
Mucus Structure, Viscoelastic Properties, and Composition in Chronic Respiratory Diseases

 , ,
, ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
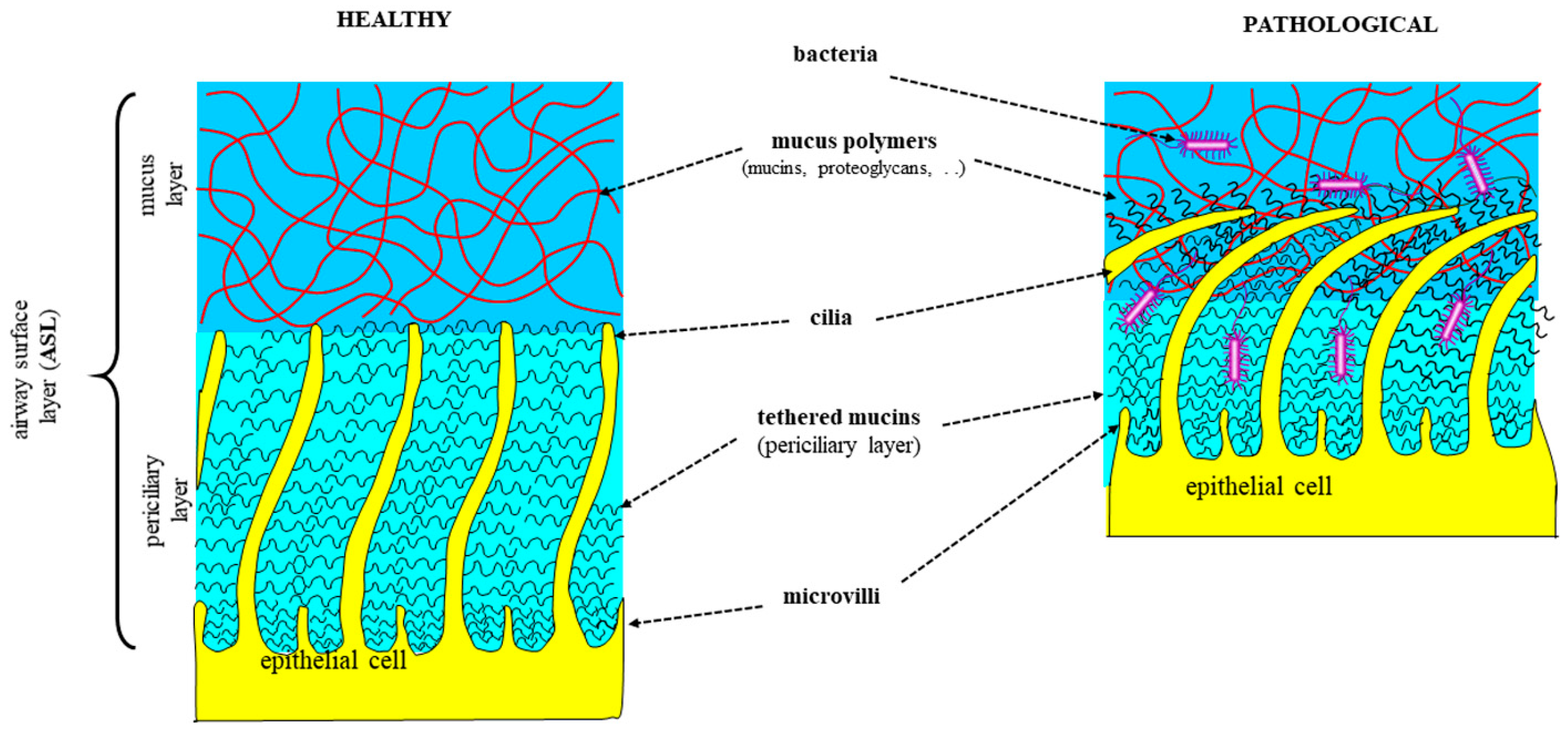
1.1. Respiratory Mucus in Healthy and Diseased Conditions
1.2. Clinical and Pathological Aspects in CRDs
1.3. Pathophysiologal Changes in CRD Mucus/Sputum
1.3.1. Physiology of Respiratory Mucus Secretion
1.3.2. Mucins, Goblet Cells, and Submucosal Glands in Pathophysiology of Mucus Secretion
1.4. Pathophysiology of Mucus Production in CRDs
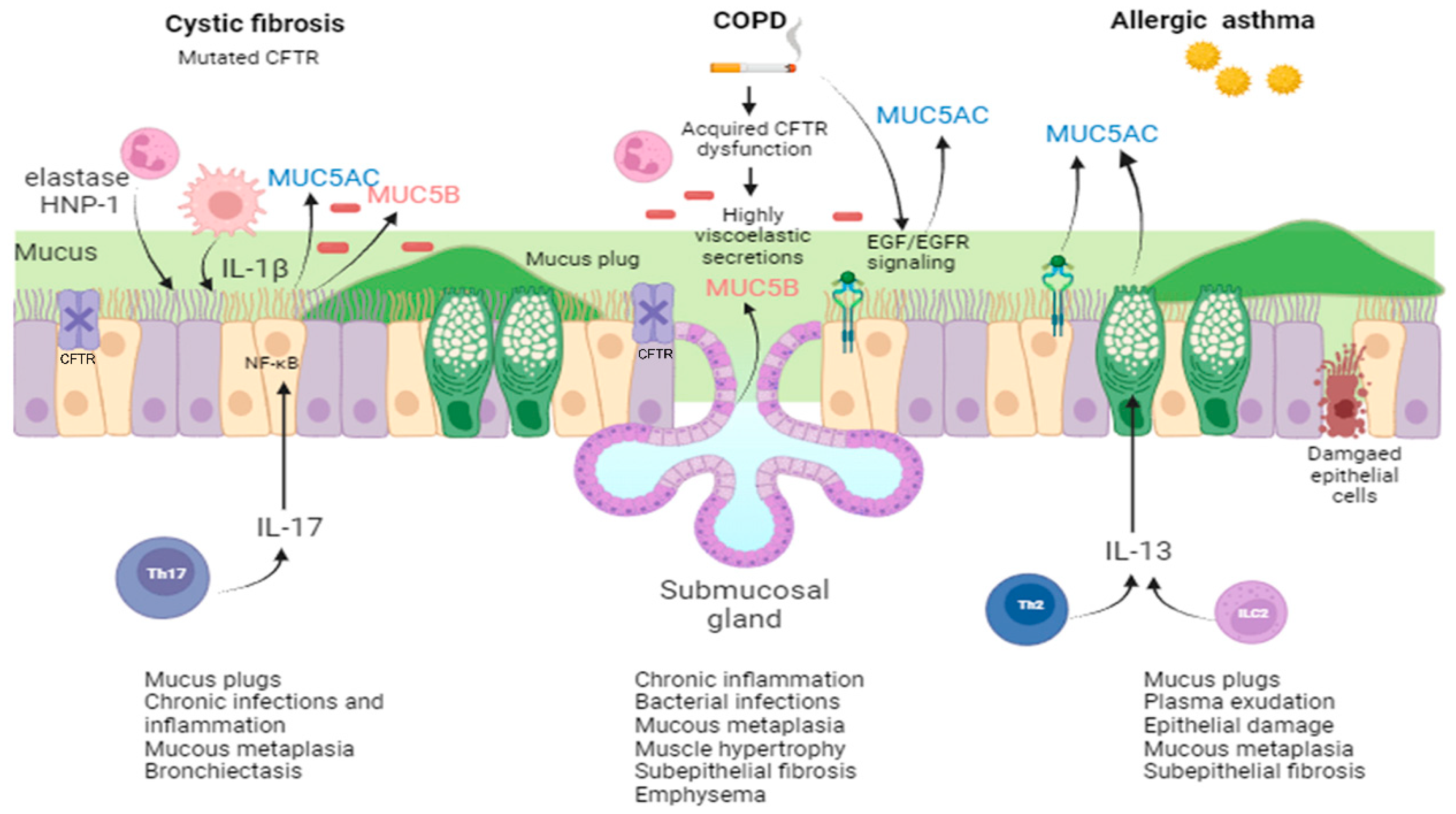
2. Mucus Structure in CRDs
2.1. Mucus in CF
2.2. Mucus in COPD
2.3. Mucus in Asthma
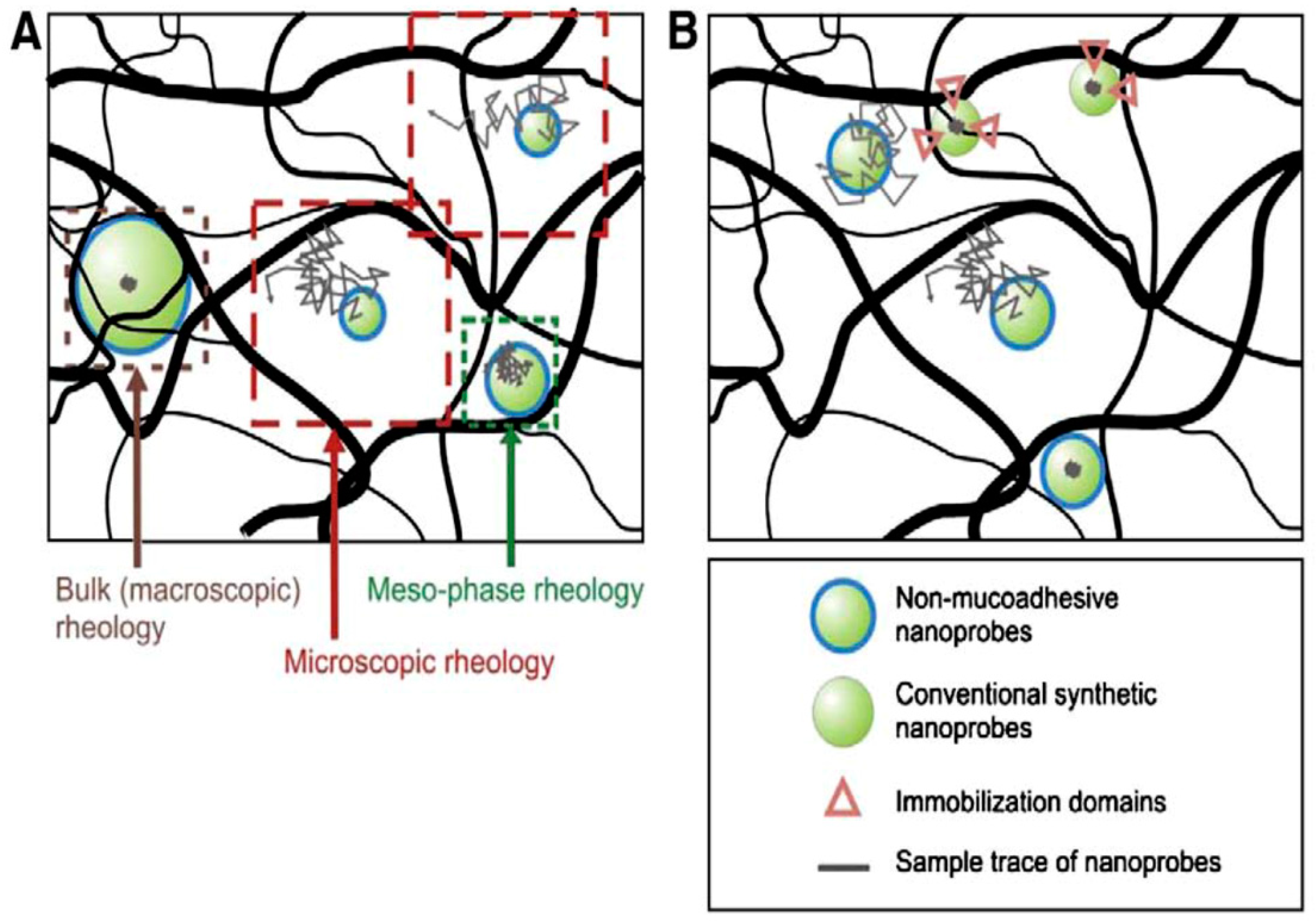
3. Mucus Viscoelastic Properties in CRDs
3.1. Mucus Viscoelasticity
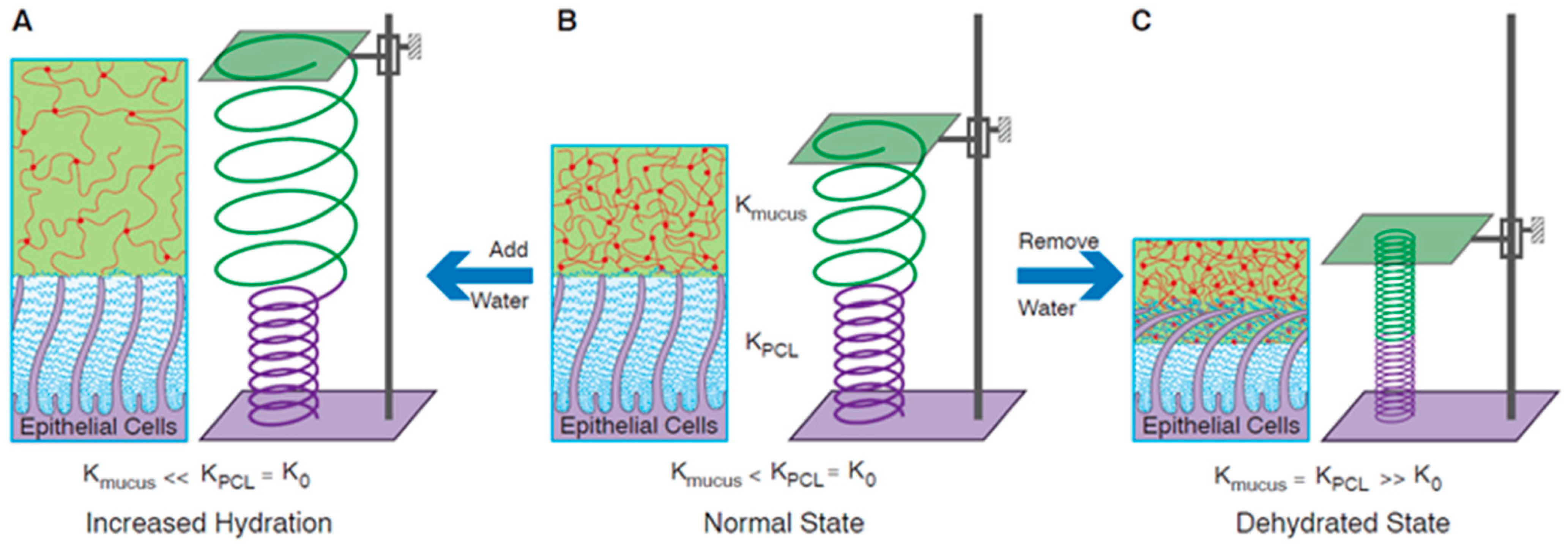
3.2. Osmotic Pressure
3.3. The Role of pH and Ionic Strength in Mucus Properties
4. Mucus/Sputum Mediators as Biomarkers in CRDs
4.1. Cystic Fibrosis (Table 2)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Cohorts | Sputum Samples | Outcomes | Reference |
|---|---|---|---|
| Three retrospective CF cohorts spanning a wide range of diseases | Induced or spontaneous sputum | Sputum resistin levels were negatively correlated with CF lung function | Forrest et al., 2019 [253] |
| CF patients (n = 47) with ppFEV1 of 67.5 (19.3) (mean (SD)) | Spontaneous sputum | NE activity correlates with chronic P. aeruginosa infection according to ELISA and fluorogenic assays, while NE activity correlates with ppFEV1 according to the fluorogenic assay | Oriano et al., 2019 [252] |
| CF patients (n = 31) 10 years and older, with at least one G551D CFTR allele | Induced or spontaneous sputum | IVA treatment did not result in significant changes in inflammatory marker levels (Il-1β, IL-6, IL-8, NE, SLPI, and AAT) | Harris et al., 2020 [273] |
| CF patients (n = 70), median age 11.8 years | Induced sputum | During acute exacerbation, sputum arginin and homoarginine levels were high and dropped after antibiotic treatment | Hanusch et al., 2020 [279] |
| Adult CF patients in two study cohorts | Spontaneous sputum | Serine trypsin-like proteases (TLP) activity inversely correlated with ppFEV1. Individuals with high TLP activity showed significantly reduced survival | Reihill et al., 2020 [255] |
| CF patients (n = 30) 12 years and older homozygous for the F508del mutation | Spontaneous sputum | LUMA-IVA decreased IL-1β levels after 8–16 weeks of treatment | Graeber et al., 2021 [274] |
| CF patients (n = 73) 10 years of age or older and with ppFEV1 of 74.1 (14.4) (mean (SD)) | Induced sputum | At baseline and after 16 weeks, NE negatively correlated with ppFEV1 | Jain et al., 2021 [258] |
| CF patients (n = 20) 29.9 (8.5) years with ppFEV1 of 49 (22) (mean (SD)) | Spontaneous sputum | For sustained-symptom responders, sputum iron was relatively stable, while for non-sustained symptom-responders, sputum iron first decreased and then increased around day 7 of treatment | Gifford et al., 2021 [280] |
| CF patients across 139 articles and 71 biomarkers | Spontaneous and induced sputum | NE, IL-8, TNF-α, and IL-1β demonstrated validity between CF and non-CF subjects, as well as responsiveness to therapies | Lepissier et al., 2022 [232] |
| Randomly chosen clinical stable CF patients (n = 114) 28 (12) years with ppFEV1 of 70 (22) (mean (SD)) | Spontaneous sputum | Biomarkers associated with time to next exacerbation: ENRAGE, MPO, sRAGE, ICAM-1, NE, YKL40, TARC, MMP-9, IL-1β, IL-5 | Liou et al., 2022 [260] |
| CF patients (n = 38) >18 years of age | Spontaneous sputum | Proteomic analysis revealed that baseline ppFEV1 severity correlated with 87 proteins; positive correlation n = 20, negative n = 67); most were either neutrophil-derived or involved in neutrophil-driven oxidant and protease activity | Maher et al., 2022 [269] |
| CF children (n = 27) with median age of 11.4 years | Spontaneous sputum | There were higher levels of hBD-2 and hCAP-18/LL-37 in the study group compared to the control group | Ishchenko et al., 2022 [282] |
| CF patients (n = 44) aged 16 years and older homozygous for the F508del mutation | Spontaneous sputum | After 12 months of LUMA–IVA treatment, significant reductions in sputum IL-6, IL-8, IL-1β, and TNF-α levels were observed | Arooj et al., 2023 [275] |
| CF patients (n = 79) aged ≥12 years with one or two F508del alleles | Spontaneous sputum | ETI improved the elastic modulus and viscous modulus of CF sputum at 3 and 12 months after initiation, reduced IL-8 and free NE activity, and shifted the CF sputum proteome toward healthy | Schaupp et al., 2023 [276] |
4.2. COPD (Table 3)
| Patient Cohorts | Sputum Samples | Outcomes | Reference |
|---|---|---|---|
| 1 male COPD patient (diagnosed according to GOLD guideline 2017) | Spontaneous sputum | In the study, a new technology was used to reduce the complexity of clinical samples so as to optimize MALDI-TOF peptidome profiling. | Terracciano et al., 2019 [289] |
| 90 COPD patients: 50 with a history of smoking (COPD/tobacco) and 40 who previously had TB (COPD/post-TB) | Spontaneous sputum | IL-1α, IL-1β, MIP-1β, sCD40L, and VEGF levels were higher in COPD patients compared to controls; IL-1α, IL-6, TNF-α, and IL-8 levels were higher in the COPD/tobacco patients compared to the COPD/post-TB patients. | Guiedem et al., 2020 [303] |
| 31 patients with different diseases | Spontaneous/ induced sputum | Identification of proteins with potential applicability in clinical practice, e.g., markers of exacerbation. | Dasgupta et al., 2021 [290] |
| COPD patients (n = 14) with mean age of 65 years; asthmatic patients (n = 21) with mean age of 55 | Induced sputum | Considering all patients, SLPI and hBD-1 were negatively correlated with ppFEV1 (p < 0.001, r = −0.610). SLPI and hBD-1 were higher in the COPD group compared to the asthma group, while elafin levels were not different. | Cane et al., 2021 [308] |
| Clinically stable COPD patients (n = 25) and COPD patients with exacerbations (n = 31) | Spontaneous sputum | The sputum cytokine signature of exacerbated patients differs from that of stable COPD patients. The observation that the levels of most cytokines do not stabilize with acute treatment of exacerbated patients suggests a prolonged effect of exacerbation on the status of COPD patients. | Barta et al., 2022 [291] |
| 341 smokers with preserved spirometry, and 562 patients with COPD (178 with GOLD stage 1 disease, 303 with GOLD stage 2 disease, and 81 with GOLD stage 3 disease) | Induced sputum | The study identified several physiologic pathways altered in the airways of patients with COPD and associated with markers of disease severity, with the strongest relationships to metabolite biomarkers of mucus hydration and adenosine metabolism. | Esther et al., 2022 [294] |
| Cohort A: COPD patients (n = 30), healthy smokers (n = 10), and healthy non-smokers (n = 10); Cohort B: COPD patients (n = 81), healthy smokers (n = 15), and healthy non-smokers (n = 26) | Spontaneous sputum | Validated immunoassays applied to sputum supernatants demonstrated differences between COPD patients and controls, the effects of current smoking, and associations between H. influenzae colonization and higher levels of selected cytokines. | Mulvanny et al., 2022 [301] |
4.3. Asthma (Table 4)
| Patient Cohorts | Sputum Samples | Outcomes | Reference |
|---|---|---|---|
| 246 patients: 118 nonsmoking patients with severe asthma; 48 current or ex-smoking patients with severe asthma; 40 patients with mild-to-moderate asthma | Induced sputum | The current classifications of asthma based on cell count (eosinophils and neutrophils) were improved through proteomic characterization. | Schofield et al., 2019 [312] |
| 102 adults and 34 children with moderate-to-severe asthma | Spontaneous or induced sputum | TNF-rcceptor2 and IL-6 receptor were the strongest discriminators of an exacerbation in both adults and children. | Ghebre et al., 2019 [325] |
| AMAZES clinical trial (ACTRN12609000197235) | Spontaneous sputum | TNF sputum marker activity correlates with asthma severity and decreases after azithromycin treatment. | Niessen et al., 2021 [314] |
| 48 children (acute asthma, n = 18; stable asthma, n = 17) | Induced sputum | CXCL-10, CCL-5, and TNF-R2 were the strongest discriminators of an asthma attack. | Ramphul et al., 2021 [326] |
| Asthmatic patients (n = 21) with mean age of 55; COPD patients (n = 14) with mean age of 65 years | Induced sputum | Considering all patients, SLPI and hBD-1 were negatively correlated with ppFEV1. SLPI and hBD-1 were lower in the asthma group compared to the COPD group, while elafin levels were not different. | Cane et al., 2021 [308] |
| 65 adult asthmatic subjects | Induced sputum | Sputum neurturin is a new marker for type 2 airway inflammation. | Sato et al., 2023 [317] |
| 52 patients with severe asthma (51 patients treated with mepolizumab and one patient treated with reslizumab) | Induced sputum | Baseline type 2 airway inflammation markers can predict remission in severe eosinophilic asthma treated with anti-IL-5 agents. | Moermans et al., 2023 [322] |
| 200 asthmatic patients | Induced sputum | Three groups of highly correlated cytokines and alarmins were identified: two T2 modules, the TSLPmod (TSLP, IL-4, IL-5, IL-9) and the IL-33mod (IL-33, IL-13, IL-21), and one non-T2 module, the IL-1βmod (IL-1β, IL-6, G-CSF). The TSLPmod was associated with asthma severity, airway obstruction, eosinophilia, and elevated FeNO. | Gautam et al., 2023 [324] |
| 65 adult asthmatic subjects | Induced sputum | Sputum neurturin is a new marker for type 2 airway inflammation. | Sato et al., 2023 [317] |
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2019 Chronic Respiratory Diseases Collaborators. Global burden of chronic respiratory diseases and risk factors, 1990–2019: An update from the Global Burden of Disease Study 2019. EClinicalMedicine 2023, 59, 101936. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chronic Obstructive Pulmonary Disease (COPD). 16 March 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/chronic-obstructive-pulmonary-disease-(copd) (accessed on 29 October 2023).
- Diseases, G.B.D.; Injuries, C. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar]
- World Health Organization. Asthma. 4 May 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/asthma (accessed on 29 October 2023).
- Scotet, V.; L’Hostis, C.; Ferec, C. The Changing Epidemiology of Cystic Fibrosis: Incidence, Survival and Impact of the CFTR Gene Discovery. Genes 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Kuhn, M.; Prettner, K.; Yu, F.; Yang, T.; Barnighausen, T.; Bloom, D.E.; Wang, C. The global economic burden of chronic obstructive pulmonary disease for 204 countries and territories in 2020-50: A health-augmented macroeconomic modelling study. Lancet Glob. Health 2023, 11, e1183–e1193. [Google Scholar] [CrossRef] [PubMed]
- Ghorani, V.; Boskabady, M.H.; Khazdair, M.R.; Kianmeher, M. Experimental animal models for COPD: A methodological review. Tob. Induc. Dis. 2017, 15, 25. [Google Scholar] [CrossRef]
- Ehre, C.; Hansson, G.C.; Thornton, D.J.; Ostedgaard, L.S. Mucus aberrant properties in CF: Insights from cells and animal models. J. Cyst. Fibros. 2023, 22 (Suppl. S1), S23–S26. [Google Scholar] [CrossRef]
- McCarron, A.; Parsons, D.; Donnelley, M. Animal and Cell Culture Models for Cystic Fibrosis: Which Model Is Right for Your Application? Am. J. Pathol. 2021, 191, 228–242. [Google Scholar] [CrossRef]
- Atanasova, K.R.; Reznikov, L.R. Strategies for measuring airway mucus and mucins. Respir. Res. 2019, 20, 261. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.K.; Wang, Y.Y.; Wirtz, D.; Hanes, J. Micro- and macrorheology of mucus. Adv. Drug Deliv. Rev. 2009, 61, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Thornton, D.J.; Sheehan, J.K. From mucins to mucus: Toward a more coherent understanding of this essential barrier. Proc. Am. Thorac. Soc. 2004, 1, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Samet, J.M.; Cheng, P.W. The role of airway mucus in pulmonary toxicology. Environ. Health Perspect. 1994, 102 (Suppl. S2), 89–103. [Google Scholar] [PubMed]
- Matthews, L.W.; Spector, S.; Lemm, J.; Potter, J.L. Studies on Pulmonary Secretions. I. The over-All Chemical Composition of Pulmonary Secretions from Patients with Cystic Fibrosis, Bronchiectasis, and Laryngectomy. Am. Rev. Respir. Dis. 1963, 88, 199–204. [Google Scholar] [PubMed]
- Mrsny, R.J.; Daugherty, A.L.; Short, S.M.; Widmer, R.; Siegel, M.W.; Keller, G.A. Distribution of DNA and alginate in purulent cystic fibrosis sputum: Implications to pulmonary targeting strategies. J. Drug Target. 1996, 4, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Schuster, B.S.; Suk, J.S.; Woodworth, G.F.; Hanes, J. Nanoparticle diffusion in respiratory mucus from humans without lung disease. Biomaterials 2013, 34, 3439–3446. [Google Scholar] [CrossRef] [PubMed]
- MacNee, W. Pathology, pathogenesis, and pathophysiology. BMJ 2006, 332, 1202–1204. [Google Scholar] [CrossRef]
- Burgel, P.R.; Nesme-Meyer, P.; Chanez, P.; Caillaud, D.; Carre, P.; Perez, T.; Roche, N.; Initiatives Bronchopneumopathie Chronique Obstructive (BPCO) Scientific Committee. Cough and sputum production are associated with frequent exacerbations and hospitalizations in COPD subjects. Chest 2009, 135, 975–982. [Google Scholar] [CrossRef]
- Wedzicha, J.A.; Singh, R.; Mackay, A.J. Acute COPD exacerbations. Clin. Chest Med. 2014, 35, 157–163. [Google Scholar] [CrossRef]
- Wedzicha, J.A. Mechanisms of Chronic Obstructive Pulmonary Disease Exacerbations. Ann. Am. Thorac. Soc. 2015, 12 (Suppl. S2), S157–S159. [Google Scholar] [CrossRef]
- Saetta, M.; Turato, G. Airway pathology in asthma. Eur. Respir. J. Suppl. 2001, 34, 18s–23s. [Google Scholar] [CrossRef]
- Gibson, R.L.; Burns, J.L.; Ramsey, B.W. Pathophysiology and management of pulmonary infections in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2003, 168, 918–951. [Google Scholar] [CrossRef]
- Thornton, C.S.; Parkins, M.D. Microbial Epidemiology of the Cystic Fibrosis Airways: Past, Present, and Future. Semin. Respir. Crit. Care Med. 2023, 44, 269–286. [Google Scholar] [CrossRef]
- Fahy, J.V.; Dickey, B.F. Airway mucus function and dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.K.; Thornton, D.J.; Somerville, M.; Carlstedt, I. Mucin structure. The structure and heterogeneity of respiratory mucus glycoproteins. Am. Rev. Respir. Dis. 1991, 144, S4-9. [Google Scholar] [CrossRef]
- Voynow, J.A.; Rubin, B.K. Mucins, mucus, and sputum. Chest 2009, 135, 505–512. [Google Scholar] [CrossRef]
- Curran, D.R.; Cohn, L. Advances in mucous cell metaplasia: A plug for mucus as a therapeutic focus in chronic airway disease. Am. J. Respir Cell Mol. Biol. 2010, 42, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Ostedgaard, L.S.; Moninger, T.O.; McMenimen, J.D.; Sawin, N.M.; Parker, C.P.; Thornell, I.M.; Powers, L.S.; Gansemer, N.D.; Bouzek, D.C.; Cook, D.P.; et al. Gel-forming mucins form distinct morphologic structures in airways. Proc. Natl. Acad. Sci. USA 2017, 114, 6842–6847. [Google Scholar] [CrossRef]
- Ermund, A.; Meiss, L.N.; Rodriguez-Pineiro, A.M.; Bähr, A.; Nilsson, H.E.; Trillo-Muyo, S.; Ridley, C.; Thornton, D.J.; Wine, J.J.; Hebert, H.; et al. The normal trachea is cleaned by MUC5B mucin bundles from the submucosal glands coated with the MUC5AC mucin. Biochem. Biophys. Res. Commun. 2017, 492, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.R.; Boucher, R.C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Investig. 2002, 109, 571–577. [Google Scholar] [CrossRef]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The Interaction between Respiratory Pathogens and Mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef]
- Hiemstra, P.S.; McCray, P.B., Jr.; Bals, R. The innate immune function of airway epithelial cells in inflammatory lung disease. Eur. Respir. J. 2015, 45, 1150–1162. [Google Scholar] [CrossRef]
- Tomazic, P.V.; Darnhofer, B.; Birner-Gruenberger, R. Nasal mucus proteome and its involvement in allergic rhinitis. Expert Rev. Proteom. 2020, 17, 191–199. [Google Scholar] [CrossRef]
- Rogers, D.F. Physiology of airway mucus secretion and pathophysiology of hypersecretion. Respir. Care 2007, 52, 1134–1146; discussion 1146–1139. [Google Scholar]
- Widdicombe, J.H.; Wine, J.J. Airway Gland Structure and Function. Physiol. Rev. 2015, 95, 1241–1319. [Google Scholar] [CrossRef]
- Bonser, L.R.; Erle, D.J. Airway Mucus and Asthma: The Role of MUC5AC and MUC5B. J. Clin. Med. 2017, 6, 112. [Google Scholar] [CrossRef]
- Ma, J.; Rubin, B.K.; Voynow, J.A. Mucins, Mucus, and Goblet Cells. Chest 2018, 154, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Whitsett, J.A. Airway Epithelial Differentiation and Mucociliary Clearance. Ann. Am. Thorac. Soc. 2018, 15, S143–S148. [Google Scholar] [CrossRef] [PubMed]
- Trillo-Muyo, S.; Nilsson, H.E.; Recktenwald, C.V.; Ermund, A.; Ridley, C.; Meiss, L.N.; Bahr, A.; Klymiuk, N.; Wine, J.J.; Koeck, P.J.B.; et al. Granule-stored MUC5B mucins are packed by the non-covalent formation of N-terminal head-to-head tetramers. J. Biol. Chem. 2018, 293, 5746–5754. [Google Scholar] [CrossRef] [PubMed]
- Montoro, D.T.; Haber, A.L.; Biton, M.; Vinarsky, V.; Lin, B.; Birket, S.E.; Yuan, F.; Chen, S.; Leung, H.M.; Villoria, J.; et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 2018, 560, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Scudieri, P.; Musante, I.; Venturini, A.; Guidone, D.; Genovese, M.; Cresta, F.; Caci, E.; Palleschi, A.; Poeta, M.; Santamaria, F.; et al. Ionocytes and CFTR Chloride Channel Expression in Normal and Cystic Fibrosis Nasal and Bronchial Epithelial Cells. Cells 2020, 9, 2090. [Google Scholar] [CrossRef] [PubMed]
- Zaragosi, L.E.; Deprez, M.; Barbry, P. Using single-cell RNA sequencing to unravel cell lineage relationships in the respiratory tract. Biochem. Soc. Trans. 2020, 48, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.B.; Button, B.; Rubinstein, M.; Boucher, R.C. Physiology and pathophysiology of human airway mucus. Physiol. Rev. 2022, 102, 1757–1836. [Google Scholar] [CrossRef] [PubMed]
- Ousingsawat, J.; Centeio, R.; Reyne, N.; McCarron, A.; Cmielewski, P.; Schreiber, R.; diStefano, G.; Romermann, D.; Seidler, U.; Donnelley, M.; et al. Inhibition of mucus secretion by niclosamide and benzbromarone in airways and intestine. Sci. Rep. 2024, 14, 1464. [Google Scholar] [CrossRef]
- Centeio, R.; Ousingsawat, J.; Schreiber, R.; Kunzelmann, K. CLCA1 Regulates Airway Mucus Production and Ion Secretion Through TMEM16A. Int. J. Mol. Sci. 2021, 22, 5133. [Google Scholar] [CrossRef]
- Thornton, D.J.; Carlstedt, I.; Howard, M.; Devine, P.L.; Price, M.R.; Sheehan, J.K. Respiratory mucins: Identification of core proteins and glycoforms. Biochem. J. 1996, 316 Pt 3, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, S.; Sheehan, J.K.; Knight, D.; Richardson, P.S.; Thornton, D.J. Heterogeneity of airways mucus: Variations in the amounts and glycoforms of the major oligomeric mucins MUC5AC and MUC5B. Biochem. J. 2002, 361, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz-Scroggins, M.E.; Yuan, S.; Kerr, S.C.; Dunican, E.M.; Yu, M.; Carrington, S.D.; Fahy, J.V. Abnormalities in MUC5AC and MUC5B Protein in Airway Mucus in Asthma. Am. J. Respir. Crit. Care Med. 2016, 194, 1296–1299. [Google Scholar] [CrossRef]
- Tajiri, T.; Matsumoto, H.; Jinnai, M.; Kanemitsu, Y.; Nagasaki, T.; Iwata, T.; Inoue, H.; Nakaji, H.; Oguma, T.; Ito, I.; et al. Pathophysiological relevance of sputum MUC5AC and MUC5B levels in patients with mild asthma. Allergol. Int. 2022, 71, 193–199. [Google Scholar] [CrossRef]
- Kirkham, S.; Kolsum, U.; Rousseau, K.; Singh, D.; Vestbo, J.; Thornton, D.J. MUC5B is the major mucin in the gel phase of sputum in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2008, 178, 1033–1039. [Google Scholar] [CrossRef]
- Caramori, G.; Di Gregorio, C.; Carlstedt, I.; Casolari, P.; Guzzinati, I.; Adcock, I.M.; Barnes, P.J.; Ciaccia, A.; Cavallesco, G.; Chung, K.F.; et al. Mucin expression in peripheral airways of patients with chronic obstructive pulmonary disease. Histopathology 2004, 45, 477–484. [Google Scholar] [CrossRef]
- Caramori, G.; Casolari, P.; Di Gregorio, C.; Saetta, M.; Baraldo, S.; Boschetto, P.; Ito, K.; Fabbri, L.M.; Barnes, P.J.; Adcock, I.M.; et al. MUC5AC expression is increased in bronchial submucosal glands of stable COPD patients. Histopathology 2009, 55, 321–331. [Google Scholar] [CrossRef]
- Rose, M.C.; Brown, C.F.; Jacoby, J.Z., 3rd; Lynn, W.S.; Kaufman, B. Biochemical properties of tracheobronchial mucins from cystic fibrosis and non-cystic fibrosis individuals. Pediatr. Res. 1987, 22, 545–551. [Google Scholar] [CrossRef]
- Henke, M.O.; Renner, A.; Huber, R.M.; Seeds, M.C.; Rubin, B.K. MUC5AC and MUC5B Mucins Are Decreased in Cystic Fibrosis Airway Secretions. Am. J. Respir. Cell Mol. Biol. 2004, 31, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.G.; Ehre, C.; Button, B.; Abdullah, L.H.; Cai, L.H.; Leigh, M.W.; DeMaria, G.C.; Matsui, H.; Donaldson, S.H.; Davis, C.W.; et al. Cystic fibrosis airway secretions exhibit mucin hyperconcentration and increased osmotic pressure. J. Clin. Investig. 2014, 124, 3047–3060. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; Muhlebach, M.S.; Ehre, C.; Hill, D.B.; Wolfgang, M.C.; Kesimer, M.; Ramsey, K.A.; Markovetz, M.R.; Garbarine, I.C.; Forest, M.G.; et al. Mucus accumulation in the lungs precedes structural changes and infection in children with cystic fibrosis. Sci. Transl. Med. 2019, 11, eaav3488. [Google Scholar] [CrossRef]
- Boucher, R.C. Muco-Obstructive Lung Diseases. N. Engl. J. Med. 2019, 380, 1941–1953. [Google Scholar] [CrossRef]
- Yuan, S.; Hollinger, M.; Lachowicz-Scroggins, M.E.; Kerr, S.C.; Dunican, E.M.; Daniel, B.M.; Ghosh, S.; Erzurum, S.C.; Willard, B.; Hazen, S.L.; et al. Oxidation increases mucin polymer cross-links to stiffen airway mucus gels. Sci. Transl. Med. 2015, 7, 276ra27. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.B.; Long, R.F.; Kissner, W.J.; Atieh, E.; Garbarine, I.C.; Markovetz, M.R.; Fontana, N.C.; Christy, M.; Habibpour, M.; Tarran, R.; et al. Pathological mucus and impaired mucus clearance in cystic fibrosis patients result from increased concentration, not altered pH. Eur. Respir. J. 2018, 52, 1801297. [Google Scholar] [CrossRef]
- Tomaiuolo, G.; Rusciano, G.; Caserta, S.; Carciati, A.; Carnovale, V.; Abete, P.; Sasso, A.; Guido, S. A new method to improve the clinical evaluation of cystic fibrosis patients by mucus viscoelastic properties. PLoS ONE 2014, 9, e82297. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.T.; Tang, C.; Kang, L.; Voynow, J.A.; Rubin, B.K. Cystic Fibrosis Sputum Rheology Correlates With Both Acute and Longitudinal Changes in Lung Function. Chest 2018, 154, 370–377. [Google Scholar] [CrossRef]
- Kato, T.; Radicioni, G.; Papanikolas, M.J.; Stoychev, G.V.; Markovetz, M.R.; Aoki, K.; Porterfield, M.; Okuda, K.; Barbosa Cardenas, S.M.; Gilmore, R.C.; et al. Mucus concentration-dependent biophysical abnormalities unify submucosal gland and superficial airway dysfunction in cystic fibrosis. Sci. Adv. 2022, 8, eabm9718. [Google Scholar] [CrossRef]
- Hoegger, M.J.; Fischer, A.J.; McMenimen, J.D.; Ostedgaard, L.S.; Tucker, A.J.; Awadalla, M.A.; Moninger, T.O.; Michalski, A.S.; Hoffman, E.A.; Zabner, J.; et al. Impaired mucus detachment disrupts mucociliary transport in a piglet model of cystic fibrosis. Science 2014, 345, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Ermund, A.; Meiss, L.N.; Dolan, B.; Bahr, A.; Klymiuk, N.; Hansson, G.C. The mucus bundles responsible for airway cleaning are retained in cystic fibrosis and by cholinergic stimulation. Eur. Respir. J. 2018, 52, 1800457. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.X.; Wen, Q.; Bennett, A.; Kim, B.; Sheils, C.A.; Bucki, R.; Janmey, P.A. Anionic poly(amino acid)s dissolve F-actin and DNA bundles, enhance DNase activity, and reduce the viscosity of cystic fibrosis sputum. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L599–L605. [Google Scholar] [CrossRef] [PubMed]
- Kater, A.; Henke, M.O.; Rubin, B.K. The role of DNA and actin polymers on the polymer structure and rheology of cystic fibrosis sputum and depolymerization by gelsolin or thymosin beta 4. Ann. N. Y Acad. Sci. 2007, 1112, 140–153. [Google Scholar] [CrossRef]
- Charman, J.; Reid, L. Sputum viscosity in chronic bronchitis, bronchiectasis, asthma and cystic fibrosis. Biorheology 1972, 9, 185–199. [Google Scholar] [CrossRef]
- Lopez-Vidriero, M.T.; Reid, L. Chemical markers of mucous and serum glycoproteins and their relation to viscosity in mucoid and purulent sputum from various hypersecretory diseases. Am. Rev. Respir. Dis. 1978, 117, 465–477. [Google Scholar]
- Sheehan, J.K.; Richardson, P.S.; Fung, D.C.; Howard, M.; Thornton, D.J. Analysis of respiratory mucus glycoproteins in asthma: A detailed study from a patient who died in status asthmaticus. Am. J. Respir. Cell Mol. Biol. 1995, 13, 748–756. [Google Scholar] [CrossRef]
- Rogers, D.F.; Evans, T.W. Plasma exudation and oedema in asthma. Br. Med. Bull. 1992, 48, 120–134. [Google Scholar] [CrossRef]
- Shimura, S.; Andoh, Y.; Haraguchi, M.; Shirato, K. Continuity of airway goblet cells and intraluminal mucus in the airways of patients with bronchial asthma. Eur. Respir. J. 1996, 9, 1395–1401. [Google Scholar] [CrossRef]
- Global Initiative for Chronic Obstructive Lung Disease. Global Strategy for Prevention, Diagnosis and Management of COPD (Report 2021). 2020. Available online: https://staging.goldcopd.org/2021-gold-reports/ (accessed on 2 January 2024).
- Kudo, M.; Ishigatsubo, Y.; Aoki, I. Pathology of asthma. Front. Microbiol. 2013, 4, 263. [Google Scholar] [CrossRef]
- Serisier, D.J.; Carroll, M.P.; Shute, J.K.; Young, S.A. Macrorheology of cystic fibrosis, chronic obstructive pulmonary disease & normal sputum. Respir. Res. 2009, 10, 63. [Google Scholar]
- King, M. Is cystic fibrosis mucus abnormal? Pediatr. Res. 1981, 15, 120–122. [Google Scholar] [CrossRef]
- Feather, E.A.; Russell, G. Sputum viscosity in cystic fibrosis of the pancreas and other pulmonary diseases. Br. J. Dis. Chest 1970, 64, 192–200. [Google Scholar] [CrossRef]
- Rogers, D.F. The airway goblet cell. Int. J. Biochem. Cell Biol. 2003, 35, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.F.; Barnes, P.J. Treatment of airway mucus hypersecretion. Ann. Med. 2006, 38, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F. The role of airway smooth muscle in the pathogenesis of airway wall remodeling in chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005, 2, 347–354; discussion 342–371. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, T.; Shimura, S.; Sasaki, H.; Takishima, T.; Yaegashi, H.; Takahashi, T. Morphometric analysis of intraluminal mucus in airways in chronic obstructive pulmonary disease. Am. Rev. Respir. Dis. 1989, 140, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Nagai, A.; West, W.W.; Thurlbeck, W.M. The National Institutes of Health Intermittent Positive-Pressure Breathing trial: Pathology studies. II. Correlation between morphologic findings, clinical findings, and evidence of expiratory air-flow obstruction. Am. Rev. Respir. Dis. 1985, 132, 946–953. [Google Scholar] [PubMed]
- Mullen, J.B.; Wright, J.L.; Wiggs, B.R.; Pare, P.D.; Hogg, J.C. Reassessment of inflammation of airways in chronic bronchitis. Br. Med. J. 1985, 291, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Adcock, I.M. Multifaceted mechanisms in COPD: Inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008, 31, 1334–1356. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, P.K. Remodeling and inflammation of bronchi in asthma and chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2004, 1, 176–183. [Google Scholar] [CrossRef]
- Aikawa, T.; Shimura, S.; Sasaki, H.; Ebina, M.; Takishima, T. Marked goblet cell hyperplasia with mucus accumulation in the airways of patients who died of severe acute asthma attack. Chest 1992, 101, 916–921. [Google Scholar] [CrossRef]
- Rogers, D.F. Airway mucus hypersecretion in asthma: An undervalued pathology? Curr. Opin. Pharmacol. 2004, 4, 241–250. [Google Scholar] [CrossRef]
- Cohn, L. Mechanisms of Mucus Induction in Asthma. In Allergy Frontiers: Clinical Manifestations; Pawankar, R., Holgate, S.T., Rosenwasser, L.J., Eds.; Springer: Tokyo, Japan, 2009; pp. 173–185. [Google Scholar] [CrossRef]
- Glynn, A.A.; Michaels, L. Bronchial biopsy in chronic bronchitis and asthma. Thorax 1960, 15, 142–153. [Google Scholar] [CrossRef]
- Evans, C.M.; Kim, K.; Tuvim, M.J.; Dickey, B.F. Mucus hypersecretion in asthma: Causes and effects. Curr. Opin. Pulm. Med. 2009, 15, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Inglis, S.K.; Wilson, S.M. Cystic fibrosis and airway submucosal glands. Pediatr. Pulmonol. 2005, 40, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Wine, J.J.; Joo, N.S. Submucosal glands and airway defense. Proc. Am. Thorac. Soc. 2004, 1, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Wine, J.J.; Hansson, G.C.; Konig, P.; Joo, N.S.; Ermund, A.; Pieper, M. Progress in understanding mucus abnormalities in cystic fibrosis airways. J. Cyst. Fibros. 2018, 17, S35–S39. [Google Scholar] [CrossRef] [PubMed]
- Dajani, R.; Zhang, Y.; Taft, P.J.; Travis, S.M.; Starner, T.D.; Olsen, A.; Zabner, J.; Welsh, M.J.; Engelhardt, J.F. Lysozyme secretion by submucosal glands protects the airway from bacterial infection. Am. J. Respir. Cell Mol. Biol. 2005, 32, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Tack, B.F.; McCray, P.B., Jr.; Welsh, M.J. Synergistic and additive killing by antimicrobial factors found in human airway surface liquid. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, L799–L805. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.S.; Evans, I.A.; Cho, H.J.; Park, I.H.; Engelhardt, J.F.; Wine, J.J. Proteomic analysis of pure human airway gland mucus reveals a large component of protective proteins. PLoS ONE 2015, 10, e0116756. [Google Scholar] [CrossRef]
- Berkebile, A.R.; Bartlett, J.A.; Abou Alaiwa, M.; Varga, S.M.; Power, U.F.; McCray, P.B., Jr. Airway Surface Liquid Has Innate Antiviral Activity That Is Reduced in Cystic Fibrosis. Am. J. Respir. Cell Mol. Biol. 2020, 62, 104–111. [Google Scholar] [CrossRef]
- Pezzulo, A.A.; Tang, X.X.; Hoegger, M.J.; Alaiwa, M.H.; Ramachandran, S.; Moninger, T.O.; Karp, P.H.; Wohlford-Lenane, C.L.; Haagsman, H.P.; van Eijk, M.; et al. Reduced airway surface pH impairs bacterial killing in the porcine cystic fibrosis lung. Nature 2012, 487, 109–113. [Google Scholar] [CrossRef]
- Rubin, B.K.; Thornton, D.J. Dropping acid: Why is cystic fibrosis mucus abnormal? Eur. Respir. J. 2018, 52, 1802057. [Google Scholar] [CrossRef]
- Morrison, C.B.; Markovetz, M.R.; Ehre, C. Mucus, mucins, and cystic fibrosis. Pediatr. Pulmonol. 2019, 54 (Suppl. S3), S84–S96. [Google Scholar] [CrossRef]
- Kreda, S.M.; Davis, C.W.; Rose, M.C. CFTR, mucins, and mucus obstruction in cystic fibrosis. Cold Spring Harb. Perspect. Med. 2012, 2, a009589. [Google Scholar] [CrossRef]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef]
- Yu, W.; Moninger, T.O.; Thurman, A.L.; Xie, Y.; Jain, A.; Zarei, K.; Powers, L.S.; Pezzulo, A.A.; Stoltz, D.A.; Welsh, M.J. Cellular and molecular architecture of submucosal glands in wild-type and cystic fibrosis pigs. Proc. Natl. Acad. Sci. USA 2022, 119, e2119759119. [Google Scholar] [CrossRef]
- Xie, Y.; Lu, L.; Tang, X.X.; Moninger, T.O.; Huang, T.J.; Stoltz, D.A.; Welsh, M.J. Acidic Submucosal Gland pH and Elevated Protein Concentration Produce Abnormal Cystic Fibrosis Mucus. Dev. Cell 2020, 54, 488–500.e485. [Google Scholar] [CrossRef]
- Voynow, J.A.; Young, L.R.; Wang, Y.; Horger, T.; Rose, M.C.; Fischer, B.M. Neutrophil elastase increases MUC5AC mRNA and protein expression in respiratory epithelial cells. Am. J. Physiol. 1999, 276, L835–L843. [Google Scholar]
- Kohri, K.; Ueki, I.F.; Nadel, J.A. Neutrophil elastase induces mucin production by ligand-dependent epidermal growth factor receptor activation. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, L531–L540. [Google Scholar] [CrossRef]
- Ishimoto, H.; Mukae, H.; Sakamoto, N.; Amenomori, M.; Kitazaki, T.; Imamura, Y.; Fujita, H.; Ishii, H.; Nakayama, S.; Yanagihara, K.; et al. Different effects of telithromycin on MUC5AC production induced by human neutrophil peptide-1 or lipopolysaccharide in NCI-H292 cells compared with azithromycin and clarithromycin. J. Antimicrob. Chemother. 2009, 63, 109–114. [Google Scholar] [CrossRef]
- Fischer, B.M.; Voynow, J.A. Neutrophil elastase induces MUC5AC gene expression in airway epithelium via a pathway involving reactive oxygen species. Am. J. Respir. Cell Mol. Biol. 2002, 26, 447–452. [Google Scholar] [CrossRef]
- Voynow, J.A.; Fischer, B.M.; Malarkey, D.E.; Burch, L.H.; Wong, T.; Longphre, M.; Ho, S.B.; Foster, W.M. Neutrophil elastase induces mucus cell metaplasia in mouse lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 287, L1293–L1302. [Google Scholar] [CrossRef]
- Park, J.A.; He, F.; Martin, L.D.; Li, Y.; Chorley, B.N.; Adler, K.B. Human neutrophil elastase induces hypersecretion of mucin from well-differentiated human bronchial epithelial cells in vitro via a protein kinase Cdelta-mediated mechanism. Am. J. Pathol. 2005, 167, 651–661. [Google Scholar] [CrossRef]
- McAllister, F.; Henry, A.; Kreindler, J.L.; Dubin, P.J.; Ulrich, L.; Steele, C.; Finder, J.D.; Pilewski, J.M.; Carreno, B.M.; Goldman, S.J.; et al. Role of IL-17A, IL-17F, and the IL-17 receptor in regulating growth-related oncogene-alpha and granulocyte colony-stimulating factor in bronchial epithelium: Implications for airway inflammation in cystic fibrosis. J. Immunol. 2005, 175, 404–412. [Google Scholar] [CrossRef]
- Decraene, A.; Willems-Widyastuti, A.; Kasran, A.; De Boeck, K.; Bullens, D.M.; Dupont, L.J. Elevated expression of both mRNA and protein levels of IL-17A in sputum of stable Cystic Fibrosis patients. Respir. Res. 2010, 11, 177. [Google Scholar] [CrossRef]
- Fujisawa, T.; Velichko, S.; Thai, P.; Hung, L.Y.; Huang, F.; Wu, R. Regulation of airway MUC5AC expression by IL-1beta and IL-17A; the NF-kappaB paradigm. J. Immunol. 2009, 183, 6236–6243. [Google Scholar] [CrossRef]
- Chen, Y.; Thai, P.; Zhao, Y.H.; Ho, Y.S.; DeSouza, M.M.; Wu, R. Stimulation of airway mucin gene expression by interleukin (IL)-17 through IL-6 paracrine/autocrine loop. J. Biol. Chem. 2003, 278, 17036–17043. [Google Scholar] [CrossRef]
- Fujisawa, T.; Chang, M.M.; Velichko, S.; Thai, P.; Hung, L.Y.; Huang, F.; Phuong, N.; Chen, Y.; Wu, R. NF-kappaB mediates IL-1beta- and IL-17A-induced MUC5B expression in airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2011, 45, 246–252. [Google Scholar] [CrossRef]
- Chen, G.; Sun, L.; Kato, T.; Okuda, K.; Martino, M.B.; Abzhanova, A.; Lin, J.M.; Gilmore, R.C.; Batson, B.D.; O’Neal, Y.K.; et al. IL-1beta dominates the promucin secretory cytokine profile in cystic fibrosis. J. Clin. Invest. 2019, 129, 4433–4450. [Google Scholar] [CrossRef]
- Dransfield, M.T.; Wilhelm, A.M.; Flanagan, B.; Courville, C.; Tidwell, S.L.; Raju, S.V.; Gaggar, A.; Steele, C.; Tang, L.P.; Liu, B.; et al. Acquired cystic fibrosis transmembrane conductance regulator dysfunction in the lower airways in COPD. Chest 2013, 144, 498–506. [Google Scholar] [CrossRef]
- Cantin, A.M. Cystic Fibrosis Transmembrane Conductance Regulator. Implications in Cystic Fibrosis and Chronic Obstructive Pulmonary Disease. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. S2), S150–S155. [Google Scholar]
- Mall, M.A.; Criner, G.J.; Miravitlles, M.; Rowe, S.M.; Vogelmeier, C.F.; Rowlands, D.J.; Schoenberger, M.; Altman, P. Cystic fibrosis transmembrane conductance regulator in COPD: A role in respiratory epithelium and beyond. Eur. Respir. J. 2023, 61, 2201307. [Google Scholar] [CrossRef]
- Takeyama, K.; Jung, B.; Shim, J.J.; Burgel, P.R.; Dao-Pick, T.; Ueki, I.F.; Protin, U.; Kroschel, P.; Nadel, J.A. Activation of epidermal growth factor receptors is responsible for mucin synthesis induced by cigarette smoke. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L165–L172. [Google Scholar] [CrossRef]
- Chen, R.; Liang, Y.; Ip, M.S.M.; Zhang, K.Y.; Mak, J.C.W. Amelioration of Cigarette Smoke-Induced Mucus Hypersecretion and Viscosity by Dendrobium officinale Polysaccharides In Vitro and In Vivo. Oxid. Med. Cell Longev. 2020, 2020, 8217642. [Google Scholar] [CrossRef]
- Guddo, F.; Vignola, A.M.; Saetta, M.; Baraldo, S.; Siena, L.; Balestro, E.; Zuin, R.; Papi, A.; Maestrelli, P.; Fabbri, L.M.; et al. Upregulation of basic fibroblast growth factor in smokers with chronic bronchitis. Eur. Respir. J. 2006, 27, 957–963. [Google Scholar] [CrossRef]
- Kim, S.; Nadel, J.A. Role of neutrophils in mucus hypersecretion in COPD and implications for therapy. Treat. Respir. Med. 2004, 3, 147–159. [Google Scholar] [CrossRef]
- Yang, R.; Wu, X.; Gounni, A.S.; Xie, J. Mucus hypersecretion in chronic obstructive pulmonary disease: From molecular mechanisms to treatment. J. Transl. Int. Med. 2023, 11, 312–315. [Google Scholar] [CrossRef]
- Raby, K.L.; Michaeloudes, C.; Tonkin, J.; Chung, K.F.; Bhavsar, P.K. Mechanisms of airway epithelial injury and abnormal repair in asthma and COPD. Front. Immunol. 2023, 14, 1201658. [Google Scholar] [CrossRef]
- Amishima, M.; Munakata, M.; Nasuhara, Y.; Sato, A.; Takahashi, T.; Homma, Y.; Kawakami, Y. Expression of epidermal growth factor and epidermal growth factor receptor immunoreactivity in the asthmatic human airway. Am. J. Respir. Crit. Care Med. 1998, 157, 1907–1912. [Google Scholar] [CrossRef]
- Fedorov, I.A.; Wilson, S.J.; Davies, D.E.; Holgate, S.T. Epithelial stress and structural remodelling in childhood asthma. Thorax 2005, 60, 389–394. [Google Scholar] [CrossRef]
- Shim, J.J.; Dabbagh, K.; Ueki, I.F.; Dao-Pick, T.; Burgel, P.R.; Takeyama, K.; Tam, D.C.; Nadel, J.A. IL-13 induces mucin production by stimulating epidermal growth factor receptors and by activating neutrophils. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L134–L140. [Google Scholar] [CrossRef]
- Zhen, G.; Park, S.W.; Nguyenvu, L.T.; Rodriguez, M.W.; Barbeau, R.; Paquet, A.C.; Erle, D.J. IL-13 and epidermal growth factor receptor have critical but distinct roles in epithelial cell mucin production. Am. J. Respir. Cell Mol. Biol. 2007, 36, 244–253. [Google Scholar] [CrossRef]
- Izuhara, K.; Ohta, S.; Shiraishi, H.; Suzuki, S.; Taniguchi, K.; Toda, S.; Tanabe, T.; Yasuo, M.; Kubo, K.; Hoshino, T.; et al. The mechanism of mucus production in bronchial asthma. Curr. Med. Chem. 2009, 16, 2867–2875. [Google Scholar] [CrossRef]
- Fahy, J.V. Goblet cell and mucin gene abnormalities in asthma. Chest 2002, 122, 320S–326S. [Google Scholar] [CrossRef]
- Komlosi, Z.I.; van de Veen, W.; Kovacs, N.; Szucs, G.; Sokolowska, M.; O’Mahony, L.; Akdis, M.; Akdis, C.A. Cellular and molecular mechanisms of allergic asthma. Mol. Aspects Med. 2022, 85, 100995. [Google Scholar] [CrossRef]
- Laoukili, J.; Perret, E.; Willems, T.; Minty, A.; Parthoens, E.; Houcine, O.; Coste, A.; Jorissen, M.; Marano, F.; Caput, D.; et al. IL-13 alters mucociliary differentiation and ciliary beating of human respiratory epithelial cells. J. Clin. Investig. 2001, 108, 1817–1824. [Google Scholar] [CrossRef]
- Kuperman, D.A.; Huang, X.; Koth, L.L.; Chang, G.H.; Dolganov, G.M.; Zhu, Z.; Elias, J.A.; Sheppard, D.; Erle, D.J. Direct effects of interleukin-13 on epithelial cells cause airway hyperreactivity and mucus overproduction in asthma. Nat. Med. 2002, 8, 885–889. [Google Scholar] [CrossRef]
- Seibold, M.A. Interleukin-13 Stimulation Reveals the Cellular and Functional Plasticity of the Airway Epithelium. Ann. Am. Thorac. Soc. 2018, 15, S98–S102. [Google Scholar] [CrossRef]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid. Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Allen, A. Structure and function of gastrointestinal mucus. In Physiology of the Gastroenterology Tract, 1st ed.; Johnson, L., Ed.; Raven Press: New York, NY, USA, 1981; pp. 617–639. [Google Scholar]
- Williams, O.W.; Sharafkhaneh, A.; Kim, V.; Dickey, B.F.; Evans, C.M. Airway mucus: From production to secretion. Am. J. Respir. Cell Mol. Biol. 2006, 34, 527–536. [Google Scholar] [CrossRef]
- Dekker, J.; Rossen, J.W.; Buller, H.A.; Einerhand, A.W. The MUC family: An obituary. Trends Biochem. Sci. 2002, 27, 126–131. [Google Scholar] [CrossRef]
- Sheehan, J.K.; Kirkham, S.; Howard, M.; Woodman, P.; Kutay, S.; Brazeau, C.; Buckley, J.; Thornton, D.J. Identification of molecular intermediates in the assembly pathway of the MUC5AC mucin. J. Biol. Chem. 2004, 279, 15698–15705. [Google Scholar] [CrossRef]
- Rajendran, R.R.; Banerjee, A. Mucus transport and distribution by steady expiration in an idealized airway geometry. Med. Eng. Phys. 2019, 66, 26–39. [Google Scholar] [CrossRef]
- Manolidis, M.; Isabey, D.; Louis, B.; Grotberg, J.B.; Filoche, M. A Macroscopic Model for Simulating the Mucociliary Clearance in a Bronchial Bifurcation: The Role of Surface Tension. J. Biomech. Eng. 2016, 138, 121005. [Google Scholar] [CrossRef]
- Olivenca, D.V.; Fonseca, L.L.; Voit, E.O.; Pinto, F.R. Thickness of the airway surface liquid layer in the lung is affected in cystic fibrosis by compromised synergistic regulation of the ENaC ion channel. J. R. Soc. Interface 2019, 16, 20190187. [Google Scholar] [CrossRef]
- Ren, S.; Li, W.; Wang, L.; Shi, Y.; Cai, M.; Hao, L.; Luo, Z.; Niu, J.; Xu, W.; Luo, Z. Numerical Analysis of Airway Mucus Clearance Effectiveness Using Assisted Coughing Techniques. Sci. Rep. 2020, 10, 2030. [Google Scholar] [CrossRef]
- Derichs, N.; Jin, B.J.; Song, Y.; Finkbeiner, W.E.; Verkman, A.S. Hyperviscous airway periciliary and mucous liquid layers in cystic fibrosis measured by confocal fluorescence photobleaching. FASEB J. 2011, 25, 2325–2332. [Google Scholar] [CrossRef]
- Button, B.; Cai, L.H.; Ehre, C.; Kesimer, M.; Hill, D.B.; Sheehan, J.K.; Boucher, R.C.; Rubinstein, M. A periciliary brush promotes the lung health by separating the mucus layer from airway epithelia. Science 2012, 337, 937–941. [Google Scholar] [CrossRef]
- Bartlett, B.A.; Feng, Y.; Fromen, C.A.; Ford Versypt, A.N. Computational fluid dynamics modeling of aerosol particle transport through lung airway mucosa. Comput. Chem. Eng. 2023, 179, 108458. [Google Scholar] [CrossRef]
- Shah, B.K.; Singh, B.; Wang, Y.; Xie, S.; Wang, C. Mucus Hypersecretion in Chronic Obstructive Pulmonary Disease and Its Treatment. Mediat. Inflamm. 2023, 2023, 8840594. [Google Scholar] [CrossRef]
- Liu, X.; Driskell, R.R.; Engelhardt, J.F. Stem cells in the lung. Methods Enzymol. 2006, 419, 285–321. [Google Scholar]
- Fischer, B.; Voynow, J. Neutrophil elastase induces MUC5AC messenger RNA expression by an oxidant-dependent mechanism. Chest 2000, 117, 317S–320S. [Google Scholar] [CrossRef]
- Okuda, K.; Chen, G.; Subramani, D.B.; Wolf, M.; Gilmore, R.C.; Kato, T.; Radicioni, G.; Kesimer, M.; Chua, M.; Dang, H.; et al. Localization of Secretory Mucins MUC5AC and MUC5B in Normal/Healthy Human Airways. Am. J. Respir. Crit. Care Med. 2019, 199, 715–727. [Google Scholar] [CrossRef]
- Chateau, S.; D’Ortona, U.; Poncet, S.; Favier, J. Transport and Mixing Induced by Beating Cilia in Human Airways. Front. Physiol. 2018, 9, 161. [Google Scholar] [CrossRef]
- Abrami, M.; Maschio, M.; Conese, M.; Confalonieri, M.; Gerin, F.; Dapas, B.; Farra, R.; Adrover, A.; Torelli, L.; Ruaro, B.; et al. Combined use of rheology and portable low-field NMR in cystic fibrosis patients. Respir. Med. 2021, 189, 106623. [Google Scholar] [CrossRef]
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Batson, B.D.; Zorn, B.T.; Radicioni, G.; Livengood, S.S.; Kumagai, T.; Dang, H.; Ceppe, A.; Clapp, P.W.; Tunney, M.; Elborn, J.S.; et al. Cystic Fibrosis Airway Mucus Hyperconcentration Produces a Vicious Cycle of Mucin, Pathogen, and Inflammatory Interactions that Promotes Disease Persistence. Am. J. Respir. Cell Mol. Biol. 2022, 67, 253–265. [Google Scholar] [CrossRef]
- Button, B.; Goodell, H.P.; Atieh, E.; Chen, Y.C.; Williams, R.; Shenoy, S.; Lackey, E.; Shenkute, N.T.; Cai, L.H.; Dennis, R.G.; et al. Roles of mucus adhesion and cohesion in cough clearance. Proc. Natl. Acad. Sci. USA 2018, 115, 12501–12506. [Google Scholar] [CrossRef]
- Abrami, M.; Maschio, M.; Conese, M.; Confalonieri, M.; Di Gioia, S.; Gerin, F.; Dapas, B.; Tonon, F.; Farra, R.; Murano, E.; et al. Use of low field nuclear magnetic resonance to monitor lung inflammation and the amount of pathological components in the sputum of cystic fibrosis patients. Magn. Reson. Med. 2020, 84, 427–436. [Google Scholar] [CrossRef]
- Abrami, M.; Maschio, M.; Conese, M.; Confalonieri, M.; Salton, F.; Gerin, F.; Dapas, B.; Farra, R.; Adrover, A.; Milcovich, G.; et al. Effect of chest physiotherapy on cystic fibrosis sputum nanostructure: An experimental and theoretical approach. Drug Deliv. Transl. Res. 2022, 12, 1943–1958. [Google Scholar] [CrossRef]
- Abrami, M.; Ascenzioni, F.; Di Domenico, E.G.; Maschio, M.; Ventura, A.; Confalonieri, M.; Di Gioia, S.; Conese, M.; Dapas, B.; Grassi, G.; et al. A novel approach based on low-field NMR for the detection of the pathological components of sputum in cystic fibrosis patients. Magn. Reson. Med. 2018, 79, 2323–2331. [Google Scholar] [CrossRef]
- Chui, M.M.; Phillips, R.J.; McCarthy, M.J. Measurement of the Porous Microstructure of Hydrogels by Nuclear Magnetic Resonance. J. Colloid. Interface Sci. 1995, 174, 336–344. [Google Scholar] [CrossRef]
- Suh, J.; Dawson, M.; Hanes, J. Real-time multiple-particle tracking: Applications to drug and gene delivery. Adv. Drug Deliv. Rev. 2005, 57, 63–78. [Google Scholar] [CrossRef]
- Duncan, G.A.; Jung, J.; Joseph, A.; Thaxton, A.L.; West, N.E.; Boyle, M.P.; Hanes, J.; Suk, J.S. Microstructural alterations of sputum in cystic fibrosis lung disease. JCI Insight 2016, 1, e88198. [Google Scholar] [CrossRef]
- National Heart, Lung and Blood Institute. Morbidity and Mortality: 2007 Chartbook on Cardiovascular, Lung, and Blood Disease; National Institutes of Health: Bethesda, MD, USA, 2007. [Google Scholar]
- Woodruff, P.G.; Agusti, A.; Roche, N.; Singh, D.; Martinez, F.J. Current concepts in targeting chronic obstructive pulmonary disease pharmacotherapy: Making progress towards personalised management. Lancet 2015, 385, 1789–1798. [Google Scholar] [CrossRef]
- Raju, S.V.; Solomon, G.M.; Dransfield, M.T.; Rowe, S.M. Acquired Cystic Fibrosis Transmembrane Conductance Regulator Dysfunction in Chronic Bronchitis and Other Diseases of Mucus Clearance. Clin. Chest Med. 2016, 37, 147–158. [Google Scholar] [CrossRef]
- Chisholm, J.F.; Shenoy, S.K.; Shade, J.K.; Kim, V.; Putcha, N.; Carson, K.A.; Wise, R.; Hansel, N.N.; Hanes, J.S.; Suk, J.S.; et al. Nanoparticle diffusion in spontaneously expectorated sputum as a biophysical tool to probe disease severity in COPD. Eur. Respir. J. 2019, 54, 1900088. [Google Scholar] [CrossRef]
- Linssen, R.S.; Chai, G.; Ma, J.; Kummarapurugu, A.B.; van Woensel, J.B.M.; Bem, R.A.; Kaler, L.; Duncan, G.A.; Zhou, L.; Rubin, B.K.; et al. Neutrophil Extracellular Traps Increase Airway Mucus Viscoelasticity and Slow Mucus Particle Transit. Am. J. Respir. Cell Mol. Biol. 2021, 64, 69–78. [Google Scholar] [CrossRef]
- Thulborn, S.J.; Mistry, V.; Brightling, C.E.; Moffitt, K.L.; Ribeiro, D.; Bafadhel, M. Neutrophil elastase as a biomarker for bacterial infection in COPD. Respir. Res. 2019, 20, 170. [Google Scholar] [CrossRef]
- Solomon, G.M.; Fu, L.; Rowe, S.M.; Collawn, J.F. The therapeutic potential of CFTR modulators for COPD and other airway diseases. Curr. Opin. Pharmacol. 2017, 34, 132–139. [Google Scholar] [CrossRef]
- Schulz, A.; Tummler, B. Non-allergic asthma as a CFTR-related disorder. J. Cyst. Fibros. 2016, 15, 641–644. [Google Scholar] [CrossRef]
- Innes, A.L.; Carrington, S.D.; Thornton, D.J.; Kirkham, S.; Rousseau, K.; Dougherty, R.H.; Raymond, W.W.; Caughey, G.H.; Muller, S.J.; Fahy, J.V. Ex vivo sputum analysis reveals impairment of protease-dependent mucus degradation by plasma proteins in acute asthma. Am. J. Respir. Crit. Care Med. 2009, 180, 203–210. [Google Scholar] [CrossRef]
- Morgan, L.E.; Jaramillo, A.M.; Shenoy, S.K.; Raclawska, D.; Emezienna, N.A.; Richardson, V.L.; Hara, N.; Harder, A.Q.; NeeDell, J.C.; Hennessy, C.E.; et al. Disulfide disruption reverses mucus dysfunction in allergic airway disease. Nat. Commun. 2021, 12, 249. [Google Scholar] [CrossRef]
- Volpato, M.; Vialaret, J.; Hirtz, C.; Petit, A.; Suehs, C.; Patarin, J.; Matzner-Lober, E.; Vachier, I.; Molinari, N.; Bourdin, A.; et al. Rheology predicts sputum eosinophilia in patients with muco-obstructive lung diseases. Biochem. Biophys. Res. Commun. 2022, 622, 64–71. [Google Scholar] [CrossRef]
- Cone, R.A. Barrier properties of mucus. Adv. Drug Deliv. Rev. 2009, 61, 75–85. [Google Scholar] [CrossRef]
- Rubin, B.K. The role of mucus in cough research. Lung 2010, 188 (Suppl. S1), S69–S72. [Google Scholar] [CrossRef]
- Ambort, D.; Johansson, M.E.; Gustafsson, J.K.; Nilsson, H.E.; Ermund, A.; Johansson, B.R.; Koeck, P.J.; Hebert, H.; Hansson, G.C. Calcium and pH-dependent packing and release of the gel-forming MUC2 mucin. Proc. Natl. Acad. Sci. USA 2012, 109, 5645–5650. [Google Scholar] [CrossRef]
- Philippe, A.-M.; Cipelletti, L.; Larobina, D. Mucus as an Arrested Phase Separation Gel. Macromolecules 2017, 50, 8221–8230. [Google Scholar] [CrossRef]
- Pednekar, D.D.; Liguori, M.A.; Marques, C.N.H.; Zhang, T.; Zhang, N.; Zhou, Z.; Amoako, K.; Gu, H. From Static to Dynamic: A Review on the Role of Mucus Heterogeneity in Particle and Microbial Transport. ACS Biomater. Sci. Eng. 2022, 8, 2825–2848. [Google Scholar] [CrossRef]
- Wang, J.; Shi, X. Molecular dynamics simulation of diffusion of nanoparticles in mucus. Acta Mech. Solida Sin. 2017, 30, 241–247. [Google Scholar] [CrossRef]
- Olmsted, S.S.; Padgett, J.L.; Yudin, A.I.; Whaley, K.J.; Moench, T.R.; Cone, R.A. Diffusion of macromolecules and virus-like particles in human cervical mucus. Biophys. J. 2001, 81, 1930–1937. [Google Scholar] [CrossRef]
- Dawson, M.; Wirtz, D.; Hanes, J. Enhanced viscoelasticity of human cystic fibrotic sputum correlates with increasing microheterogeneity in particle transport. J. Biol. Chem. 2003, 278, 50393–50401. [Google Scholar] [CrossRef]
- Larobina, D.; Pommella, A.; Philippe, A.M.; Nagazi, M.Y.; Cipelletti, L. Enhanced microscopic dynamics in mucus gels under a mechanical load in the linear viscoelastic regime. Proc. Natl. Acad. Sci. USA 2021, 118, e2103995118. [Google Scholar] [CrossRef]
- Ghanem, R.; Roquefort, P.; Ramel, S.; Laurent, V.; Haute, T.; Le Gall, T.; Aubry, T.; Montier, T. Apparent Yield Stress of Sputum as a Relevant Biomarker in Cystic Fibrosis. Cells 2021, 10, 3107. [Google Scholar] [CrossRef]
- Suk, J.S.; Lai, S.K.; Wang, Y.Y.; Ensign, L.M.; Zeitlin, P.L.; Boyle, M.P.; Hanes, J. The penetration of fresh undiluted sputum expectorated by cystic fibrosis patients by non-adhesive polymer nanoparticles. Biomaterials 2009, 30, 2591–2597. [Google Scholar] [CrossRef]
- Sanders, N.N.; De Smedt, S.C.; Van Rompaey, E.; Simoens, P.; De Baets, F.; Demeester, J. Cystic fibrosis sputum. A barrier to the transport of nanospheres. Am. J. Respir. crit. Care Med. 2000, 162, 1905–1911. [Google Scholar] [CrossRef]
- Meldrum, O.W.; Yakubov, G.E.; Bonilla, M.R.; Deshmukh, O.; McGuckin, M.A.; Gidley, M.J. Mucin gel assembly is controlled by a collective action of non-mucin proteins, disulfide bridges, Ca(2+)-mediated links, and hydrogen bonding. Sci. Rep. 2018, 8, 5802. [Google Scholar] [CrossRef]
- Broughton-Head, V.J.; Shur, J.; Carroll, M.P.; Smith, J.R.; Shute, J.K. Unfractionated heparin reduces the elasticity of sputum from patients with cystic fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L1240–L1249. [Google Scholar] [CrossRef]
- Rubin, B.K. Mucus structure and properties in cystic fibrosis. Paediatr. Respir. Rev. 2007, 8, 4–7. [Google Scholar] [CrossRef]
- Voynow, J.A.; Gendler, S.J.; Rose, M.C. Regulation of mucin genes in chronic inflammatory airway diseases. Am. J. Respir. Cell Mol. Biol. 2006, 34, 661–665. [Google Scholar] [CrossRef]
- Rubin, B.K.; Ramirez, O.; King, M. Mucus rheology and transport in neonatal respiratory distress syndrome and the effect of surfactant therapy. Chest 1992, 101, 1080–1085. [Google Scholar] [CrossRef]
- Cone, R.A. Mucus. In Mucosal Immunology; Mestecky, J., Lamm, M.E., Ogra, P.L., Strober, W., Bienenstock, J., McGhee, J.R., Mayer, L., Eds.; Elsevier Academic Press: Burlington, MA, USA; San Diego, CA, USA; London, UK, 2005; pp. 49–72. [Google Scholar]
- Hattori, M.; Majima, Y.; Ukai, K.; Sakakura, Y. Effects of nasal allergen challenge on dynamic viscoelasticity of nasal mucus. Ann. Otol. Rhinol. Laryngol. 1993, 102, 314–317. [Google Scholar] [CrossRef]
- Rubin, B.K.; Ramirez, O.; Zayas, J.G.; Finegan, B.; King, M. Respiratory mucus from asymptomatic smokers is better hydrated and more easily cleared by mucociliary action. Am. Rev. Respir. Dis. 1992, 145, 545–547. [Google Scholar] [CrossRef]
- Zayas, J.G.; Man, G.C.; King, M. Tracheal mucus rheology in patients undergoing diagnostic bronchoscopy. Interrelations with smoking and cancer. Am. Rev. Respir. Dis. 1990, 141, 1107–1113. [Google Scholar] [CrossRef]
- Walker, P.S.; Conway, J.H.; Fleming, J.S.; Bondesson, E.; Borgstrom, L. Pulmonary clearance rate of two chemically different forms of inhaled pertechnetate. J. Aerosol Med. 2001, 14, 209–215. [Google Scholar] [CrossRef]
- Norton, M.M.; Robinson, R.J.; Weinstein, S.J. Model of ciliary clearance and the role of mucus rheology. Phys. Rev. E Stat. Nonlin Soft Matter Phys. 2011, 83, 011921. [Google Scholar] [CrossRef]
- King, M. The role of mucus viscoelasticity in cough clearance. Biorheology 1987, 24, 589–597. [Google Scholar] [CrossRef]
- Chatelin, R.; Anne-Archard, D.; Murris-Espin, M.; Thiriet, M.; Poncet, P. Numerical and experimental investigation of mucociliary clearance breakdown in cystic fibrosis. J. Biomech. 2017, 53, 56–63. [Google Scholar] [CrossRef]
- Anderson, W.H.; Coakley, R.D.; Button, B.; Henderson, A.G.; Zeman, K.L.; Alexis, N.E.; Peden, D.B.; Lazarowski, E.R.; Davis, C.W.; Bailey, S.; et al. The Relationship of Mucus Concentration (Hydration) to Mucus Osmotic Pressure and Transport in Chronic Bronchitis. Am. J. Respir. Crit. Care Med. 2015, 192, 182–190. [Google Scholar] [CrossRef]
- Parlato, R.M.; Greco, F.; Maffettone, P.L.; Larobina, D. Effect of pH on the viscoelastic properties of pig gastric mucus. J. Mech. Behav. Biomed. Mater. 2019, 98, 195–199. [Google Scholar] [CrossRef]
- Tang, X.X.; Ostedgaard, L.S.; Hoegger, M.J.; Moninger, T.O.; Karp, P.H.; McMenimen, J.D.; Choudhury, B.; Varki, A.; Stoltz, D.A.; Welsh, M.J. Acidic pH increases airway surface liquid viscosity in cystic fibrosis. J. Clin. Investig. 2016, 126, 879–891. [Google Scholar] [CrossRef]
- Zajac, M.; Dreano, E.; Edwards, A.; Planelles, G.; Sermet-Gaudelus, I. Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge. Int. J. Mol. Sci. 2021, 22, 3384. [Google Scholar] [CrossRef]
- Ambort, D.; van der Post, S.; Johansson, M.E.; Mackenzie, J.; Thomsson, E.; Krengel, U.; Hansson, G.C. Function of the CysD domain of the gel-forming MUC2 mucin. Biochem. J. 2011, 436, 61–70. [Google Scholar] [CrossRef]
- Nadel, J.A. Mucous hypersecretion and relationship to cough. Pulm. Pharmacol. Ther. 2013, 26, 510–513. [Google Scholar] [CrossRef]
- Nystrom, B.; Kjoniksen, A.L.; Beheshti, N.; Maleki, A.; Zhu, K.; Knudsen, K.D.; Pamies, R.; Hernandez Cifre, J.G.; Garcia de la Torre, J. Characterization of polyelectrolyte features in polysaccharide systems and mucin. Adv. Colloid. Interface Sci. 2010, 158, 108–118. [Google Scholar] [CrossRef]
- Quinton, P.M. Cystic fibrosis: Impaired bicarbonate secretion and mucoviscidosis. Lancet 2008, 372, 415–417. [Google Scholar] [CrossRef]
- Goralski, J.L.; Wu, D.; Thelin, W.R.; Boucher, R.C.; Button, B. The in vitro effect of nebulised hypertonic saline on human bronchial epithelium. Eur. Respir. J. 2018, 51, 1702652. [Google Scholar] [CrossRef]
- Stoltz, D.A.; Meyerholz, D.K.; Pezzulo, A.A.; Ramachandran, S.; Rogan, M.P.; Davis, G.J.; Hanfland, R.A.; Wohlford-Lenane, C.; Dohrn, C.L.; Bartlett, J.A.; et al. Cystic fibrosis pigs develop lung disease and exhibit defective bacterial eradication at birth. Sci. Transl. Med. 2010, 2, 29ra31. [Google Scholar] [CrossRef]
- Birket, S.E.; Davis, J.M.; Fernandez, C.M.; Tuggle, K.L.; Oden, A.M.; Chu, K.K.; Tearney, G.J.; Fanucchi, M.V.; Sorscher, E.J.; Rowe, S.M. Development of an airway mucus defect in the cystic fibrosis rat. JCI Insight 2018, 3, e97199. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, S.H.; Bennett, W.D.; Zeman, K.L.; Knowles, M.R.; Tarran, R.; Boucher, R.C. Mucus clearance and lung function in cystic fibrosis with hypertonic saline. N. Engl. J. Med. 2006, 354, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Geitani, R.; Moubareck, C.A.; Xu, Z.; Karam Sarkis, D.; Touqui, L. Expression and Roles of Antimicrobial Peptides in Innate Defense of Airway Mucosa: Potential Implication in Cystic Fibrosis. Front. Immunol. 2020, 11, 1198. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Antimicrobial polypeptides in host defense of the respiratory tract. J. Clin. Investig. 2002, 109, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Comstock, A.T.; Sajjan, U.S. Barrier function of airway tract epithelium. Tissue Barriers 2013, 1, e24997. [Google Scholar] [CrossRef]
- Lecaille, F.; Lalmanach, G.; Andrault, P.M. Antimicrobial proteins and peptides in human lung diseases: A friend and foe partnership with host proteases. Biochimie 2016, 122, 151–168. [Google Scholar] [CrossRef]
- Felgentreff, K.; Beisswenger, C.; Griese, M.; Gulder, T.; Bringmann, G.; Bals, R. The antimicrobial peptide cathelicidin interacts with airway mucus. Peptides 2006, 27, 3100–3106. [Google Scholar] [CrossRef]
- Herasimenka, Y.; Benincasa, M.; Mattiuzzo, M.; Cescutti, P.; Gennaro, R.; Rizzo, R. Interaction of antimicrobial peptides with bacterial polysaccharides from lung pathogens. Peptides 2005, 26, 1127–1132. [Google Scholar] [CrossRef]
- Weiner, D.J.; Bucki, R.; Janmey, P.A. The antimicrobial activity of the cathelicidin LL37 is inhibited by F-actin bundles and restored by gelsolin. Am. J. Respir. Cell Mol. Biol. 2003, 28, 738–745. [Google Scholar] [CrossRef]
- Xiao, W.; Hsu, Y.P.; Ishizaka, A.; Kirikae, T.; Moss, R.B. Sputum cathelicidin, urokinase plasminogen activation system components, and cytokines discriminate cystic fibrosis, COPD, and asthma inflammation. Chest 2005, 128, 2316–2326. [Google Scholar] [CrossRef]
- Bucki, R.; Byfield, F.J.; Janmey, P.A. Release of the antimicrobial peptide LL-37 from DNA/F-actin bundles in cystic fibrosis sputum. Eur. Respir. J. 2007, 29, 624–632. [Google Scholar] [CrossRef]
- Bergsson, G.; Reeves, E.P.; McNally, P.; Chotirmall, S.H.; Greene, C.M.; Greally, P.; Murphy, P.; O’Neill, S.J.; McElvaney, N.G. LL-37 complexation with glycosaminoglycans in cystic fibrosis lungs inhibits antimicrobial activity, which can be restored by hypertonic saline. J. Immunol. 2009, 183, 543–551. [Google Scholar] [CrossRef]
- Thompson, A.B.; Bohling, T.; Payvandi, F.; Rennard, S.I. Lower respiratory tract lactoferrin and lysozyme arise primarily in the airways and are elevated in association with chronic bronchitis. J. Lab. Clin. Med. 1990, 115, 148–158. [Google Scholar]
- Golec, M.; Reichel, C.; Mackiewicz, B.; Skorska, C.; Curzytek, K.; Lemieszek, M.; Dutkiewicz, J.; Gora, A.; Ziesche, R.; Boltuc, J.; et al. Cathelicidin LL-37, granzymes, TGF-beta1 and cytokines levels in induced sputum from farmers with and without COPD. Ann. Agric. Environ. Med. 2009, 16, 289–297. [Google Scholar]
- Jiang, Y.Y.; Xiao, W.; Zhu, M.X.; Yang, Z.H.; Pan, X.J.; Zhang, Y.; Sun, C.C.; Xing, Y. The effect of human antibacterial peptide LL-37 in the pathogenesis of chronic obstructive pulmonary disease. Respir. Med. 2012, 106, 1680–1689. [Google Scholar] [CrossRef]
- Tsoumakidou, M.; Bouloukaki, I.; Thimaki, K.; Tzanakis, N.; Siafakas, N.M. Innate immunity proteins in chronic obstructive pulmonary disease and idiopathic pulmonary fibrosis. Exp. Lung Res. 2010, 36, 373–380. [Google Scholar] [CrossRef]
- Persson, L.J.; Aanerud, M.; Hardie, J.A.; Miodini Nilsen, R.; Bakke, P.S.; Eagan, T.M.; Hiemstra, P.S. Antimicrobial peptide levels are linked to airway inflammation, bacterial colonisation and exacerbations in chronic obstructive pulmonary disease. Eur. Respir. J. 2017, 49, 1601328. [Google Scholar] [CrossRef]
- Hill, A.T.; Campbell, E.J.; Bayley, D.L.; Hill, S.L.; Stockley, R.A. Evidence for excessive bronchial inflammation during an acute exacerbation of chronic obstructive pulmonary disease in patients with alpha(1)-antitrypsin deficiency (PiZ). Am. J. Respir. Crit. Care Med. 1999, 160, 1968–1975. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, G.I.; Wrona, C.T.; Murphy, T.F.; Sethi, S. Moraxella catarrhalis acquisition, airway inflammation and protease-antiprotease balance in chronic obstructive pulmonary disease. BMC Infect. Dis. 2009, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, G.I.; Sethi, S.; Murphy, T.F. Effects of bacterial infection on airway antimicrobial peptides and proteins in COPD. Chest 2011, 140, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Mallia, P.; Footitt, J.; Sotero, R.; Jepson, A.; Contoli, M.; Trujillo-Torralbo, M.B.; Kebadze, T.; Aniscenko, J.; Oleszkiewicz, G.; Gray, K.; et al. Rhinovirus infection induces degradation of antimicrobial peptides and secondary bacterial infection in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2012, 186, 1117–1124. [Google Scholar] [CrossRef]
- Baines, K.J.; Wright, T.K.; Simpson, J.L.; McDonald, V.M.; Wood, L.G.; Parsons, K.S.; Wark, P.A.; Gibson, P.G. Airway beta-Defensin-1 Protein Is Elevated in COPD and Severe Asthma. Mediators Inflamm. 2015, 2015, 407271. [Google Scholar] [CrossRef]
- Sagel, S.D.; Chmiel, J.F.; Konstan, M.W. Sputum biomarkers of inflammation in cystic fibrosis lung disease. Proc. Am. Thorac. Soc. 2007, 4, 406–417. [Google Scholar] [CrossRef]
- Conese, M.; Tirelli, A.S.; Alicandro, G.; Di Gioia, S.; Carbone, A.; Castellani, S.; Colombo, C. Biomarkers of Inflammation and Remodelling in Cystic Fibrosis. Clin. Immunol. Endocr. Metab. Drugs 2016, 3, 92–108. [Google Scholar] [CrossRef]
- Lepissier, A.; Addy, C.; Hayes, K.; Noel, S.; Bui, S.; Burgel, P.R.; Dupont, L.; Eickmeier, O.; Fayon, M.; Leal, T.; et al. Inflammation biomarkers in sputum for clinical trials in cystic fibrosis: Current understanding and gaps in knowledge. J. Cyst. Fibros. 2022, 21, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Giddings, O.; Esther, C.R., Jr. Mapping targetable inflammation and outcomes with cystic fibrosis biomarkers. Pediatr. Pulmonol. 2017, 52, S21–S28. [Google Scholar] [CrossRef] [PubMed]
- Paone, G.; Leone, V.; Conti, V.; De Marchis, L.; Ialleni, E.; Graziani, C.; Salducci, M.; Ramaccia, M.; Munafo, G. Blood and sputum biomarkers in COPD and asthma: A review. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 698–708. [Google Scholar] [PubMed]
- Moon, J.Y.; Leitao Filho, F.S.; Shahangian, K.; Takiguchi, H.; Sin, D.D. Blood and sputum protein biomarkers for chronic obstructive pulmonary disease (COPD). Expert. Rev. Proteomics 2018, 15, 923–935. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, M.; Iadarola, P.; Viglio, S. Proteomic Analysis of Human Sputum for the Diagnosis of Lung Disorders: Where Are We Today? Int. J. Mol. Sci. 2022, 23, 5692. [Google Scholar] [CrossRef]
- Dong, T.; Santos, S.; Yang, Z.; Yang, S.; Kirkhus, N.E. Sputum and salivary protein biomarkers and point-of-care biosensors for the management of COPD. Analyst 2020, 145, 1583–1604. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Liu, Y.; Yang, Q.; Han, S.; Zhang, H. Prognostic Biomarkers Based on Proteomic Technology in COPD: A Recent Review. Int. J. Chron. Obstruct Pulmon Dis. 2023, 18, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.F.; Ko, F.W.; Wong, G.W. Recent advances in asthma biomarker research. Ther. Adv. Respir. Dis. 2013, 7, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Diamant, Z.; Vijverberg, S.; Alving, K.; Bakirtas, A.; Bjermer, L.; Custovic, A.; Dahlen, S.E.; Gaga, M.; Gerth van Wijk, R.; Giacco, S.D.; et al. Toward clinically applicable biomarkers for asthma: An EAACI position paper. Allergy 2019, 74, 1835–1851. [Google Scholar] [CrossRef] [PubMed]
- Szczesniak, R.; Heltshe, S.L.; Stanojevic, S.; Mayer-Hamblett, N. Use of FEV(1) in cystic fibrosis epidemiologic studies and clinical trials: A statistical perspective for the clinical researcher. J. Cyst. Fibros. 2017, 16, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.L.; O’Callaghan, L.; Pelligra, C.; Konstan, M.W.; Ward, A.; Ishak, J.K.; Chandler, C.; Liou, T.G. Modeling long-term health outcomes of patients with cystic fibrosis homozygous for F508del-CFTR treated with lumacaftor/ivacaftor. Ther. Adv. Respir. Dis. 2019, 13, 1753466618820186. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Hamblett, N.; Aitken, M.L.; Accurso, F.J.; Kronmal, R.A.; Konstan, M.W.; Burns, J.L.; Sagel, S.D.; Ramsey, B.W. Association between pulmonary function and sputum biomarkers in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2007, 175, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Fayon, M.; Kent, L.; Bui, S.; Dupont, L.; Sermet, I.; European Cystic Fibrosis Society Clinical Trial Network (ECFS-CTN) Standardisation Committee. Clinimetric properties of bronchoalveolar lavage inflammatory markers in cystic fibrosis. Eur. Respir. J. 2014, 43, 610–626. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, K.A.; Schultz, A.; Stick, S.M. Biomarkers in Paediatric Cystic Fibrosis Lung Disease. Paediatr. Respir. Rev. 2015, 16, 213–218. [Google Scholar] [CrossRef]
- Ronchetti, K.; Tame, J.D.; Paisey, C.; Thia, L.P.; Doull, I.; Howe, R.; Mahenthiralingam, E.; Forton, J.T. The CF-Sputum Induction Trial (CF-SpIT) to assess lower airway bacterial sampling in young children with cystic fibrosis: A prospective internally controlled interventional trial. Lancet Respir. Med. 2018, 6, 461–471. [Google Scholar] [CrossRef]
- Henig, N.R.; Tonelli, M.R.; Pier, M.V.; Burns, J.L.; Aitken, M.L. Sputum induction as a research tool for sampling the airways of subjects with cystic fibrosis. Thorax 2001, 56, 306–311. [Google Scholar] [CrossRef]
- De Boeck, K.; Alifier, M.; Vandeputte, S. Sputum induction in young cystic fibrosis patients. Eur. Respir. J. 2000, 16, 91–94. [Google Scholar] [CrossRef]
- Hovold, G.; Palmcrantz, V.; Kahn, F.; Egesten, A.; Pahlman, L.I. Heparin-binding protein in sputum as a marker of pulmonary inflammation, lung function, and bacterial load in children with cystic fibrosis. BMC Pulm. Med. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, C.L.; Henig, N.R.; Mayer-Hamblett, N.; Accurso, F.J.; Burns, J.L.; Chmiel, J.F.; Daines, C.L.; Gibson, R.L.; McNamara, S.; Retsch-Bogart, Z.; et al. Inflammatory and microbiologic markers in induced sputum after intravenous antibiotics in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2003, 168, 171–1475. [Google Scholar] [CrossRef] [PubMed]
- Sagel, S.D.; Wagner, B.D.; Anthony, M.M.; Emmett, P.; Zemanick, E.T. Sputum biomarkers of inflammation and lung function decline in children with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2012, 186, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Oriano, M.; Terranova, L.; Sotgiu, G.; Saderi, L.; Bellofiore, A.; Retucci, M.; Marotta, C.; Gramegna, A.; Miglietta, D.; Carnini, C.; et al. Evaluation of active neutrophil elastase in sputum of bronchiectasis and cystic fibrosis patients: A comparison among different techniques. Pulm. Pharmacol. Ther. 2019, 59, 101856. [Google Scholar] [CrossRef]
- Forrest, O.A.; Chopyk, D.M.; Gernez, Y.; Brown, M.R.; Conrad, C.K.; Moss, R.B.; Tangpricha, V.; Peng, L.; Tirouvanziam, R. Resistin is elevated in cystic fibrosis sputum and correlates negatively with lung function. J. Cyst. Fibros. 2019, 18, 64–70. [Google Scholar] [CrossRef]
- Bokarewa, M.; Nagaev, I.; Dahlberg, L.; Smith, U.; Tarkowski, A. Resistin, an adipokine with potent proinflammatory properties. J. Immunol. 2005, 174, 5789–5795. [Google Scholar] [CrossRef]
- Reihill, J.; Moffitt, K.; Douglas, L.; Stuart Elborn, J.; Jones, A.; Lorraine Martin, S. Sputum trypsin-like protease activity relates to clinical outcome in cystic fibrosis. J. Cyst. Fibros. 2020, 19, 647–653. [Google Scholar] [CrossRef]
- Reihill, J.A.; Walker, B.; Hamilton, R.A.; Ferguson, T.E.; Elborn, J.S.; Stutts, M.J.; Harvey, B.J.; Saint-Criq, V.; Hendrick, S.M.; Martin, S.L. Inhibition of Protease-ENaC Signaling Improves Mucociliary Function in Cystic Fibrosis Airways. Am. J. Respir. Crit. Care Med. 2016, 194, 701–710. [Google Scholar] [CrossRef]
- McMahon, D.B.; Workman, A.D.; Kohanski, M.A.; Carey, R.M.; Freund, J.R.; Hariri, B.M.; Chen, B.; Doghramji, L.J.; Adappa, N.D.; Palmer, J.N.; et al. Protease-activated receptor 2 activates airway apical membrane chloride permeability and increases ciliary beating. FASEB J. 2018, 32, 155–167. [Google Scholar] [CrossRef]
- Jain, R.; Baines, A.; Khan, U.; Wagner, B.D.; Sagel, S.D. Evaluation of airway and circulating inflammatory biomarkers for cystic fibrosis drug development. J. Cyst. Fibros. 2021, 20, 50–56. [Google Scholar] [CrossRef]
- Tirelli, A.S.; Colombo, C.; Torresani, E.; Cariani, L.; Arnaboldi, E.; Conese, M. Validation of an automated sensitive immunoassay for quantitation of cytokines in the sputum of cystic fibrosis patients. Clin. Chem. Lab. Med. 2007, 45, 108–111. [Google Scholar] [CrossRef]
- Liou, T.G.; Argel, N.; Asfour, F.; Brown, P.S.; Chatfield, B.A.; Cox, D.R.; Daines, C.L.; Durham, D.; Francis, J.A.; Glover, B.; et al. Associations of Sputum Biomarkers with Clinical Outcomes in People with Cystic Fibrosis. medRxiv 2022. [Google Scholar] [CrossRef]
- Pattison, S.H.; Gibson, D.S.; Johnston, E.; Peacock, S.; Rivera, K.; Tunney, M.M.; Pappin, D.J.; Elborn, J.S. Proteomic profile of cystic fibrosis sputum cells in adults chronically infected with Pseudomonas aeruginosa. Eur. Respir. J. 2017, 50, 1601569. [Google Scholar] [CrossRef] [PubMed]
- Sagel, S.D.; Kapsner, R.K.; Osberg, I. Induced sputum matrix metalloproteinase-9 correlates with lung function and airway inflammation in children with cystic fibrosis. Pediatr. Pulmonol. 2005, 39, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Okamoto, K.; Rubin, B.K. Pulmonary function is negatively correlated with sputum inflammatory markers and cough clearability in subjects with cystic fibrosis but not those with chronic bronchitis. Chest 2006, 129, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Sloane, A.J.; Lindner, R.A.; Prasad, S.S.; Sebastian, L.T.; Pedersen, S.K.; Robinson, M.; Bye, P.T.; Nielson, D.W.; Harry, J.L. Proteomic analysis of sputum from adults and children with cystic fibrosis and from control subjects. Am. J. Respir. Crit. Care Med. 2005, 172, 1416–1426. [Google Scholar] [CrossRef]
- Hector, A.; Kormann, M.S.; Mack, I.; Latzin, P.; Casaulta, C.; Kieninger, E.; Zhou, Z.; Yildirim, A.O.; Bohla, A.; Rieber, N.; et al. The chitinase-like protein YKL-40 modulates cystic fibrosis lung disease. PLoS ONE 2011, 6, e24399. [Google Scholar] [CrossRef]
- Gray, R.D.; Imrie, M.; Boyd, A.C.; Porteous, D.; Innes, J.A.; Greening, A.P. Sputum and serum calprotectin are useful biomarkers during CF exacerbation. J. Cyst. Fibros. 2010, 9, 193–198. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Halbwachs-Mecarelli, L.; Schuster, A.; Nusbaum, P.; Ueki, I.; Canteloup, S.; Lenoir, G.; Descamps-Latscha, B.; Nadel, J.A. Proteinase 3, a potent secretagogue in airways, is present in cystic fibrosis sputum. Am. J. Respir. Cell Mol. Biol. 1999, 20, 729–736. [Google Scholar] [CrossRef]
- Gray, R.D.; MacGregor, G.; Noble, D.; Imrie, M.; Dewar, M.; Boyd, A.C.; Innes, J.A.; Porteous, D.J.; Greening, A.P. Sputum proteomics in inflammatory and suppurative respiratory diseases. Am. J. Respir. Crit. Care Med. 2008, 178, 444–452. [Google Scholar] [CrossRef]
- Maher, R.E.; Barrett, E.; Beynon, R.J.; Harman, V.M.; Jones, A.M.; McNamara, P.S.; Smith, J.A.; Lord, R.W. The relationship between lung disease severity and the sputum proteome in cystic fibrosis. Respir. Med. 2022, 204, 107002. [Google Scholar] [CrossRef]
- Hisert, K.B.; Heltshe, S.L.; Pope, C.; Jorth, P.; Wu, X.; Edwards, R.M.; Radey, M.; Accurso, F.J.; Wolter, D.J.; Cooke, G.; et al. Restoring Cystic Fibrosis Transmembrane Conductance Regulator Function Reduces Airway Bacteria and Inflammation in People with Cystic Fibrosis and Chronic Lung Infections. Am. J. Respir. Crit. Care Med. 2017, 195, 1617–1628. [Google Scholar] [CrossRef]
- Gianotti, A.; Capurro, V.; Scudieri, P.; Galietta, L.J.; Moran, O.; Zegarra-Moran, O. Pharmacological rescue of mutant CFTR protein improves the viscoelastic properties of CF mucus. J. Cyst. Fibros. 2016, 15, 295–301. [Google Scholar] [CrossRef]
- Morrison, C.B.; Shaffer, K.M.; Araba, K.C.; Markovetz, M.R.; Wykoff, J.A.; Quinney, N.L.; Hao, S.; Delion, M.F.; Flen, A.L.; Morton, L.C.; et al. Treatment of cystic fibrosis airway cells with CFTR modulators reverses aberrant mucus properties via hydration. Eur. Respir. J. 2022, 59, 2100185. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.K.; Wagner, B.D.; Zemanick, E.T.; Robertson, C.E.; Stevens, M.J.; Heltshe, S.L.; Rowe, S.M.; Sagel, S.D. Changes in Airway Microbiome and Inflammation with Ivacaftor Treatment in Patients with Cystic Fibrosis and the G551D Mutation. Ann. Am. Thorac. Soc. 2020, 17, 212–220. [Google Scholar] [CrossRef]
- Graeber, S.Y.; Boutin, S.; Wielputz, M.O.; Joachim, C.; Frey, D.L.; Wege, S.; Sommerburg, O.; Kauczor, H.U.; Stahl, M.; Dalpke, A.H.; et al. Effects of Lumacaftor-Ivacaftor on Lung Clearance Index, Magnetic Resonance Imaging, and Airway Microbiome in Phe508del Homozygous Patients with Cystic Fibrosis. Ann. Am. Thorac. Soc. 2021, 18, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Arooj, P.; Morrissy, D.V.; McCarthy, Y.; Vagg, T.; McCarthy, M.; Fleming, C.; Daly, M.; Eustace, J.A.; Murphy, D.M.; Plant, B.J. ROCK STUDY in CF: Sustained anti-inflammatory effects of lumacaftor-ivacaftor in sputum and peripheral blood samples of adult patients with cystic fibrosis-an observational study. BMJ Open Respir. Res. 2023, 10, e001590. [Google Scholar] [CrossRef] [PubMed]
- Schaupp, L.; Addante, A.; Voller, M.; Fentker, K.; Kuppe, A.; Bardua, M.; Duerr, J.; Piehler, L.; Rohmel, J.; Thee, S.; et al. Longitudinal effects of elexacaftor/tezacaftor/ivacaftor on sputum viscoelastic properties, airway infection and inflammation in patients with cystic fibrosis. Eur. Respir. J. 2023, 62, 2202153. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Haas, D.; Bouchard, M.; Quon, B.S. Metabolomic Biomarkers to Predict and Diagnose Cystic Fibrosis Pulmonary Exacerbations: A Systematic Review. Front. Pediatr. 2022, 10, 896439. [Google Scholar] [CrossRef]
- Twomey, K.B.; Alston, M.; An, S.Q.; O’Connell, O.J.; McCarthy, Y.; Swarbreck, D.; Febrer, M.; Dow, J.M.; Plant, B.J.; Ryan, R.P. Microbiota and metabolite profiling reveal specific alterations in bacterial community structure and environment in the cystic fibrosis airway during exacerbation. PLoS ONE 2013, 8, e82432. [Google Scholar] [CrossRef]
- Hanusch, B.; Brinkmann, F.; Mayorandan, S.; Chobanyan-Jurgens, K.; Wiemers, A.; Jansen, K.; Ballmann, M.; Schmidt-Choudhury, A.; Bollenbach, A.; Derichs, N.; et al. Local and Systemic Alterations of the L-Arginine/Nitric Oxide Pathway in Sputum, Blood, and Urine of Pediatric Cystic Fibrosis Patients and Effects of Antibiotic Treatment. J. Clin. Med. 2020, 9, 3802. [Google Scholar] [CrossRef] [PubMed]
- Gifford, A.H.; Polineni, D.; He, J.; D’Amico, J.L.; Dorman, D.B.; Williams, M.A.; Nymon, A.B.; Balwan, A.; Budden, T.; Zuckerman, J.B. A pilot study of cystic fibrosis exacerbation response phenotypes reveals contrasting serum and sputum iron trends. Sci. Rep. 2021, 11, 4897. [Google Scholar] [CrossRef]
- Reid, D.W.; Carroll, V.; O’May, C.; Champion, A.; Kirov, S.M. Increased airway iron as a potential factor in the persistence of Pseudomonas aeruginosa infection in cystic fibrosis. Eur. Respir. J. 2007, 30, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Ishchenko, O.; Koshevaya, I.; Zhernosekova, I.; Garets, V.; Stepanskyi, D. The Levels of the Human-β-Defensin-2 and LL-37 in the Sputum of Children with Cystic Fibrosis: A Case–control Study and Literature Review. Open Access Maced. J. Med. Sci. 2022, 10, 2367–2373. [Google Scholar] [CrossRef]
- Chen, C.I.; Schaller-Bals, S.; Paul, K.P.; Wahn, U.; Bals, R. Beta-defensins and LL-37 in bronchoalveolar lavage fluid of patients with cystic fibrosis. J. Cyst. Fibros. 2004, 3, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Vestbo, J.; Hurd, S.S.; Agusti, A.G.; Jones, P.W.; Vogelmeier, C.; Anzueto, A.; Barnes, P.J.; Fabbri, L.M.; Martinez, F.J.; Nishimura, M.; et al. Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease: GOLD executive summary. Am. J. Respir. Crit. Care Med. 2013, 187, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Agnew, M. Spirometry in clinical use: Practical issues. Breathe 2010, 6, 196–203. [Google Scholar]
- Naranjo-Hernandez, D.; Talaminos-Barroso, A.; Reina-Tosina, J.; Roa, L.M.; Barbarov-Rostan, G.; Cejudo-Ramos, P.; Marquez-Martin, E.; Ortega-Ruiz, F. Smart Vest for Respiratory Rate Monitoring of COPD Patients Based on Non-Contact Capacitive Sensing. Sensors 2018, 18, 2144. [Google Scholar] [CrossRef]
- Biomarkers Definitions Working, G. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar]
- Fleming, T.R.; Powers, J.H. Biomarkers and surrogate endpoints in clinical trials. Stat. Med. 2012, 31, 2973–2984. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, R.; Preiano, M.; Maggisano, G.; Pelaia, C.; Savino, R. Hexagonal Mesoporous Silica as a Rapid, Efficient and Versatile Tool for MALDI-TOF MS Sample Preparation in Clinical Peptidomics Analysis: A Pilot Study. Molecules 2019, 24, 2311. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Chakraborty, R.; Saha, B.; Suri, H.; Singh, P.; Raj, A.; Taneja, B.; Dash, D.; Sengupta, S.; Agrawal, A. Sputum Protein Biomarkers in Airway Diseases: A Pilot Study. Int. J. Chron. Obstruct Pulmon Dis. 2021, 16, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Barta, I.; Paska, C.; Antus, B. Sputum Cytokine Profiling in COPD: Comparison Between Stable Disease and Exacerbation. Int. J. Chron. Obstruct Pulmon Dis. 2022, 17, 1897–1908. [Google Scholar] [CrossRef]
- Barnes, P.J. The cytokine network in chronic obstructive pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2009, 41, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Yasuo, M.; Urushibata, K.; Hanaoka, M.; Koizumi, T.; Kubo, K. Airway inflammation during stable and acutely exacerbated chronic obstructive pulmonary disease. Eur. Respir. J. 2005, 25, 640–646. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; O’Neal, W.K.; Anderson, W.H.; Kesimer, M.; Ceppe, A.; Doerschuk, C.M.; Alexis, N.E.; Hastie, A.T.; Barr, R.G.; Bowler, R.P.; et al. Identification of Sputum Biomarkers Predictive of Pulmonary Exacerbations in COPD. Chest 2022, 161, 1239–1249. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; Turkovic, L.; Rosenow, T.; Muhlebach, M.S.; Boucher, R.C.; Ranganathan, S.; Stick, S.M.; Arest, C.F. Metabolomic biomarkers predictive of early structural lung disease in cystic fibrosis. Eur. Respir. J. 2016, 48, 1612–1621. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; Coakley, R.D.; Henderson, A.G.; Zhou, Y.H.; Wright, F.A.; Boucher, R.C. Metabolomic Evaluation of Neutrophilic Airway Inflammation in Cystic Fibrosis. Chest 2015, 148, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, G.J.; Lamb, N.J.; Tilley, R.; Evans, T.W.; Gutteridge, J.M. Plasma hypoxanthine levels in ARDS: Implications for oxidative stress, morbidity, and mortality. Am. J. Respir. Crit. Care Med. 1997, 155, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal 2012, 8, 359–373. [Google Scholar] [CrossRef]
- Lazarowski, E.R.; Sesma, J.I.; Seminario, L.; Esther, C.R., Jr.; Kreda, S.M. Nucleotide release by airway epithelia. Subcell. Biochem. 2011, 55, 1–15. [Google Scholar]
- Kreda, S.M.; Okada, S.F.; van Heusden, C.A.; O’Neal, W.; Gabriel, S.; Abdullah, L.; Davis, C.W.; Boucher, R.C.; Lazarowski, E.R. Coordinated release of nucleotides and mucin from human airway epithelial Calu-3 cells. J. Physiol. 2007, 584, 245–259. [Google Scholar] [CrossRef]
- Mulvanny, A.; Pattwell, C.; Beech, A.; Southworth, T.; Singh, D. Validation of Sputum Biomarker Immunoassays and Cytokine Expression Profiles in COPD. Biomedicines 2022, 10, 1949. [Google Scholar] [CrossRef] [PubMed]
- Eickmeier, O.; Huebner, M.; Herrmann, E.; Zissler, U.; Rosewich, M.; Baer, P.C.; Buhl, R.; Schmitt-Grohe, S.; Zielen, S.; Schubert, R. Sputum biomarker profiles in cystic fibrosis (CF) and chronic obstructive pulmonary disease (COPD) and association between pulmonary function. Cytokine 2010, 50, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Guiedem, E.; Pefura-Yone, E.W.; Ikomey, G.M.; Nkenfou, C.N.; Mesembe, M.; Yivala, M.M.; Chendi, B.H.; Jacobs, G.B.; Chegou, N.N.; Okomo, M.C.A. Cytokine profile in the sputum of subjects with post-tuberculosis airflow obstruction and in those with tobacco related chronic obstructive pulmonary disease. BMC Immunol. 2020, 21, 52. [Google Scholar] [CrossRef] [PubMed]
- Aaron, S.D.; Vandemheen, K.L.; Ramsay, T.; Zhang, C.; Avnur, Z.; Nikolcheva, T.; Quinn, A. Multi analyte profiling and variability of inflammatory markers in blood and induced sputum in patients with stable COPD. Respir. Res. 2010, 11, 41. [Google Scholar] [CrossRef]
- Moermans, C.; Bonnet, C.; Willems, E.; Baron, F.; Nguyen, M.; Henket, M.; Sele, J.; Corhay, J.L.; Beguin, Y.; Louis, R. Sputum cytokine levels in patients undergoing hematopoietic SCT and comparison with healthy subjects and COPD: A pilot study. Bone Marrow Transplant. 2014, 49, 1382–1388. [Google Scholar] [CrossRef]
- Aaron, S.D.; Angel, J.B.; Lunau, M.; Wright, K.; Fex, C.; Le Saux, N.; Dales, R.E. Granulocyte inflammatory markers and airway infection during acute exacerbation of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2001, 163, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Bafadhel, M.; McKenna, S.; Terry, S.; Mistry, V.; Reid, C.; Haldar, P.; McCormick, M.; Haldar, K.; Kebadze, T.; Duvoix, A.; et al. Acute exacerbations of chronic obstructive pulmonary disease: Identification of biologic clusters and their biomarkers. Am. J. Respir. Crit. Care Med. 2011, 184, 662–671. [Google Scholar] [CrossRef]
- Cane, J.; Tregidgo, L.; Thulborn, S.; Finch, D.; Bafadhel, M. Antimicrobial Peptides SLPI and Beta Defensin-1 in Sputum are Negatively Correlated with FEV(1). Int. J. Chron. Obstruct Pulmon Dis. 2021, 16, 1437–1447. [Google Scholar] [CrossRef]
- Hastie, A.T.; Moore, W.C.; Meyers, D.A.; Vestal, P.L.; Li, H.; Peters, S.P.; Bleecker, E.R.; National Heart, L.; Blood Institute Severe Asthma Research, P. Analyses of asthma severity phenotypes and inflammatory proteins in subjects stratified by sputum granulocytes. J. Allergy Clin. Immunol. 2010, 125, 1028–1036. [Google Scholar] [CrossRef]
- Gharib, S.A.; Nguyen, E.V.; Lai, Y.; Plampin, J.D.; Goodlett, D.R.; Hallstrand, T.S. Induced sputum proteome in healthy subjects and asthmatic patients. J. Allergy Clin. Immunol. 2011, 128, 1176–1184.e1176. [Google Scholar] [CrossRef]
- Lefaudeux, D.; De Meulder, B.; Loza, M.J.; Peffer, N.; Rowe, A.; Baribaud, F.; Bansal, A.T.; Lutter, R.; Sousa, A.R.; Corfield, J.; et al. U-BIOPRED clinical adult asthma clusters linked to a subset of sputum omics. J. Allergy Clin. Immunol. 2017, 139, 1797–1807. [Google Scholar] [CrossRef]
- Schofield, J.P.R.; Burg, D.; Nicholas, B.; Strazzeri, F.; Brandsma, J.; Staykova, D.; Folisi, C.; Bansal, A.T.; Xian, Y.; Guo, Y.; et al. Stratification of asthma phenotypes by airway proteomic signatures. J. Allergy Clin. Immunol. 2019, 144, 70–82. [Google Scholar] [CrossRef]
- Gibson, P.G.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; Peters, M.J.; Marks, G.B.; Baraket, M.; et al. Effect of azithromycin on asthma exacerbations and quality of life in adults with persistent uncontrolled asthma (AMAZES): A randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 659–668. [Google Scholar] [CrossRef]
- Niessen, N.M.; Gibson, P.G.; Baines, K.J.; Barker, D.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; et al. Sputum TNF markers are increased in neutrophilic and severe asthma and are reduced by azithromycin treatment. Allergy 2021, 76, 2090–2101. [Google Scholar] [CrossRef]
- Baines, K.J.; Simpson, J.L.; Wood, L.G.; Scott, R.J.; Gibson, P.G. Transcriptional phenotypes of asthma defined by gene expression profiling of induced sputum samples. J. Allergy Clin. Immunol. 2011, 127, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.S.; Pavlidis, S.; Loza, M.; Baribaud, F.; Rowe, A.; Pandis, I.; Sousa, A.; Corfield, J.; Djukanovic, R.; Lutter, R.; et al. T-helper cell type 2 (Th2) and non-Th2 molecular phenotypes of asthma using sputum transcriptomics in U-BIOPRED. Eur. Respir. J. 2017, 49, 1602135. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Suzuki, Y.; Kikuchi, M.; Rikimaru, M.; Saito, J.; Shibata, Y. Sputum Neurturin Levels in Adult Asthmatic Subjects. J. Asthma Allergy 2023, 16, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Michel, T.; Theresine, M.; Poli, A.; Domingues, O.; Ammerlaan, W.; Brons, N.H.; Hentges, F.; Zimmer, J. Increased Th2 cytokine secretion, eosinophilic airway inflammation, and airway hyperresponsiveness in neurturin-deficient mice. J. Immunol. 2011, 186, 6497–6504. [Google Scholar] [CrossRef]
- Mauffray, M.; Domingues, O.; Hentges, F.; Zimmer, J.; Hanau, D.; Michel, T. Neurturin influences inflammatory responses and airway remodeling in different mouse asthma models. J. Immunol. 2015, 194, 1423–1433. [Google Scholar] [CrossRef]
- Chung, Y.; Katial, R.; Mu, F.; Cook, E.E.; Young, J.; Yang, D.; Betts, K.A.; Carstens, D.D. Real-world effectiveness of benralizumab: Results from the ZEPHYR 1 Study. Ann. Allergy Asthma Immunol. 2022, 128, 669–676.e666. [Google Scholar] [CrossRef]
- Schleich, F.; Graff, S.; Nekoee, H.; Moermans, C.; Henket, M.; Sanchez, C.; Paulus, V.; Guissard, F.; Donneau, A.F.; Louis, R. Real-word experience with mepolizumab: Does it deliver what it has promised? Clin. Exp. Allergy 2020, 50, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Moermans, C.; Brion, C.; Bock, G.; Graff, S.; Gerday, S.; Nekoee, H.; Poulet, C.; Bricmont, N.; Henket, M.; Paulus, V.; et al. Sputum Type 2 Markers Could Predict Remission in Severe Asthma Treated With Anti-IL-5. Chest 2023, 163, 1368–1379. [Google Scholar] [CrossRef]
- Fahy, J.V. Type 2 inflammation in asthma—Present in most, absent in many. Nat. Rev. Immunol. 2015, 15, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Chu, J.H.; Cohen, A.J.; Kaur, R.; Lee, S.; Wilson, G.; Liu, Q.; Gomez, J.; Rajaveen, H.; Yan, X.; et al. Sputum alarmins delineate distinct T2 cytokine pathways and unique subtypes of patients with asthma. Allergy 2023, 78, 3274–3277. [Google Scholar] [CrossRef] [PubMed]
- Ghebre, M.A.; Pang, P.H.; Desai, D.; Hargadon, B.; Newby, C.; Woods, J.; Rapley, L.; Cohen, S.E.; Herath, A.; Gaillard, E.A.; et al. Severe exacerbations in moderate-to-severe asthmatics are associated with increased pro-inflammatory and type 1 mediators in sputum and serum. BMC Pulm. Med. 2019, 19, 144. [Google Scholar] [CrossRef]
- Ramphul, M.; Welsh, K.G.; May, R.D.; Ghebre, M.A.; Rapley, L.; Cohen, E.S.; Herath, A.; Monteiro, W.; Rousseau, K.; Thornton, D.J.; et al. Sputum biomarkers during acute severe asthma attacks in children-a case-control study. Acta Paediatr. 2022, 111, 620–627. [Google Scholar] [CrossRef]
- Kaczynska, K.; Zajac, D.; Wojciechowski, P.; Jampolska, M. Regulatory Peptides in Asthma. Int. J. Mol. Sci. 2021, 22, 13656. [Google Scholar] [CrossRef]





| Pathological Changes/Disease | COPD | Asthma | CF |
|---|---|---|---|
| Mucus amount | Increased luminal content | Increased luminal content | Increased luminal content |
| Mucins | - Increased amounts of MUC5AC and MUC5B - Small amount of MUC2 | - Increased content of MUC5AC and variable content of MUC5B - Small amount of MUC2 | Spontaneous or induced sputum: - Decreased amounts of MUC5AC and MUC5B - Small amounts of MUC2 - Increased levels of MUC5AC and MUC5B BAL: - Increased amounts of MUC5B and MUC5AC |
| Other components of mucus | Albumin | DNA, actin | DNA, actin |
| “Tethering” of mucus to goblet cells | No | Yes | No |
| Mucus viscosity | High | High | Low |
| Airway pathology | - Goblet cell hyperplasia - Submucosal gland hypertrophy | - Loss of ciliated cells - Marked goblet cell hyperplasia - Submucosal gland hypertrophy | - Goblet cell hyperplasia - Submucosal gland hypertrophy |
| Plasma exudation | No | Yes | No |
| Airway inflammation | Macrophages and neutrophils | - Eosinophils and Th2 cells - Neutrophils in severe disease | Macrophages and neutrophils |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrami, M.; Biasin, A.; Tescione, F.; Tierno, D.; Dapas, B.; Carbone, A.; Grassi, G.; Conese, M.; Di Gioia, S.; Larobina, D.; et al. Mucus Structure, Viscoelastic Properties, and Composition in Chronic Respiratory Diseases. Int. J. Mol. Sci. 2024, 25, 1933. https://doi.org/10.3390/ijms25031933
Abrami M, Biasin A, Tescione F, Tierno D, Dapas B, Carbone A, Grassi G, Conese M, Di Gioia S, Larobina D, et al. Mucus Structure, Viscoelastic Properties, and Composition in Chronic Respiratory Diseases. International Journal of Molecular Sciences. 2024; 25(3):1933. https://doi.org/10.3390/ijms25031933
Chicago/Turabian StyleAbrami, Michela, Alice Biasin, Fabiana Tescione, Domenico Tierno, Barbara Dapas, Annalucia Carbone, Gabriele Grassi, Massimo Conese, Sante Di Gioia, Domenico Larobina, and et al. 2024. "Mucus Structure, Viscoelastic Properties, and Composition in Chronic Respiratory Diseases" International Journal of Molecular Sciences 25, no. 3: 1933. https://doi.org/10.3390/ijms25031933
APA StyleAbrami, M., Biasin, A., Tescione, F., Tierno, D., Dapas, B., Carbone, A., Grassi, G., Conese, M., Di Gioia, S., Larobina, D., & Grassi, M. (2024). Mucus Structure, Viscoelastic Properties, and Composition in Chronic Respiratory Diseases. International Journal of Molecular Sciences, 25(3), 1933. https://doi.org/10.3390/ijms25031933