
Fertilization- and Irrigation-Modified Bacterial Community Composition and Stimulated Enzyme Activity of Eucalyptus Plantations Soil

Abstract
:1. Introduction
2. Results
2.1. Effects of Fertilization and Irrigation on Enzyme Activity in the Soil of Eucalyptus Plantations
2.2. Effects of Fertilization and Irrigation on the Bacterial Diversity in the Soil of the Eucalyptus Plantations
2.3. Effects of Fertilization and Irrigation on Bacterial Community Composition in the Soil of the Eucalyptus Plantations
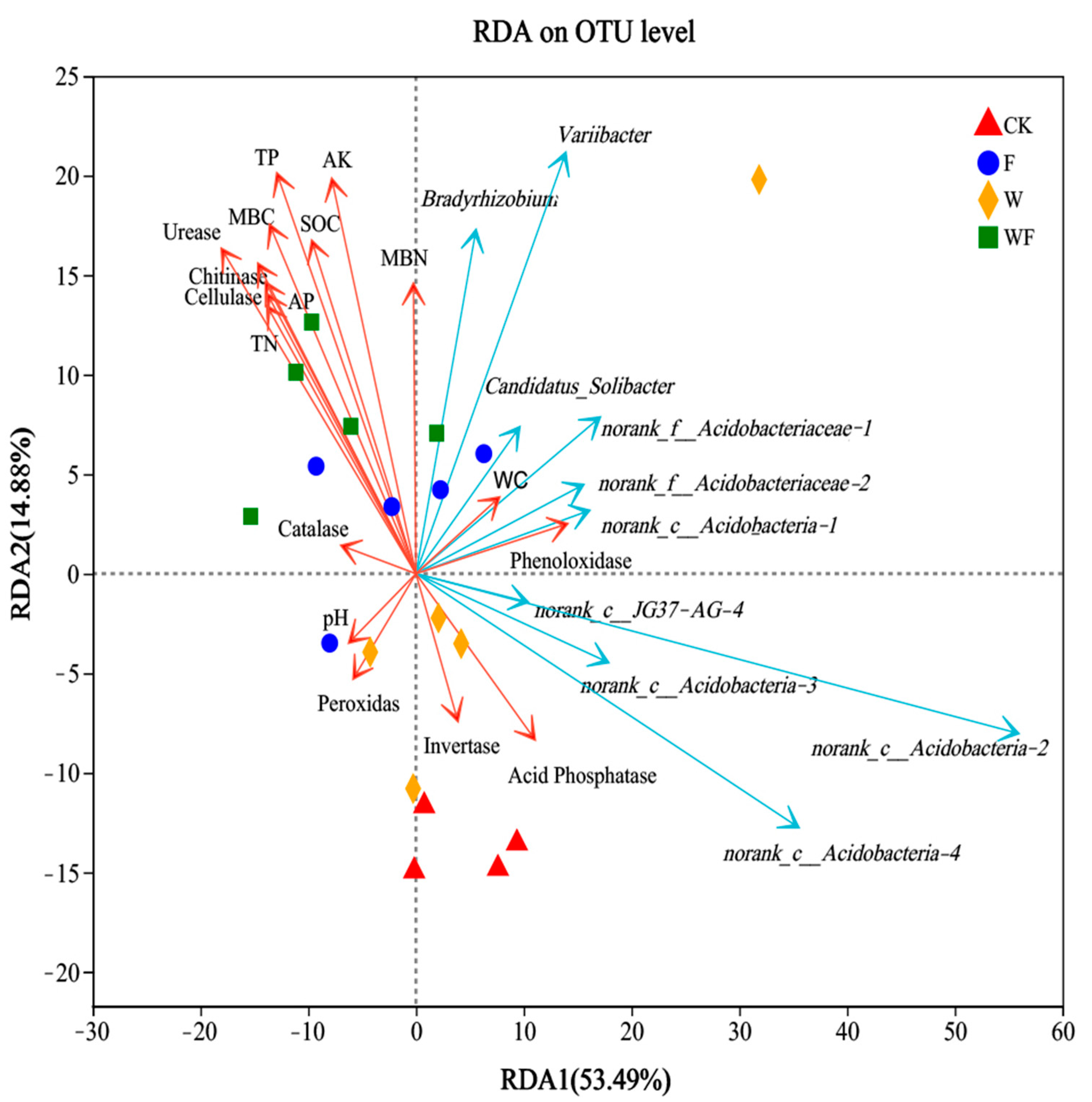
2.4. Effect of Physicochemical Factors and Enzyme Activities on the Bacterial Community Structure in the Soil of the Eucalyptus Plantations
2.5. Correlation Analysis among Management Practices–Soil Factors–Bacterial Diversity
2.6. Functional Analysis in the Soil of the Eucalyptus Plantations
3. Discussion
3.1. Effect of Irrigation and Fertilization on the Enzyme Activity in the Soil of Forests
3.2. Effect of Irrigation and Fertilization on Soil Bacterial Community Composition
3.3. Effect of Irrigation and Fertilization on the Soil Bacterial Function
4. Materials and Methods
4.1. Study Site and Experimental Materials
4.2. Experimental Method
4.3. Soil Enzyme Activity and Physicochemical Properties
4.4. Soil DNA Extraction and ILLUMINA Sequencing
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nacke, H.; Engelhaupt, M.; Brady, S.; Fischer, C.; Tautzt, J.; Daniel, R. Identification and characterization of novel cellulolytic and hemicellulolytic genes and enzymes derived from German grassland soil metagenomes. Biotechnol. Lett. 2012, 34, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Llado, S.; Lopez-Mondejar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [PubMed]
- Eichorst, S.A.; Kuske, C.R. Identification of cellulose-responsive bacterial and fungal communities in geographically and edaphically different soils by using stable isotope probing. Appl. Environ. Microbiol. 2012, 78, 2316–2327. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Mondejar, R.; Zuehlke, D.; Becher, D.; Riedel, K.; Baldrian, P. Cellulose and hemicellulose decomposition by forest soil bacteria proceeds by the action of structurally variable enzymatic systems. Sci. Rep. 2016, 6, 25279. [Google Scholar] [CrossRef] [PubMed]
- Nemergut, D.R.; Cleveland, C.C.; Wieder, W.R.; Washenberger, C.L.; Townsend, A.R. Plot-scale manipulations of organic matter inputs to soils correlate with shifts in microbial community composition in a lowland tropical rain forest. Soil Biol. Biochem. 2010, 42, 2153–2160. [Google Scholar] [CrossRef]
- Lin, Y.T.; Jangid, K.; Whitman, W.B.; Coleman, D.C.; Chiu, C.Y. Change in bacterial community structure in response to disturbance of natural hardwood and secondary coniferous forest soils in central Taiwan. Microb. Ecol. 2011, 61, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yang, X.; Li, J.; Wen, S.; Yang, L.; Ji, L.; He, G. Divergent effects of fertilizer regimes on taxonomic and functional compositions of rhizosphere bacteria and fungi in phoebe bournei Young plantations are associated with root exudates. Forests 2023, 14, 126. [Google Scholar] [CrossRef]
- Hu, X.J.; Liu, J.J.; Wei, D.; Zhu, P.; Cui, X.A.; Zhou, B.K.; Chen, X.L.; Jin, J.; Liu, X.B.; Wang, G.H. Soil bacterial communities under different long-term fertilization regimes in three locations across the black soil region of northeast China. Pedosphere 2018, 28, 751–763. [Google Scholar] [CrossRef]
- Yang, G.; Ryo, M.; Roy, J.; Hempel, S.; Rillig, M.C. Plant and soil biodiversity have non-substitutable stabilising effects on biomass production. Ecol. Lett. 2021, 24, 1582–1593. [Google Scholar] [CrossRef]
- Sui, X.; Li, M.; Frey, B.; Wang, M.; Weng, X.; Wang, X.; Chen, F.; Li, X.; Du, Z.; Yang, L. Climax forest has a higher soil bacterial diversity but lower soil nutrient contents than degraded forests in temperate northern China. Ecol. Evol. 2022, 12, e9535. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends. Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Sessitsch, A.; Howieson, J.G.; Perret, X.; Antoun, H.; Martinez-Romero, E. Advances in Rhizobium Research. Crit. Rev. Plant Sci. 2002, 21, 323–378. [Google Scholar] [CrossRef]
- Krause, S.M.B.; Dohrmann, A.B.; Gillor, O.; Christensen, B.T.; Merbach, I.; Tebbe, C.C. Soil properties and habitats determine the response of bacterial communities to agricultural wastewater irrigation. Pedosphere 2020, 30, 146–158. [Google Scholar] [CrossRef]
- Liu, Z.; Beskrovnaya, P.; Melnyk, R.A.; Hossain, S.S.; Khorasani, S.; O’Sullivan, L.R.; Wiesmann, C.L.; Bush, J.; Richard, J.D.; Haney, C.H. A genome-wide screen identifies genes in rhizosphere-associated Pseudomonas required to evade plant defenses. Mbio 2018, 9, e00433-18. [Google Scholar] [CrossRef]
- Cole, B.J.; Feltcher, M.E.; Waters, R.J.; Wetmore, K.M.; Mucyn, T.S.; Ryan, E.M.; Wang, G.; Ul-Hasan, S.; McDonald, M.; Yoshikuni, Y.; et al. Genome-wide identification of bacterial plant colonization genes. PLoS Biol. 2017, 15, 2002860. [Google Scholar] [CrossRef]
- He, J.Z.; Zheng, Y.; Chen, C.R.; He, Y.Q.; Zhang, L.M. Microbial composition and diversity of an upland red soil under long-term fertilization treatments as revealed by culture-dependent and culture-independent approaches. J. Soils Sediments 2008, 8, 349–358. [Google Scholar] [CrossRef]
- Schuetz, K.; Kandeler, E.; Nagel, P.; Scheu, S.; Ruess, L. Functional microbial community response to nutrient pulses by artificial groundwater recharge practice in surface soils and subsoils. FEMS Microbiol. Ecol. 2010, 72, 445–455. [Google Scholar] [CrossRef]
- Luo, X.; Yang, Y.; Wang, L.; Zhang, Y.; Liao, H.; Peng, S.; Chen, W.; Huang, Q. Relations between bacterial communities and enzyme functions of two paddy soils. Eur. J. Soil Sci. 2018, 69, 655–665. [Google Scholar] [CrossRef]
- Nayak, D.R.; Babu, Y.J.; Adhya, T.K. Long-term application of compost influences microbial biomass and enzyme activities in a tropical Aeric Endoaquept planted to rice under flooded condition. Soil Biol. Biochem. 2007, 39, 1897–1906. [Google Scholar] [CrossRef]
- Mei, N.; Zhang, X.; Wang, X.; Peng, C.; Gao, H.; Zhu, P.; Gu, Y. Effects of 40 years applications of inorganic and organic fertilization on soil bacterial community in a maize agroecosystem in northeast China. Eur. J. Agron. 2021, 130, 126332. [Google Scholar] [CrossRef]
- Tang, Q.; Xia, Y.Q.; Ti, C.P.; Shan, J.; Zhou, W.; Li, C.L.; Yan, X.; Yan, X.Y. Partial organic fertilizer substitution promotes soil multifunctionality by increasing microbial community diversity and complexity. Pedosphere 2023, 33, 407–420. [Google Scholar] [CrossRef]
- Yan, T.; Xue, J.; Zhou, Z.; Wu, Y. Biochar-based fertilizer amendments improve the soil microbial community structure in a karst mountainous area. Sci. Total Environment. 2021, 794, 148757. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Liu, J.; Yu, Z.; Li, Y.; Jin, J.; Liu, X.; Wang, G. Changes of bacterial community compositions after three years of biochar application in a black soil of northeast China. Appl. Soil Ecol. 2017, 113, 11–21. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Zhang, Q.; Han, Y.; Chen, W.; Guo, Y.; Wu, M.; Wang, Y.; Hui, L. Soil type and pH mediated arable soil bacterial compositional variation across geographic distance in North China Plain. Appl. Soil Ecol. 2022, 169, 104220. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, X.; Liu, B.; Wan, J.; Jin, H.; Tao, K. Taiping HouDeciphering environmental factors driving soil microbial elevational distribution in the southeastern Qinghai-Tibetan Plateau. Eur. J. Soil Biol. 2022, 113, 103444. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Y.; Wang, S.; Sun, M.; Wang, M.; Ren, X.; Gao, Z.; Zhou, Y.; Zhang, J.; Zhuang, W.; et al. Temperature and phosphorus: The main environmental factors affecting the seasonal variation of soil bacterial diversity in in Nansi Lake Wetland. Front. Microbiol. 2023, 13, 147. [Google Scholar] [CrossRef]
- Feng, M.; Adams, J.M.; Fan, K.; Shi, Y.; Sun, R.; Wang, D.; Guo, X.; Chu, H. Long-term fertilization influences community assembly processes of soil diazotrophs. Soil Biol. Biochem. 2018, 126, 151–158. [Google Scholar] [CrossRef]
- Sradnick, A.; Murugan, R.; Oltmanns, M.; Raupp, J.; Joergensen, R.G. Changes in functional diversity of the soil microbial community in a heterogeneous sandy soil after long-term fertilization with cattle manure and mineral fertilizer. Appl. Soil Ecol. 2013, 63, 23–28. [Google Scholar] [CrossRef]
- Hu, J.; Lin, X.; Wang, J.; Dai, J.; Chen, R.; Zhang, J.; Wong, M.H. Microbial functional diversity, metabolic quotient, and invertase activity of a sandy loam soil as affected by long-term application of organic amendment and mineral fertilizer. J. Soils Sediments 2011, 11, 271–280. [Google Scholar] [CrossRef]
- Preece, C.; Verbruggen, E.; Liu, L.; Weedon, J.T.; Penuelas, J. Effects of past and current drought on the composition and diversity of soil microbial communities. Soil Biol. Biochem. 2019, 131, 28–39. [Google Scholar] [CrossRef]
- Wang, R.; Dorodnikov, M.; Yang, S.; Zhang, Y.; Filley, T.R.; Turco, R.F.; Zhang, Y.; Xu, Z.; Li, H.; Jiang, Y. Responses of enzymatic activities within soil aggregates to 9-year nitrogen and water addition in a semi-arid grassland. Soil Biol. Biochem. 2015, 81, 159–167. [Google Scholar] [CrossRef]
- Bastida, F.; Torres, I.F.; Hernandez, T.; Garcia, C. The impacts of organic amendments: Do they confer stability against drought on the soil microbial community? Soil Biol. Biochem. 2017, 113, 173–183. [Google Scholar] [CrossRef]
- Arnold, R.J.; Xie, Y.J.; Luo, J.Z.; Wang, H.R.; Midgley, S.J. A tale of two genera: Exotic Eucalyptus and Acacia species in China. 1. Domestication and Research. Int. For. Rev. 2020, 22, 1–18. [Google Scholar] [CrossRef]
- Booth, T.H. Eucalypt plantations and climate change. For. Ecol. Manag. 2013, 301, 28–34. [Google Scholar] [CrossRef]
- Madejon, P.; Alaejos, J.; Garcia-Albala, J.; Fernandez, M.; Madejon, E. Three-year study of fast-growing trees in degraded soils amended with composts: Effects on soil fertility and productivity. J. Environ. Manag. 2016, 169, 18–26. [Google Scholar] [CrossRef]
- Li, C.F.; Cao, C.G.; Wang, J.P.; Ming, Z.; Yuan, W.l.; Ahmad, S. Nitrogen losses from integrated rice-duck and rice-fish ecosystems in southern China. Plant Soil 2008, 307, 207–217. [Google Scholar]
- Reardon, C.L.; Klein, A.M.; Melle, C.J.; Hagerty, C.H.; Klarer, E.R.; Machado, S.; Paulitz, T.; Pritchett, L.; Schlatter, D.; Smith, S.F.; et al. Enzyme activities distinguish long-term fertilizer effects under different soil storage methods. Appl. Soil Ecol. 2022, 177, 104518. [Google Scholar] [CrossRef]
- Tuo, Y.; Wang, Z.; Zheng, Y.; Shi, X.; Liu, X.; Ding, M.; Yang, Q. Effect of water and fertilizer regulation on the soil microbial biomass carbon and nitrogen, enzyme activity, and saponin content of Panax notoginseng. Agric. Water Manag. 2023, 278, 108145. [Google Scholar] [CrossRef]
- Wei, Z.X.; Liang, Y.L.; Inoue, M.; Zhou, M.J.; Huang, M.L.; Gu, J.F.; Wu, Y. Effects of different water and fertilizer supply on cucumber soil nutrient content, enzyme activity, and microbial diversity. Chin. J. Appl. Ecol. 2009, 20, 1678–1684. [Google Scholar]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, J.; Mehandia, S.; Singh, G.; Raina, A.; Arya, S.K. Catalase Enzyme: Application in bioremediation and food industry. Biocatal. Agric. Biotechnol. 2018, 16, 192–199. [Google Scholar] [CrossRef]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol. Biochem. 2010, 42, 2182–2191. [Google Scholar] [CrossRef]
- Xun, W.; Xu, Z.; Li, W.; Ren, Y.; Huang, T.; Ran, W.; Wang, B.; Shen, Q.; Zhang, R. Long-term organic-inorganic fertilization ensures great soil productivity and bacterial diversity after natural-to-agricultural ecosystem conversion. J. Microbiol. 2016, 54, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Bastida, F.; Torres, I.F.; Romero-Trigueros, C.; Baldrian, P.; Vetrovsky, T.; Bayona, J.M.; Alarcon, J.J.; Hernandez, T.; Garcia, C.; Nicolas, E. Combined effects of reduced irrigation and water quality on the soil microbial community of a citrus orchard under semi-arid conditions. Soil Biol. Biochem. 2017, 104, 226–237. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Niu, W. Responses of bacterial community, root-soil interaction and tomato yield to different practices in subsurface drip irrigation. Sustainability 2020, 12, 2338. [Google Scholar] [CrossRef]
- Dangi, S.R.; Zhang, H.; Wang, D.; Gerik, J.; Hanson, B.D. Soil microbial community composition in a peach orchard under different irrigation methods and postharvest deficit irrigation. Soil Sci. 2016, 181, 208–215. [Google Scholar] [CrossRef]
- Mercier, A.; Dictor, M.C.; Harris-Hellal, J.; Breeze, D.; Mouvet, C. Distinct bacterial community structure of 3 tropical volcanic soils from banana plantations contaminated with chlordecone in Guadeloupe (French West Indies). Chemosphere 2013, 92, 787–794. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Li, J.H.; Friedman, C.R.; Wang, H.F. Variation of soil bacterial communities in a chronosequence of rubber tree (Hevea brasiliensis) plantations. Front. Plant Sci. 2017, 8, 849. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil Acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Wang, Y.; Ye, Y.; Zhao, Y.; Huang, Y.; Fu, W.; Chu, X. Effects of continuous nitrogen fertilizer application on the diversity and composition of Rhizosphere soil bacteria. Front. Microbiol. 2020, 11, 1948. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; He, Q.; Huang, D.; Wang, Z.; Mao, J.; Xie, X.; Su, Y.; Qiu, Q.; Li, J.; Chen, Z. Responses of fungal community structure and functional composition to short-term fertilization and dry season irrigation in Eucalyptus urophylla × Eucalyptus grandis plantation soils. Forests 2022, 13, 854. [Google Scholar] [CrossRef]
- Huber, K.J.; Vieira, S.; Sikorski, J.; Wuest, P.K.; Foesel, B.U.; Groengroeft, A.; Overmann, J. Differential response of Acidobacteria to water content, soil type, and land use during an extended drought in African savannah soils. Front. Microbiol. 2022, 13, 750456. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Brunner, I.; Hagedorn, F.; Bardgett, R.D.; Stierli, B.; Herzog, C.; Chen, X.; Zingg, A.; Graf-Pannatier, E.; Rigling, A.; et al. A decade of irrigation transforms the soil microbiome of a semi-arid pine forest. Mol. Ecol. 2017, 26, 1190–1206. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Su, W.; Chen, H.; Barberan, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Acuna, J.J.; Marileo, L.G.; Araya, M.A.; Rilling, J.I.; Larama, G.A.; Mora, M.L.; Epstein, S.; Jorquera, M.A. In Situ Cultivation approach to increase the culturable bacterial diversity in the rhizobiome of plants. J. Soil Sci. Plant Nutr. 2020, 20, 1411–1426. [Google Scholar] [CrossRef]
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 2019, 219, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, H.; Ferreira, R.; Gonzalez, J.M.; da Clara, M.I.; Santana, M.M. Influence of temperature and copper on Oxalobacteraceae in soil enrichments. Curr. Microbiol. 2015, 72, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Li, R.C.; Ren, C.Y.; Wu, L.K.; Zhang, X.X.; Mao, X.Y.; Fan, Z.; Cui, W.L.; Zhang, W.; Wei, G.H.; Shu, D.T. Fertilizing induced alterations of microbial functional profiles in soil nitrogen cycling closely associate with crop yield. Environ. Res. 2023, 231, 116194. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, Y.; Zhang, S.; Wei, W.; Kuzyakov, Y.; Ding, X. Fertilization effects on microbial community composition and aggregate formation in saline-alkaline Soil. Plant Soil 2021, 463, 523–535. [Google Scholar] [CrossRef]
- Wang, C.; Ma, H.; Feng, Z.; Yan, Z.; Song, B.; Wang, J.; Zheng, Y.; Hao, W.; Zhang, W.; Yao, M.; et al. Integrated organic and inorganic fertilization and reduced irrigation altered prokaryotic microbial community and diversity in different compartments of wheat root zone contributing to improved nitrogen uptake and wheat yield. Sci. Total Environ. 2022, 842, 156952. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Trivedi, P.; Osanai, Y.; Liu, Y.R.; Hamonts, K.; Jeffries, T.C.; Singh, B.K. Carbon content and climate variability drive global soil bacterial diversity patterns. Ecol. Monogr. 2016, 86, 373–390. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil bacterial community composition altered by increased nutrient availability in Arctic tundra soils. Front. Microbiol. 2014, 5, 516. [Google Scholar] [CrossRef]
- Fierer, N.; Ladau, J.; Clemente, J.C.; Leff, J.W.; Owens, S.M.; Pollard, K.S.; Knight, R.; Gilbert, J.A.; McCulley, R.L. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the United States. Science 2013, 342, 621–624. [Google Scholar] [CrossRef]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Hu, C.; Yuan, Z. Soil metagenomics reveals effects of continuous sugarcane cropping on the structure and functional pathway of rhizospheric microbial community. Front. Microbiol. 2021, 12, 627569. [Google Scholar] [CrossRef]
- Yu, F.; Thuy Van, T.; He, Q.; Hua, L.; Su, Y.; Li, J. Dry season irrigation promotes leaf growth in Eucalyptus Urophylla × E. Grandis Under Fertilization. Forests 2019, 10, 67. [Google Scholar]
- Hua, L.; Yu, F.; Qiu, Q.; He, Q.; Su, Y.; Liu, X.; Li, J. Relationships between diurnal and seasonal variation of photosynthetic characteristics of Eucalyptus plantation and environmental factors under dry-season irrigation with fertilization. Agric. Water Manag. 2021, 248, 106737. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis. Part 2: Microbiological and Biochemical Properties; Weaver, R.W., Angel, J.S., Bottomley, P.S., Eds.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 775–833. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry. J. Appl. Ecol. 1995, 33, 178. [Google Scholar]
- Jiang, J.P.; Xiong, Y.C.; Jiang, H.M.; Ye, D.Y.; Song, Y.J.; Li, F.M. Soil microbial activity during secondary vegetation succession. Pedosphere 2009, 19, 735–747. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Nanniperi, P.; Gelsomino, A.; Felici, M. Method to determine guaiacol oxidase activity in Soil. Soil Sci. Soc. Am. J. 1991, 55, 1347–1352. [Google Scholar] [CrossRef]
- Finzi, A.C.; Berthrong, S.T. The uptake of amino acids by microbes and trees in three cold-temperate Forests. Ecology 2005, 86, 3345–3353. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2010; pp. 39–114. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Ji, L.; Tanunchai, B.; Wahdan, S.F.M.; Schaedler, M.; Purahong, W. Future climate change enhances the complexity of plastisphere microbial co-occurrence networks, but does not significantly affect the community assembly. Sci. Total Environ. 2022, 844, 157016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | CK | F | W | WF |
|---|---|---|---|---|
| Urease (μg g−1 h−1) | 13.32 ± 1.12 c | 18.72 ± 2.92 b | 14.33 ± 1.31 c | 25.20 ± 2.38 a |
| Acid Phosphatase (μmol g−1 h−1) | 5.77 ± 0.50 a | 4.14 ± 0.65 b | 5.83 ± 0.46 a | 5.20 ± 0.19 a |
| Cellulase (μmol g−1 h−1) | 0.07 ± 0.01 b | 0.09 ± 0.01 a | 0.08 ± 0.01 b | 0.09 ± 0.01 a |
| Chitinase (μmol g−1 h−1) | 0.09 ± 0.01 c | 0.11 ± 0.01 b | 0.11 ± 0.01 b | 0.20 ± 0.02 a |
| Invertase (mg g−1 d−1) | 4.88 ± 0.93 a | 3.31 ± 0.30 b | 5.39 ± 1.57 a | 4.79 ± 0.38 a |
| Catalase (mg g−1 20 min−1) | 0.70 ± 0.04 b | 0.69 ± 0.04 b | 0.72 ± 0.02 ab | 0.76 ± 0.04 a |
| Phenoloxidase (μmol kg−1 h−1) | 4.07 ± 1.45 ab | 4.43 ± 0.87 ab | 5.41 ± 1.22 a | 3.63 ± 1.06 b |
| Peroxidase (μmol kg−1 h−1) | 7.55 ± 1.00 a | 4.53 ± 0.80 b | 5.94 ± 1.01 b | 7.44 ± 1.41 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, C.; Mao, J.; Zhang, J.; Yang, X.; Gao, S.; Li, J.; He, Q.; Tang, G.; Xie, X.; Chen, Z. Fertilization- and Irrigation-Modified Bacterial Community Composition and Stimulated Enzyme Activity of Eucalyptus Plantations Soil. Int. J. Mol. Sci. 2024, 25, 1385. https://doi.org/10.3390/ijms25031385
Huo C, Mao J, Zhang J, Yang X, Gao S, Li J, He Q, Tang G, Xie X, Chen Z. Fertilization- and Irrigation-Modified Bacterial Community Composition and Stimulated Enzyme Activity of Eucalyptus Plantations Soil. International Journal of Molecular Sciences. 2024; 25(3):1385. https://doi.org/10.3390/ijms25031385
Chicago/Turabian StyleHuo, Chunyu, Jianhui Mao, Jianlang Zhang, Xinzhu Yang, Shangkun Gao, Jiyue Li, Qian He, Guangda Tang, Xianan Xie, and Zujing Chen. 2024. "Fertilization- and Irrigation-Modified Bacterial Community Composition and Stimulated Enzyme Activity of Eucalyptus Plantations Soil" International Journal of Molecular Sciences 25, no. 3: 1385. https://doi.org/10.3390/ijms25031385
APA StyleHuo, C., Mao, J., Zhang, J., Yang, X., Gao, S., Li, J., He, Q., Tang, G., Xie, X., & Chen, Z. (2024). Fertilization- and Irrigation-Modified Bacterial Community Composition and Stimulated Enzyme Activity of Eucalyptus Plantations Soil. International Journal of Molecular Sciences, 25(3), 1385. https://doi.org/10.3390/ijms25031385