
The Antioxidant Properties of Glucosinolates in Cardiac Cells Are Independent of H2S Signaling



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
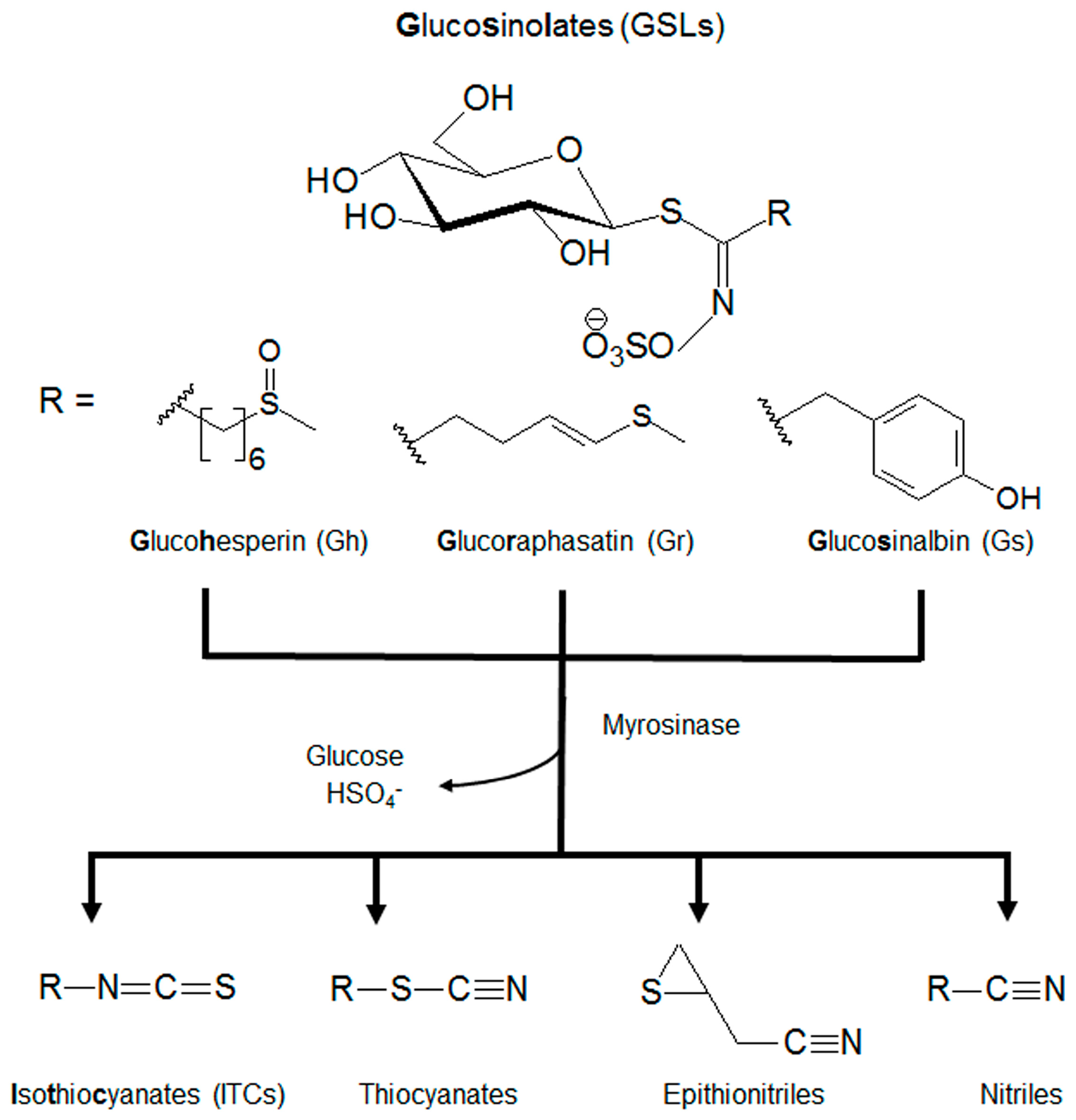
:1. Introduction
2. Results
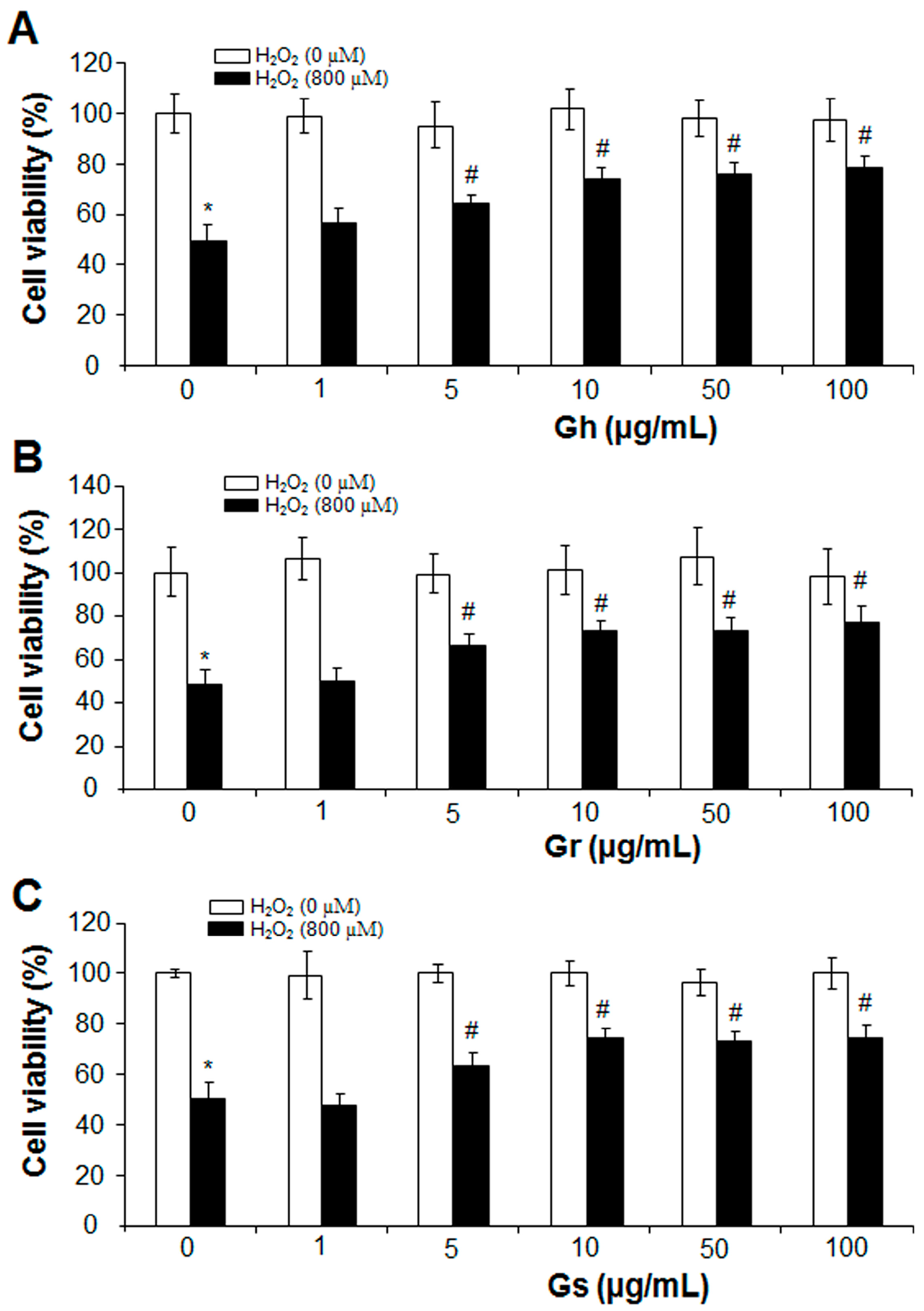
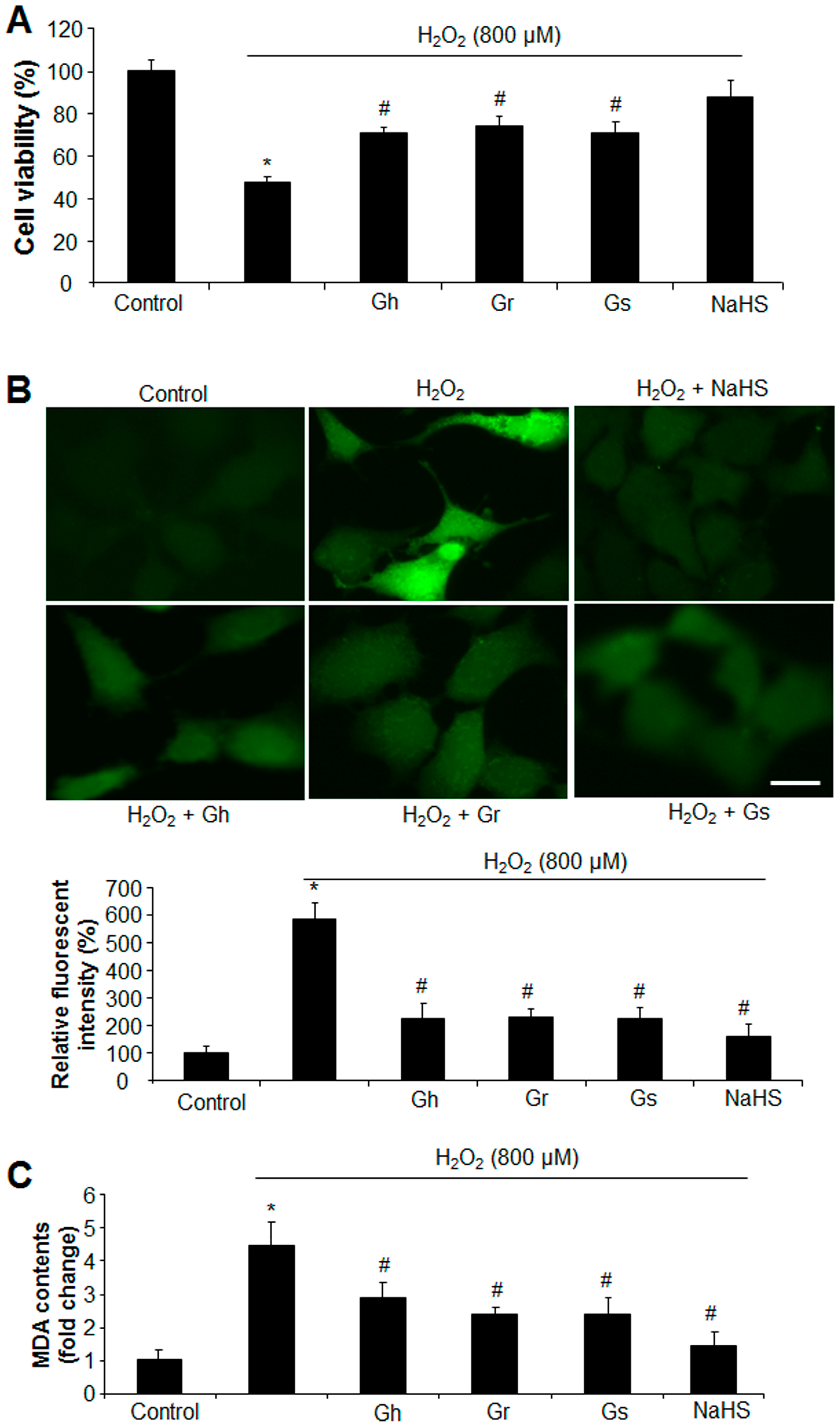
2.1. GSLs and H2S Protected H2O2-Induced Cell Death and Oxidative Stress
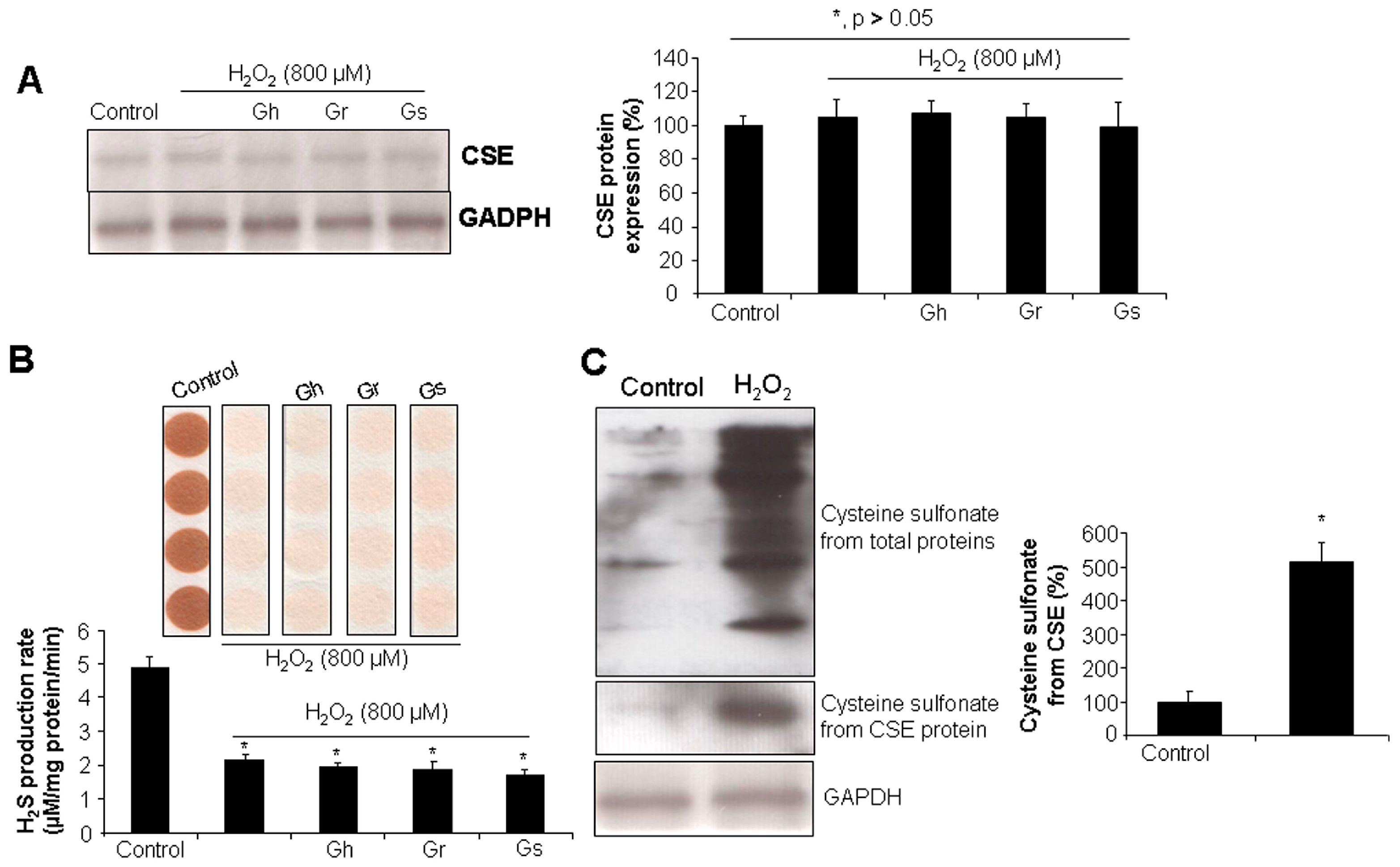
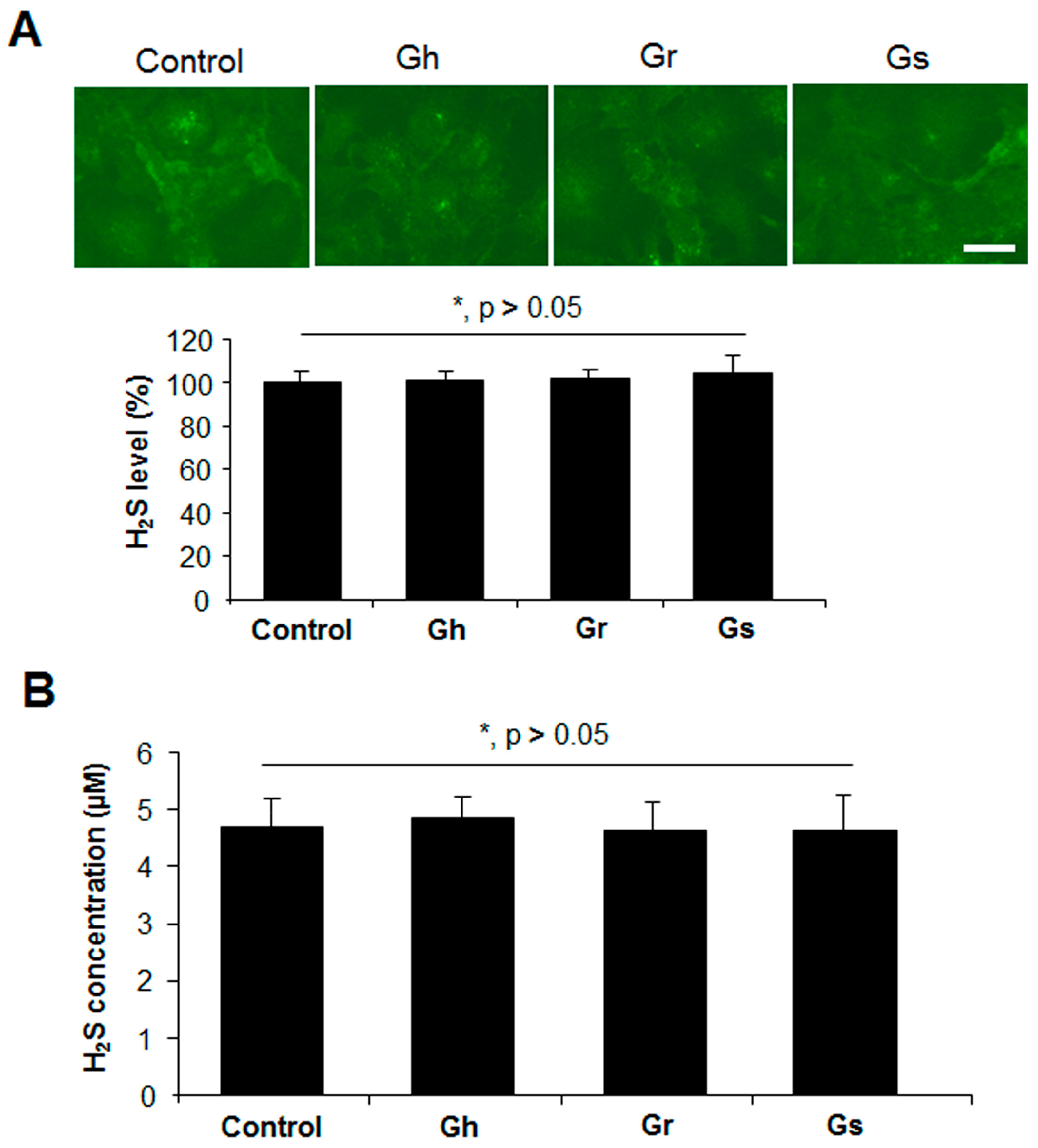
2.2. GSLs Had No Effect on CSE Protein Expression and H2S Generation in H9C2 Cells
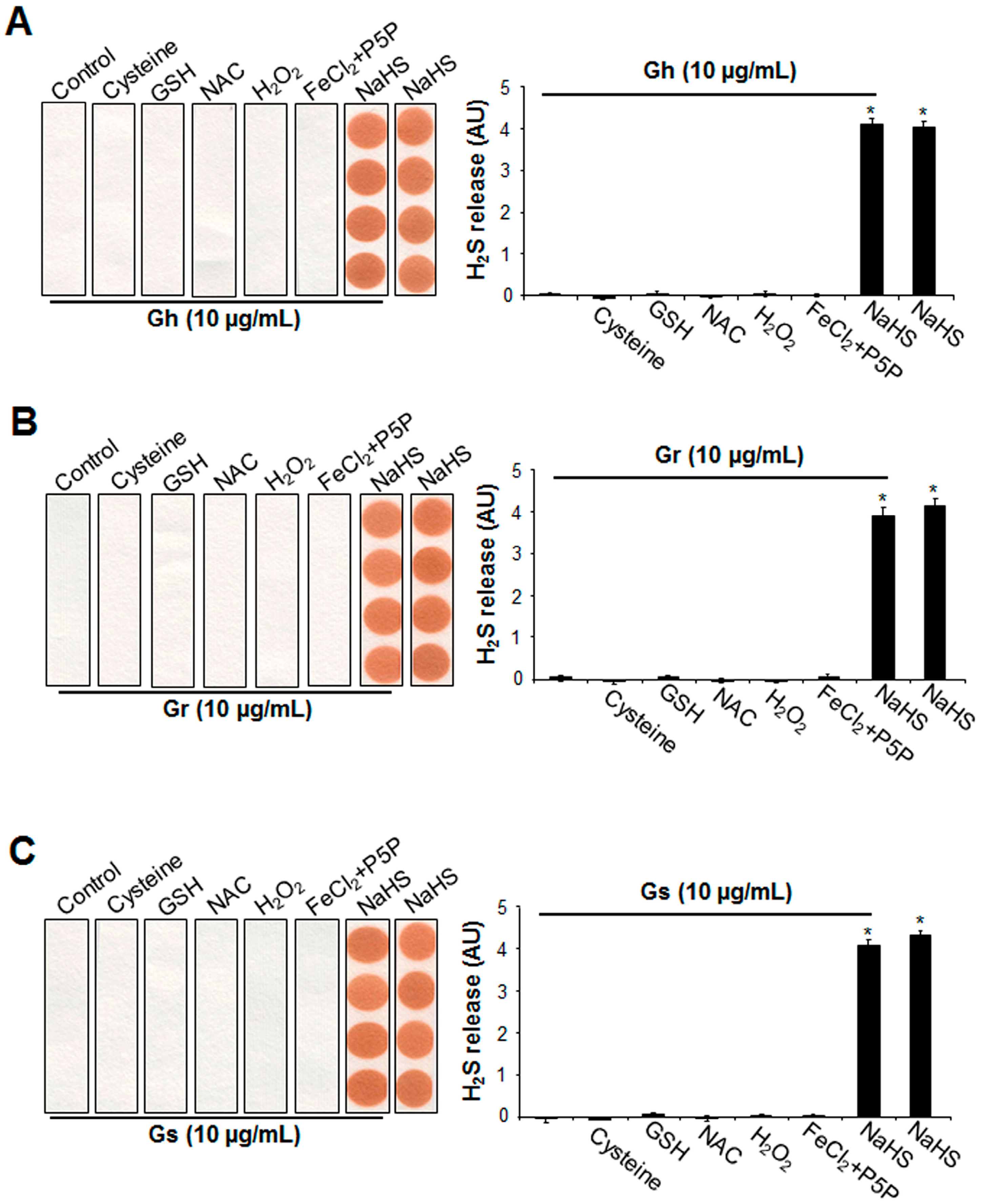
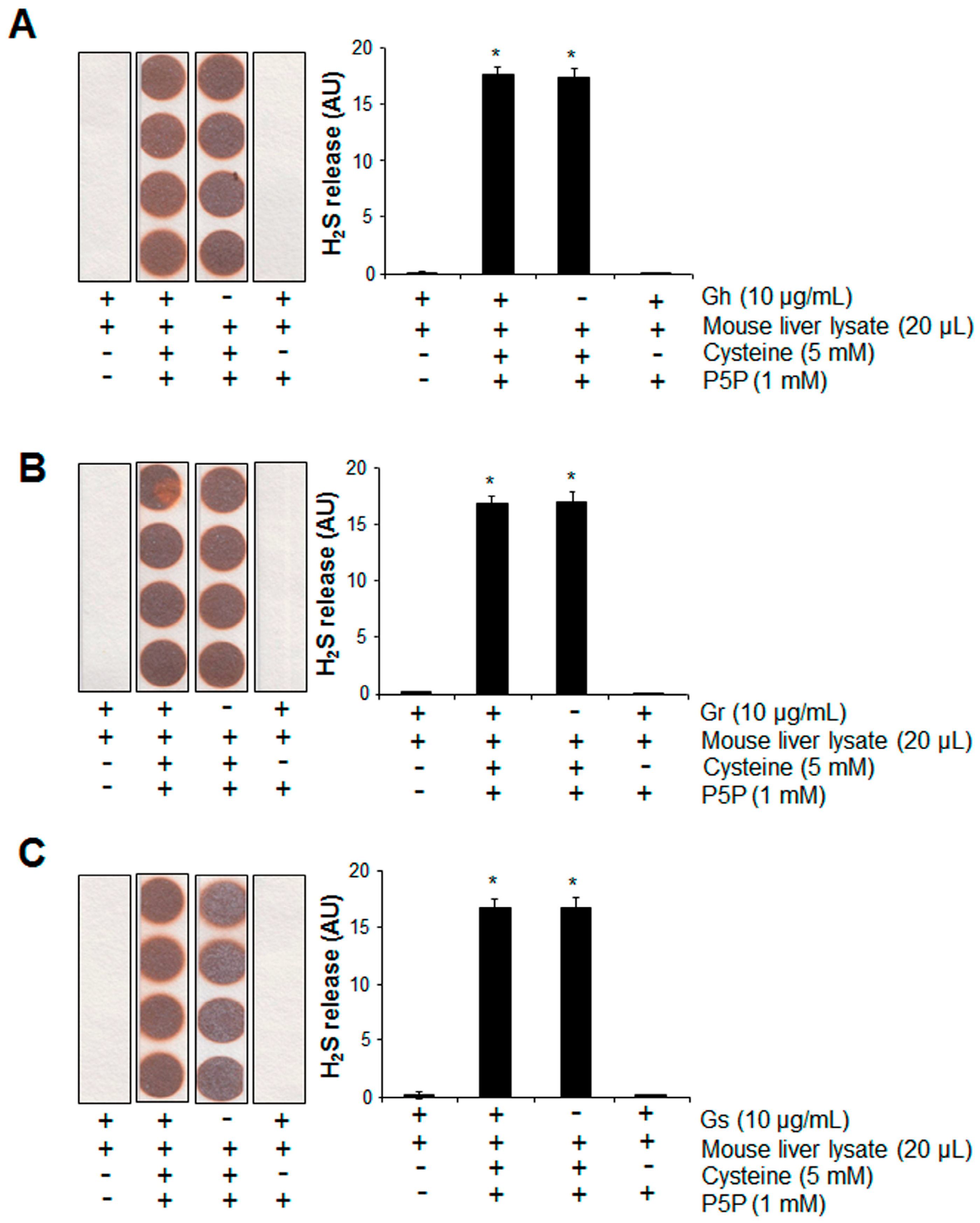
2.3. No H2S Was Released from GSLs under Various Tested Conditions
3. Discussion
4. Materials and Methods
4.1. Isolation of the 3 GSLs (Gh, Gr, and Gs)
4.2. Cell Culture
4.3. Animal Care and Tissue Collection
4.4. Cell Viability
4.5. Measurement of H2S Production
4.6. Western Blotting
4.7. Detection of Oxidative Stress
4.8. Co-IP Assay
4.9. Malondialdehyde (MDA) Content
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health benefits of plant-derived sulfur compounds, glucosinolates, and organosulfur compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.M.; Abdull Razis, A.F.; Mohd Sukri, N.S.; Perimal, E.K.; Ahmad, H.; Rollin, P.; Djedaini-Pilard, F.; Mazzon, E.; Rigaud, S. Beneficial health effects of glucosinolates-derived isothiocyanates on cardiovascular and neurodegenerative diseases. Molecules 2022, 27, 624. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Subramanian, P.; Hahn, B.S. Glucosinolate diversity analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for functional food breeding. Foods 2023, 12, 2400. [Google Scholar] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Armah, C.N.; Derdemezis, C.; Traka, M.H.; Dainty, J.R.; Doleman, J.F.; Saha, S.; Leung, W.; Potter, J.F.; Lovegrove, J.A.; Mithen, R.F. Diet rich in high glucoraphanin broccoli reduces plasma LDL cholesterol: Evidence from randomised controlled trials. Mol. Nutr. Food Res. 2015, 59, 918–926. [Google Scholar] [CrossRef] [PubMed]
- García-Ibañez, P.; Yepes-Molina, L.; Ruiz-Alcaraz, A.J.; Martínez-Esparza, M.; Moreno, D.A.; Carvajal, M.; García-Peñarrubia, P. Brassica bioactives could ameliorate the chronic inflammatory condition of endometriosis. Int. J. Mol. Sci. 2020, 21, 9397. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. Anti-inflammatory and anti-apoptotic effects of (RS)-glucoraphanin bioactivated with myrosinase in murine sub-acute and acute MPTP-induced Parkinson’s disease. Bioorg. Med. Chem. 2013, 21, 5532–5547. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef]
- Nishida, M.; Sawa, T.; Kitajima, N.; Ono, K.; Inoue, H.; Ihara, H.; Motohashi, H.; Yamamoto, M.; Suematsu, M.; Kurose, H.; et al. Hydrogen sulfide anion regulates redox signaling via electrophile sulfhydration. Nat. Chem. Biol. 2012, 8, 714–724. [Google Scholar]
- Sen, U.; Sathnur, P.B.; Kundu, S.; Givvimani, S.; Coley, D.M.; Mishra, P.K.; Qipshidze, N.; Tyagi, N.; Metreveli, N.; Tyagi, S.C. Increased endogenous H2S generation by CBS, CSE, and 3MST gene therapy improves ex vivo renovascular relaxation in hyperhomocysteinemia. Am. J. Physiol. Cell Physiol. 2012, 303, C41–C51. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Kurboniyon, M.S.; Zou, Y.; Luo, K.; Fang, S.; Xia, P.; Ning, S.; Zhang, L.; Wang, C. GSH-triggered/photothermal-enhanced H2S signaling molecule release for gas therapy. Pharmaceutics 2023, 15, 2443. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef] [PubMed]
- Predmore, B.L.; Kondo, K.; Bhushan, S.; Zlatopolsky, M.A.; King, A.L.; Aragon, J.P.; Grinsfelder, D.B.; Condit, M.E.; Lefer, D.J. The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H2410–H2418. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.; Ghimenti, G.; Breschi, M.; Calderone, V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Micheli, L.; Trallori, E.; Citi, V.; Martelli, A.; Testai, L.; De Nicola, G.R.; Iori, R.; Calderone, V.; Ghelardini, C.; et al. Effect of glucoraphanin and sulforaphane against chemotherapy-induced neuropathic pain: Kv7 potassium channels modulation by H2S release in vivo. Phytother. Res. 2018, 32, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Mensah, G.A.; Roth, G.A.; Fuster, V. The global burden of cardiovascular diseases and risk factors: 2020 and beyond. J. Am. Coll. Cardiol. 2019, 74, 2529–2532. [Google Scholar] [CrossRef]
- Wojtasińska, A.; Kućmierz, J.; Tokarek, J.; Dybiec, J.; Rodzeń, A.; Młynarska, E.; Rysz, J.; Franczyk, B. New insights into cardiovascular diseases treatment based on molecular targets. Int. J. Mol. Sci. 2023, 24, 16735. [Google Scholar] [CrossRef]
- Stojanovic, D.; Stojanovic, M.; Milenkovic, J.; Velickov, A.; Ignjatovic, A.; Milojkovic, M. The multi-faceted nature of renalase for mitochondrial dysfunction improvement in cardiac disease. Cells 2023, 12, 1607. [Google Scholar] [CrossRef]
- Su, X.; Zhou, M.; Li, Y.; Zhang, J.; An, N.; Yang, F.; Zhang, G.; Yuan, C.; Chen, H.; Wu, H.; et al. Protective effects of natural products against myocardial ischemia/reperfusion: Mitochondria-targeted therapeutics. Biomed. Pharmacother. 2022, 149, 112893. [Google Scholar] [CrossRef]
- Connolly, E.L.; Sim, M.; Travica, N.; Marx, W.; Beasy, G.; Lynch, G.S.; Bondonno, C.P.; Lewis, J.R.; Hodgson, J.M.; Blekkenhorst, L.C. Glucosinolates from cruciferous vegetables and their potential role in chronic disease: Investigating the preclinical and clinical evidence. Front. Pharmacol. 2021, 12, 767975. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, G.; Zong, G.; Sampson, L.; Hu, F.B.; Willett, W.C.; Rimm, E.B.; Manson, J.E.; Rexrode, K.M.; Sun, Q. Intake of glucosinolates and risk of coronary heart disease in three large prospective cohorts of US men and women. Clin. Epidemiol. 2018, 10, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Tan, B.; Sun, Y.; Hu, Q. Cystathionine γ lyase S-sulfhydrates Drp1 to ameliorate heart dysfunction. Redox. Biol. 2022, 58, 102519. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, L.; Lu, B.; Wen, J.; Wang, M.; Zhang, S.; Li, Q.; Shu, F.; Lu, F.; Liu, N.; et al. Hydrogen sulphide reduced the accumulation of lipid droplets in cardiac tissues of db/db mice via Hrd1 S-sulfhydration. J. Cell. Mol. Med. 2021, 25, 9154–9167. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Piragine, E.; Testai, L.; Breschi, M.C.; Calderone, V.; Martelli, A. The role of hydrogen sulfide and H2S-donors in myocardial protection against ischemia/reperfusion injury. Curr. Med. Chem. 2018, 25, 4380–4401. [Google Scholar] [CrossRef] [PubMed]
- Bechelli, C.; Macabrey, D.; Deglise, S.; Allagnat, F. Clinical potential of hydrogen sulfide in peripheral arterial disease. Int. J. Mol. Sci. 2023, 24, 9955. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Mitidieri, E.; Turnaturi, C.; Vanacore, D.; Ciampi, C.; Lucarini, E.; Cirino, G.; Ghelardini, C.; Sorrentino, R.; Di Cesare Mannelli, L.; et al. Beneficial effect of H2S-releasing molecules in an in vitro model of sarcopenia: Relevance of glucoraphanin. Int. J. Mol. Sci. 2022, 23, 5955. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, X.; Lu, Y.; Liang, D.; Huang, D. Isothiocyanates as H2S donors triggered by cysteine: Reaction mechanism and structure and activity relationship. Org. Lett. 2019, 21, 5977–5980. [Google Scholar] [CrossRef]
- Gambari, L.; Barone, M.; Amore, E.; Grigolo, B.; Filardo, G.; Iori, R.; Citi, V.; Calderone, V.; Grassi, F. Glucoraphanin increases intracellular hydrogen sulfide (H2S) levels and stimulates osteogenic differentiation in human mesenchymal stromal cell. Nutrients 2022, 14, 435. [Google Scholar] [CrossRef]
- Ahn, B.K.; Ahn, Y.J.; Lee, Y.J.; Lee, Y.H.; Lee, G.J. Simple and sensitive detection of bacterial hydrogen sulfide production using a paper-based colorimetric assay. Sensors 2022, 22, 5928. [Google Scholar] [CrossRef]
- Jeddi, S.; Gholami, H.; Gheibi, S.; Kashfi, K.; Ghasemi, A. Altered gene expression of hydrogen sulfide-producing enzymes in the liver and muscles tissues of hyperthyroid rats. J. Cell. Physiol. 2019, 234, 17937–17945. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Citi, V.; Calderone, V. Vascular Effects of H2S-donors: Fluorimetric detection of H2S generation and ion channel activation in human aortic smooth muscle cells. Methods Mol. Biol. 2019, 2007, 79–87. [Google Scholar] [PubMed]
- Bibli, S.I.; Luck, B.; Zukunft, S.; Wittig, J.; Chen, W.; Xian, M.; Papapetropoulos, A.; Hu, J.; Fleming, I. A selective and sensitive method for quantification of endogenous polysulfide production in biological samples. Redox Biol. 2018, 18, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Davies, M.J.; Krämer, A.C.; Miotto, G.; Zaccarin, M.; Zhang, H.; Ursini, F. Protein cysteine oxidation in redox signaling: Caveats on sulfenic acid detection and quantification. Arch. Biochem. Biophys. 2017, 617, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, G.; Li, H.; Yan, S.; Qian, J.; Guo, X.; Li, G.; Qi, H.; Zhu, Z.; Wu, Y.; et al. H2O2-mediated oxidative stress enhances cystathionine γ-lyase-derived H2S synthesis via a sulfenic acid intermediate. Antioxidants 2021, 10, 1488. [Google Scholar] [CrossRef]
- Wen, Y.D.; Wang, H.; Kho, S.H.; Rinkiko, S.; Sheng, X.; Shen, H.M.; Zhu, Y.Z. Hydrogen sulfide protects HUVECs against hydrogen peroxide induced mitochondrial dysfunction and oxidative stress. PLoS ONE 2013, 8, e53147. [Google Scholar] [CrossRef]
- Montaut, S.; Benson, H.J.; Kay, M.; Guido, B.S.; Mahboob, S.S.; Chénier, J.; Gasparetto, J.L.; Joly, H.A. Probing the free-radical scavenging activity of the extract, the major glucosinolate and isothiocyanate of Eruca sativa Mill and Lepidium densiflorum Schrad. seeds. J. Food Compos. Anal. 2017, 61, 52–58. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, F.; Aromokunola, B.; Montaut, S.; Yang, G. The Antioxidant Properties of Glucosinolates in Cardiac Cells Are Independent of H2S Signaling. Int. J. Mol. Sci. 2024, 25, 696. https://doi.org/10.3390/ijms25020696
Harvey F, Aromokunola B, Montaut S, Yang G. The Antioxidant Properties of Glucosinolates in Cardiac Cells Are Independent of H2S Signaling. International Journal of Molecular Sciences. 2024; 25(2):696. https://doi.org/10.3390/ijms25020696
Chicago/Turabian StyleHarvey, Félix, Boluwaji Aromokunola, Sabine Montaut, and Guangdong Yang. 2024. "The Antioxidant Properties of Glucosinolates in Cardiac Cells Are Independent of H2S Signaling" International Journal of Molecular Sciences 25, no. 2: 696. https://doi.org/10.3390/ijms25020696
APA StyleHarvey, F., Aromokunola, B., Montaut, S., & Yang, G. (2024). The Antioxidant Properties of Glucosinolates in Cardiac Cells Are Independent of H2S Signaling. International Journal of Molecular Sciences, 25(2), 696. https://doi.org/10.3390/ijms25020696