

Canonical and Non-Canonical Localization of Tight Junction Proteins during Early Murine Cranial Development


Abstract
:1. Introduction
2. Results
2.1. Expression Profile of Tight-Junction-Related Proteins in Embryonic Mouse Heads
2.2. Localization of the Zona Occludens-1 (ZO-1) Protein during Mouse Cranial Neurulation
2.3. Claudin-4 Is Specific to the Non-Neuronal Ectoderm in Cranial Mouse Neural Folds
2.4. Non-Junctional Localization of Claudin-3 in the Early Developing Neural Tube
2.5. Claudin-1 and Occludin Are Localized to the Neuroepithelium and to the Non-Neuronal Ectoderm
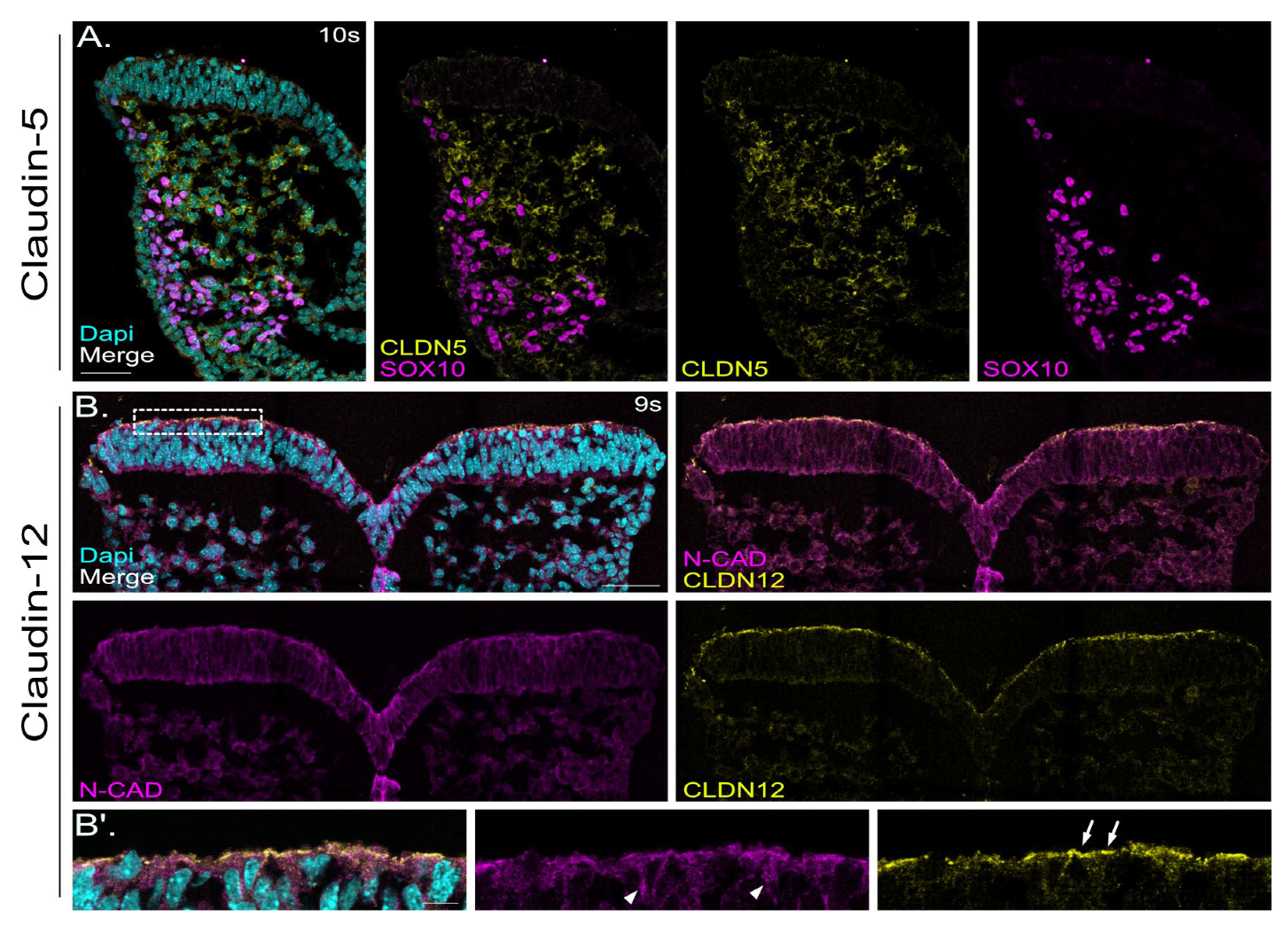
2.6. Localization of Claudin-12 Exclusively in Neuronal Tissue and Localization of Claudin-5 Only in SOX10-Negative Mesenchymal Cells
3. Discussion
3.1. Expression Profile of Genes Encoding Tight Junction Complex Proteins
3.2. Tight-Junction-Associated Proteins in the Neural Tube
3.3. Occludin in the Cranial Neural Tube
3.4. Claudins in the Cranial Neural Tube
3.4.1. Non-Neuronal Ectoderm-Specific Claudin
3.4.2. Non-Canonical Localization of Claudins in the Neural Tube
3.4.3. Mesoderm-Specific Claudins in the Cranial Neural Tube
3.4.4. Classical Tight Junction Localization of Claudins and Occludin in the Non-Neuronal and Neuronal Ectoderm
3.4.5. Neuronal Ectoderm-Specific Claudins in the Cranial Neural Tube
4. Materials and Methods
4.1. Animal Work and Embryo Collection
4.2. Embryo Fixation and Cryosectioning
4.3. RNA Library Generation and Sequencing
4.4. Antibodies
4.5. Immunofluorescence
4.5.1. Sections
4.5.2. Whole-Mount
4.6. Microscopy and Image Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colas, J.-F.; Schoenwolf, G.C. Towards a Cellular and Molecular Understanding of Neurulation. Dev. Dyn. 2001, 221, 117–145. [Google Scholar] [CrossRef] [PubMed]
- Lowery, L.A.; Sive, H. Strategies of Vertebrate Neurulation and a Re-Evaluation of Teleost Neural Tube Formation. Mech. Dev. 2004, 121, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Nikolopoulou, E.; Galea, G.L.; Rolo, A.; Greene, N.D.E.; Copp, A.J. Neural Tube Closure: Cellular, Molecular and Biomechanical Mechanisms. Development 2017, 144, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Copp, A.J.; Greene, N.D.E.; Murdoch, J.N. The Genetic Basis of Mammalian Neurulation. Nat. Rev. Genet. 2003, 4, 784–793. [Google Scholar] [CrossRef]
- Greene, N.D.E.; Copp, A.J. Development of the Vertebrate Central Nervous System: Formation of the Neural Tube. Prenat. Diagn. 2009, 29, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.D.E.; Copp, A.J. Neural Tube Defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.J.; Juriloff, D.M. Mouse Mutants with Neural Tube Closure Defects and Their Role in Understanding Human Neural Tube Defects. Birth Defects Res. Part. A Clin. Mol. Teratol. 2007, 79, 187–210. [Google Scholar] [CrossRef] [PubMed]
- Szabó, A.; Mayor, R. Mechanisms of Neural Crest Migration. Annu. Rev. Genet. 2018, 52, 43–63. [Google Scholar] [CrossRef]
- Martik, M.L.; Bronner, M.E. Riding the Crest to Get a Head: Neural Crest Evolution in Vertebrates. Nat. Rev. Neurosci. 2021, 22, 616–626. [Google Scholar] [CrossRef]
- Stundl, J.; Bertucci, P.Y.; Lauri, A.; Arendt, D.; Bronner, M.E. Evolution of New Cell Types at the Lateral Neural Border. Curr. Top. Dev. Biol. 2021, 141, 173–205. [Google Scholar] [CrossRef]
- Koontz, A.; Urrutia, H.A.; Bronner, M.E. Making a Head: Neural Crest and Ectodermal Placodes in Cranial Sensory Development. Semin. Cell Dev. Biol. 2023, 138, 15–27. [Google Scholar] [CrossRef]
- Vega-Lopez, G.A.; Cerrizuela, S.; Tribulo, C.; Aybar, M.J. Neurocristopathies: New Insights 150 Years after the Neural Crest Discovery. Dev. Biol. 2018, 444, S110–S143. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, M.G.; Palade, G.E. Junctional Complexes in Various Epithelia. J. Cell Biol. 1963, 17, 375–412. [Google Scholar] [CrossRef] [PubMed]
- Claude, P.; Goodenough, D.A. Fracture Faces of Zonulae Occludentes from “Tight” and “Leaky” Epithelia. J. Cell Biol. 1973, 58, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Rouaud, F.; Sluysmans, S.; Flinois, A.; Shah, J.; Vasileva, E.; Citi, S. Scaffolding Proteins of Vertebrate Apical Junctions: Structure, Functions and Biophysics. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183399. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Tanaka, H.; Tamura, A. The Claudins: From Tight Junctions to Biological Systems. Trends Biochem. Sci. 2019, 44, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin Multigene Family Encoding Four-Transmembrane Domain Protein Components of Tight Junction Strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D.; Yu, A.S.L. Claudins and the Modulation of Tight Junction Permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel Integral Membrane Proteins Localizing at Tight Junctions with No Sequence Similarity to Occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Occludin: A Novel Integral Membrane Protein Localizing at Tight Junctions. J. Cell Biol. 1993, 123, 1777–1788. [Google Scholar] [CrossRef]
- Ando-Akatsuka, Y.; Saitou, M.; Hirase, T.; Kishi, M.; Sakakibara, A.; Itoh, M.; Yonemura, S.; Furuse, M.; Tsukita, S. Interspecies Diversity of the Occludin Sequence: cDNA Cloning of Human, Mouse, Dog, and Rat-Kangaroo Homologues. J. Cell Biol. 1996, 133, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Schwietzer, Y.A.; Otani, T.; Furuse, M.; Ebnet, K. Physiological Functions of Junctional Adhesion Molecules (JAMs) in Tight Junctions. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183299. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight Junction Proteins Occludin and ZO-1 as Regulators of Epithelial Proliferation and Survival. Ann. N. Y Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef] [PubMed]
- McNeil, E.; Capaldo, C.T.; Macara, I.G. Zonula Occludens-1 Function in the Assembly of Tight Junctions in Madin-Darby Canine Kidney Epithelial Cells. Mol. Biol. Cell 2006, 17, 1922–1932. [Google Scholar] [CrossRef]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A High Molecular Weight Polypeptide Associated with the Tight Junction (Zonula Occludens) in a Variety of Epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Nguyen, T.P.; Tokuda, S.; Sugihara, K.; Sugawara, T.; Furuse, K.; Miura, T.; Ebnet, K.; Furuse, M. Claudins and JAM-A Coordinately Regulate Tight Junction Formation and Epithelial Polarity. J. Cell Biol. 2019, 218, 3372–3396. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Itallie, C.M.V. Physiology and Function of the Tight Junction. Cold Spring Harb. Perspect. Biol. 2009, 1, a002584. [Google Scholar] [CrossRef]
- Powell, D.W. Barrier Function of Epithelia. Am. J. Physiol. 1981, 241, G275–G288. [Google Scholar] [CrossRef]
- Diamond, J.M. Channels in Epithelial Cell Membranes and Junctions. Fed. Proc. 1978, 37, 2639–2643. [Google Scholar]
- Huber, J.D.; Egleton, R.D.; Davis, T.P. Molecular Physiology and Pathophysiology of Tight Junctions in the Blood–Brain Barrier. Trends Neurosci. 2001, 24, 719–725. [Google Scholar] [CrossRef]
- Balda, M.S.; Matter, K. Tight Junctions. J. Cell Sci. 1998, 111 Pt 5, 541–547. [Google Scholar] [CrossRef]
- Madara, J.L. Regulation of the Movement of Solutes across Tight Junctions. Annu. Rev. Physiol. 1998, 60, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.B.N.; Huang, E.; Ward, H.J. Tight Junction Biology and Kidney Dysfunction. Am. J. Physiol. Ren. Physiol. 2006, 290, F20–F34. [Google Scholar] [CrossRef] [PubMed]
- Bleich, M.; Günzel, D. Physiology, Pathophysiology, and Clinical Impact of Claudins. Pflug. Arch Eur. J. Physiol. 2017, 469, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Bleich, M.; Shan, Q.; Himmerkus, N. Calcium Regulation of Tight Junction Permeability. Ann. N. Y Acad. Sci. 2012, 1258, 93–99. [Google Scholar] [CrossRef]
- Breiderhoff, T.; Himmerkus, N.; Meoli, L.; Fromm, A.; Sewerin, S.; Kriuchkova, N.; Nagel, O.; Ladilov, Y.; Krug, S.M.; Quintanova, C.; et al. Claudin-10a Deficiency Shifts Proximal Tubular Cl- Permeability to Cation Selectivity via Claudin-2 Redistribution. J. Am. Soc. Nephrol. 2022, 33, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Fromm, M.; Piontek, J.; Rosenthal, R.; Günzel, D.; Krug, S.M. Tight Junctions of the Proximal Tubule and Their Channel Proteins. Pflug. Arch. 2017, 469, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.A. The Role of Tight Junctions in Cancer Metastasis. Semin. Cell Dev. Biol. 2014, 36, 224–231. [Google Scholar] [CrossRef]
- Krug, S.M.; Schulzke, J.D.; Fromm, M. Tight Junction, Selective Permeability, and Related Diseases. Semin. Cell Dev. Biol. 2014, 36, 166–176. [Google Scholar] [CrossRef]
- Parikh, K.; Antanaviciute, A.; Fawkner-Corbett, D.; Jagielowicz, M.; Aulicino, A.; Lagerholm, C.; Davis, S.; Kinchen, J.; Chen, H.H.; Alham, N.K.; et al. Colonic Epithelial Cell Diversity in Health and Inflammatory Bowel Disease. Nature 2019, 567, 49–55. [Google Scholar] [CrossRef]
- Horowitz, A.; Chanez-Paredes, S.D.; Haest, X.; Turner, J.R. Paracellular Permeability and Tight Junction Regulation in Gut Health and Disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 417–432. [Google Scholar] [CrossRef]
- Gupta, I.; Ryan, A. Claudins: Unlocking the Code to Tight Junction Function during Embryogenesis and in Disease. Clin. Genet. 2010, 77, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The Blood-Brain Barrier in Health and Chronic Neurodegenerative Disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Syed, N.; Therachiyil, L.; Nisar, S.; Hashem, S.; Macha, M.A.; Yadav, S.K.; Krishnankutty, R.; Muralitharan, S.; Al-Naemi, H.; et al. Claudin-1, A Double-Edged Sword in Cancer. Int. J. Mol. Sci. 2020, 21, 569. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Baumholtz, A.I.; Gupta, I.R.; Ryan, A.K. Claudins in Morphogenesis: Forming an Epithelial Tube. Tissue Barriers 2017, 5, e1361899. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Moriwaki, K. The Role of Claudin-Based Tight Junctions in Morphogenesis. Ann. N. Y. Acad. Sci. 2009, 1165, 58–61. [Google Scholar] [CrossRef]
- Du, D.; Xu, F.; Yu, L.; Zhang, C.; Lu, X.; Yuan, H.; Huang, Q.; Zhang, F.; Bao, H.; Jia, L.; et al. The Tight Junction Protein, Occludin, Regulates the Directional Migration of Epithelial Cells. Dev. Cell 2010, 18, 52–63. [Google Scholar] [CrossRef]
- Balda, M.S.; Matter, K. Tight Junctions as Regulators of Tissue Remodelling. Curr. Opin. Cell Biol. 2016, 42, 94–101. [Google Scholar] [CrossRef]
- Baumholtz, A.I.; Simard, A.; Nikolopoulou, E.; Oosenbrug, M.; Collins, M.M.; Piontek, A.; Krause, G.; Piontek, J.; Greene, N.D.E.; Ryan, A.K. Claudins Are Essential for Cell Shape Changes and Convergent Extension Movements during Neural Tube Closure. Dev. Biol. 2017, 428, 25–38. [Google Scholar] [CrossRef]
- Friedl, P.; Mayor, R. Tuning Collective Cell Migration by Cell-Cell Junction Regulation. Cold Spring Harb. Perspect. Biol. 2017, 9, a029199. [Google Scholar] [CrossRef]
- Matter, K.; Balda, M.S. Signalling to and from Tight Junctions. Nat. Rev. Mol. Cell Biol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight Junctions: From Simple Barriers to Multifunctional Molecular Gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Baumholtz, A.I.; De Marco, P.; Capra, V.; Ryan, A.K. Functional Validation of CLDN Variants Identified in a Neural Tube Defect Cohort Demonstrates Their Contribution to Neural Tube Defects. Front. Neurosci. 2020, 14, 664. [Google Scholar] [CrossRef] [PubMed]
- Gamero-Estevez, E.; Baumholtz, A.I.; Ryan, A.K. Developing a Link between Toxicants, Claudins and Neural Tube Defects. Reprod. Toxicol. 2018, 81, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Mecklenburg, N.; Kowalczyk, I.; Witte, F.; Görne, J.; Laier, A.; Mamo, T.M.; Gonschior, H.; Lehmann, M.; Richter, M.; Sporbert, A.; et al. Identification of Disease-Relevant Modulators of the SHH Pathway in the Developing Brain. Development 2021, 148, dev199307. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and Function of Claudins. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Mineta, K.; Yamamoto, Y.; Yamazaki, Y.; Tanaka, H.; Tada, Y.; Saito, K.; Tamura, A.; Igarashi, M.; Endo, T.; Takeuchi, K.; et al. Predicted Expansion of the Claudin Multigene Family. FEBS Lett. 2011, 585, 606–612. [Google Scholar] [CrossRef]
- Hagen, S.J. Non-Canonical Functions of Claudin Proteins: Beyond the Regulation of Cell-Cell Adhesions. Tissue Barriers 2017, 5, e1327839. [Google Scholar] [CrossRef]
- Gumbiner, B.; Lowenkopf, T.; Apatira, D. Identification of a 160-kDa Polypeptide That Binds to the Tight Junction Protein ZO-1. Proc. Natl. Acad. Sci. USA 1991, 88, 3460–3464. [Google Scholar] [CrossRef]
- Balda, M.S.; Gonzalez-Mariscal, L.; Matter, K.; Cereijido, M.; Anderson, J.M. Assembly of the Tight Junction: The Role of Diacylglycerol. J. Cell Biol. 1993, 123, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.C.; Kaschube, M.; Wieschaus, E.F. Pulsed Contractions of an Actin–Myosin Network Drive Apical Constriction. Nature 2009, 457, 495–499. [Google Scholar] [CrossRef]
- Martin, A.C.; Goldstein, B. Apical Constriction: Themes and Variations on a Cellular Mechanism Driving Morphogenesis. Development 2014, 141, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, N.; Skourides, P.A. Cell-Autonomous Ca2+ Flashes Elicit Pulsed Contractions of an Apical Actin Network to Drive Apical Constriction during Neural Tube Closure. Cell Rep. 2015, 13, 2189–2202. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara-Tani, R.; Mori, S.; Ogata, R.; Sasaki, R.; Ikemoto, A.; Kishi, S.; Kondoh, M.; Kuniyasu, H. Claudin-4: A New Molecular Target for Epithelial Cancer Therapy. Int. J. Mol. Sci. 2023, 24, 5494. [Google Scholar] [CrossRef] [PubMed]
- Lameris, A.L.L.; Huybers, S.; Burke, J.R.; Monnens, L.A.; Bindels, R.J.M.; Hoenderop, J.G.J. Involvement of Claudin 3 and Claudin 4 in Idiopathic Infantile Hypercalcaemia: A Novel Hypothesis? Nephrol. Dial. Transpl. 2010, 25, 3504–3509. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; Kopplin, K.; Blasig, R.; Stuiver, M.; Koning, N.; Goverse, G.; van der Pol, S.M.A.; van het Hof, B.; Gollasch, M.; Drexhage, J.A.R.; et al. Disturbed Function of the Blood–Cerebrospinal Fluid Barrier Aggravates Neuro-Inflammation. Acta Neuropathol. 2014, 128, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-Based Tight Junctions Are Crucial for the Mammalian Epidermal Barrier: A Lesson from Claudin-1-Deficient Mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef]
- Feldman, G.J.; Mullin, J.M.; Ryan, M.P. Occludin: Structure, Function and Regulation. Adv. Drug Deliv. Rev. 2005, 57, 883–917. [Google Scholar] [CrossRef]
- Pusch, C.; Hustert, E.; Pfeifer, D.; Südbeck, P.; Kist, R.; Roe, B.; Wang, Z.; Balling, R.; Blin, N.; Scherer, G. The SOX10/Sox10 Gene from Human and Mouse: Sequence, Expression, and Transactivation by the Encoded HMG Domain Transcription Factor. Hum. Genet. 1998, 103, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.T.-C.; Hu, X.; Otto, I.U.C.; Schürger, C.; von Bieberstein, B.R.; Doppler, K.; Krug, S.M.; Hankir, M.K.; Blasig, R.; Sommer, C.; et al. Myelin Barrier Breakdown, Mechanical Hypersensitivity, and Painfulness in Polyneuropathy with Claudin-12 Deficiency. Neurobiol. Dis. 2023, 185, 106246. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Schoenwolf, G.C. Further Evidence of Extrinsic Forces in Bending of the Neural Plate. J. Comp. Neurol. 1991, 307, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Aaku-Saraste, E.; Hellwig, A.; Huttner, W.B. Loss of Occludin and Functional Tight Junctions, but Not ZO-1, during Neural Tube Closure—Remodeling of the Neuroepithelium Prior to Neurogenesis. Dev. Biol. 1996, 180, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Fishwick, K.J.; Neiderer, T.; Jhingory, S.; Bronner, M.; Taneyhill, L.A. The Tight Junction Protein Claudin-1 Influences Cranial Neural Crest Cell Emigration. Mech. Dev. 2012, 129, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Grego-Bessa, J.; Hildebrand, J.; Anderson, K.V. Morphogenesis of the Mouse Neural Plate Depends on Distinct Roles of Cofilin 1 in Apical and Basal Epithelial Domains. Development 2015, 142, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Vacca, B.; Sanchez-Heras, E.; Steed, E.; Busson, S.L.; Balda, M.S.; Ohnuma, S.-I.; Sasai, N.; Mayor, R.; Matter, K. Control of Neural Crest Induction by MarvelD3-Mediated Attenuation of JNK Signalling. Sci. Rep. 2018, 8, 1204. [Google Scholar] [CrossRef]
- Legere, E.-A.; Baumholtz, A.I.; Lachance, J.-F.B.; Archer, M.; Piontek, J.; Ryan, A.K. Claudin-3 in the Non-Neural Ectoderm Is Essential for Neural Fold Fusion in Chicken Embryos. Dev. Biol. 2024, 507, 20–33. [Google Scholar] [CrossRef]
- Mackensen, A.; Haanen, J.B.A.G.; Koenecke, C.; Alsdorf, W.; Wagner-Drouet, E.; Borchmann, P.; Heudobler, D.; Ferstl, B.; Klobuch, S.; Bokemeyer, C.; et al. CLDN6-Specific CAR-T Cells plus Amplifying RNA Vaccine in Relapsed or Refractory Solid Tumors: The Phase 1 BNT211-01 Trial. Nat. Med. 2023, 29, 2844–2853. [Google Scholar] [CrossRef]
- Troy, T.-C.; Arabzadeh, A.; Yerlikaya, S.; Turksen, K. Claudin Immunolocalization in Neonatal Mouse Epithelial Tissues. Cell Tissue Res. 2007, 330, 381–388. [Google Scholar] [CrossRef]
- Moriwaki, K.; Tsukita, S.; Furuse, M. Tight Junctions Containing Claudin 4 and 6 Are Essential for Blastocyst Formation in Preimplantation Mouse Embryos. Dev. Biol. 2007, 312, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Ichikawa-Tomikawa, N.; Satohisa, S.; Akashi, Y.; Kanai, R.; Saito, T.; Sawada, N.; Chiba, H. The Tight-Junction Protein Claudin-6 Induces Epithelial Differentiation from Mouse F9 and Embryonic Stem Cells. PLoS ONE 2013, 8, e75106. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, F.R.; Lewis, J.B.; Belgique, S.T.; Wood, T.T.; Reynolds, P.R. Developmental Lung Expression and Transcriptional Regulation of Claudin-6 by TTF-1, Gata-6, and FoxA2. Respir. Res. 2014, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Episkopou, V. SOX2 Functions in Adult Neural Stem Cells. Trends Neurosci. 2005, 28, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Nudel, N.; Benvenisty, N. Immunologic and Chemical Targeting of the Tight-Junction Protein Claudin-6 Eliminates Tumorigenic Human Pluripotent Stem Cells. Nat. Commun. 2013, 4, 1992. [Google Scholar] [CrossRef] [PubMed]
- Bronstein, J.M.; Tiwari-Woodruff, S.; Buznikov, A.G.; Stevens, D.B. Involvement of OSP/Claudin-11 in Oligodendrocyte Membrane Interactions: Role in Biology and Disease. J. Neurosci. Res. 2000, 59, 706–711. [Google Scholar] [CrossRef]
- Gow, A.; Southwood, C.M.; Li, J.S.; Pariali, M.; Riordan, G.P.; Brodie, S.E.; Danias, J.; Bronstein, J.M.; Kachar, B.; Lazzarini, R.A. CNS Myelin and Sertoli Cell Tight Junction Strands Are Absent in Osp/Claudin-11 Null Mice. Cell 1999, 99, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Tiwari-Woodruff, S.K.; Kaplan, R.; Kornblum, H.I.; Bronstein, J.M. Developmental Expression of OAP-1/Tspan-3, a Member of the Tetraspanin Superfamily. J. Neurosci. Res. 2004, 77, 166–173. [Google Scholar] [CrossRef]
- Yan, H.; Kanki, H.; Matsumura, S.; Kawano, T.; Nishiyama, K.; Sugiyama, S.; Takemori, H.; Mochizuki, H.; Sasaki, T. MiRNA-132/212 Regulates Tight Junction Stabilization in Blood-Brain Barrier after Stroke. Cell Death Discov. 2021, 7, 380. [Google Scholar] [CrossRef]
- Burek, M.; König, A.; Lang, M.; Fiedler, J.; Oerter, S.; Roewer, N.; Bohnert, M.; Thal, S.C.; Blecharz-Lang, K.G.; Woitzik, J.; et al. Hypoxia-Induced MicroRNA-212/132 Alter Blood-Brain Barrier Integrity through Inhibition of Tight Junction-Associated Proteins in Human and Mouse Brain Microvascular Endothelial Cells. Transl. Stroke Res. 2019, 10, 672–683. [Google Scholar] [CrossRef]
- Wyss, L.; Schäfer, J.; Liebner, S.; Mittelbronn, M.; Deutsch, U.; Enzmann, G.; Adams, R.H.; Aurrand-Lions, M.; Plate, K.H.; Imhof, B.A.; et al. Junctional Adhesion Molecule (JAM)-C Deficient C57BL/6 Mice Develop a Severe Hydrocephalus. PLoS ONE 2012, 7, e45619. [Google Scholar] [CrossRef] [PubMed]
- Katsuno, T.; Umeda, K.; Matsui, T.; Hata, M.; Tamura, A.; Itoh, M.; Takeuchi, K.; Fujimori, T.; Nabeshima, Y.; Noda, T.; et al. Deficiency of Zonula Occludens-1 Causes Embryonic Lethal Phenotype Associated with Defected Yolk Sac Angiogenesis and Apoptosis of Embryonic Cells. Mol. Biol. Cell 2008, 19, 2465–2475. [Google Scholar] [CrossRef] [PubMed]
- Simard, A.; Di Pietro, E.; Ryan, A.K. Gene Expression Pattern of Claudin-1 during Chick Embryogenesis. Gene Expr. Patterns 2005, 5, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Bendriem, R.M.; Singh, S.; Aleem, A.A.; Antonetti, D.A.; Ross, M.E. Tight Junction Protein Occludin Regulates Progenitor Self-Renewal and Survival in Developing Cortex. Elife 2019, 8, e49376. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, I.; Lickert, H. Foxa2 Regulates Polarity and Epithelialization in the Endoderm Germ Layer of the Mouse Embryo. Development 2009, 136, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J. Claudin Proteins in Human Cancer: Promising New Targets for Diagnosis and Therapy. Cancer Res. 2005, 65, 9603–9606. [Google Scholar] [CrossRef] [PubMed]
- Neesse, A.; Griesmann, H.; Gress, T.M.; Michl, P. Claudin-4 as Therapeutic Target in Cancer. Arch. Biochem. Biophys. 2012, 524, 64–70. [Google Scholar] [CrossRef]
- Francke, U. Williams-Beuren Syndrome: Genes and Mechanisms. Hum. Mol. Genet. 1999, 8, 1947–1954. [Google Scholar] [CrossRef]
- Werth, M.; Walentin, K.; Aue, A.; Schönheit, J.; Wuebken, A.; Pode-Shakked, N.; Vilianovitch, L.; Erdmann, B.; Dekel, B.; Bader, M.; et al. The Transcription Factor Grainyhead-like 2 Regulates the Molecular Composition of the Epithelial Apical Junctional Complex. Development 2010, 137, 3835–3845. [Google Scholar] [CrossRef]
- Todd, M.C.; Petty, H.M.; King, J.M.; Piana Marshall, B.N.; Sheller, R.A.; Cuevas, M.E. Overexpression and Delocalization of Claudin-3 Protein in MCF-7 and MDA-MB-415 Breast Cancer Cell Lines. Oncol. Lett. 2015, 10, 156–162. [Google Scholar] [CrossRef]
- Tokuhara, Y.; Morinishi, T.; Matsunaga, T.; Sakai, M.; Sakai, T.; Ohsaki, H.; Kadota, K.; Kushida, Y.; Haba, R.; Hirakawa, E. Nuclear Expression of Claudin-3 in Human Colorectal Adenocarcinoma Cell Lines and Tissues. Oncol. Lett. 2018, 15, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.-R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 Regulates Cellular Transformation and Metastatic Behavior in Colon Cancer. J. Clin. Investig. 2005, 115, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Schilpp, C.; Lochbaum, R.; Braubach, P.; Jonigk, D.; Frick, M.; Dietl, P.; Wittekindt, O.H. TGF-Β1 Increases Permeability of Ciliated Airway Epithelia via Redistribution of Claudin 3 from Tight Junction into Cell Nuclei. Pflug. Arch. 2021, 473, 287–311. [Google Scholar] [CrossRef]
- Balda, M.S.; Matter, K. Tight Junctions and the Regulation of Gene Expression. Biochim. Biophys. Acta 2009, 1788, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Sourisseau, T.; Georgiadis, A.; Tsapara, A.; Ali, R.R.; Pestell, R.; Matter, K.; Balda, M.S. Regulation of PCNA and Cyclin D1 Expression and Epithelial Morphogenesis by the ZO-1-Regulated Transcription Factor ZONAB/DbpA. Mol. Cell Biol. 2006, 26, 2387–2398. [Google Scholar] [CrossRef]
- Spadaro, D.; Tapia, R.; Pulimeno, P.; Citi, S. The Control of Gene Expression and Cell Proliferation by the Epithelial Apical Junctional Complex. Essays Biochem. 2012, 53, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, T.; Misteli, T. The Lamin Protein Family. Genome Biol. 2011, 12, 222. [Google Scholar] [CrossRef]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of Neurological Function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-Selective Loosening of the Blood-Brain Barrier in Claudin-5-Deficient Mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Hewitt, K.J.; Agarwal, R.; Morin, P.J. The Claudin Gene Family: Expression in Normal and Neoplastic Tissues. BMC Cancer 2006, 6, 186. [Google Scholar] [CrossRef]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial Claudin: Claudin-5/Tmvcf Constitutes Tight Junction Strands in Endothelial Cells. J. Cell Biol. 1999, 147, 185–194. [Google Scholar] [CrossRef]
- Ben-Yosef, T.; Belyantseva, I.A.; Saunders, T.L.; Hughes, E.D.; Kawamoto, K.; Van Itallie, C.M.; Beyer, L.A.; Halsey, K.; Gardner, D.J.; Wilcox, E.R.; et al. Claudin 14 Knockout Mice, a Model for Autosomal Recessive Deafness DFNB29, Are Deaf Due to Cochlear Hair Cell Degeneration. Hum. Mol. Genet. 2003, 12, 2049–2061. [Google Scholar] [CrossRef]
- Anderson, W.J.; Zhou, Q.; Alcalde, V.; Kaneko, O.F.; Blank, L.J.; Sherwood, R.I.; Guseh, J.S.; Rajagopal, J.; Melton, D.A. Genetic Targeting of the Endoderm with Claudin-6CreER. Dev. Dyn. 2008, 237, 504–512. [Google Scholar] [CrossRef]
- Fujita, H.; Hamazaki, Y.; Noda, Y.; Oshima, M.; Minato, N. Claudin-4 Deficiency Results in Urothelial Hyperplasia and Lethal Hydronephrosis. PLoS ONE 2012, 7, e52272. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Weber, C.R.; Turner, J.R. The Tight Junction Protein Complex Undergoes Rapid and Continuous Molecular Remodeling at Steady State. J. Cell Biol. 2008, 181, 683–695. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Manufacturer, Catalog No. | Dilution |
|---|---|---|
| Mouse Claudin-1 | Santa Cruz Biotechnology, Heidelberg, Germany, sc166338 | 1:100 |
| Mouse E-cadherin | Abcam, Cambridge, UK, ab76055 | 1:100 |
| Mouse Lamin B1 | Proteintech, Manchester, UK, 66095-1-Ig | 1:200 |
| Mouse ZO-1 | Invitrogen GmbH, Karlsruhe, Germany, 33-9100 | 1:100 |
| Rabbit Claudin-3 | Invitrogen, 34-1700 | 1:100 |
| Rabbit Claudin-4 | Invitrogen, 36-4800 | 1:100 |
| Rabbit Claudin-5 | Invitrogen, 1548773A | 1:100 |
| Rabbit Claudin-12 | IBL International, Hamburg, Germany, 18801 | 1:100 |
| Rabbit ITGA4 | Cusabio, Houston TX, USA, CSB-PA011867LA01HU | 1:200 |
| Rabbit Occludin | Proteintech, 13409-1-AP | 1:200 |
| Goat N-cadherin | Santa Cruz, sc31030 | 1:100 |
| Goat SOX10 | R&D Systems, Wiesbaden, Germany, AF2864 | 1:200 |
| Donkey anti-mouse Alexa Fluor 488 | Abcam, ab150109 | 1:500 |
| Donkey anti-rabbit Alexa Fluor 555 | Abcam, ab150074 | 1:500 |
| Donkey anti-goat Alexa Fluor 647 | Abcam, ab150131 | 1:500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mak, S.; Hammes, A. Canonical and Non-Canonical Localization of Tight Junction Proteins during Early Murine Cranial Development. Int. J. Mol. Sci. 2024, 25, 1426. https://doi.org/10.3390/ijms25031426
Mak S, Hammes A. Canonical and Non-Canonical Localization of Tight Junction Proteins during Early Murine Cranial Development. International Journal of Molecular Sciences. 2024; 25(3):1426. https://doi.org/10.3390/ijms25031426
Chicago/Turabian StyleMak, Shermin, and Annette Hammes. 2024. "Canonical and Non-Canonical Localization of Tight Junction Proteins during Early Murine Cranial Development" International Journal of Molecular Sciences 25, no. 3: 1426. https://doi.org/10.3390/ijms25031426
APA StyleMak, S., & Hammes, A. (2024). Canonical and Non-Canonical Localization of Tight Junction Proteins during Early Murine Cranial Development. International Journal of Molecular Sciences, 25(3), 1426. https://doi.org/10.3390/ijms25031426