


Differential Gene Expression following DHX36/G4R1 Knockout Is Associated with G-Quadruplex Content and Cancer

 , , ,
, , ,
Abstract
:1. Introduction
1.1. Evidence for a Role of G4s in Gene Regulation
1.2. Potential Effects of G4s on Transcription
1.3. G4 Binding and Stabilization by Ligands Can Alter Gene Expression
1.4. Helicases Modulate G4 Effects on Expression
2. Results
2.1. Data Features
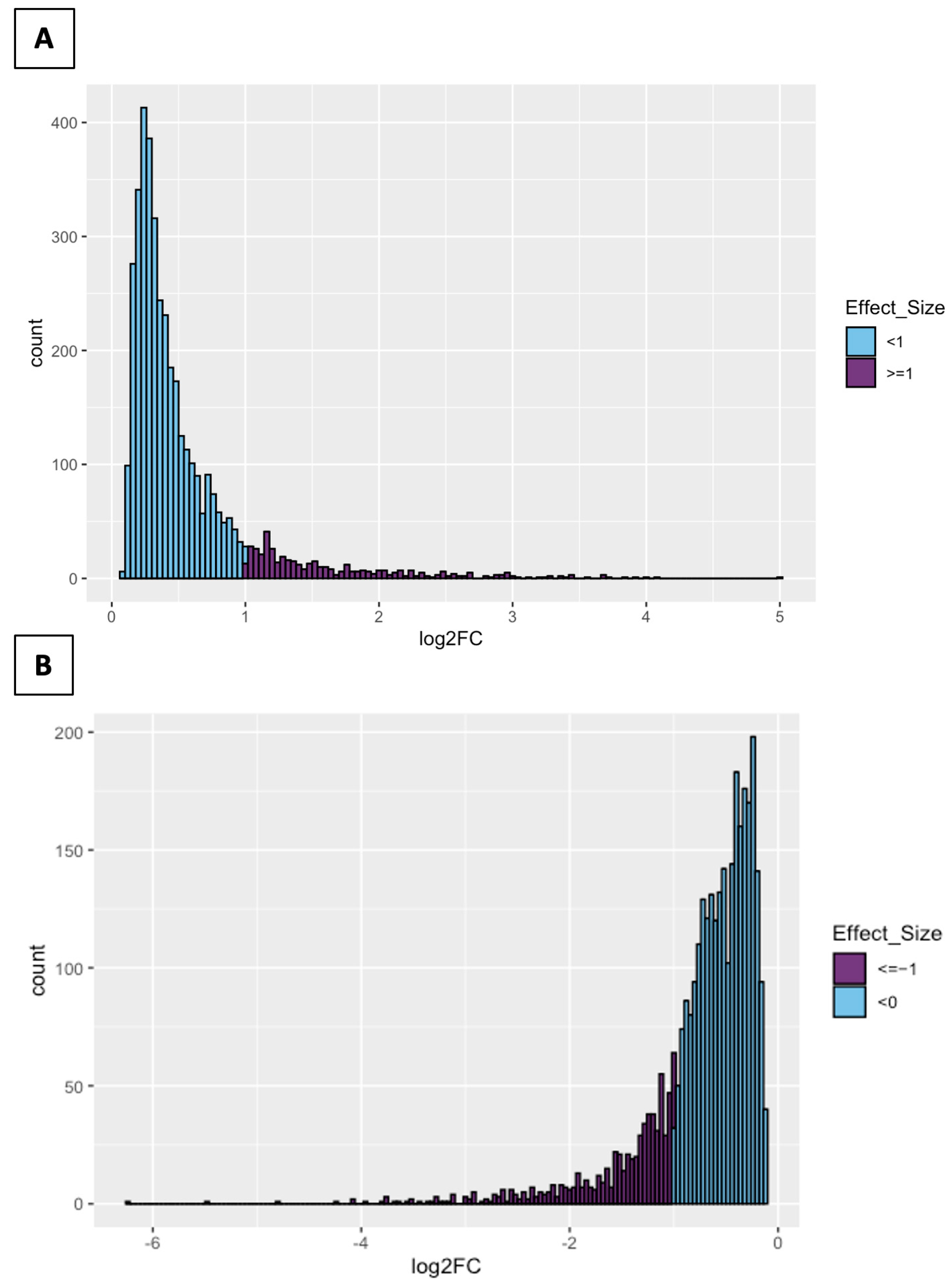
2.2. Differential Expression
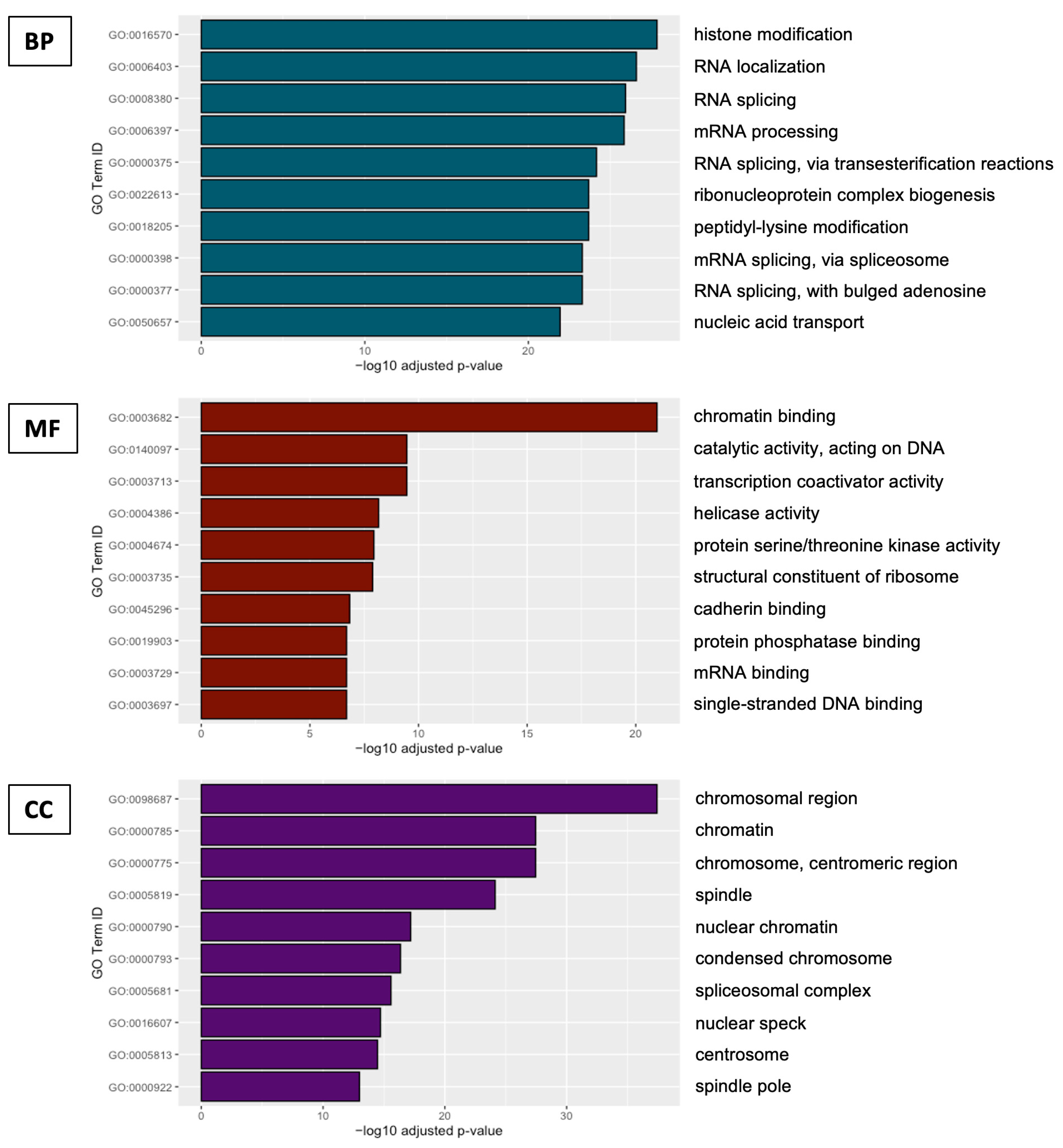
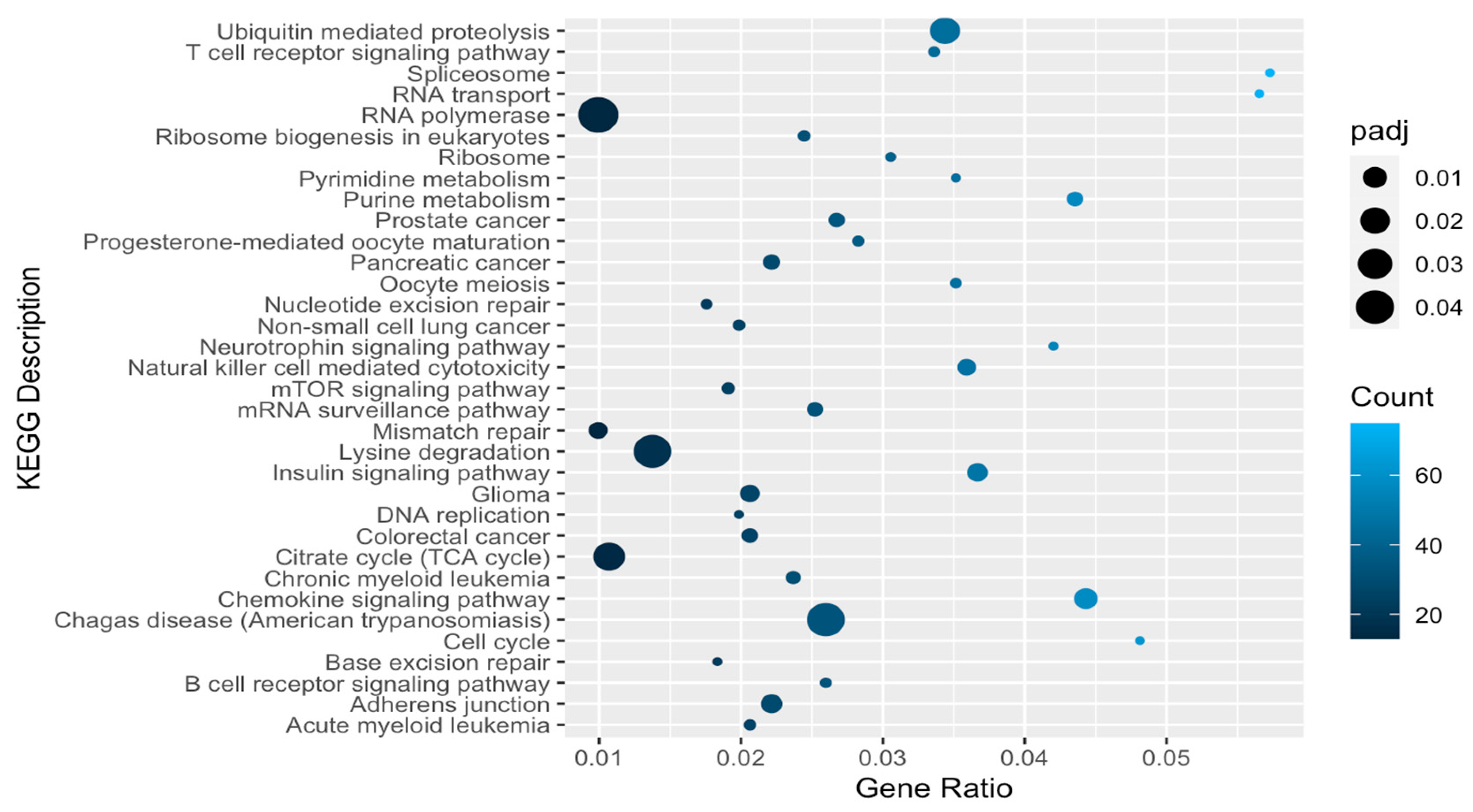
2.3. Functional Genomics
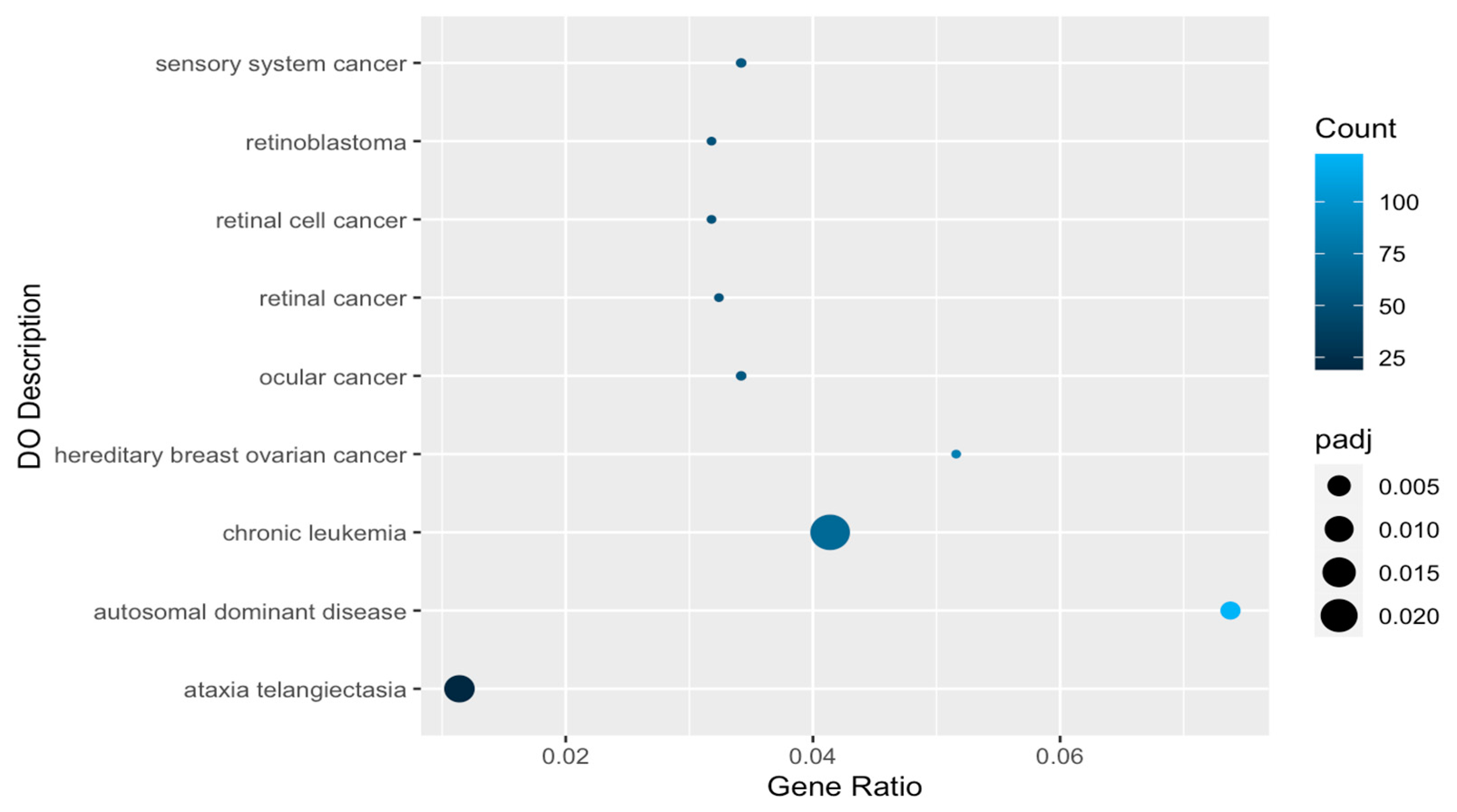
2.4. Disease Associations of Differentially Expressed Genes
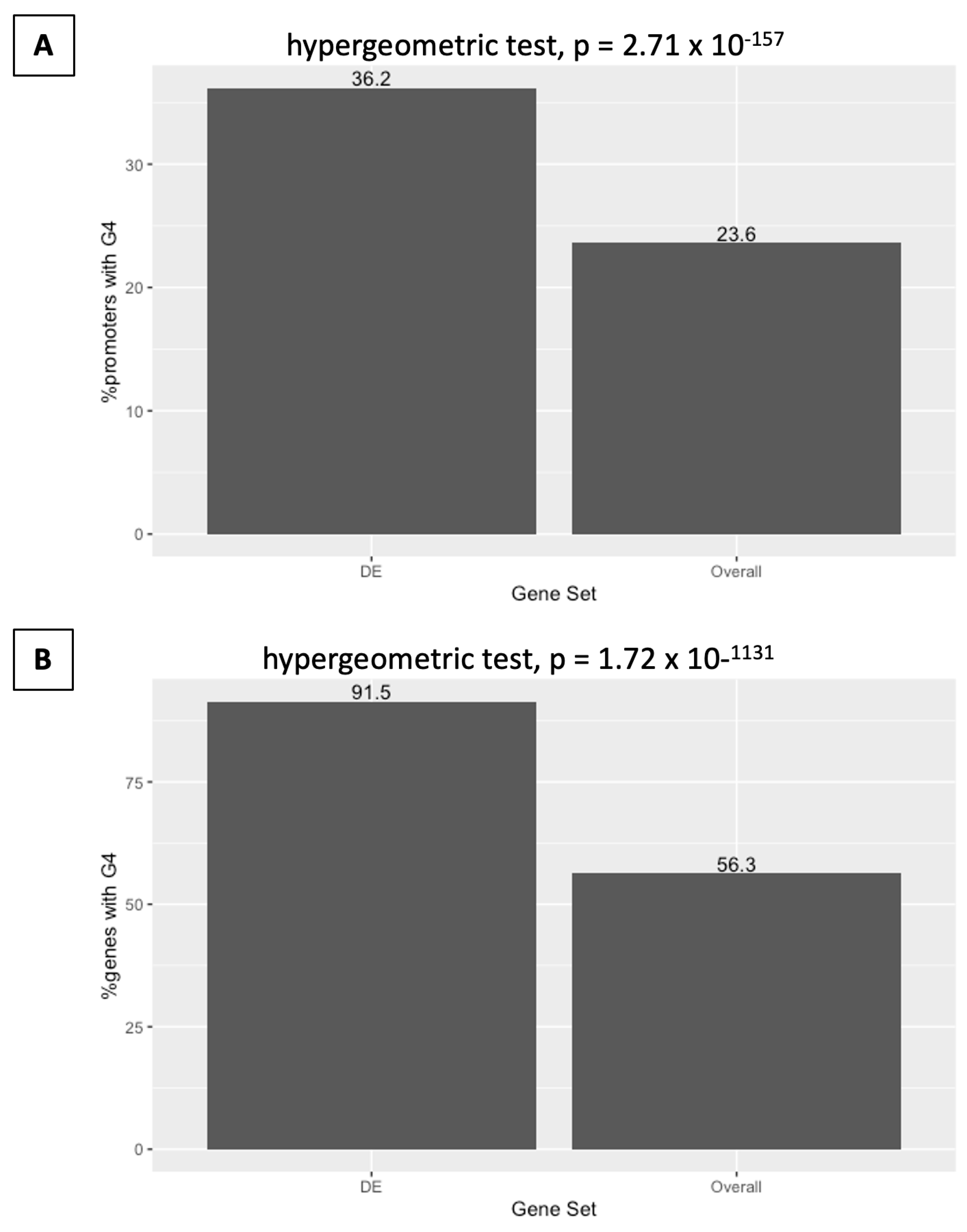
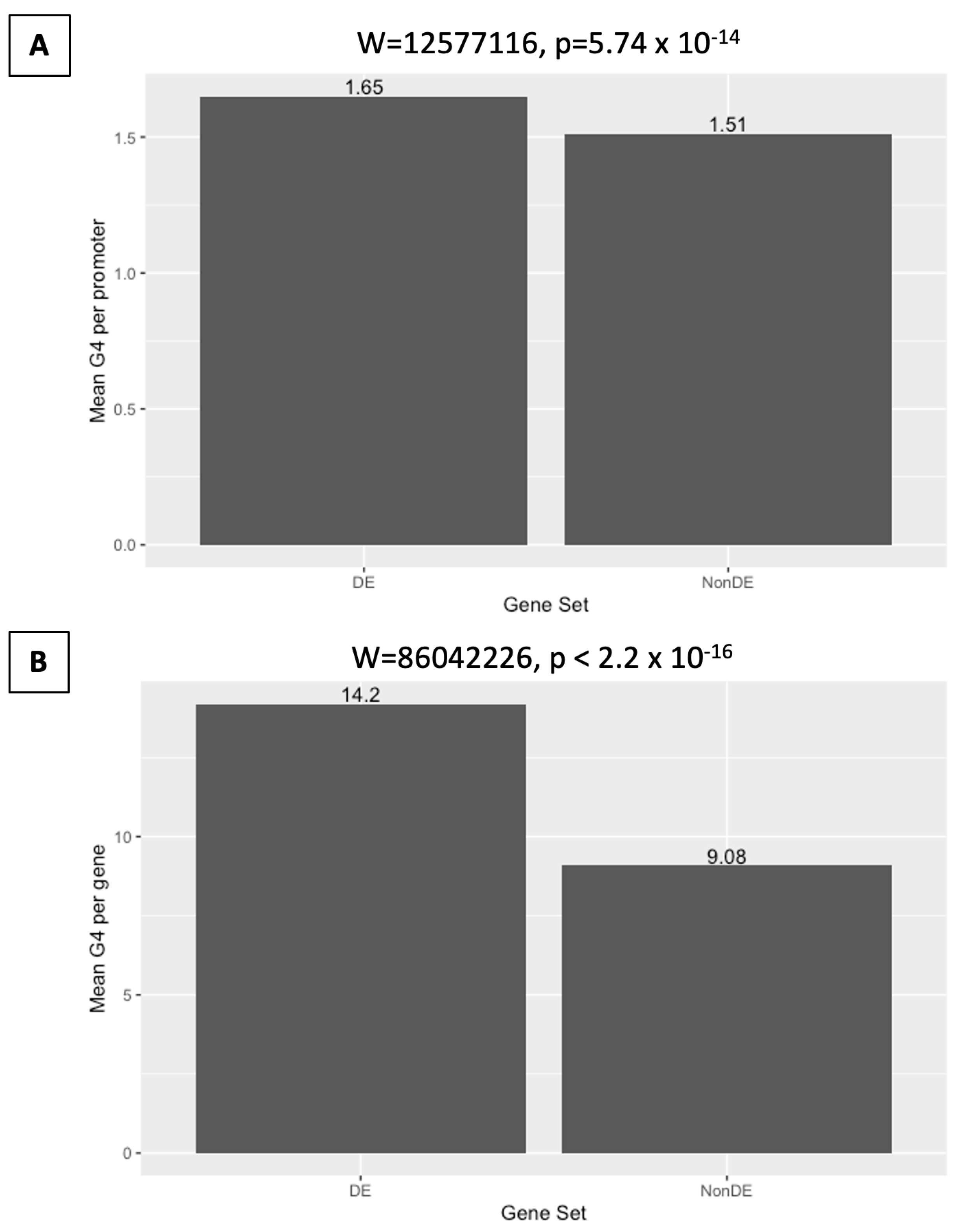
2.5. Differential Expression on DHX36 Knockout Is Associated with G-Quadruplex Content in Promoters
2.6. Differential Expression on DHX36 Knockout Is Associated with G-Quadruplex Content in Gene Regions
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eddy, J.; Maizels, N. Gene function correlates with potential for G4 DNA formation in the human genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef]
- Maizels, N.; Gray, L.T. The G4 genome. PLoS Genet. 2013, 9, e1003468. [Google Scholar] [CrossRef]
- Mendoza, O.; Bourdoncle, A.; Boule, J.B.; Brosh, R.M., Jr.; Mergny, J.L. G-quadruplexes and helicases. Nucleic Acids Res. 2016, 44, 1989–2006. [Google Scholar] [CrossRef]
- Maizels, N. G4 motifs in human genes. Ann. N. Y. Acad. Sci. 2012, 1267, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Kim, N.; Kumari, M.; Verma, S.; Sharma, T.K.; Yadav, V.; Kumar, A. G-Quadruplex Structures in Bacteria: Biological Relevance and Potential as an Antimicrobial Target. J. Bacteriol. 2021, 203. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Smaldino, P.J.; Routh, E.D.; Kim, J.H.; Giri, B.; Creacy, S.D.; Hantgan, R.R.; Akman, S.A.; Vaughn, J.P. Mutational Dissection of Telomeric DNA Binding Requirements of G4 Resolvase 1 Shows that G4-Structure and Certain 3′-Tail Sequences Are Sufficient for Tight and Complete Binding. PLoS ONE 2015, 10, e0132668. [Google Scholar] [CrossRef]
- Bochman, M.L.; Paeschke, K.; Zakian, V.A. DNA secondary structures: Stability and function of G-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Harrington, C.; Lan, Y.; Akman, S.A. The identification and characterization of a G4-DNA resolvase activity. J. Biol. Chem. 1997, 272, 24631–24636. [Google Scholar] [CrossRef]
- Vaughn, J.P.; Creacy, S.D.; Routh, E.D.; Joyner-Butt, C.; Jenkins, G.S.; Pauli, S.; Nagamine, Y.; Akman, S.A. The DEXH protein product of the DHX36 gene is the major source of tetramolecular quadruplex G4-DNA resolving activity in HeLa cell lysates. J. Biol. Chem. 2005, 280, 38117–38120. [Google Scholar] [CrossRef] [PubMed]
- Sauer, M.; Paeschke, K. G-quadruplex unwinding helicases and their function in vivo. Biochem. Soc. Trans. 2017, 45, 1173–1182. [Google Scholar] [CrossRef]
- Halder, K.; Halder, R.; Chowdhury, S. Genome-wide analysis predicts DNA structural motifs as nucleosome exclusion signals. Mol. Biosyst. 2009, 5, 1703–1712. [Google Scholar] [CrossRef]
- Wong, H.M.; Huppert, J.L. Stable G-quadruplexes are found outside nucleosome-bound regions. Mol. Biosyst. 2009, 5, 1713–1719. [Google Scholar] [CrossRef]
- Andersson, R.; Sandelin, A. Determinants of enhancer and promoter activities of regulatory elements. Nat. Rev. Genet. 2020, 21, 71–87. [Google Scholar] [CrossRef]
- Zheng, K.W.; Zhang, J.Y.; He, Y.D.; Gong, J.Y.; Wen, C.J.; Chen, J.N.; Hao, Y.H.; Zhao, Y.; Tan, Z. Detection of genomic G-quadruplexes in living cells using a small artificial protein. Nucleic Acids Res. 2020, 48, 11706–11720. [Google Scholar] [CrossRef]
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef]
- Huppert, J.L.; Balasubramanian, S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef]
- Rawal, P.; Kummarasetti, V.B.; Ravindran, J.; Kumar, N.; Halder, K.; Sharma, R.; Mukerji, M.; Das, S.K.; Chowdhury, S. Genome-wide prediction of G4 DNA as regulatory motifs: Role in Escherichia coli global regulation. Genome Res. 2006, 16, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Hershman, S.G.; Chen, Q.; Lee, J.Y.; Kozak, M.L.; Yue, P.; Wang, L.S.; Johnson, F.B. Genomic distribution and functional analyses of potential G-quadruplex-forming sequences in Saccharomyces cerevisiae. Nucleic Acids Res. 2008, 36, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.K.; Neidle, S. The relationship of potential G-quadruplex sequences in cis-upstream regions of the human genome to SP1-binding elements. Nucleic Acids Res. 2008, 36, 2700–2704. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Gertz, E.M.; Wojtowicz, D.; Zhabinskaya, D.; Levens, D.; Benham, C.J.; Schäffer, A.A.; Przytycka, T.M. Potential non-B DNA regions in the human genome are associated with higher rates of nucleotide mutation and expression variation. Nucleic Acids Res. 2014, 42, 12367–12379. [Google Scholar] [CrossRef] [PubMed]
- Fernando, H.; Sewitz, S.; Darot, J.; Tavaré, S.; Huppert, J.L.; Balasubramanian, S. Genome-wide analysis of a G-quadruplex-specific single-chain antibody that regulates gene expression. Nucleic Acids Res. 2009, 37, 6716–6722. [Google Scholar] [CrossRef] [PubMed]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.; Zhang, R.; Xiao, K.; Yang, J.; Sun, X. G-Quadruplex-Binding Proteins: Promising Targets for Drug Design. Biomolecules 2022, 12, 648. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, J.; Cuesta, S.M.; Adhikari, S.; Hänsel-Hertsch, R.; Tannahill, D.; Balasubramanian, S. G-quadruplexes are transcription factor binding hubs in human chromatin. Genome Biol. 2021, 22, 117. [Google Scholar] [CrossRef]
- Hurley, L.H.; Von Hoff, D.D.; Siddiqui-Jain, A.; Yang, D. Drug targeting of the c-MYC promoter to repress gene expression via a G-quadruplex silencer element. Semin. Oncol. 2006, 33, 498–512. [Google Scholar] [CrossRef]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef]
- Lam, E.Y.; Beraldi, D.; Tannahill, D.; Balasubramanian, S. G-quadruplex structures are stable and detectable in human genomic DNA. Nat. Commun. 2013, 4, 1796. [Google Scholar] [CrossRef]
- Fry, R.C.; Sambandan, T.G.; Rha, C. DNA damage and stress transcripts in Saccharomyces cerevisiae mutant sgs1. Mech. Ageing Dev. 2003, 124, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.H.; Tang, W.; Robles, A.I.; Beyer, R.P.; Gray, L.T.; Welsh, J.A.; Schetter, A.J.; Kumamoto, K.; Wang, X.W.; Hickson, I.D.; et al. Regulation of gene expression by the BLM helicase correlates with the presence of G-quadruplex DNA motifs. Proc. Natl. Acad. Sci. USA 2014, 111, 9905–9910. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.H.; Liu, X.; Canterel-Thouennon, L.; Li, L.; Edmonson, C.; Rennert, O.M. Telomerase protects werner syndrome lineage-specific stem cells from premature aging. Stem Cell Rep. 2014, 2, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Creacy, S.D.; Routh, E.D.; Iwamoto, F.; Nagamine, Y.; Akman, S.A.; Vaughn, J.P. G4 resolvase 1 binds both DNA and RNA tetramolecular quadruplex with high affinity and is the major source of tetramolecular quadruplex G4-DNA and G4-RNA resolving activity in HeLa cell lysates. J. Biol. Chem. 2008, 283, 34626–34634. [Google Scholar] [CrossRef]
- Giri, B.; Smaldino, P.J.; Thys, R.G.; Creacy, S.D.; Routh, E.D.; Hantgan, R.R.; Lattmann, S.; Nagamine, Y.; Akman, S.A.; Vaughn, J.P. G4 resolvase 1 tightly binds and unwinds unimolecular G4-DNA. Nucleic Acids Res. 2011, 39, 7161–7178. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Tippana, R.; Demeshkina, N.A.; Murat, P.; Balasubramanian, S.; Myong, S.; Ferre-D’Amare, A.R. Structural basis of G-quadruplex unfolding by the DEAH/RHA helicase DHX36. Nature 2018, 558, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Smaldino, P.J.; Zhang, Q.; Miller, L.D.; Cao, P.; Stadelman, K.; Wan, M.; Giri, B.; Lei, M.; Nagamine, Y.; et al. Yin Yang 1 contains G-quadruplex structures in its promoter and 5′-UTR and its expression is modulated by G4 resolvase 1. Nucleic Acids Res. 2012, 40, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Sauer, M.; Juranek, S.A.; Marks, J.; De Magis, A.; Kazemier, H.G.; Hilbig, D.; Benhalevy, D.; Wang, X.; Hafner, M.; Paeschke, K. DHX36 prevents the accumulation of translationally inactive mRNAs with G4-structures in untranslated regions. Nat. Commun. 2019, 10, 2421. [Google Scholar] [CrossRef]
- Murat, P.; Marsico, G.; Herdy, B.; Ghanbarian, A.T.; Portella, G.; Balasubramanian, S. RNA G-quadruplexes at upstream open reading frames cause DHX36- and DHX9-dependent translation of human mRNAs. Genome Biol. 2018, 19, 229. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, X.; Wang, K.; Zhang, B.; Qiu, S. Corrigendum: The Cellular Functions and Molecular Mechanisms of G-Quadruplex Unwinding Helicases in Humans. Front. Mol. Biosci. 2022, 8, 833430. [Google Scholar] [CrossRef]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Res 2015, 4, 1521. [Google Scholar] [CrossRef]
- Guo, J.U.; Bartel, D.P. RNA G-quadruplexes are globally unfolded in eukaryotic cells and depleted in bacteria. Science 2016, 353, aaf5371. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Sandwith, S.N.; Green, K.M.; Chambers, A.E.; Krans, A.; Raimer, H.M.; Sharlow, M.E.; Reisinger, M.A.; Richardson, A.E.; Routh, E.D.; et al. The RNA helicase DHX36-G4R1 modulates C9orf72 GGGGCC hexanucleotide repeat-associated translation. J. Biol. Chem. 2021, 297, 100914. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Fulton, D.L.; Sundararajan, S.; Badis, G.; Hughes, T.R.; Wasserman, W.W.; Roach, J.C.; Sladek, R. TFCat: The curated catalog of mouse and human transcription factors. Genome Biol. 2009, 10, R29. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Treatment | Raw Reads | Filtered Reads | % Q20 | % Uniquely Mapped | % Multi-Mapping |
|---|---|---|---|---|---|---|
| A11_1 | DHX36 KO | 25,210,874 | 24,430,301 | 98.11 | 93.58 | 2.24 |
| A11_2 | DHX36 KO | 28,718,452 | 27,622,299 | 98.05 | 93.26 | 2.26 |
| A11_3 | DHX36 KO | 28,926,801 | 28,216,080 | 98.16 | 93.87 | 2.14 |
| A28_1 | Wild-type | 27,734,054 | 27,130,315 | 98.12 | 93.59 | 2.14 |
| A28_2 | Wild-type | 26,521,784 | 25,939,496 | 98.13 | 93.87 | 2.14 |
| A28_3 | Wild-type | 28,081,209 | 27,206,143 | 98.23 | 93.56 | 2.23 |
| ID | Description | Gene Ratio | Background Ratio | Raw p-Value | Adjusted p-Value |
|---|---|---|---|---|---|
| DOID:4645 | retinal cancer | 54/1667 | 113/7405 | 2.29 × 10−9 | 6.79 × 10−7 |
| DOID:5683 | hereditary breast ovarian cancer | 86/1667 | 214/7405 | 3.04 × 10−9 | 6.79 × 10−7 |
| DOID:768 | retinoblastoma | 53/1667 | 111/7405 | 3.34 × 10−9 | 6.79 × 10−7 |
| DOID:771 | retinal cell cancer | 53/1667 | 111/7405 | 3.34 × 10−9 | 6.79 × 10−7 |
| DOID:0060116 | sensory system cancer | 57/1667 | 134/7405 | 1.49 × 10−7 | 2.01 × 10−5 |
| DOID:2174 | ocular cancer | 57/1667 | 134/7405 | 1.49 × 10−7 | 2.01 × 10−5 |
| DOID:0050736 | autosomal dominant disease | 123/1667 | 394/7405 | 2.56 × 10−5 | 0.00297116 |
| DOID:12704 | ataxia telangiectasia | 19/1667 | 37/7405 | 0.0001142 | 0.01159172 |
| DOID:1036 | ataxia telangiectasia | 69/1667 | 209/7405 | 0.00026725 | 0.02411196 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gumina, J.M.; Richardson, A.E.; Shojiv, M.H.; Chambers, A.E.; Sandwith, S.N.; Reisinger, M.A.; Karns, T.J.; Osborne, T.L.; Alashi, H.N.; Anderson, Q.T.; et al. Differential Gene Expression following DHX36/G4R1 Knockout Is Associated with G-Quadruplex Content and Cancer. Int. J. Mol. Sci. 2024, 25, 1753. https://doi.org/10.3390/ijms25031753
Gumina JM, Richardson AE, Shojiv MH, Chambers AE, Sandwith SN, Reisinger MA, Karns TJ, Osborne TL, Alashi HN, Anderson QT, et al. Differential Gene Expression following DHX36/G4R1 Knockout Is Associated with G-Quadruplex Content and Cancer. International Journal of Molecular Sciences. 2024; 25(3):1753. https://doi.org/10.3390/ijms25031753
Chicago/Turabian StyleGumina, Joseph M., Adam E. Richardson, Mahmudul Hasan Shojiv, Antonio E. Chambers, Siara N. Sandwith, Michael A. Reisinger, Taylor J. Karns, Tyler L. Osborne, Hasna N. Alashi, Quinn T. Anderson, and et al. 2024. "Differential Gene Expression following DHX36/G4R1 Knockout Is Associated with G-Quadruplex Content and Cancer" International Journal of Molecular Sciences 25, no. 3: 1753. https://doi.org/10.3390/ijms25031753
APA StyleGumina, J. M., Richardson, A. E., Shojiv, M. H., Chambers, A. E., Sandwith, S. N., Reisinger, M. A., Karns, T. J., Osborne, T. L., Alashi, H. N., Anderson, Q. T., Sharlow, M. E., Seiler, D. C., Rogers, E. M., Bartosik, A. R., Smaldino, M. A., Vaughn, J. P., Wang, Y. -H., Smaldino, P. J., & Haney, R. A. (2024). Differential Gene Expression following DHX36/G4R1 Knockout Is Associated with G-Quadruplex Content and Cancer. International Journal of Molecular Sciences, 25(3), 1753. https://doi.org/10.3390/ijms25031753