
OsACA9, an Autoinhibited Ca2+-ATPase, Synergically Regulates Disease Resistance and Leaf Senescence in Rice

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Evolution Analysis and Expression Profile Suggest the Potential Role of OsACA9 in Various Biological Processes
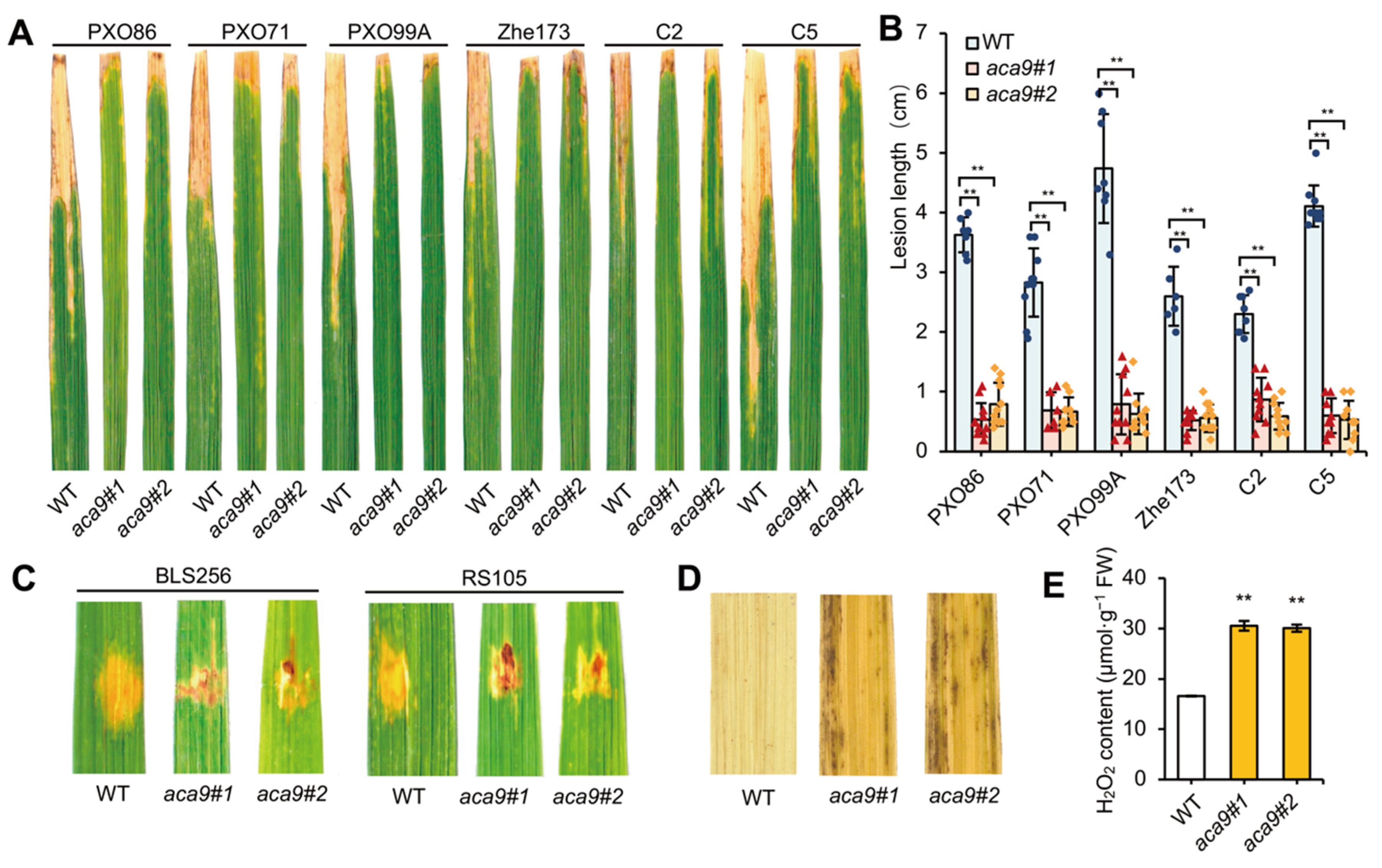
2.2. Knockout of OsACA9 Broadly Enhanced the Bacterial Disease Resistance of Rice
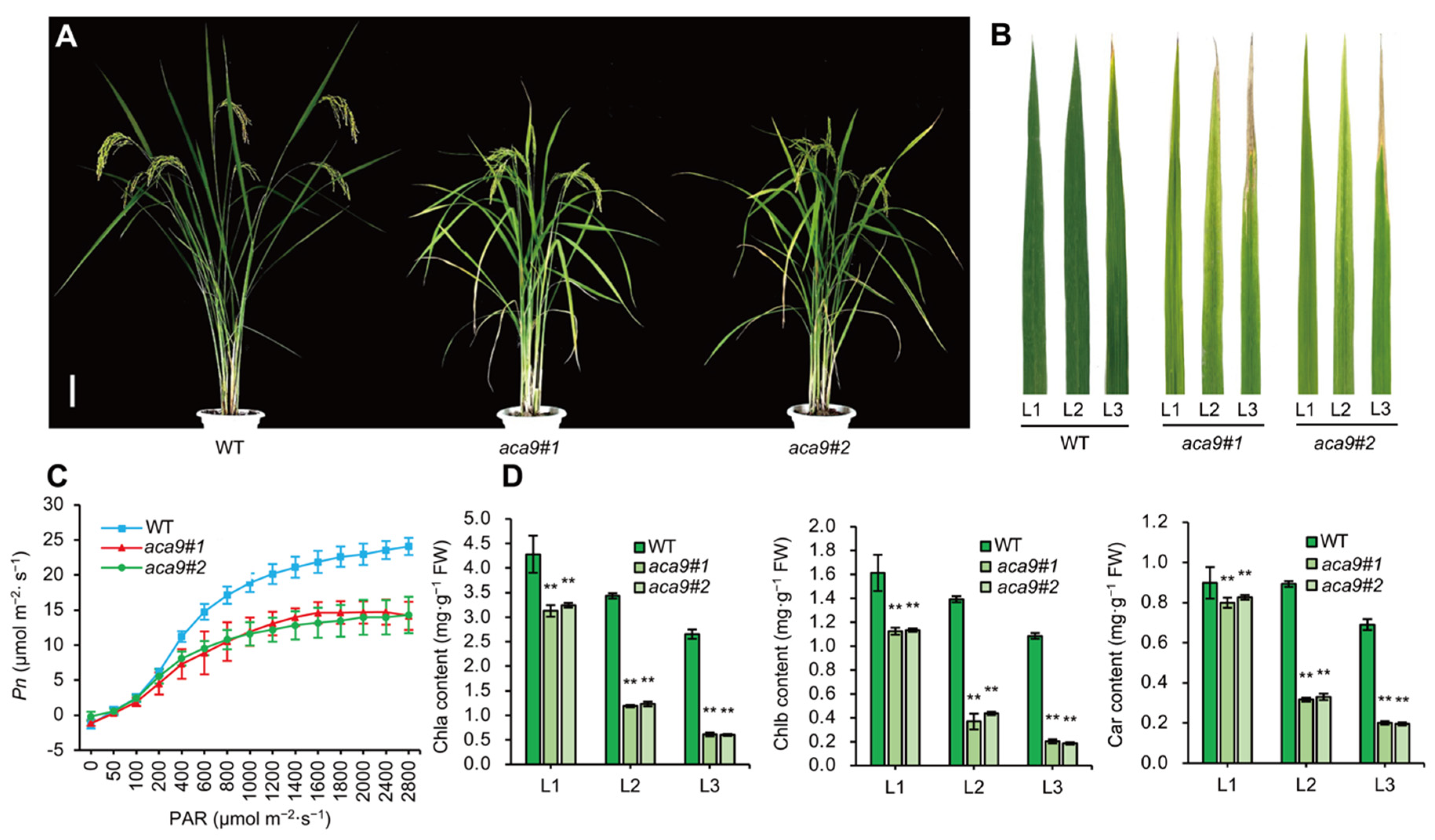
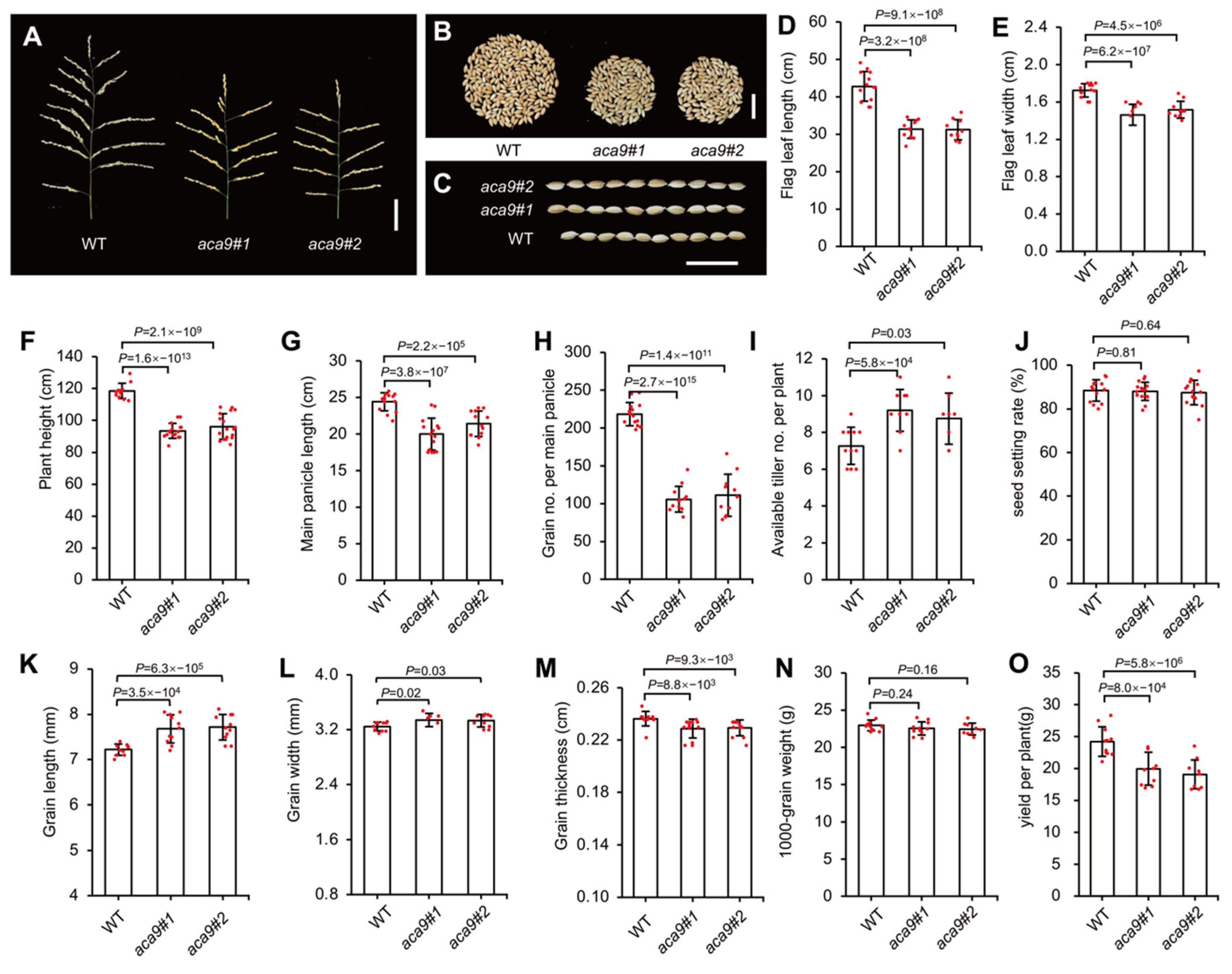
2.3. OsACA9 Regulated the Leaf Senescence and Development of Rice
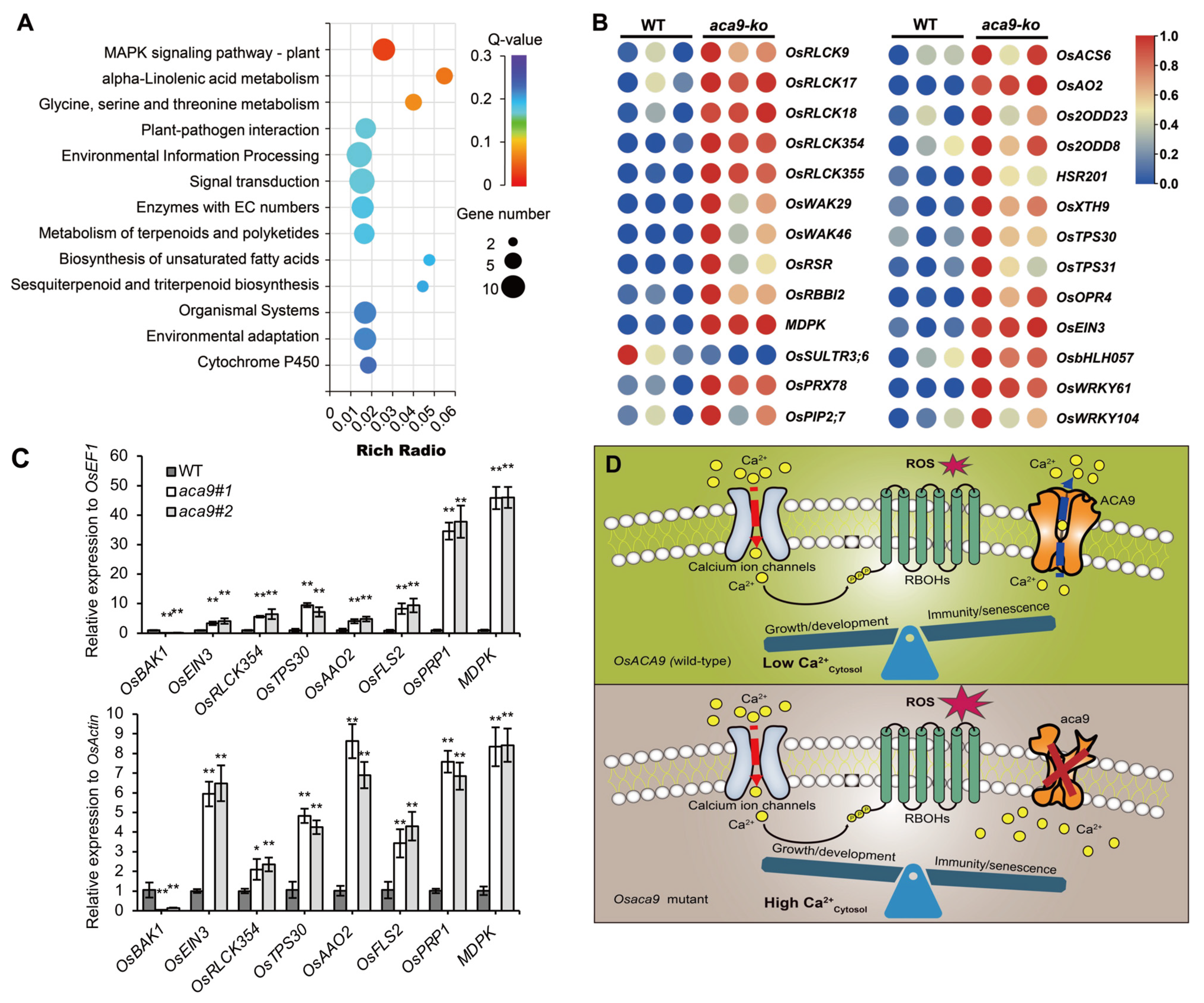
2.4. Transcriptomic Analysis Suggested OsACA9 Involved in MAPK Cascades in PAMP-Triggered Defense Response
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Phylogenetic Analysis of OsACA Gene Family
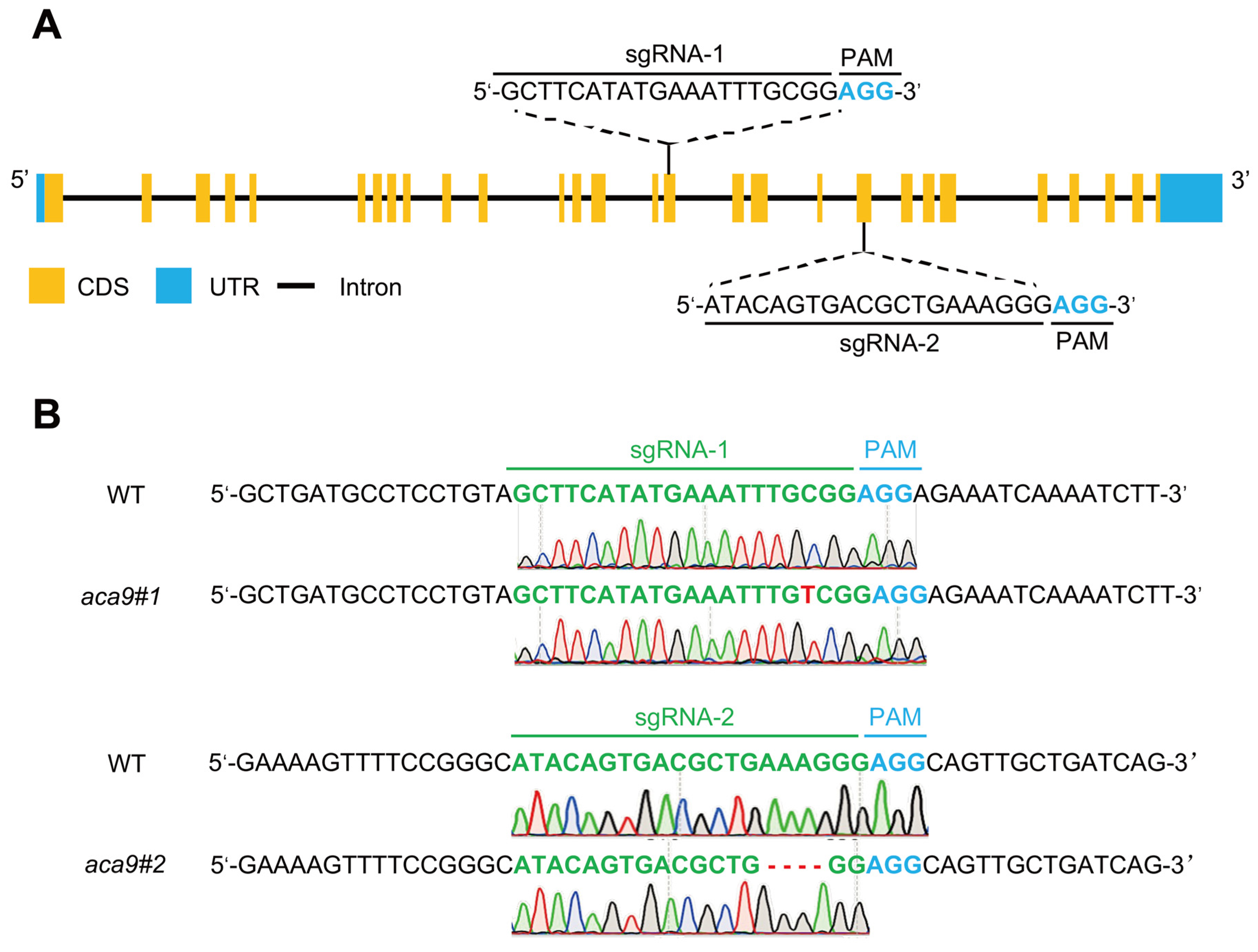
4.3. Construction of OsACA9 Knockout Rice Plants by CRISPR/Cas9
4.4. Assay of Chlorophyll Content and Photosynthesis
4.5. Disease Resistance Identification of Rice Plants
4.6. DAB Staining and H2O2 Assay
4.7. RNA-Seq and Data Analysis
4.8. RNA Extraction and RT-qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium Spikes, Waves and Oscillations in Plant Development and Biotic Interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Köster, P.; DeFalco, T.A.; Zipfel, C. Ca2+ Signals in Plant Immunity. EMBO J. 2022, 41, e110741. [Google Scholar] [CrossRef]
- Upadhyay, S.K. Calcium Channels, OST1 and Stomatal Defence: Current Status and Beyond. Cells 2022, 12, 127. [Google Scholar] [CrossRef]
- Dong, X.; Gifford, M.L.; Su, C. Ca2+ Signatures in Symbiosis: Another Level of Dynamism for This Key Messenger. J. Exp. Bot. 2024, 75, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic Plant Aequorin Reports the Effects of Touch and Cold-Shock and Elicitors on Cytoplasmic Calcium. Nature 1991, 352, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Calcium Signalling in Arabidopsis thaliana Responding to Drought and Salinity. Plant J. 1997, 12, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K. CPK12 and Ca2+-Mediated Hypoxia Signaling. Plant Signal Behav. 2023, 18, 2273593. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Knight, H.; Brandt, S.; Knight, M.R. A History of Stress Alters Drought Calcium Signalling Pathways in Arabidopsis. Plant J. 1998, 16, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Møller, J.V.; Juul, B.; le Maire, M. Structural Organization, Ion Transport, and Energy Transduction of P-Type ATPases. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1996, 1286, 1–51. [Google Scholar] [CrossRef]
- García Bossi, J.; Kumar, K.; Barberini, M.L.; Domínguez, G.D.; Rondón Guerrero, Y.D.C.; Marino-Buslje, C.; Obertello, M.; Muschietti, J.P.; Estevez, J.M. The Role of P-Type IIA and P-Type IIB Ca2+-ATPases in Plant Development and Growth. J. Exp. Bot. 2020, 71, 1239–1248. [Google Scholar] [CrossRef]
- Wdowikowska, A.; Kłobus, G. Plant P-Type ATPases. Postepy Biochem. 2011, 57, 85–91. [Google Scholar]
- Pedersen, C.N.S.; Axelsen, K.B.; Harper, J.F.; Palmgren, M.G. Evolution of Plant P-Type ATPases. Front. Plant Sci. 2012, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Curran, A.C.; Hwang, I.; Corbin, J.; Martinez, S.; Rayle, D.; Sze, H.; Harper, J.F. Autoinhibition of a Calmodulin-Dependent Calcium Pump Involves a Structure in the Stalk That Connects the Transmembrane Domain to the ATPase Catalytic Domain. J. Biol. Chem. 2000, 275, 30301–30308. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Harper, J.F.; Liang, F.; Sze, H. Calmodulin Activation of an Endoplasmic Reticulum-Located Calcium Pump Involves an Interaction with the N-Terminal Autoinhibitory Domain. Plant Physiol. 2000, 122, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Axelsen, K.B.; Palmgren, M.G. Inventory of the Superfamily of P-Type Ion Pumps in Arabidopsis. Plant Physiol. 2001, 126, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Shu, Y.; Chen, X.L.; Wang, A.X. Identification and Analysis of Ca2+-ATPase Gene Family in Solanaceae. Mol. Plant Breed. 2021, 4268–4277. [Google Scholar]
- Jiang, Q.; Wu, X.; Zhang, X.; Ji, Z.; Cao, Y.; Duan, Q.; Huang, J. Genome-Wide Identification and Expression Analysis of AS2 Genes in Brassica Rapa Reveal Their Potential Roles in Abiotic Stress. Int. J. Mol. Sci. 2023, 24, 10534. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, C.; Geng, L.; Chen, Y.; Zhang, L.; Xiang, D.; Liu, P. Identification and Expression Pattern Analysis of RcACA Gene Family in Castor under Abiotic Stresses. Chin. J. Biotechnol. 2023, 39, 2861–2873. [Google Scholar]
- Chen, C.; Zhang, M.; Zhang, M.; Yang, M.; Dai, S.; Meng, Q.; Lv, W.; Zhuang, K. Ethylene-Insensitive 3-LIKE 2 Regulates β-Carotene and Ascorbic Acid Accumulation in Tomatoes during Ripening. Plant Physiol. 2023, 192, 2067–2080. [Google Scholar] [CrossRef]
- Taneja, M.; Upadhyay, S.K. Molecular Characterization and Differential Expression Suggested Diverse Functions of P-Type II Ca2+ ATPases in Triticum aestivum L. BMC Genom. 2018, 19, 389. [Google Scholar] [CrossRef]
- Huang, L.; Berkelman, T.; Franklin, A.E.; Hoffman, N.E. Characterization of a Gene Encoding a Ca(2+)-ATPase-like Protein in the Plastid Envelope. Proc. Natl. Acad. Sci. USA 1993, 90, 10066–10070. [Google Scholar] [CrossRef]
- Rahmati Ishka, M.; Brown, E.; Rosenberg, A.; Romanowsky, S.; Davis, J.A.; Choi, W.-G.; Harper, J.F. Arabidopsis Ca2+-ATPases 1, 2, and 7 in the Endoplasmic Reticulum Contribute to Growth and Pollen Fitness. Plant Physiol. 2021, 185, 1966–1985. [Google Scholar] [CrossRef]
- Schiøtt, M.; Romanowsky, S.M.; Baekgaard, L.; Jakobsen, M.K.; Palmgren, M.G.; Harper, J.F. A Plant Plasma Membrane Ca2+ Pump Is Required for Normal Pollen Tube Growth and Fertilization. Proc. Natl. Acad. Sci. USA 2004, 101, 9502–9507. [Google Scholar] [CrossRef] [PubMed]
- Iwano, M.; Igarashi, M.; Tarutani, Y.; Kaothien-Nakayama, P.; Nakayama, H.; Moriyama, H.; Yakabe, R.; Entani, T.; Shimosato-Asano, H.; Ueki, M.; et al. A Pollen Coat-Inducible Autoinhibited Ca2+-ATPase Expressed in Stigmatic Papilla Cells Is Required for Compatible Pollination in the Brassicaceae. Plant Cell 2014, 26, 636–649. [Google Scholar] [CrossRef] [PubMed]
- George, L.; Romanowsky, S.M.; Harper, J.F.; Sharrock, R.A. The ACA10 Ca2+-ATPase Regulates Adult Vegetative Development and Inflorescence Architecture in Arabidopsis. Plant Physiol. 2008, 146, 716–728. [Google Scholar] [CrossRef] [PubMed]
- Kamrul Huda, K.M.; Yadav, S.; Akhter Banu, M.S.; Trivedi, D.K.; Tuteja, N. Genome-Wide Analysis of Plant-Type II Ca(2+)ATPases Gene Family from Rice and Arabidopsis: Potential Role in Abiotic Stresses. Plant Physiol. Biochem. 2013, 65, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Anil, V.S.; Rajkumar, P.; Kumar, P.; Mathew, M.K. A Plant Ca2+ Pump, ACA2, Relieves Salt Hypersensitivity in Yeast. Modulation of Cytosolic Calcium Signature and Activation of Adaptive Na+ Homeostasis. J. Biol. Chem. 2008, 283, 3497–3506. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.S.; Lee, S.H.; Kim, J.C.; Heo, W.D.; Kim, M.C.; Park, C.Y.; Park, H.C.; Lim, C.O.; Kim, W.B.; Harper, J.F.; et al. Identification of a Calmodulin-Regulated Soybean Ca(2+)-ATPase (SCA1) That Is Located in the Plasma Membrane. Plant Cell 2000, 12, 1393–1407. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Jia, B.; Cui, N.; Wen, Y.; Duanmu, H.; Yu, Q.; Xiao, J.; Sun, X.; Zhu, Y. Functional Characterization of a Glycine Soja Ca(2+)ATPase in Salt-Alkaline Stress Responses. Plant Mol. Biol. 2016, 90, 419–434. [Google Scholar] [CrossRef]
- Qudeimat, E.; Faltusz, A.M.C.; Wheeler, G.; Lang, D.; Holtorf, H.; Brownlee, C.; Reski, R.; Frank, W. A PIIB-Type Ca2+-ATPase Is Essential for Stress Adaptation in Physcomitrella Patens. Proc. Natl. Acad. Sci. USA 2008, 105, 19555–19560. [Google Scholar] [CrossRef]
- Boursiac, Y.; Lee, S.M.; Romanowsky, S.; Blank, R.; Sladek, C.; Chung, W.S.; Harper, J.F. Disruption of the Vacuolar Calcium-ATPases in Arabidopsis Results in the Activation of a Salicylic Acid-Dependent Programmed Cell Death Pathway. Plant Physiol. 2010, 154, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Benschop, J.J.; Mohammed, S.; O’Flaherty, M.; Heck, A.J.R.; Slijper, M.; Menke, F.L.H. Quantitative Phosphoproteomics of Early Elicitor Signaling in Arabidopsis. Mol. Cell Proteom. 2007, 6, 1198–1214. [Google Scholar] [CrossRef]
- Hilleary, R.; Paez-Valencia, J.; Vens, C.S.; Toyota, M.; Palmgren, M.; Gilroy, S. Tonoplast-Localized Ca2+ Pumps Regulate Ca2+ Signals during Pattern-Triggered Immunity in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2020, 117, 18849–18857. [Google Scholar] [CrossRef] [PubMed]
- Frei dit Frey, N.; Mbengue, M.; Kwaaitaal, M.; Nitsch, L.; Altenbach, D.; Häweker, H.; Lozano-Duran, R.; Njo, M.F.; Beeckman, T.; Huettel, B.; et al. Plasma Membrane Calcium ATPases Are Important Components of Receptor-Mediated Signaling in Plant Immune Responses and Development. Plant Physiol. 2012, 159, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.; Tchieu, J.; Sussman, M.R.; Boutry, M.; Palmgren, M.G.; Gribskov, M.; Harper, J.F.; Axelsen, K.B. Genomic Comparison of P-Type ATPase Ion Pumps in Arabidopsis and Rice. Plant Physiol. 2003, 132, 618–628. [Google Scholar] [CrossRef]
- Singh, A.; Kanwar, P.; Yadav, A.K.; Mishra, M.; Jha, S.K.; Baranwal, V.; Pandey, A.; Kapoor, S.; Tyagi, A.K.; Pandey, G.K. Genome-Wide Expressional and Functional Analysis of Calcium Transport Elements during Abiotic Stress and Development in Rice. FEBS J. 2014, 281, 894–915. [Google Scholar] [CrossRef]
- Kan, C.-C.; Chung, T.-Y.; Wu, H.-Y.; Juo, Y.-A.; Hsieh, M.-H. Exogenous Glutamate Rapidly Induces the Expression of Genes Involved in Metabolism and Defense Responses in Rice Roots. BMC Genom. 2017, 18, 186. [Google Scholar] [CrossRef]
- Bang, S.W.; Lee, D.-K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Do Choi, Y.; Suh, J.-W.; Kim, J.-K. Overexpression of OsTF1L, a Rice HD-Zip Transcription Factor, Promotes Lignin Biosynthesis and Stomatal Closure That Improves Drought Tolerance. Plant Biotechnol. J. 2019, 17, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Huda, K.M.K.; Banu, M.S.A.; Garg, B.; Tula, S.; Tuteja, R.; Tuteja, N. OsACA6, a P-Type IIB Ca2+ ATPase Promotes Salinity and Drought Stress Tolerance in Tobacco by ROS Scavenging and Enhancing the Expression of Stress-Responsive Genes. Plant J. 2013, 76, 997–1015. [Google Scholar] [CrossRef]
- Um, T.; Park, T.; Shim, J.S.; Kim, Y.S.; Lee, G.-S.; Choi, I.-Y.; Kim, J.-K.; Seo, J.S.; Park, S.C. Application of Upstream Open Reading Frames (UORFs) Editing for the Development of Stress-Tolerant Crops. Int. J. Mol. Sci. 2021, 22, 3743. [Google Scholar] [CrossRef]
- Manivannan, A.; Ahn, Y.-K. Silicon Regulates Potential Genes Involved in Major Physiological Processes in Plants to Combat Stress. Front. Plant Sci. 2017, 8, 1346. [Google Scholar] [CrossRef]
- Amirbakhtiar, N.; Ismaili, A.; Ghaffari, M.-R.; Mirdar Mansuri, R.; Sanjari, S.; Shobbar, Z.-S. Transcriptome Analysis of Bread Wheat Leaves in Response to Salt Stress. PLoS ONE 2021, 16, e0254189. [Google Scholar]
- Kadota, Y.; Liebrand, T.W.H.; Goto, Y.; Sklenar, J.; Derbyshire, P.; Menke, F.L.H.; Torres, M.-A.; Molina, A.; Zipfel, C.; Coaker, G.; et al. Quantitative Phosphoproteomic Analysis Reveals Common Regulatory Mechanisms between Effector- and PAMP-Triggered Immunity in Plants. New Phytol. 2019, 221, 2160–2175. [Google Scholar] [CrossRef]
- van Dongen, J.T.; Licausi, F. Oxygen Sensing and Signaling. Annu. Rev. Plant Biol. 2015, 66, 345–367. [Google Scholar] [CrossRef]
- Song, G.; Kwon, C.-T.; Kim, S.-H.; Shim, Y.; Lim, C.; Koh, H.-J.; An, G.; Kang, K.; Paek, N.-C. The Rice SPOTTED LEAF4 (SPL4) Encodes a Plant Spastin That Inhibits ROS Accumulation in Leaf Development and Functions in Leaf Senescence. Front. Plant Sci. 2019, 9, 1925. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, J.-M. Receptor-Like Cytoplasmic Kinases: Central Players in Plant Receptor Kinase–Mediated Signaling. Annu. Rev. Plant Biol. 2018, 69, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Cernadas, R.A.; Doyle, E.L.; Niño-Liu, D.O.; Wilkins, K.E.; Bancroft, T.; Wang, L.; Schmidt, C.L.; Caldo, R.; Yang, B.; White, F.F.; et al. Code-Assisted Discovery of TAL Effector Targets in Bacterial Leaf Streak of Rice Reveals Contrast with Bacterial Blight and a Novel Susceptibility Gene. PLoS Pathog. 2014, 10, e1003972. [Google Scholar] [CrossRef] [PubMed]
- Vij, S.; Giri, J.; Dansana, P.K.; Kapoor, S.; Tyagi, A.K. The Receptor-Like Cytoplasmic Kinase (OsRLCK) Gene Family in Rice: Organization, Phylogenetic Relationship, and Expression during Development and Stress. Mol. Plant 2008, 1, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Han, T.; Song, Q.; Ye, W.; Song, X.; Chu, J.; Li, J.; Chen, Z.J. The Rice Circadian Clock Regulates Tiller Growth and Panicle Development Through Strigolactone Signaling and Sugar Sensing. Plant Cell 2020, 32, 3124–3138. [Google Scholar] [CrossRef]
- Wang, A.; Shu, X.; Jing, X.; Jiao, C.; Chen, L.; Zhang, J.; Ma, L.; Jiang, Y.; Yamamoto, N.; Li, S.; et al. Identification of Rice (Oryza Sativa L.) Genes Involved in Sheath Blight Resistance via a Genome-wide Association Study. Plant Biotechnol. J. 2021, 19, 1553–1566. [Google Scholar] [CrossRef]
- Cui, Z.; Xue, C.; Mei, Q.; Xuan, Y. Malectin Domain Protein Kinase (MDPK) Promotes Rice Resistance to Sheath Blight via IDD12, IDD13, and IDD14. Int. J. Mol. Sci. 2022, 23, 8214. [Google Scholar] [CrossRef]
- Xu, X.; Xu, Z.; Li, Z.; Zakria, M.; Zou, L.; Chen, G. Increasing Resistance to Bacterial Leaf Streak in Rice by Editing the Promoter of Susceptibility Gene OsSULRT3;6. Plant Biotechnol. J. 2021, 19, 1101–1103. [Google Scholar] [CrossRef]
- Kumar, K.; Mosa, K.A.; Chhikara, S.; Musante, C.; White, J.C.; Dhankher, O.P. Two Rice Plasma Membrane Intrinsic Proteins, OsPIP2;4 and OsPIP2;7, Are Involved in Transport and Providing Tolerance to Boron Toxicity. Planta 2014, 239, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Back, K. Molecular Cloning of Melatonin 2-hydroxylase Responsible for 2-hydroxymelatonin Production in Rice (ORyza Sativa). J. Pineal Res. 2015, 58, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Li, Y. Genome-Wide Identification and Expression Profiles of 13 Key Structural Gene Families Involved in the Biosynthesis of Rice Flavonoid Scaffolds. Genes 2022, 13, 410. [Google Scholar] [CrossRef]
- Yang, L.; Wang, C.C.; Guo, W.D.; Li, X.B.; Lu, M.; Yu, C.L. Differential Expression of Cell Wall Related Genes in the Elongation Zone of Rice Roots under Water Deficit. Russ. J. Plant Physiol. 2006, 53, 390–395. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.-J.; Cao, G.-C.; Gu, S.-H.; Wu, K.-M.; Gao, X.-W.; Guo, Y.-Y. Rice Gene Expression Profiles Responding to Larval Feeding of the Striped Stem Borer at the 1st to 2nd Instar Stage. Insect Sci. 2011, 18, 273–281. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, X.; Ning, Y.; Jing, W.; Bruce, T.J.A.; Qi, F.; Xu, Q.; Wu, K.; Zhang, Y.; Guo, Y. TPS46, a Rice Terpene Synthase Conferring Natural Resistance to Bird Cherry-Oat Aphid, Rhopalosiphum Padi (Linnaeus). Front. Plant Sci. 2017, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiang, Z.; Li, J.; Wang, S.; Chen, Y.; Liu, Y.; Mao, D.; Luan, S.; Chen, L. BHLH57 Confers Chilling Tolerance and Grain Yield Improvement in Rice. Plant Cell Environ. 2023, 46, 1402–1418. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Cho, K.; Shibato, J.; Han, O.; Iwahashi, H.; Tamogami, S.; Majeed Zargar, S.; Kubo, A.; Masuo, Y.; Agrawal, G.K.; et al. Rice OsOPRs: Transcriptional Profiling Responses to Diverse Environmental Stimuli and Biochemical Analysis of OsOPR1. J. Plant Biol. 2009, 52, 229–243. [Google Scholar] [CrossRef]
- Kuroha, T.; Nagai, K.; Gamuyao, R.; Wang, D.R.; Furuta, T.; Nakamori, M.; Kitaoka, T.; Adachi, K.; Minami, A.; Mori, Y.; et al. Ethylene-Gibberellin Signaling Underlies Adaptation of Rice to Periodic Flooding. Science 2018, 361, 181–186. [Google Scholar] [CrossRef]
- Wang, H.; Hao, J.; Chen, X.; Hao, Z.; Wang, X.; Lou, Y.; Peng, Y.; Guo, Z. Overexpression of Rice WRKY89 Enhances Ultraviolet B Tolerance and Disease Resistance in Rice Plants. Plant Mol. Biol. 2007, 65, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Tasaki, K.; Komori, H.; Katou, S. Hypersensitivity-Related Genes HSR201 and HSR203J Are Regulated by Calmodulin-Binding Protein 60-Type Transcription Factors and Required for Pathogen Signal-Induced Salicylic Acid Synthesis. Plant Cell Physiol. 2022, 63, 1008–1022. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Niu, R.; Yu, H.; Guo, J.; Du, C.; Zhang, Z.; Wei, Y.; Li, J.; Zhang, S. OsSLA1Functions in Leaf Angle Regulation by Enhancing the Interaction between OsBRI1and OsBAK1in Rice. Plant J. 2022, 110, 1111–1127. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Tian, Q.; Deng, P.; Zhang, W.; Jing, W. The Rice Aldehyde Oxidase OsAO3 Gene Regulates Plant Growth, Grain Yield, and Drought Tolerance by Participating in ABA Biosynthesis. Biochem. Biophys. Res. Commun. 2021, 548, 189–195. [Google Scholar] [CrossRef]
- Takai, R.; Isogai, A.; Takayama, S.; Che, F.-S. Analysis of Flagellin Perception Mediated by Flg22 Receptor OsFLS2 in Rice. Mol. Plant-Microbe Interact. 2008, 21, 1635–1642. [Google Scholar] [CrossRef]
- Yu, H.; Yan, J.; Du, X.; Hua, J. Overlapping and Differential Roles of Plasma Membrane Calcium ATPases in Arabidopsis Growth and Environmental Responses. J. Exp. Bot. 2018, 69, 2693–2703. [Google Scholar] [CrossRef] [PubMed]
- Schwessinger, B.; Roux, M.; Kadota, Y.; Ntoukakis, V.; Sklenar, J.; Jones, A.; Zipfel, C. Phosphorylation-Dependent Differential Regulation of Plant Growth, Cell Death, and Innate Immunity by the Regulatory Receptor-like Kinase BAK1. PLoS Genet. 2011, 7, e1002046. [Google Scholar] [CrossRef]
- Albrecht, C.; Boutrot, F.; Segonzac, C.; Schwessinger, B.; Gimenez-Ibanez, S.; Chinchilla, D.; Rathjen, J.P.; de Vries, S.C.; Zipfel, C. Brassinosteroids Inhibit Pathogen-Associated Molecular Pattern-Triggered Immune Signaling Independent of the Receptor Kinase BAK1. Proc. Natl. Acad. Sci. USA 2012, 109, 303–308. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, S. MAPK Cascades in Plant Disease Resistance Signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Guo, P.; Xia, X.; Guo, H.; Li, Z. Multiple Layers of Regulation on Leaf Senescence: New Advances and Perspectives. Front. Plant Sci. 2021, 12, 788996. [Google Scholar] [CrossRef]
- Li, Z.; Peng, J.; Wen, X.; Guo, H. Ethylene-Insensitive3 Is a Senescence-Associated Gene That Accelerates Age-Dependent Leaf Senescence by Directly Repressing MiR164 Transcription in Arabidopsis. Plant Cell 2013, 25, 3311–3328. [Google Scholar] [CrossRef]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.; Penfold, C.A.; Jenkins, D.; et al. High-Resolution Temporal Profiling of Transcripts during Arabidopsis Leaf Senescence Reveals a Distinct Chronology of Processes and Regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef]
- Ahmad, S.; Guo, Y. Signal Transduction in Leaf Senescence: Progress and Perspective. Plants 2019, 8, 405. [Google Scholar] [CrossRef]
- Liu, M.; Shi, Z.; Zhang, X.; Wang, M.; Zhang, L.; Zheng, K.; Liu, J.; Hu, X.; Di, C.; Qian, Q.; et al. Inducible Overexpression of Ideal Plant Architecture1 Improves Both Yield and Disease Resistance in Rice. Nat. Plants 2019, 5, 389–400. [Google Scholar] [CrossRef]
- Lecourieux, D.; Ranjeva, R.; Pugin, A. Calcium in Plant Defence-Signalling Pathways. New Phytologist 2006, 171, 249–269. [Google Scholar] [CrossRef]
- Costa, A.; Navazio, L.; Szabo, I. The Contribution of Organelles to Plant Intracellular Calcium Signalling. J. Exp. Bot. 2018, 69, 4175–4193. [Google Scholar] [CrossRef]
- Kaur, A.; Madhu; Taneja, M.; Upadhyay, S.K. Applications of Calcium Transport Elements in Plant Improvement. In Calcium Transport Elements in Plants; Elsevier: Amsterdam, The Netherlands, 2021; pp. 427–445. [Google Scholar]
- Chand, V.; Gupta, V. Interaction between Ca2+ and ROS Signaling in Plants. In Calcium Transport Elements in Plants; Elsevier: Amsterdam, The Netherlands, 2021; pp. 387–410. [Google Scholar]
- Ranf, S.; Eschen-Lippold, L.; Pecher, P.; Lee, J.; Scheel, D. Interplay between Calcium Signalling and Early Signalling Elements during Defence Responses to Microbe- or Damage-Associated Molecular Patterns. Plant J. 2011, 68, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Aichi, I.; Matsuoka, M. A Protocol for Agrobacterium-Mediated Transformation in Rice. Nat. Protoc. 2006, 1, 2796–2802. [Google Scholar] [CrossRef] [PubMed]
- Hartmut, K. Lichtenthaler Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 350–382. [Google Scholar]
- Kauffman, H.E.; Reddy, A.P.; Hsieh, S.P.; Merca, S.D. An Improved Technique for Evaluating Resistance of Rice Varieties to Xanthomonas Oryzae. Plant Dis. Report. 1973, 57, 537–541. [Google Scholar]
- Wu, T.; Bi, Y.; Yu, Y.; Zhou, Z.; Yuan, B.; Ding, X.; Zhang, Q.; Chen, X.; Yang, H.; Liu, H.; et al. Activated Expression of Rice DMR6-like Gene OsS3H Partially Explores the Susceptibility to Bacterial Leaf Streak Mediated by Knock-Out OsF3H04g. Int. J. Mol. Sci. 2023, 24, 13263. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular Localization of H2O2 in Plants. H2O2 Accumulation in Papillae and Hypersensitive Response during the Barley—Powdery Mildew Interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Definition | Fold (log2) | Function | Reference |
|---|---|---|---|---|---|
| Os01g0113500 | OsRLCK9 | receptor-like kinase | 2.642 | ||
| Os01g0116900 | OsRLCK17 | LRK-type protein; protein kinase domain containing protein | 3.050 | Resistance to Xanthomonas oryzae | [49] |
| Os01g0116800 | OsRLCK18 | TAK14 | 4.914 | Resistance to abiotic stress | [49] |
| Os11g0695000 | OsRLCK354 | leucine-rich repeat receptor protein kinase EXS precursor | 6.877 | ||
| Os11g0691800 | OsRLCK355 | receptor-like protein kinase precursor | 4.635 | ||
| Os04g0127500 | OsWAK29 | OsWAK receptor-like protein kinase | 6.259 | ||
| Os04g0370100 | OsWAK46 | OsWAK receptor-like protein kinase | 7.100 | ||
| Os02g0618200 | OsRSR1 | two-component response regulator | 3.034 | Regulate tiller growth and panicle development | [50] |
| Os11g0229500 | OsRSR | disease resistance protein | 9.878 | Resistance to SHB | [51] |
| Os01g0124000 | OsRBBI2 | BBTI2-Bowman-Birk type bran trypsin inhibitor precursor | 6.995 | ||
| Os09g0355400 | MDPK | malectin domain protein kinase | 9.393 | Resistance to SHB | [52] |
| Os01g0719300 | OsSULTR3;6 | secondary active sulfate transmembrane transporter activity | −4.383 | Resistance to bacterial leaf streak | [53] |
| Os06g0306300 | OsPRX78 | peroxidase precursor | 3.878 | ||
| Os09g0541000 | OsPIP2;7 | aquaporin, plasma membrane intrinsic protein | 4.199 | Tolerance to boron toxicity | [54] |
| Os01g0901600 | OsACS6 | AMP-binding domain containing protein | 2.457 | Phenylpropanoid biosynthesis | |
| Os04g0182200 | Os2ODD23 | 1-aminocyclopropane-1-carboxylate oxidase homolog 2 | 2.366 | Benzoxazinoid biosynthesis | [55] |
| Os03g0289800 | Os2ODD8 | leucoanthocyanidin dioxygenase | 2.387 | Response to low-temperature stress and salt stress | [56] |
| Os04g0604300 | OsXTH9 | glycosyl hydrolases family 16 | 3.210 | Cell wall modification | [57] |
| Os08g0167800 | TPS30;46 | terpene synthase | 4.295 | Key gene for biosynthesis of limonene, methyl salicylate | [58] |
| Os08g0168000 | OsTPS31 | terpene synthase activity | 4.549 | Resistance to insect | [59] |
| Os07g0543000 | OsbHLH057 | bHLH transcription factor | 1.557 | Stress tolerance and yield | [60] |
| Os06g0215900 | OsOPR4 | 12-oxophytodienoate reductase | 8.469 | Plant hormone response related; Jasmonic Acid biosynthesis related | [61] |
| Os03g0324200 | OsEIN3 | ethylene signaling regulation factors | 6.398 | Involved in environmental humidity regulation of rice blast resistance in rice | [62] |
| Os11g0117400 | OsWRKY104 | WRKY transcription factor | 2.929 | Response to biotic and abiotic stresses | [63] |
| Os11g0685700 | OsWRKY61 | DNA-binding transcription factor activity | 9.098 | ||
| Os12g0458100 | HSR201 | transferase family protein, putative, expressed | 5.811 | Hypersensitivity-related; required for pathogen signal-induced salicylic acid synthesis | [64] |
| Os11g051450 | OsBAK1 | brassinosteroid insensitive 1-associated receptor kinase 1 precursor | −6.408 | plant growth and development; PAMP triggered immune response | [65] |
| Os10g0138100 | OsAO2, AAO2 | aldehyde oxidase | 12.345 | Regulates plant growth, grain yield, and drought tolerance | [66] |
| Os04g0618700 | OsFLS2 | leucine-rich repeat receptor protein kinase EXS precursor; flg22 Receptor | 2.101 | PAMP triggered immune response | [67] |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| OsActin | GTTACTCATTCACCACAACGGC | CCTTTCAGGAGGGGCGACC |
| OsEF1 | AAGAGGAAGTCAGCGGCTAAG | CAGAATGGGCAGGAAAATACA |
| OsACA9 | CGGGACGACGACGATGGC | CGCGCTTGGCGGGGATGT |
| OsACA11 | TGGCGGCAAGCAGCTCTAGT | CGGAATGCAGCCCTGACAAC |
| OsBAK1 | ATCAACTGGGTGGAGAGTGA | ATCTCCCAAGGTGTGTGGTA |
| OsEIN3 | CCTCAAGAAGGCCTGGAAGG | CTTGGCGGTCATCTTGTCCT |
| OsRLCK354 | TCCGTCCTTAACCTCACCAT | GGGATTTGTCCAGAGAGCTG |
| OsTPS30 | TACCGCTGGCTATACTCACA | CTCATGGCTTCTAGGCTTGG |
| OsAO2 | TGCTGGGAGAAAGCCGAAAT | TGGCACTCTGCACTTGACTT |
| OsFLS2 | AGAGATTGTCGCTCCATGCC | AGAGCGAGTTGTTCTGGACG |
| OsRSR1 | AACACACTCCCAATCCGCTT | AGGGTGGTTTTCCCCAATCC |
| MDPK | GGACGCAACACCTGGATACT | CGTGTATGGAGTGGTGCCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, Z.; Lu, Y.; Huang, J.; Hu, Z.; Lou, J.; Fan, X.; Gu, Z.; Liu, P.; Ma, B.; et al. OsACA9, an Autoinhibited Ca2+-ATPase, Synergically Regulates Disease Resistance and Leaf Senescence in Rice. Int. J. Mol. Sci. 2024, 25, 1874. https://doi.org/10.3390/ijms25031874
Wang X, Wang Z, Lu Y, Huang J, Hu Z, Lou J, Fan X, Gu Z, Liu P, Ma B, et al. OsACA9, an Autoinhibited Ca2+-ATPase, Synergically Regulates Disease Resistance and Leaf Senescence in Rice. International Journal of Molecular Sciences. 2024; 25(3):1874. https://doi.org/10.3390/ijms25031874
Chicago/Turabian StyleWang, Xinyu, Ziyao Wang, Yiduo Lu, Jiani Huang, Zhuoer Hu, Junlei Lou, Xinyue Fan, Zhimin Gu, Pengcheng Liu, Bojun Ma, and et al. 2024. "OsACA9, an Autoinhibited Ca2+-ATPase, Synergically Regulates Disease Resistance and Leaf Senescence in Rice" International Journal of Molecular Sciences 25, no. 3: 1874. https://doi.org/10.3390/ijms25031874
APA StyleWang, X., Wang, Z., Lu, Y., Huang, J., Hu, Z., Lou, J., Fan, X., Gu, Z., Liu, P., Ma, B., & Chen, X. (2024). OsACA9, an Autoinhibited Ca2+-ATPase, Synergically Regulates Disease Resistance and Leaf Senescence in Rice. International Journal of Molecular Sciences, 25(3), 1874. https://doi.org/10.3390/ijms25031874