
Safe Procedure for Efficient Hydrodynamic Gene Transfer to Isolated Porcine Liver in Transplantation

 , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
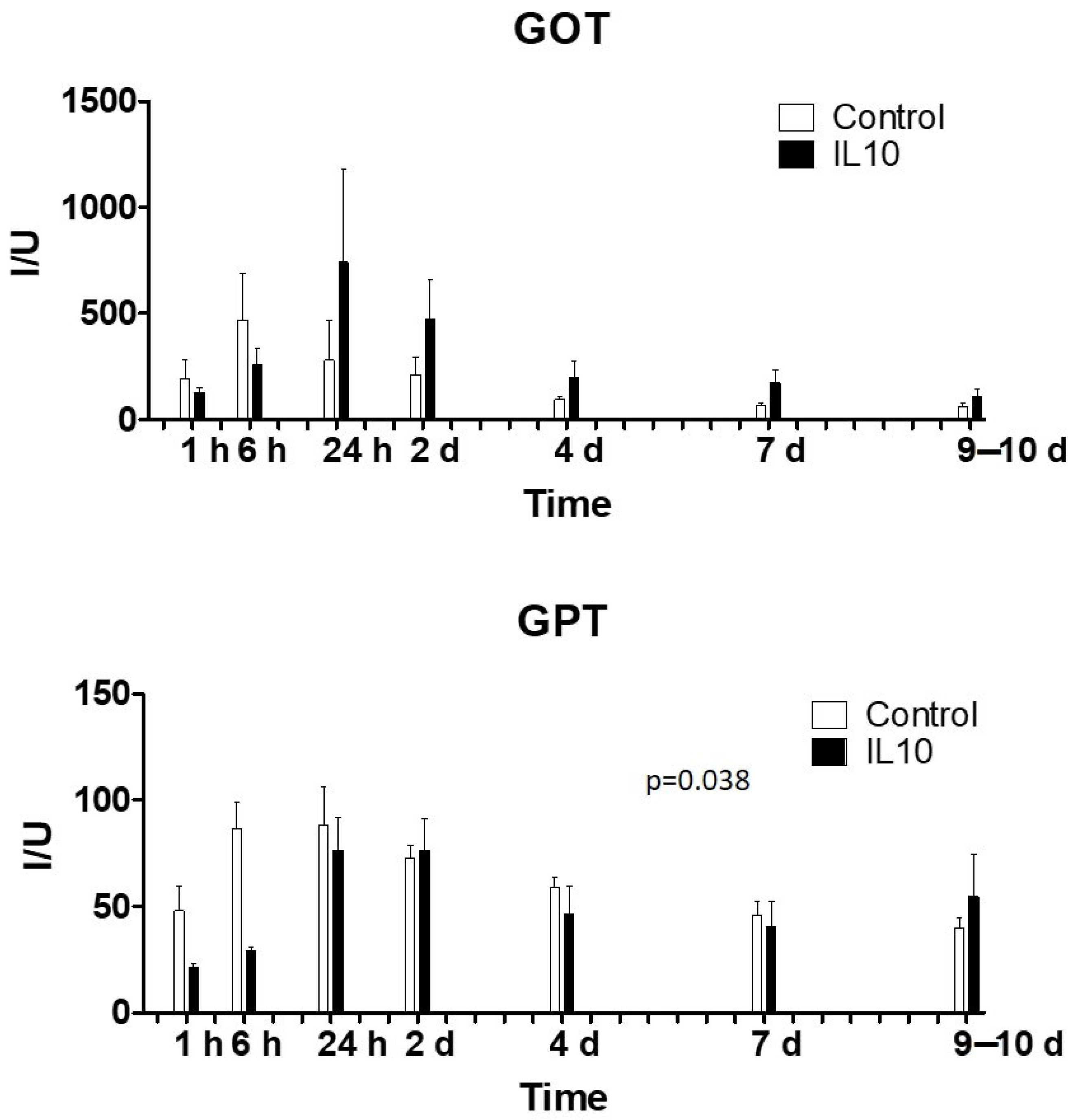
2.1. Biochemical Variables and Liver Function
2.2. Human IL-10 Protein Concentration in Plasma
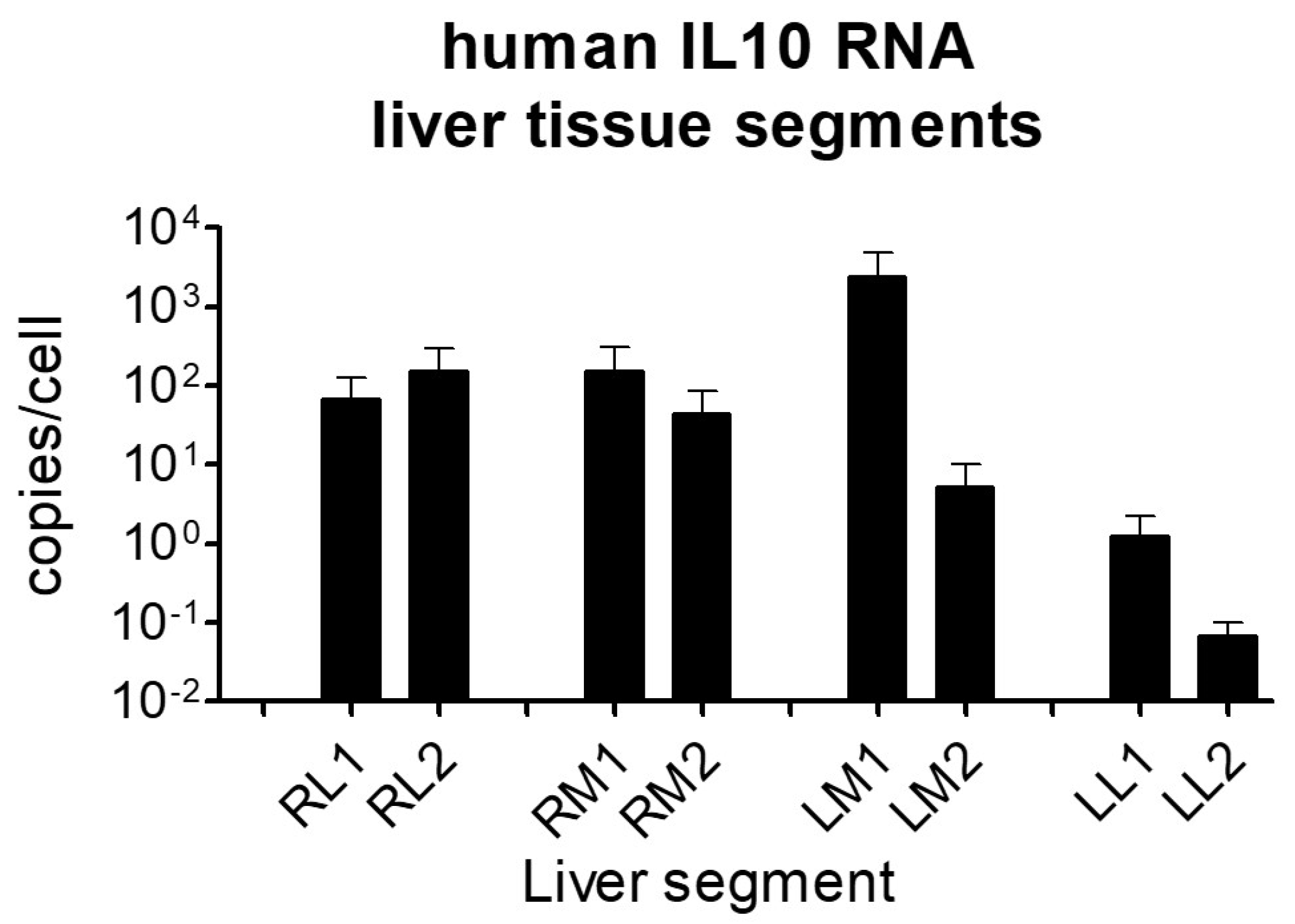
2.3. Human IL-10 RNA Expression in Liver Tissue Samples
2.4. Expression of Porcine Interleukin-10
2.5. Plasma Levels of Other Porcine Cytokines
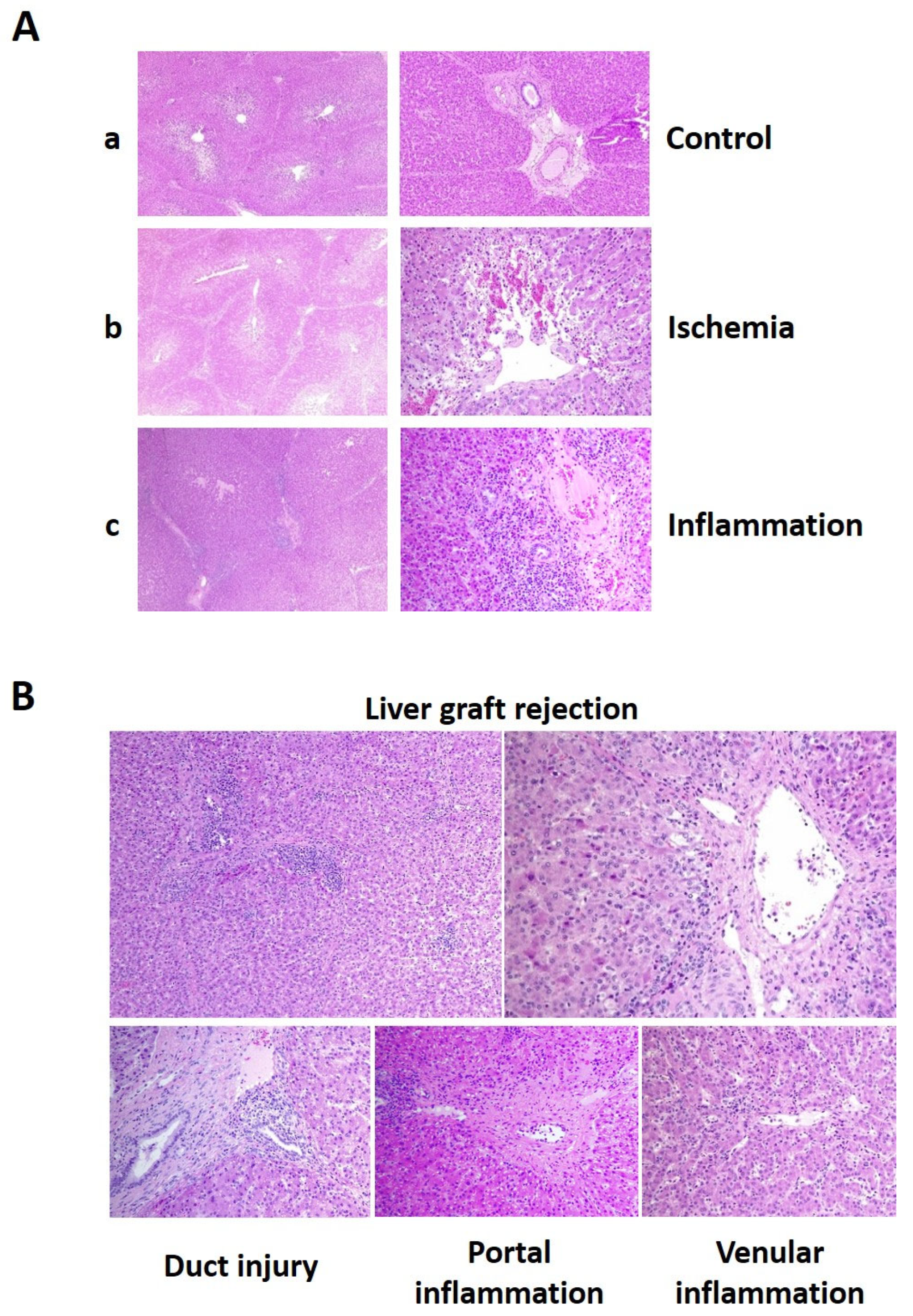
2.6. Histological Evaluation of Liver Injury
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Surgical Procedure
4.2.1. Donor
4.2.2. Bench Surgery and Gene Transfer
4.2.3. Recipient
4.3. Post-Transplantation Care
4.4. Plasmid
4.5. Biochemical Parameters and Liver Enzymes Determination
4.6. Quantitative PCR and RT-PCR
4.7. Cytokine Determination
4.7.1. hIL-10 ELISA
4.7.2. Determination of Porcine Cytokines
4.8. Anatomopathological Evaluation of Tissue Samples
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchwald, J.E.; Martins, P.N. Designer organs: The future of personalized transplantation. Artif. Organs 2022, 46, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Sun, R.; Huang, Q.; Tian, Z. Technical Improvement and Application of Hydrodynamic Gene Delivery in Study of Liver Diseases. Front. Pharmacol. 2017, 8, 591. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Yokoo, T.; Kanefuji, T.; Kamimura, K.; Zhang, G.; Liu, D. Hydrodynamic Delivery: Characteristics, Applications, and Technological Advances. Pharmaceutics 2023, 15, 1111. [Google Scholar] [CrossRef] [PubMed]
- Kumbhari, V.; Li, L.; Piontek, K.; Ishida, M.; Fu, R.; Khalil, B.; Garrett, C.M.; Liapi, E.; Kalloo, A.N.; Selaru, F.M. Successful liver-directed gene delivery by ERCP-guided hydrodynamic injection (with videos). Gastrointest. Endosc. 2018, 88, 755–763.e5. [Google Scholar] [CrossRef]
- Hickman, M.A.; Malone, R.W.; Lehmann-Bruinsma, K.; Sih, T.R.; Knoell, D.; Szoka, F.C.; Walzem, R.L.; Carlson, D.M.; Powell, J.S.; Shekhar, S.; et al. Gene expression following direct injection of DNA into liver. Hum. Gene Ther. 1994, 5, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Budker, V.; Zhang, G.; Knechtle, S.; A Wolff, J. Naked DNA delivered intraportally expresses efficiently in hepatocytes. Gene Ther. 1996, 3, 593–598. [Google Scholar] [PubMed]
- Liu, F.; Song, Y.K.; Liu, D. Hydrodynamics-based transfection in animals by systemic administration of plasmid DNA. Gene Ther. 1999, 6, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Budker, V.; Wolff, J.A.; Lee, H.-O.; Gallego-Villar, L.; Grisch-Chan, H.M.; Häberle, J.; Thöny, B.; Kruger, W.D.; Hyland, K.A.; Aronovich, E.L.; et al. High Levels of Foreign Gene Expression in Hepatocytes after Tail Vein Injections of Naked Plasmid DNA. Hum. Gene Ther. 1999, 10, 1735–1737. [Google Scholar] [CrossRef]
- Aliño, S.F.; Crespo, A.; Dasí, F. Long-term therapeutic levels of human alpha-1 antitrypsin in plasma after hydrodynamic injection of nonviral DNA. Gene Ther. 2003, 10, 1672–1679. [Google Scholar] [CrossRef]
- Yoshino, H.; Hashizume, K.; Kobayashi, E. Naked plasmid DNA transfer to the porcine liver using rapid injection with large volume. Gene Ther. 2006, 13, 1696–1702. [Google Scholar] [CrossRef]
- Aliño, S.F.; Herrero, M.J.; Noguera, I.; Dasí, F.; Sánchez, M. Pig liver gene therapy by noninvasive interventionist catheterism. Gene Ther. 2007, 14, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Suda, K.; Liu, D. Computer-assisted Hydrodynamic Gene Delivery. Mol. Ther. 2008, 16, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Fabre, J.W.; Whitehorne, M.; Grehan, A.; Sawyer, G.J.; Zhang, X.; Davenport, M.; Rela, M.; Eastman, S.J.; Baskin, K.M.; Hodges, B.L.; et al. Critical Physiological and Surgical Considerations for Hydrodynamic Pressurization of Individual Segments of the Pig Liver. Hum. Gene Ther. 2011, 22, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Sendra, L.; Herrero, M.J.; Aliño, S.F. Translational Advances of Hydrofection by Hydrodynamic Injection. Genes 2018, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Sendra, L.; Miguel, A.; Pérez-Enguix, D.; Herrero, M.J.; Montalvá, E.; García-Gimeno, M.A.; Noguera, I.; Díaz, A.; Pérez, J.; Sanz, P.; et al. Studying Closed Hydrodynamic Models of “In Vivo” DNA Perfusion in Pig Liver for Gene Therapy Translation to Humans. PLoS ONE 2016, 11, e0163898. [Google Scholar] [CrossRef] [PubMed]
- Carreño, O.; Sendra, L.; Montalvá, E.; Miguel, A.; Orbis, F.; Herrero, M.; Noguera, I.; Aliño, S.; Lopez-Andujar, R. A Surgical Model for Isolating the Pig Liver in vivo for Gene Therapy. Eur. Surg. Res. 2013, 51, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Sendra, L.; Herrero, M.J.; Montalvá, E.M.; Noguera, I.; Orbis, F.; Díaz, A.; Fernández-Delgado, R.; López-Andújar, R.; Aliño, S.F. Efficacy of interleukin 10 gene hydrofection in pig liver vascular isolated ‘in vivo’ by surgical procedure with interest in liver transplantation. PLoS ONE 2019, 14, e0224568. [Google Scholar] [CrossRef]
- Kamimura, K.; Kanefuji, T.; Suda, T.; Yokoo, T.; Zhang, G.; Aoyagi, Y.; Liu, D. Liver lobe-specific hydrodynamic gene delivery to baboons: A preclinical trial for hemophilia gene therapy. Mol. Ther. Nucleic Acids 2023, 32, 903–913. [Google Scholar] [CrossRef]
- Gisbert, L.S.; Matas, A.M.; Ortí, L.S.; Herrero, M.J.; Olivas, L.S.; Orón, E.M.M.; Frasson, M.; López, R.A.; López-Andújar, R.; Ximénez, E.G.; et al. Efficacy of hydrodynamic interleukin 10 gene transfer in human liver segments with interest in transplantation. Liver Transplant. 2017, 23, 50–62. [Google Scholar] [CrossRef]
- Gao, Q.; DeLaura, I.F.; Anwar, I.J.; Kesseli, S.J.; Kahan, R.; Abraham, N.; Asokan, A.; Barbas, A.S.; Hartwig, M.G. Gene Therapy: Will the Promise of Optimizing Lung Allografts Become Reality? Front. Immunol. 2022, 13, 931524. [Google Scholar] [CrossRef]
- Hirschfield, G.M.; Gibbs, P.; Griffiths, W.J.H. Adult liver transplantation: What non-specialists need to know. BMJ 2009, 338, b1670. [Google Scholar] [CrossRef]
- Rodríguez-Perálvarez, M.; Rico-Juri, J.M.; Tsochatzis, E.; Burra, P.; De la Mata, M.; Lerut, J. Biopsy-proven acute cellular rejection as an efficacy endpoint of randomized trials in liver transplantation: A systematic review and critical appraisal. Transpl. Int. 2016, 29, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Carlini, V.; Noonan, D.M.; Abdalalem, E.; Goletti, D.; Sansone, C.; Calabrone, L.; Albini, A. The multifaceted nature of IL-10: Regulation, role in immunological homeostasis and its relevance to cancer, COVID-19 and post-COVID conditions. Front. Immunol. 2023, 14, 1161067. [Google Scholar] [CrossRef] [PubMed]
- Mendiola Pla, M.; Chiang, Y.; Roan, J.N.; Bowles, D.E. Gene Therapy for Cardiac Transplantation [Internet]. Heart Transplantation—New Insights in Therapeutic Strategies; IntechOpen Limited: London, UK, 2022. [Google Scholar] [CrossRef]
- Blancho, G.; Gianello, P.; Germana, S.; Baetscher, M.; Sachs, D.H.; LeGuern, C. Molecular identification of porcine interleukin 10: Regulation of expression in a kidney allograft model. Proc. Natl. Acad. Sci. USA 1995, 92, 2800–2804. [Google Scholar] [CrossRef]
- Liu, K.S.; Fan, X.Q.; Zhang, L.; NA Wen, Q.; Feng, J.H.; Chen, F.C.; Luo, J.M.; Sun, W.B. Effects of recombinant human interleukin-10 on Treg cells, IL-10 and TGF-β in transplantation of rabbit skin. Mol. Med. Rep. 2013, 9, 639–644. [Google Scholar] [CrossRef]
- Martins, S.; de Perrot, M.; Imai, Y.; Yamane, M.; Quadri, S.; Segall, L.; Dutly, A.; Sakiyama, S.; Chaparro, A.; Davidson, B.; et al. Transbronchial administration of adenoviral-mediated interleukin-10 gene to the donor improves function in a pig lung transplant model. Gene Ther. 2004, 11, 1786–1796. [Google Scholar] [CrossRef]
- Hartog, G.D.; Savelkoul, H.F.J.; Schoemaker, R.; Tijhaar, E.; Westphal, A.H.; de Ruiter, T.; van de Weg-Schrijver, E.; van Neerven, R.J.J. Modulation of Human Immune Responses by Bovine Interleukin-10. PLoS ONE 2011, 6, e18188. [Google Scholar] [CrossRef]
- Kondo, T.; Kobayashi, J.; Saitoh, T.; Maruyama, K.; Ishii, K.J.; Barber, G.N.; Komatsu, K.; Akira, S.; Kawai, T. DNA damage sensor MRE11 recognizes cytosolic double-stranded DNA and induces type i interferon by regulating STING trafficking. Proc. Natl. Acad. Sci. USA 2013, 110, 2969–2974. [Google Scholar] [CrossRef]
- Katashiba, Y.; Miyamoto, R.; Hyo, A.; Shimamoto, K.; Murakami, N.; Ogata, M.; Amakawa, R.; Inaba, M.; Nomura, S.; Fukuhara, S.; et al. Interferon-α and interleukin-12 are induced, respectively, by double-stranded DNA and single-stranded RNA in human myeloid dendritic cells. Immunology 2011, 132, 165–173. [Google Scholar] [CrossRef]
- Seldon, P.M.; Barnes, P.J.; Giembycz, M.A. Interleukin-10 does not mediate the inhibitory effect of PDE-4 inhibitors and other cAMP-elevating drugs on lipopolysaccharide-induced tumors necrosis factor-α generation from human peripheral blood monocytes. Cell Biochem. Biophys. 1998, 29, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Assadiasl, S.; Mooney, N.; Nicknam, M.H. Cytokines in Liver Transplantation. Cytokine 2021, 148, 155705. [Google Scholar] [CrossRef]
- Saxton, R.A.; Tsutsumi, N.; Su, L.L.; Abhiraman, G.C.; Mohan, K.; Henneberg, L.T.; Aduri, N.G.; Gati, C.; Garcia, K.C. Structure-based decoupling of the pro- and anti-inflammatory functions of interleukin-10. Science 2021, 371, 1222. [Google Scholar] [CrossRef]
- Nagata, K.; Nishiyama, C. IL-10 in Mast Cell-Mediated Immune Responses: Anti-Inflammatory and Proinflammatory Roles. Int. J. Mol. Sci. 2021, 22, 4972. [Google Scholar] [CrossRef]
- Lu, L.; Barbi, J.; Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703–717. [Google Scholar] [CrossRef]
- Whitehouse, G.; Gray, E.; Mastoridis, S.; Merritt, E.; Kodela, E.; Yang, J.H.M.; Danger, R.; Mairal, M.; Christakoudi, S.; Lozano, J.J.; et al. IL-2 therapy restores regulatory T-cell dysfunction induced by calcineurin inhibitors. Proc. Natl. Acad. Sci. USA 2017, 114, 7083–7088. [Google Scholar] [CrossRef]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of Interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Liu, M.; Nixon, B.G.; Kang, D.; Toure, A.; Bivona, M.; Li, M.O. Foxp3-independent mechanism by which TGF-β controls peripheral T cell tolerance. Proc. Natl. Acad. Sci. USA 2017, 114, E7536–E7544. [Google Scholar] [CrossRef] [PubMed]
- Zorn, E.; Nelson, E.A.; Mohseni, M.; Porcheray, F.; Kim, H.; Litsa, D.; Bellucci, R.; Raderschall, E.; Canning, C.; Soiffer, R.J.; et al. IL-2 regulates FOXP3 expression in human CD4+CD25+ regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo. Blood 2006, 108, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Skartsis, N.; Peng, Y.; Ferreira, L.M.R.; Nguyen, V.; Ronin, E.; Muller, Y.D.; Vincenti, F.; Tang, Q. IL-6 and TNFα Drive Extensive Proliferation of Human Tregs Without Compromising Their Lineage Stability or Function. Front. Immunol. 2021, 12, 783282. [Google Scholar] [CrossRef]
- Bagheri, Y.; Babaha, F.; Falak, R.; Yazdani, R.; Azizi, G.; Sadri, M.; Abolhassani, H.; Shekarabi, M.; Aghamohammadi, A. IL-10 induces TGF-β secretion, TGF-β receptor II upregulation, and IgA secretion in B cells. Eur. Cytokine Netw. 2019, 30, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Santner-Nanan, B.; Hu, M.; Skarratt, K.; Lee, C.H.; Stormon, M.; Wong, M.; Fuller, S.J.; Nanan, R. IL-10 Potentiates Differentiation of Human Induced Regulatory T Cells via STAT3 and Foxo1. J. Immunol. 2015, 195, 3665–3674. [Google Scholar] [CrossRef] [PubMed]
- Fondevila, C.; Hessheimer, A.J.; Flores, E.; Vendrell, M.; Muñoz, J.; Escobar, B.; Calatayud, D.; Taurá, P.; Fuster, J.; García-Valdecasas, J.C. Step-by-step guide for a simplified model of porcine orthotopic liver transplant. J. Surg. Res. 2011, 167, e39–e45. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Creatinine (mg/dL) | Lactate (mmol/L) | Hemoglobin (g/dL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | IL-10-Treated | Control | IL-10-Treated | Control | IL-10-Treated | Control | IL-10-Treated | ||||||||||
| Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | ||
| Pre-surgery | 7.39 | 0.04 | 7.06 | 0.50 | 1.20 | 0.14 | ND | ND | 1.18 | 0.32 | 0.57 | 0.00 | 8.85 | 1.91 | ND | 0.00 | |
| 1 d | 7.49 | 0.05 | 7.44 | 0.07 | 1.70 | 0.10 | 1.33 | 0.15 | 1.15 | 0.53 | 1.44 | 0.21 | 8.60 | 1.39 | 9.14 | 0.93 | |
| 2 d | 7.52 | 0.03 | 7.48 | 0.01 | 1.25 | 0.07 | 1.15 | 0.17 | 0.79 | 0.17 | 0.72 | 0.18 | 7.80 | 0.00 | 8.30 | 1.04 | |
| 4 d | 7.47 | 0.05 | 7.48 | 0.01 | 1.13 | 0.10 | 1.17 | 0.06 | 0.77 | 0.19 | 1.13 | 0.32 | 8.30 | 0.81 | 9.88 | 1.96 | |
| 7 d | 7.49 | 0.03 | 7.48 | 0.04 | 1.15 | 0.31 | 1.05 | 0.06 | 1.01 | 0.48 | 1.17 | 0.34 | 7.93 | 1.12 | 10.40 | 2.08 | |
| 9–10 d | 7.42 | 0.10 | 7.41 | 0.08 | 1.07 | 0.21 | 0.97 | 0.15 | 1.84 | 1.44 | 1.27 | 0.73 | 8.27 | 0.50 | 9.63 | 1.40 | |
| Bilirubin (mg/dL) | Glucose (mg/dL) | Albumin (g/L) | |||||||||||||||
| Control | IL-10-Treated | Control | IL-10-Treated | Control | IL-10-Treated | ||||||||||||
| Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | Average | SD | ||||||
| 1 d | 0.46 | 0.18 | 0.33 | 0.13 | 88.25 | 9.50 | 104.60 | 31.39 | 25.50 | 5.80 | 25.80 | 2.86 | |||||
| 2 d | 0.35 | 0.24 | 0.47 | 0.18 | 101.00 | 37.42 | 88.40 | 11.63 | 24.20 | 1.92 | 28.20 | 2.28 | |||||
| 4 d | 0.50 | 0.33 | 0.64 | 0.36 | 87.00 | 10.93 | 63.60 | 34.34 | 24.60 | 2.51 | 26.20 | 2.05 | |||||
| 7 d | 0.54 | 0.28 | 0.84 | 0.64 | 83.60 | 9.34 | 52.00 | 37.40 | 23.80 | 3.56 | 25.20 | 2.77 | |||||
| 9–10 d | 0.78 | 0.55 | 0.91 | 0.65 | 78.67 | 17.16 | 69.50 | 20.86 | 25.67 | 2.52 | 28.75 | 5.25 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sendra, L.; Navasquillo, M.; Montalvá, E.M.; Calatayud, D.; Pérez-Rojas, J.; Maupoey, J.; Carmona, P.; Zarragoikoetxea, I.; López-Cantero, M.; Herrero, M.J.; et al. Safe Procedure for Efficient Hydrodynamic Gene Transfer to Isolated Porcine Liver in Transplantation. Int. J. Mol. Sci. 2024, 25, 1491. https://doi.org/10.3390/ijms25031491
Sendra L, Navasquillo M, Montalvá EM, Calatayud D, Pérez-Rojas J, Maupoey J, Carmona P, Zarragoikoetxea I, López-Cantero M, Herrero MJ, et al. Safe Procedure for Efficient Hydrodynamic Gene Transfer to Isolated Porcine Liver in Transplantation. International Journal of Molecular Sciences. 2024; 25(3):1491. https://doi.org/10.3390/ijms25031491
Chicago/Turabian StyleSendra, Luis, Mireia Navasquillo, Eva M. Montalvá, David Calatayud, Judith Pérez-Rojas, Javier Maupoey, Paula Carmona, Iratxe Zarragoikoetxea, Marta López-Cantero, María José Herrero, and et al. 2024. "Safe Procedure for Efficient Hydrodynamic Gene Transfer to Isolated Porcine Liver in Transplantation" International Journal of Molecular Sciences 25, no. 3: 1491. https://doi.org/10.3390/ijms25031491
APA StyleSendra, L., Navasquillo, M., Montalvá, E. M., Calatayud, D., Pérez-Rojas, J., Maupoey, J., Carmona, P., Zarragoikoetxea, I., López-Cantero, M., Herrero, M. J., Aliño, S. F., & López-Andújar, R. (2024). Safe Procedure for Efficient Hydrodynamic Gene Transfer to Isolated Porcine Liver in Transplantation. International Journal of Molecular Sciences, 25(3), 1491. https://doi.org/10.3390/ijms25031491