthocyanidin content | [172] |
| 5 | P. trichocarpa | S. populiperda | PtDef | NO | A | ↑ | ↑ | Defensin toxicity to fungus, increased SA and JA activity, increased PRs and nsLTPs accumulation, increased HR amplitude and H2O2 accumulation | [42] |
| 6, 7 | P. × euramericana | NO * | A | ↑ | ↑ | [174,311] | |||
| 8 | P. tomentosa | A. alternata | LJAMP2 | YES | A | ↑ | ↑ | LJAMP2 antifungal toxicity, maybe plant immunity activation as nsLTP | [176] |
| C. gloeosporioides | ↑ | ||||||||
| 9 | P. nigra × P. maximowiczii | S. musiva | MsrA2 | YES | A | ↑ | ↑ | MsrA2 (modified dermaseptin β1) is toxic to fungi | [179] |
| 10 | P. tomentosa | C. chrysosperma | BbChit1 | YES | A | ↑ | ↑ | Chitinase activity => fungal cell wall degradation | [181] |
| 11 | P. tomentosa | A. alternata | LJAMP2 or BbChit1 | YES | A | ↑ | ↑ | Same to articles 10 or 11 | [182] |
| LJAMP2 + BbChit1 | A | ↑ + ↑ | ↑ ** | Combined action of LJAMP2 and BbChit1 | |||||
| 12 | P. deltoides × P. euramericana | M. brunnea | PeTLP | NO | A | ↑ | ↑ | Activation of other defense proteins | [183] |
| 13 | P. davidiana × P. alba | C. chrysosperma | TaHsp24 | YES | A | ↑ | ↑ | Increased activity of SA and JA pathways and PRs content | [186] |
| A. alternata | ↑ | ||||||||
| 14 | P. deltoides × P. euroamericana | B. dothidea or A. alternata | PdePrx12 | NO | A | ↑ | ↓ | H2O2 scavenging => H2O2 content reduced in overexpressor and increased in underexpressor line | [187] |
| A | ↓ | ↑ | |||||||
| 15 | P. tomentosa | D. gregaria | PtrWRKY89 | NO * | A | ↑ | ↑ | Activation of SA signaling and PR genes expression with no effect on JA signaling | [189] |
| 16 | P. trichocarpa | NO | A | ↑ | ↑ | [192] | |||
| M. brunnea | ↑ | ||||||||
| 17, 18 | P. simonii × P. nigra | A. alternata | PsnWRKY70 | NO | A | ↑ | ↑ | Activation of PTI, ETI and SA | [193] |
| 19 | P. tomentosa | D. gregaria | PtrWRKY40 | NO * | A | ↑ | ↓ | Activation of JA, but inactivation of SA signaling => increased resistance to necrotrophs, but decreased to biotrophs | [194] |
| Arabidopsis thaliana | B. cinerea | YES | A | ↑ | ↑ | ||||
| 20 | P. tomentosa | D. gregaria | PtoWRKY60 | NO | A | ↑ | ↑ | Activation of SA signaling, no effect on JA | [195] |
| 21 | Arabidopsis thaliana | P. syringae | PtrWRKY73 | YES | A | ↑ | ↑ | Activation of SA => increased resistance to biotrophs, but decreased to necrotrophs | [196] |
| B. cinerea | A | ↑ | ↓ | ||||||
| 22 | P. tremula × P. alba | Melampsora sp. | PtWRKY23 | NO | A | ↓ (RNAi) | ↓ | WRKY23 expression is at an optimal level in the WT plant and should be subject to adequate and flexible regulation? | [197] |
| A | ↑ | ||||||||
| 23 | P. simonii | A. alternata | PsnWRKY25 | NO | A | ↑ | ↑ | Activation of PsCERK1, PR1 and secondary metabolism | [198] |
| 24 | P. tremula × P. tremuloides | - | MYB182 | NO | A | ↑ | - | Repression of flavonoid biosynthesis pathway => decreased flavonoid content in overexpressors and increased in knockout plants *** | [202] |
| 25 | P. tremula × P. tremuloides | - | MYB165 | NO | A | ↑ | - | [203] | |
| MYB194 | NO | A | ↑ | - | |||||
| 26 | P. tomentosa | - | PtrMYB57 | NO * | A | ↑ | - | [204] | |
| NO | CRISPR/Cas9 | ↓ (knockout) | |||||||
| 27 | P. tremula × P. alba | M. larici-populina, M. aecidiodes | MYB134 | NO | A | ↑ | ↑ | Upregulation of flavonoid biosynthesis pathway => increased flavonoid content *** | [170] |
| 28 | P. tremula × P. alba | - | NO | A | ↑ | - | [205] | ||
| P. tremula × P. tremuloides | A | ||||||||
| 29 | P. tomentosa | D. gregaria | PtoMYB115 | NO | A | ↑ | ↑ | [55] | |
| Nicotiana benthamiana | - | PtoMYB115 + PtoTT8 + PtoTGA1 | YES | A | ↑ + ↑ + ↑ | - | |||
| 30 | P. alba | B. cinerea | PalMYB90 + PalbHLH1 | NO | A | ↑ + ↑ | ↑ | [206] | |
| ↑ | |||||||||
| 31 | P. alba × P. tremula | - | PtrMYB119 | NO * | A | ↑ | - | [207] | |
| PtrMYB120 | NO * | A | ↑ | ||||||
| 32 | P. alba × P. glandulosa | - | NO * | A | ↑ | - | [208] | ||
| A | ↓ | ||||||||
| 33 | P. tomentosa | - | PtoMYB6 | NO | A | ↑ | - | [209] | |
| 34 | P. deltoides. | - | PdeMYB118 | NO | A | ↑ | - | [210] | |
| 35 | P. tremula × P. tremuloides | - | MYB117 | NO | A | ↑ | - | [211] | |
| 36 | P. tomentosa | - | PtoMYB142 | NO | A | ↑ | - | Increased wax content *** | [212] |
| 37 | P. davidiana × P. alba | F. oxysporum | PdPapERF109 | NO | A | ↑ | ↑ | Increase in ROS-scavenging activity => more adequate immune response | [219] |
| 38 | P. trichocarpa | B. salicis | AtVIP1 | YES | A | ↑ | ↑ | PR1 activation; full molecular mechanism is unknown | [85] |
| PtVIP1 | NO | A | ↑ | ||||||
| 39 | P. tomentosa | C. gloeosporioides | PeTGA1 | NO * | ↑ | ↑ | PeSARD activation => upregulation of SA biosynthesis | [223] | |
| 40 | P. trichocarpa | C. gloeosporioides | ptc-miR472 | NO | A | ↑ | ↓ | Fail to activate ETI, NBS-LRRs, ROS => susceptible to biotrophs; active JA/ET signaling => resistant to necrotrophs | [241] |
| C. chrysosperma | ↑ | ||||||||
| C. gloeosporioides | A | ↓ (via STTM) | ↑ | Hyperactivated NBS-LRRs => quick and robust ETI and SA response | |||||
| C. chrysosperma | ~unchanged | ||||||||
| 41 | P. alba × P. grandidentata | M. aecidiodes | AtGolS + CsRFS | YES | A | ↑ | ↓ | Suppressed SA, Ca2+, phosphatidic acid, activated JA signaling => failed ROS and PR1 accumulation => increased susceptibility to biotrophs (but resistance to necrotrophs?) | [262] |
| 42 | P. davidiana × P. bollena | A. alternata | PdbLOX2 | NO | A | ↑ | ↑ | Hyperaccumulation of JA => hyper-resistance to necrotrophs | [263] |
| 43 | P. tremula × P. alba, P. deltoides | - | LecRLK-G, TPX2 | NO | CRISPRa (dCas9) | ↑ | - | More active immune response *** | [312] |
| - | PLATZ | NO | CBE (nCas9) | ↓ | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovalev, M.A.; Gladysh, N.S.; Bogdanova, A.S.; Bolsheva, N.L.; Popchenko, M.I.; Kudryavtseva, A.V. Editing Metabolism, Sex, and Microbiome: How Can We Help Poplar Resist Pathogens? Int. J. Mol. Sci. 2024, 25, 1308. https://doi.org/10.3390/ijms25021308
Kovalev MA, Gladysh NS, Bogdanova AS, Bolsheva NL, Popchenko MI, Kudryavtseva AV. Editing Metabolism, Sex, and Microbiome: How Can We Help Poplar Resist Pathogens? International Journal of Molecular Sciences. 2024; 25(2):1308. https://doi.org/10.3390/ijms25021308
Chicago/Turabian StyleKovalev, Maxim A., Natalya S. Gladysh, Alina S. Bogdanova, Nadezhda L. Bolsheva, Mikhail I. Popchenko, and Anna V. Kudryavtseva. 2024. "Editing Metabolism, Sex, and Microbiome: How Can We Help Poplar Resist Pathogens?" International Journal of Molecular Sciences 25, no. 2: 1308. https://doi.org/10.3390/ijms25021308
APA StyleKovalev, M. A., Gladysh, N. S., Bogdanova, A. S., Bolsheva, N. L., Popchenko, M. I., & Kudryavtseva, A. V. (2024). Editing Metabolism, Sex, and Microbiome: How Can We Help Poplar Resist Pathogens? International Journal of Molecular Sciences, 25(2), 1308. https://doi.org/10.3390/ijms25021308