
Inhibitory Effect of a Tankyrase Inhibitor on Mechanical Stress-Induced Protease Expression in Human Articular Chondrocytes

 , , , , and
, , , , and 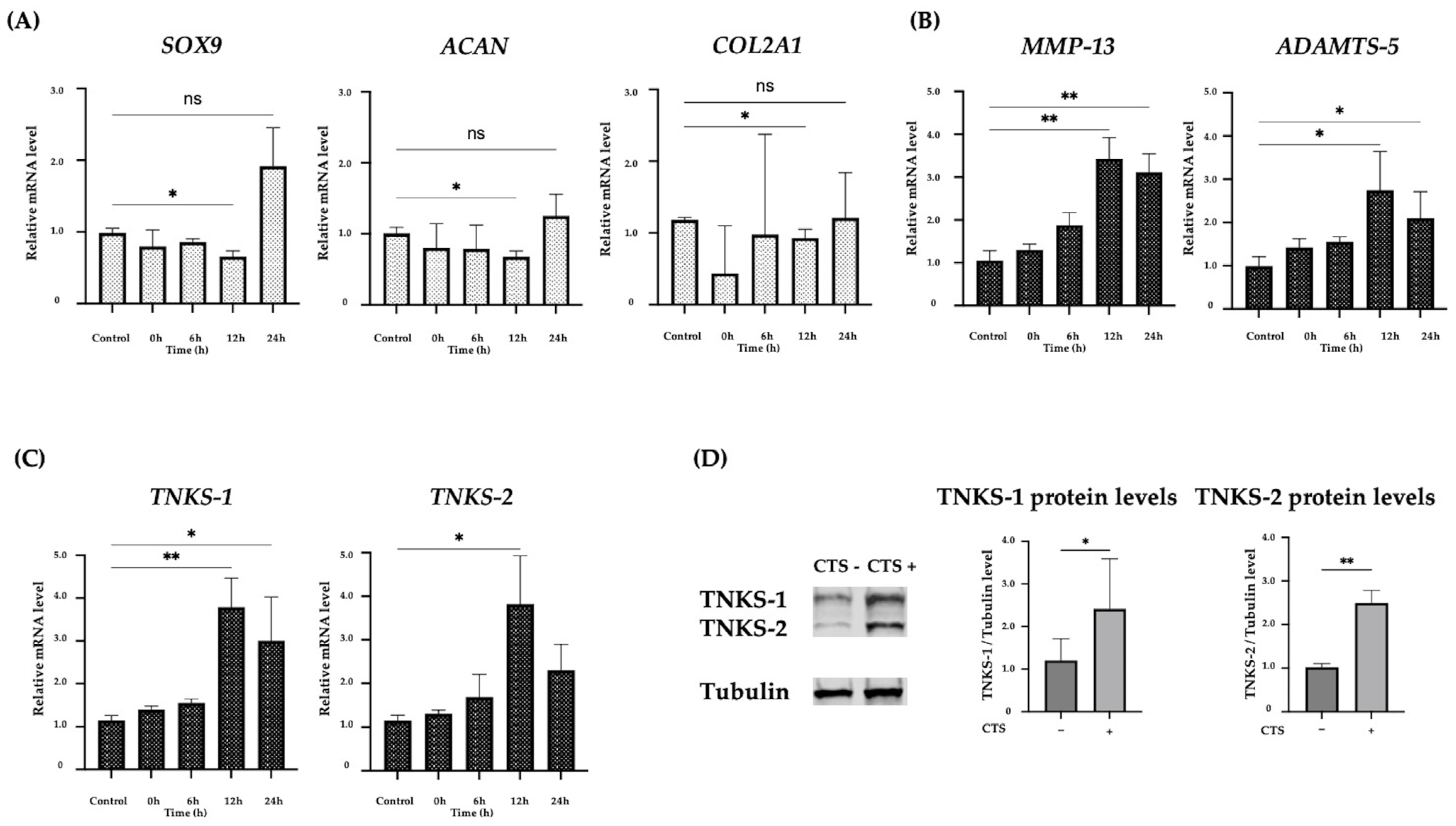{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effect of CTS Stimulation in Normal Human Chondrocytes
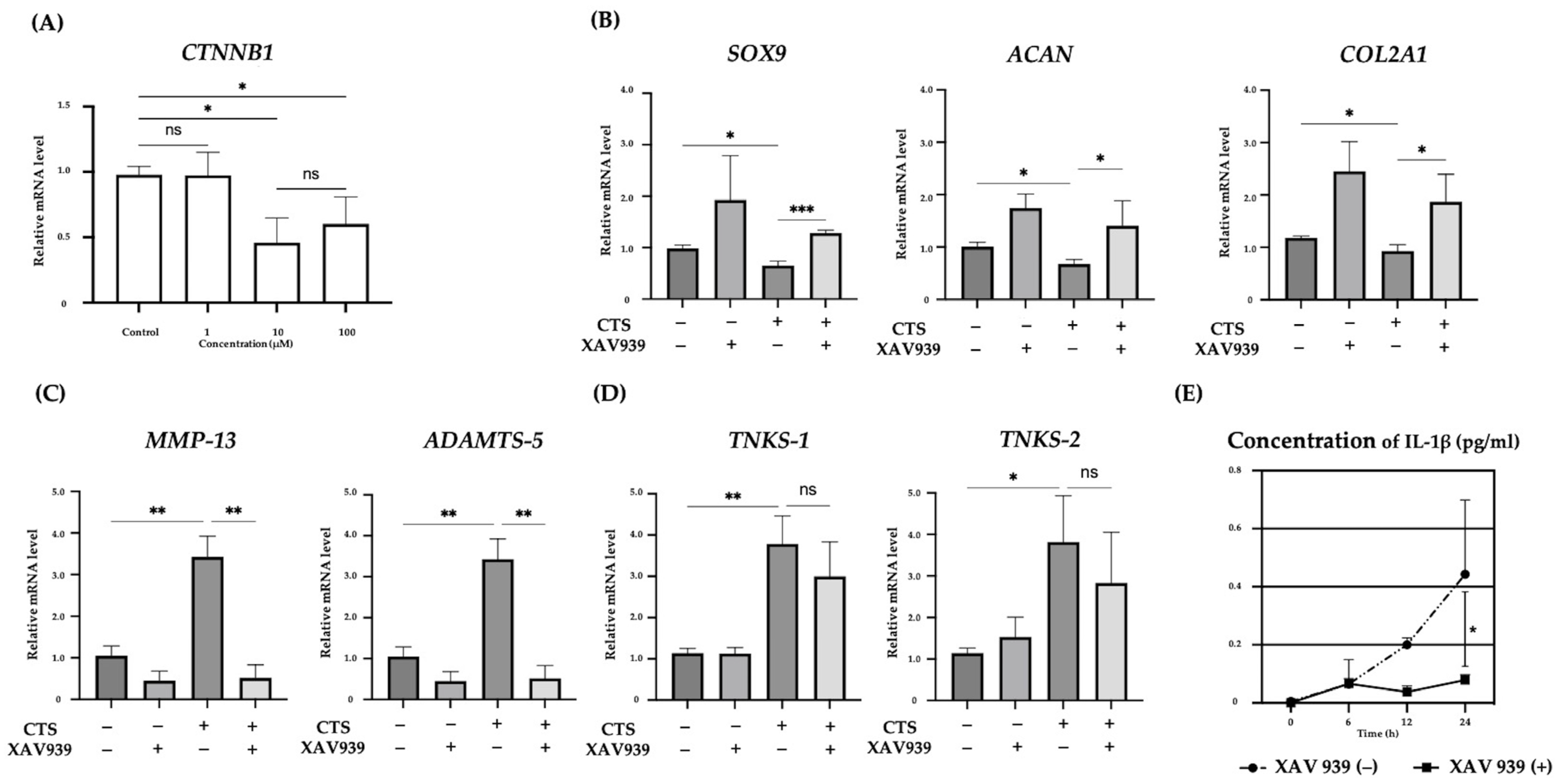
2.2. Effect of the TNKS-1/2 Inhibitor XAV939 on CTS-Induced Gene Expression and Concentration of IL-1β in the Supernatant in Normal Human Chondrocytes
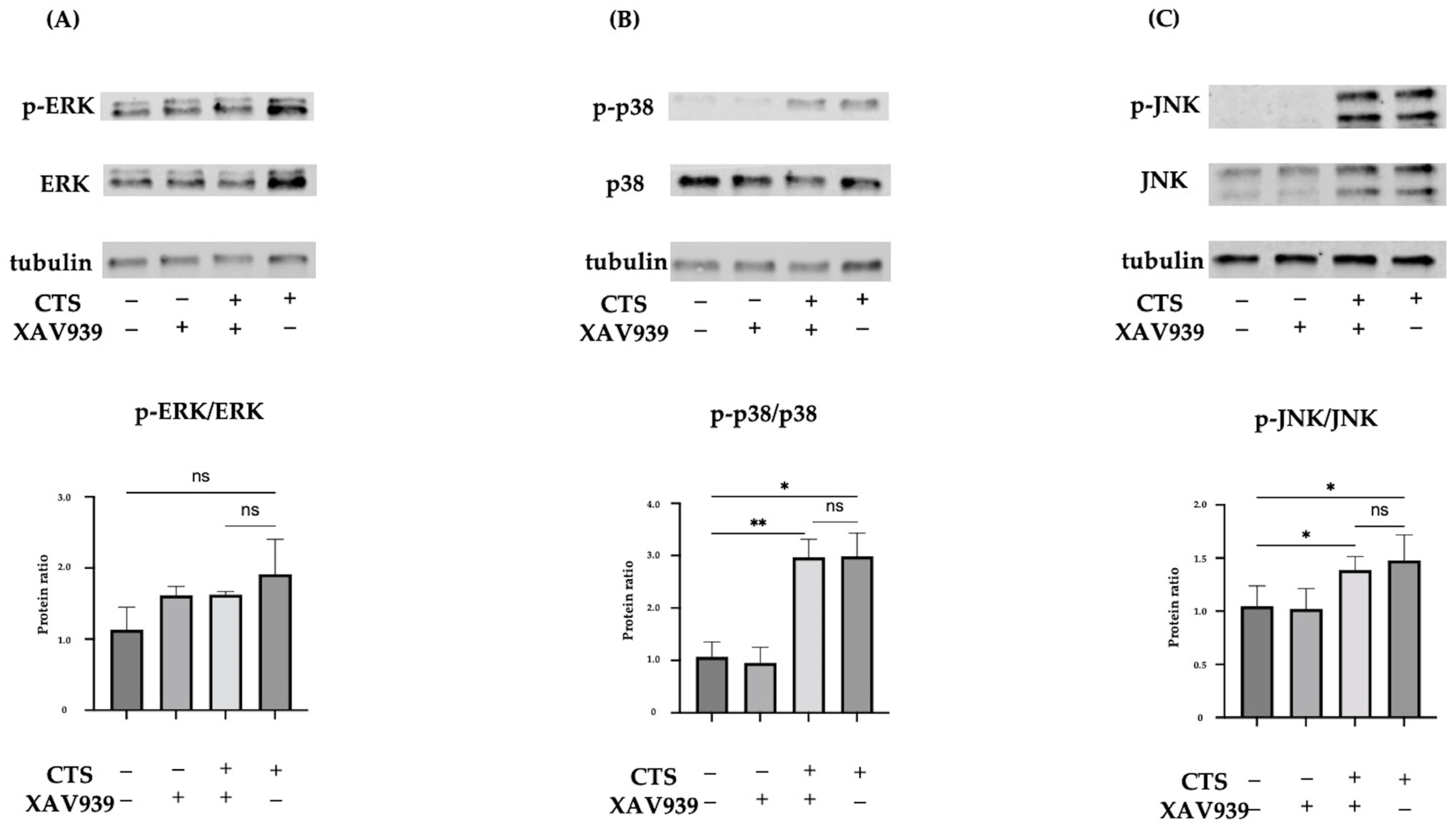
2.3. Effect of XAV939 on Activation of the MAPK Pathway in Normal Human Chondrocytes
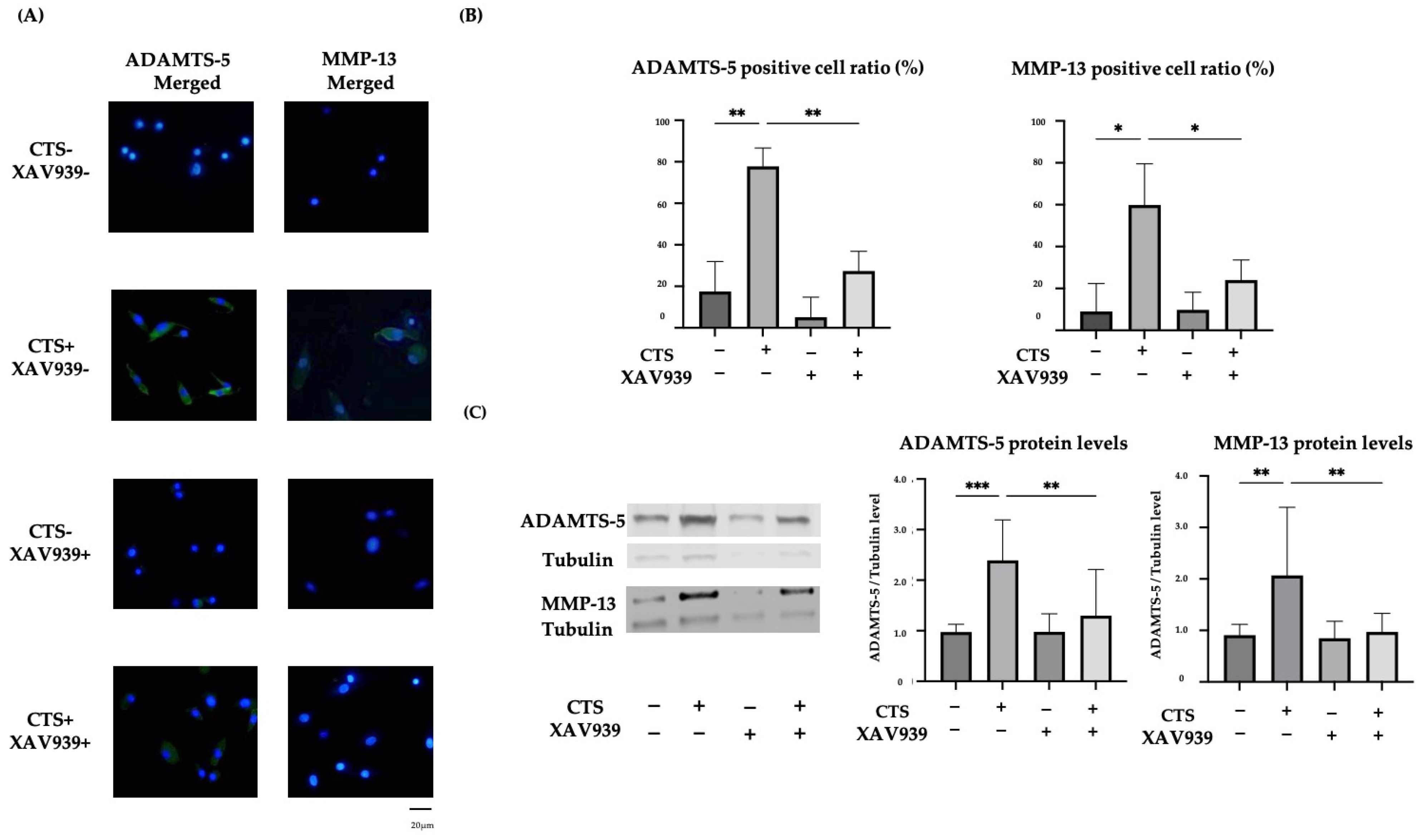
2.4. The TNKS-1/2 Inhibitor XAV939 Suppressed CTS-Induced ADAMTS-5 and MMP-13 Protein Expression in Normal Human Chondrocytes
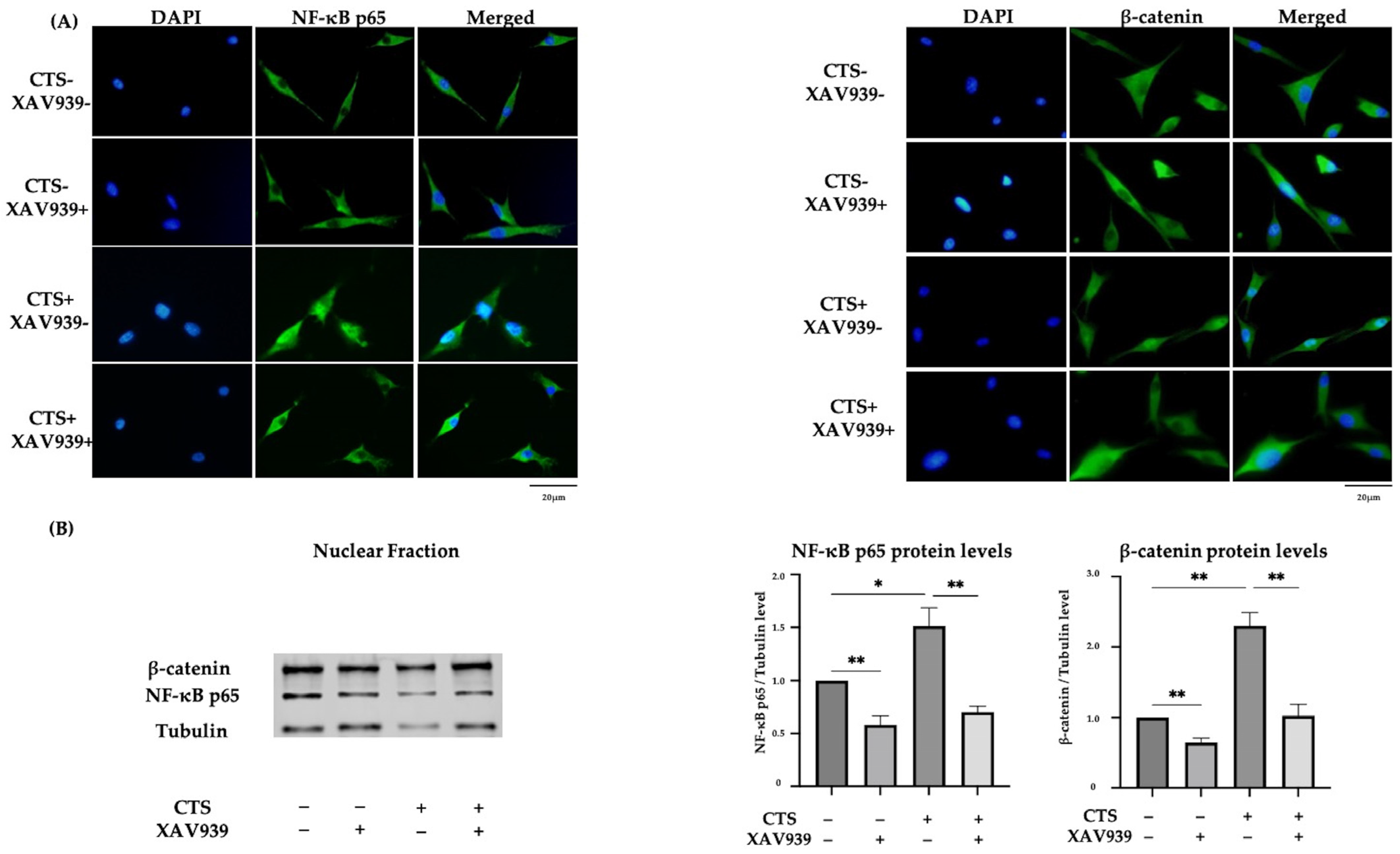
2.5. Effect of XAV939 on CTS-Induced Nuclear Translocation of NF-κB p65 and β-Catenin in Normal Human Chondrocytes
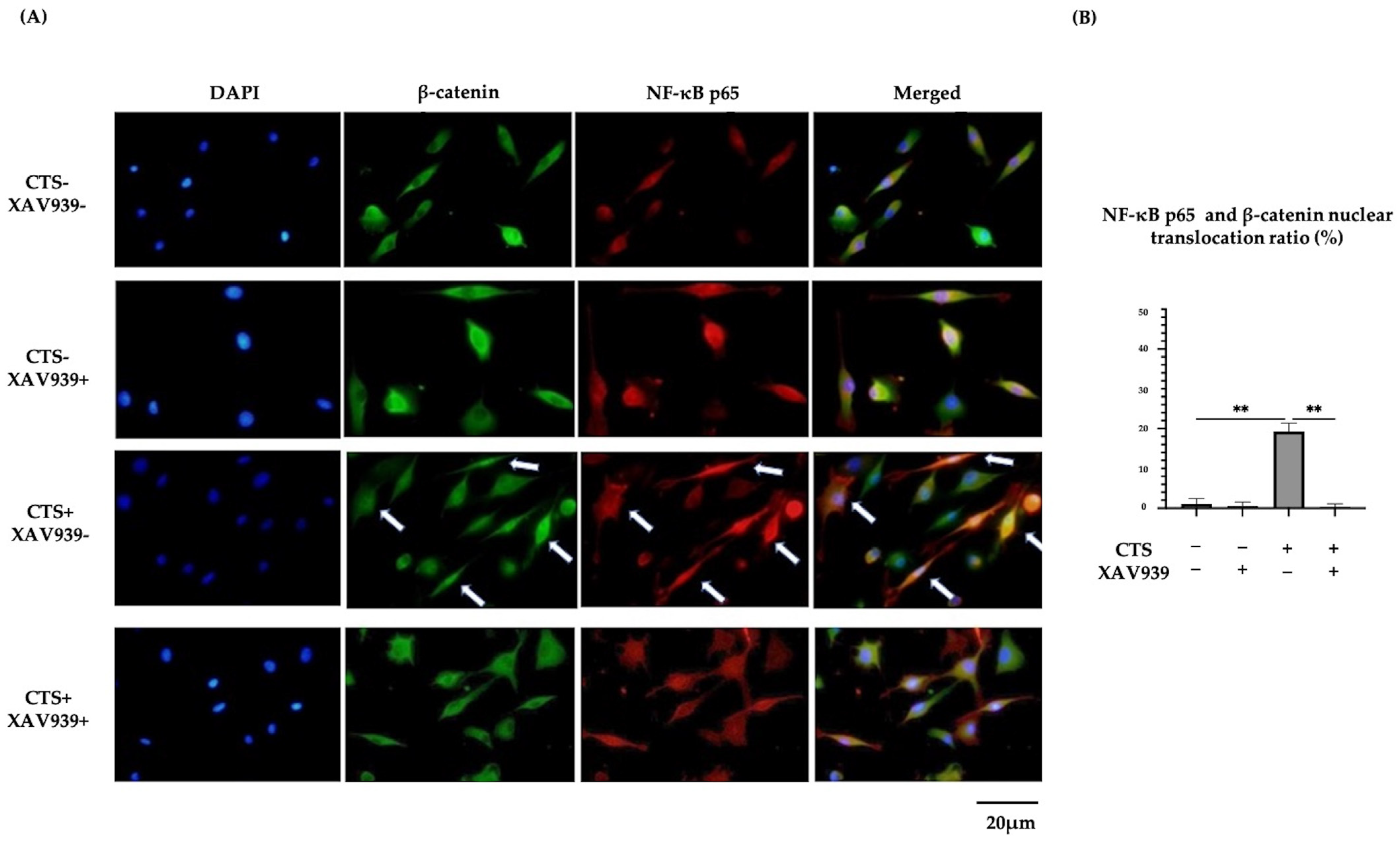
2.6. Double Immunocytochemical Staining of NF-κB p65 and β-Catenin in Normal Human Chondrocytes
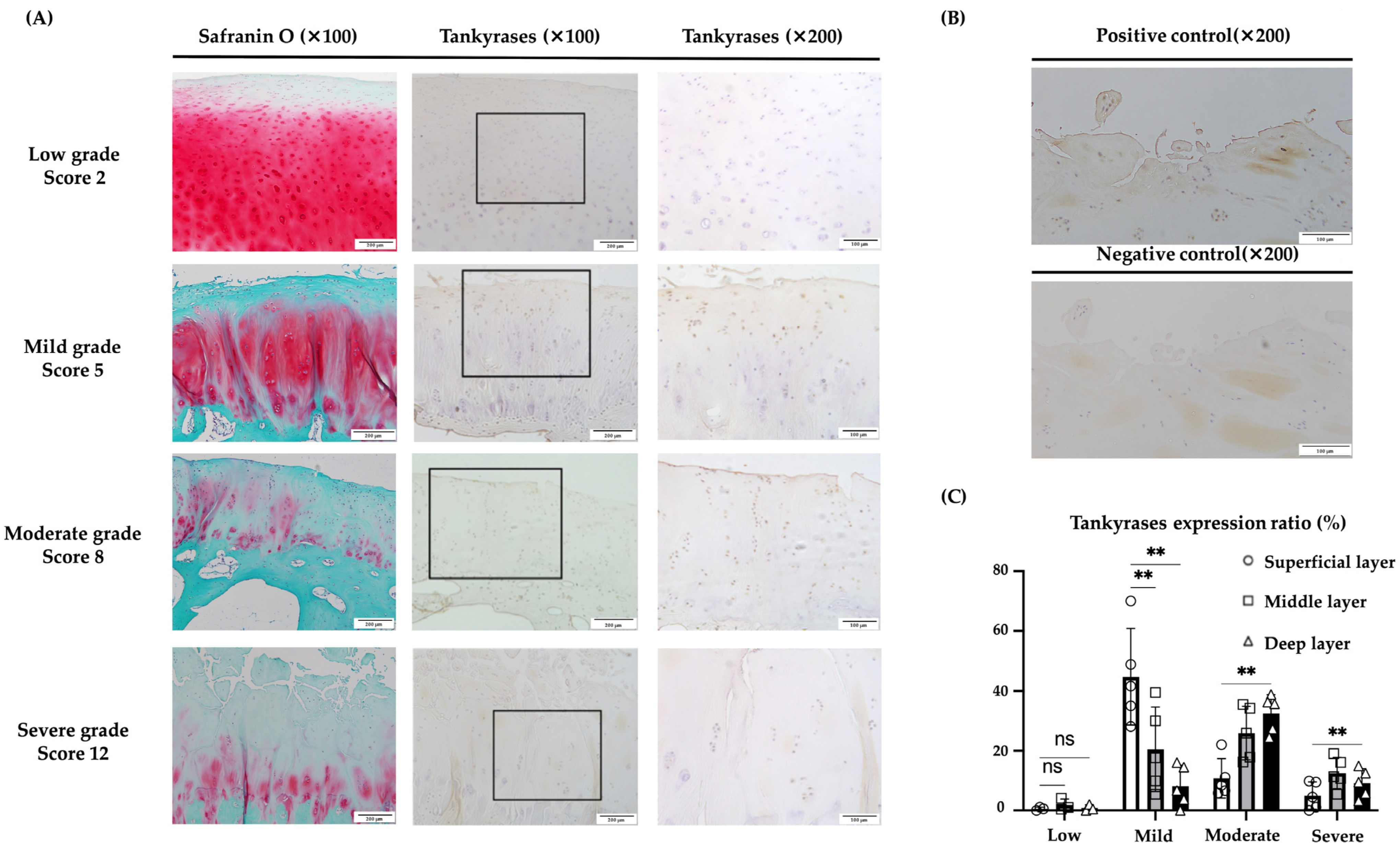
2.7. Immunohistochemical Evaluation of TNKS-1/2 in Human Articular Cartilage Tissues
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Culture
4.2. Cyclic Tensile Strain
4.3. Treatment with TNKS-1/2 Inhibitor
4.4. ELISA for IL-1β in Culture Medium
4.5. Quantitative Real-Time PCR Analysis
4.6. Immunocytochemistry
4.7. Western Blot Analysis
4.8. Clinical Samples of Human Cartilage Tissue
4.9. Histological Evaluation of Cartilage Destruction
4.10. Immunohistochemical Evaluation of TNKS-1/2 in Articular Cartilage and the Expression Ratio in Chondrocytes
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dequeker, J.; Luyten, F.P. The history of osteoarthritis-osteoarthrosis. Ann. Rheum. Dis. 2008, 67, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F. The Role of Aging in the Development of Osteoarthritis. Trans. Am. Clin. Climatol. Assoc. 2017, 128, 44–54. [Google Scholar] [PubMed]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Ushiyama, T.; Chano, T.; Inoue, K.; Matsusue, Y. Cytokine production in the infrapatellar fat pad: Another source of cytokines in knee synovial fluids. Ann. Rheum. Dis. 2003, 62, 108–112. [Google Scholar] [CrossRef]
- Mow, V.C.; Ratcliffe, A.; Poole, A.R. Cartilage and diarthrodial joints as paradigms for hierarchical materials and structures. Biomaterials 1992, 13, 67–97. [Google Scholar] [CrossRef]
- Hashimoto, M.; Nakasa, T.; Hikata, T.; Asahara, H. Molecular network of cartilage homeostasis and osteoarthritis. Med. Res. Rev. 2008, 28, 464–481. [Google Scholar] [CrossRef]
- Lefebvre, V.; de Crombrugghe, B. Toward understanding SOX9 function in chondrocyte differentiation. Matrix Biol. 1998, 16, 529–540. [Google Scholar] [CrossRef]
- Fukui, N.; Ikeda, Y.; Ohnuki, T.; Tanaka, N.; Hikita, A.; Mitomi, H.; Mori, T.; Juji, T.; Katsuragawa, Y.; Yamamoto, S.; et al. Regional differences in chondrocyte metabolism in osteoarthritis: A detailed analysis by laser capture microdissection. Arthritis Rheum. 2008, 58, 154–163. [Google Scholar] [CrossRef]
- Zhu, J.; Zhu, Y.; Xiao, W.; Hu, Y.; Li, Y. Instability and excessive mechanical loading mediate subchondral bone changes to induce osteoarthritis. Ann. Transl. Med. 2020, 8, 350. [Google Scholar] [CrossRef]
- Fang, T.; Zhou, X.; Jin, M.; Nie, J.; Li, X. Molecular mechanisms of mechanical load-induced osteoarthritis. Int. Orthop. 2021, 45, 1125–1136. [Google Scholar] [CrossRef]
- Belluzzi, E.; Todros, S.; Pozzuoli, A.; Ruggieri, P.; Carniel, E.L.; Berardo, A. Human Cartilage Biomechanics: Experimental and Theoretical Approaches towards the Identification of Mechanical Properties in Healthy and Osteoarthritic Conditions. Processes 2023, 11, 1014. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, S.; Su, H.; Cheng, J. Isoliquiritigenin Inhibits IL-1β-Induced Production of Matrix Metalloproteinase in Articular Chondrocytes. Mol. Ther. Methods Clin. Dev. 2018, 9, 153–159. [Google Scholar] [CrossRef]
- Tetsunaga, T.; Nishida, K.; Furumatsu, T.; Naruse, K.; Hirohata, S.; Yoshida, A.; Saito, T.; Ozaki, T. Regulation of mechanical stress-induced MMP-13 and ADAMTS-5 expression by RUNX-2 transcriptional factor in SW1353 chondrocyte-like cells. Osteoarthr. Cartil. 2011, 19, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sampson, E.R.; Jin, H.; Li, J.; Ke, Q.H.; Im, H.J.; Chen, D. MMP13 is a critical target gene during the progression of osteoarthritis. Arthritis Res. Ther. 2013, 15, R5. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Askew, R.; Sheppard, B.; Carito, B.; Blanchet, T.; Ma, H.L.; Flannery, C.R.; Peluso, D.; Kanki, K.; Yang, Z.; et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature 2005, 434, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, A.M.; Rome, B.N.; Reichmann, W.M.; Collins, J.E.; Burbine, S.A.; Thornhill, T.S.; Wright, J.; Katz, J.N.; Losina, E. Estimating the burden of total knee replacement in the United States. J. Bone Jt. Surg. Am. 2013, 95, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Jeong, S.; Kim, H.; Kang, D.; Lee, J.; Kang, S.B.; Kim, J.H. Disease-modifying therapeutic strategies in osteoarthritis: Current status and future directions. Exp. Mol. Med. 2021, 53, 1689–1696. [Google Scholar] [CrossRef]
- Riffell, J.L.; Lord, C.J.; Ashworth, A. Tankyrase-targeted therapeutics: Expanding opportunities in the PARP family. Nat. Rev. Drug Discov. 2012, 11, 923–936. [Google Scholar] [CrossRef]
- Seimiya, H.; Muramatsu, Y.; Ohishi, T.; Tsuruo, T. Tankyrase 1 as a target for telomere-directed molecular cancer therapeutics. Cancer Cell 2005, 7, 25–37. [Google Scholar] [CrossRef]
- Mariotti, L.; Pollock, K.; Guettler, S. Regulation of Wnt/β-catenin signalling by tankyrase-dependent poly(ADP-ribosyl)ation and scaffolding. Br. J. Pharmacol. 2017, 174, 4611–4636. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.Y.; Sbodio, J.I.; Tsun, Z.Y.; Luo, B.; Chi, N.W. Insulin-stimulated exocytosis of GLUT4 is enhanced by IRAP and its partner tankyrase. Biochem. J. 2007, 402, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Levaot, N.; Voytyuk, O.; Dimitriou, I.; Sircoulomb, F.; Chandrakumar, A.; Deckert, M.; Krzyzanowski, P.M.; Scotter, A.; Gu, S.; Janmohamed, S.; et al. Loss of Tankyrase-mediated destruction of 3BP2 is the underlying pathogenic mechanism of cherubism. Cell 2011, 147, 1324–1339. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, S.J.; Smith, S. Tankyrase function at telomeres, spindle poles, and beyond. Biochimie 2008, 90, 83–92. [Google Scholar] [CrossRef]
- Chiang, Y.J.; Hsiao, S.J.; Yver, D.; Cushman, S.W.; Tessarollo, L.; Smith, S.; Hodes, R.J. Tankyrase 1 and tankyrase 2 are essential but redundant for mouse embryonic development. PLoS ONE 2008, 3, e2639. [Google Scholar] [CrossRef]
- Kim, S.; Han, S.; Kim, Y.; Kim, H.S.; Gu, Y.R.; Kang, D.; Cho, Y.; Kim, H.; Lee, J.; Seo, Y.; et al. Tankyrase inhibition preserves osteoarthritic cartilage by coordinating cartilage matrix anabolism via effects on SOX9 PARylation. Nat. Commun. 2019, 10, 4898. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Nishida, K.; Furumatsu, T.; Yoshida, A.; Ozawa, M.; Ozaki, T. Histone deacetylase inhibitors suppress mechanical stress-induced expression of RUNX-2 and ADAMTS-5 through the inhibition of the MAPK signaling pathway in cultured human chondrocytes. Osteoarthr. Cartil. 2013, 21, 165–174. [Google Scholar] [CrossRef]
- Ozawa, M.; Nishida, K.; Yoshida, A.; Saito, T.; Harada, R.; Machida, T.; Ozaki, T. Hyaluronan suppresses mechanical stress-induced expression of catabolic enzymes by human chondrocytes via inhibition of IL-1β production and subsequent NF-κB activation. Inflamm. Res. 2015, 64, 243–252. [Google Scholar] [CrossRef]
- Machida, T.; Nishida, K.; Nasu, Y.; Nakahara, R.; Ozawa, M.; Harada, R.; Horita, M.; Takeshita, A.; Kaneda, D.; Yoshida, A.; et al. Inhibitory effect of JAK inhibitor on mechanical stress-induced protease expression by human articular chondrocytes. Inflamm. Res. 2017, 66, 999–1009. [Google Scholar] [CrossRef]
- Pfander, D.; Cramer, T.; Weseloh, G.; Pullig, O.; Schuppan, D.; Bauer, M.; Swoboda, B. Hepatocyte growth factor in human osteoarthritic cartilage. Osteoarthr. Cartil. 1999, 7, 548–559. [Google Scholar] [CrossRef]
- Gabay, O.; Hall, D.J.; Berenbaum, F.; Henrotin, Y.; Sanchez, C. Osteoarthritis and obesity: Experimental models. Jt. Bone Spine 2008, 75, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Bleuel, J.; Zaucke, F.; Brüggemann, G.P.; Niehoff, A. Effects of cyclic tensile strain on chondrocyte metabolism: A systematic review. PLoS ONE 2015, 10, e0119816. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, Q.; Wang, X.; Kang, L.; Fu, Y.; Yin, Y.; Li, Z.; Liu, Y.; Xu, X.; Wang, Y. SOX9 is a regulator of ADAMTSs-induced cartilage degeneration at the early stage of human osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2259–2268. [Google Scholar] [CrossRef]
- Kobayashi, H.; Hirata, M.; Saito, T.; Itoh, S.; Chung, U.I.; Kawaguchi, H. Transcriptional induction of ADAMTS5 protein by nuclear factor-κB (NF-κB) family member RelA/p65 in chondrocytes during osteoarthritis development. J. Biol. Chem. 2013, 288, 28620–28629. [Google Scholar] [CrossRef] [PubMed]
- Mengshol, J.A.; Vincenti, M.P.; Coon, C.I.; Barchowsky, A.; Brinckerhoff, C.E. Interleukin-1 induction of collagenase 3 (matrix metalloproteinase 13) gene expression in chondrocytes requires p38, c-Jun N-terminal kinase, and nuclear factor kappaB: Differential regulation of collagenase 1 and collagenase 3. Arthritis Rheum. 2000, 43, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, J.; El Mabrouk, M.; Ahmad, R.; Chaudry, A.; Zafarullah, M. Interleukin-1 induction of aggrecanase gene expression in human articular chondrocytes is mediated by mitogen-activated protein kinases. Cell Physiol. Biochem. 2012, 30, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Miclea, R.L.; Siebelt, M.; Finos, L.; Goeman, J.J.; Löwik, C.W.; Oostdijk, W.; Weinans, H.; Wit, J.M.; Robanus-Maandag, E.C.; Karperien, M. Inhibition of Gsk3β in cartilage induces osteoarthritic features through activation of the canonical Wnt signaling pathway. Osteoarthr. Cartil. 2011, 19, 1363–1372. [Google Scholar] [CrossRef]
- Jiang, L.; Lin, J.; Zhao, S.; Wu, J.; Jin, Y.; Yu, L.; Wu, N.; Wu, Z.; Wang, Y.; Lin, M. ADAMTS5 in Osteoarthritis: Biological Functions, Regulatory Network, and Potential Targeting Therapies. Front. Mol. Biosci. 2021, 8, 703110. [Google Scholar] [CrossRef]
- Attur, M.; Yang, Q.; Shimada, K.; Tachida, Y.; Nagase, H.; Mignatti, P.; Statman, L.; Palmer, G.; Kirsch, T.; Beier, F.; et al. Elevated expression of periostin in human osteoarthritic cartilage and its potential role in matrix degradation via matrix metalloproteinase-13. FASEB J. 2015, 29, 4107–4121. [Google Scholar] [CrossRef]
- Kaminker, P.G.; Kim, S.H.; Taylor, R.D.; Zebarjadian, Y.; Funk, W.D.; Morin, G.B.; Yaswen, P.; Campisi, J. TANK2, a new TRF1-associated poly(ADP-ribose) polymerase, causes rapid induction of cell death upon overexpression. J. Biol. Chem. 2001, 276, 35891–35899. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Yang, Y.; Ueberheide, B.; Smith, S. Whole proteome analysis of human tankyrase knockout cells reveals targets of tankyrase-mediated degradation. Nat. Commun. 2017, 8, 2214. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Haikarainen, T.; Krauss, S.; Lehtio, L. Tankyrases: Structure, function and therapeutic implications in cancer. Curr. Pharm. Des. 2014, 20, 6472–6488. [Google Scholar] [CrossRef] [PubMed]
- Lehtiö, L.; Chi, N.W.; Krauss, S. Tankyrases as drug targets. FEBS J. 2013, 280, 3576–3593. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K. Novel insight into the function of tankyrase. Oncol. Lett. 2018, 16, 6895–6902. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small molecule-mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Song, C.X.; Liu, S.Y.; Zhu, W.T.; Xu, S.Y.; Ni, G.X. Excessive mechanical stretch-mediated osteoblasts promote the catabolism and apoptosis of chondrocytes via the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2021, 24, 593. [Google Scholar] [CrossRef]
- Ma, B.; van Blitterswijk, C.A.; Karperien, M. A Wnt/β-catenin negative feedback loop inhibits interleukin-1-induced matrix metalloproteinase expression in human articular chondrocytes. Arthritis Rheum. 2012, 64, 2589–2600. [Google Scholar] [CrossRef]
- Yun, K.; So, J.S.; Jash, A.; Im, S.H. Lymphoid enhancer binding factor 1 regulates transcription through gene looping. J. Immunol. 2009, 183, 5129–5137. [Google Scholar] [CrossRef]
- Ma, B.; Zhong, L.; van Blitterswijk, C.A.; Post, J.N.; Karperien, M. T cell factor 4 is a pro-catabolic and apoptotic factor in human articular chondrocytes by potentiating nuclear factor κB signaling. J. Biol. Chem. 2013, 288, 17552–17558. [Google Scholar] [CrossRef]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef]
- Yun, K.; Choi, Y.D.; Nam, J.H.; Park, Z.; Im, S.H. NF-kappaB regulates Lef1 gene expression in chondrocytes. Biochem. Biophys. Res. Commun. 2007, 357, 589–595. [Google Scholar] [CrossRef]
- Li, Z.; Dai, A.; Yang, M.; Chen, S.; Deng, Z.; Li, L. p38MAPK Signaling Pathway in Osteoarthritis: Pathological and Therapeutic Aspects. J. Inflamm. Res. 2022, 15, 723–734. [Google Scholar] [CrossRef]
- Zhang, Y.; Pizzute, T.; Pei, M. A review of crosstalk between MAPK and Wnt signals and its impact on cartilage regeneration. Cell Tissue Res. 2014, 358, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Ma, X.; Meng, J.; Zhang, C.; Ma, K.; Zhou, C. Role of Wnt-5A in interleukin-1beta-induced matrix metalloproteinase expression in rabbit temporomandibular joint condylar chondrocytes. Arthritis Rheum. 2009, 60, 2714–2722. [Google Scholar] [CrossRef]
- Jaeger, K.; Selent, C.; Jaehme, W.; Mahr, S.; Goebel, U.; Ibrahim, S.; Vollmar, B.; Mueller-Hilke, B. The genetics of osteoarthritis in STR/ort mice. Osteoarthr. Cartil. 2008, 16, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Keam, B.; Hwang, M.Y.; Lee, Y.; Park, S.; Oh, J.H.; Kim, Y.J.; Lee, H.S.; Kim, N.H.; Kim, Y.J.; et al. A genome-wide association study of copy-number variation identifies putative loci associated with osteoarthritis in Koreans. BMC Musculoskelet. Disord. 2015, 16, 76. [Google Scholar] [CrossRef]
- Fukui, N.; Miyamoto, Y.; Nakajima, M.; Ikeda, Y.; Hikita, A.; Furukawa, H.; Mitomi, H.; Tanaka, N.; Katsuragawa, Y.; Yamamoto, S.; et al. Zonal gene expression of chondrocytes in osteoarthritic cartilage. Arthritis Rheum. 2008, 58, 3843–3853. [Google Scholar] [CrossRef]
- Li, H.; Wang, D.; Yuan, Y.; Min, J. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Res. Ther. 2017, 19, 248. [Google Scholar] [CrossRef]
- Kouri, J.B.; Jiménez, S.A.; Quintero, M.; Chico, A. Ultrastructural study of chondrocytes from fibrillated and non-fibrillated human osteoarthritic cartilage. Osteoarthr. Cartil. 1996, 4, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Naruse, K.; Yamada, T.; Sokabe, M. Involvement of SA channels in orienting response of cultured endothelial cells to cyclic stretch. Am. J. Physiol. 1998, 274, H1532–H1538. [Google Scholar] [CrossRef] [PubMed]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J. Bone Jt. Surg. Am. 1971, 53, 523–537. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotta, Y.; Nishida, K.; Yoshida, A.; Nasu, Y.; Nakahara, R.; Naniwa, S.; Shimizu, N.; Ichikawa, C.; Lin, D.; Fujiwara, T.; et al. Inhibitory Effect of a Tankyrase Inhibitor on Mechanical Stress-Induced Protease Expression in Human Articular Chondrocytes. Int. J. Mol. Sci. 2024, 25, 1443. https://doi.org/10.3390/ijms25031443
Hotta Y, Nishida K, Yoshida A, Nasu Y, Nakahara R, Naniwa S, Shimizu N, Ichikawa C, Lin D, Fujiwara T, et al. Inhibitory Effect of a Tankyrase Inhibitor on Mechanical Stress-Induced Protease Expression in Human Articular Chondrocytes. International Journal of Molecular Sciences. 2024; 25(3):1443. https://doi.org/10.3390/ijms25031443
Chicago/Turabian StyleHotta, Yoshifumi, Keiichiro Nishida, Aki Yoshida, Yoshihisa Nasu, Ryuichi Nakahara, Shuichi Naniwa, Noriyuki Shimizu, Chinatsu Ichikawa, Deting Lin, Tomohiro Fujiwara, and et al. 2024. "Inhibitory Effect of a Tankyrase Inhibitor on Mechanical Stress-Induced Protease Expression in Human Articular Chondrocytes" International Journal of Molecular Sciences 25, no. 3: 1443. https://doi.org/10.3390/ijms25031443
APA StyleHotta, Y., Nishida, K., Yoshida, A., Nasu, Y., Nakahara, R., Naniwa, S., Shimizu, N., Ichikawa, C., Lin, D., Fujiwara, T., & Ozaki, T. (2024). Inhibitory Effect of a Tankyrase Inhibitor on Mechanical Stress-Induced Protease Expression in Human Articular Chondrocytes. International Journal of Molecular Sciences, 25(3), 1443. https://doi.org/10.3390/ijms25031443