
ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice


Abstract
:1. Introduction
2. Results
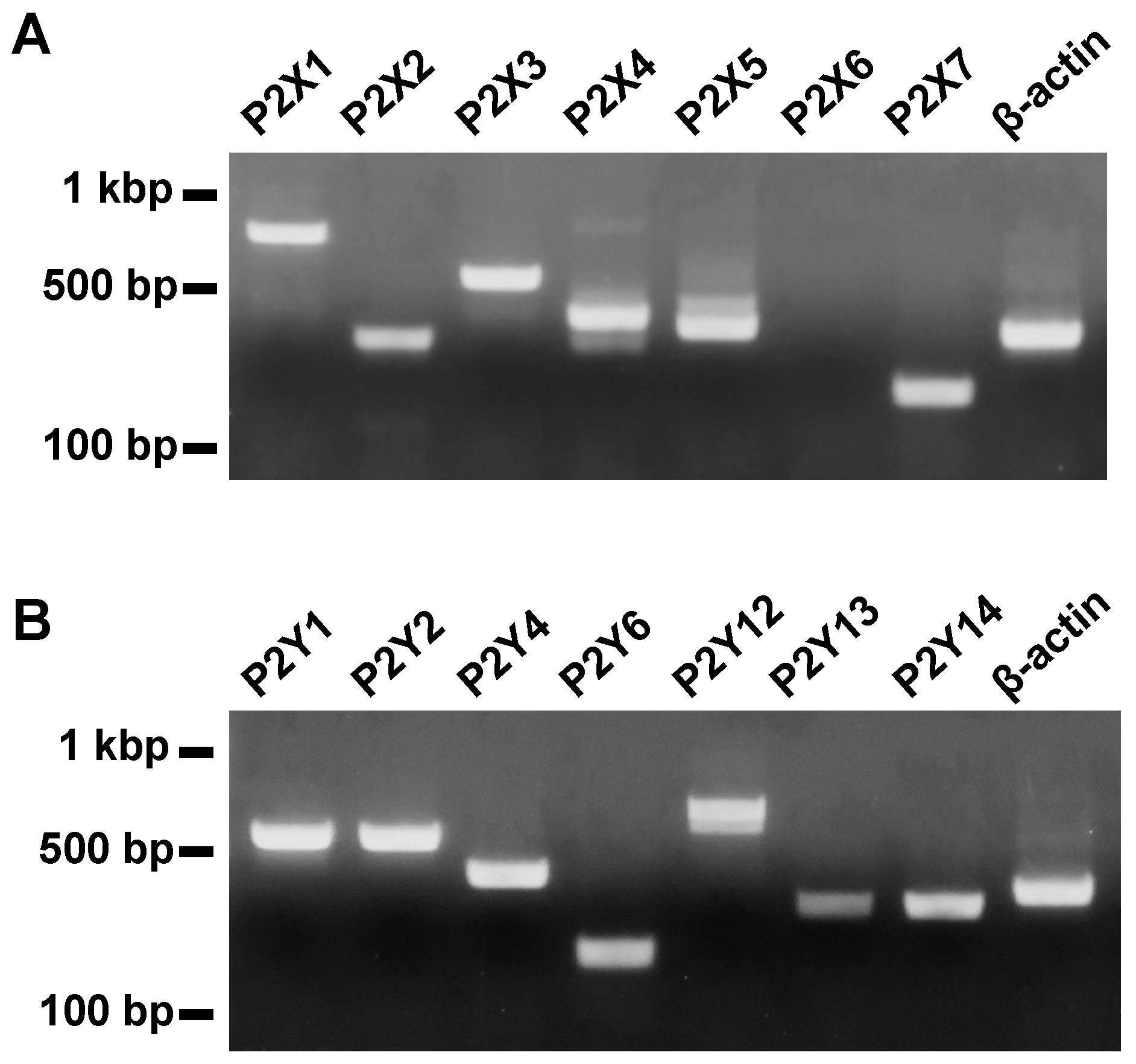
2.1. Molecular Identification of P2 Receptors in the Mouse Esophagus
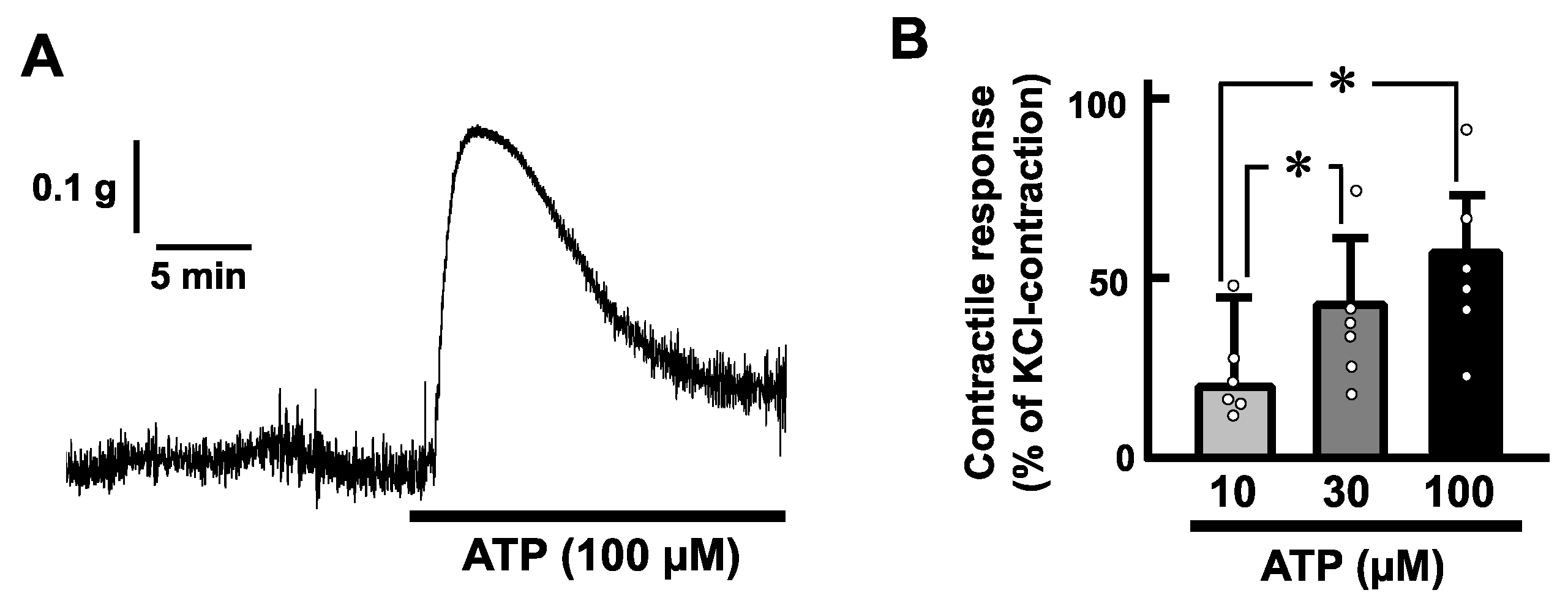
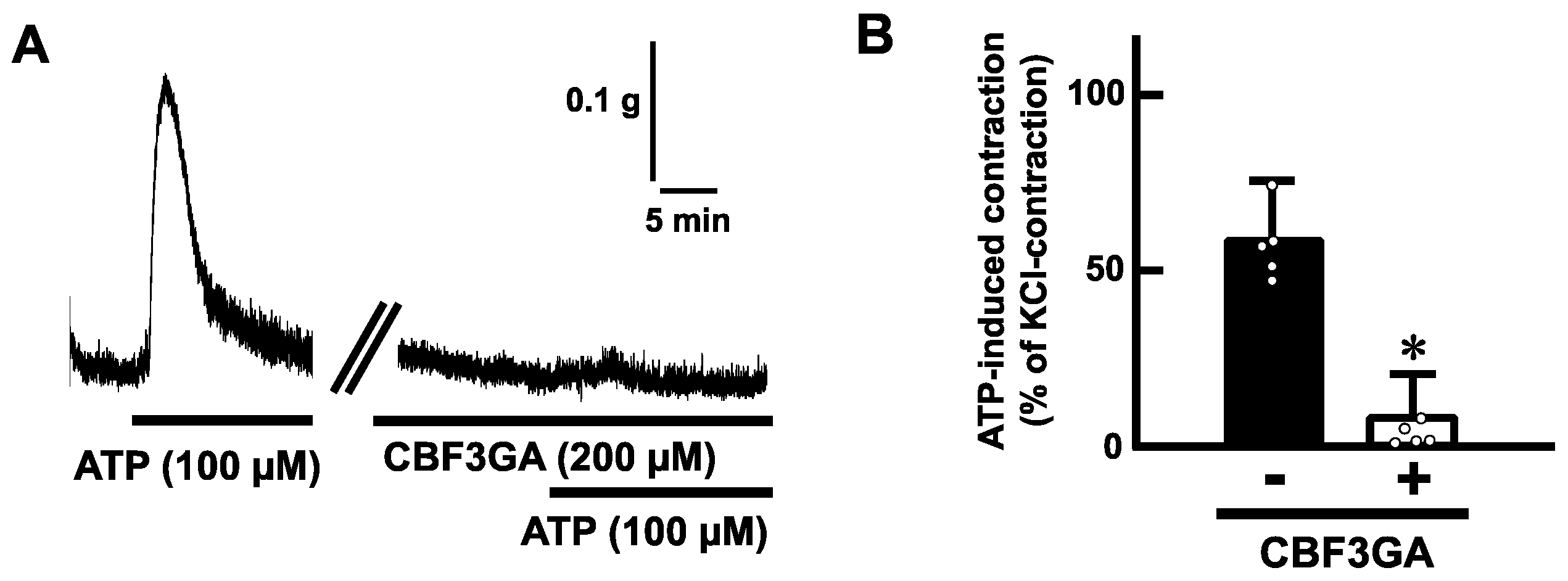
2.2. Effects of ATP on the Mechanical Activity of Mouse Esophageal Segments
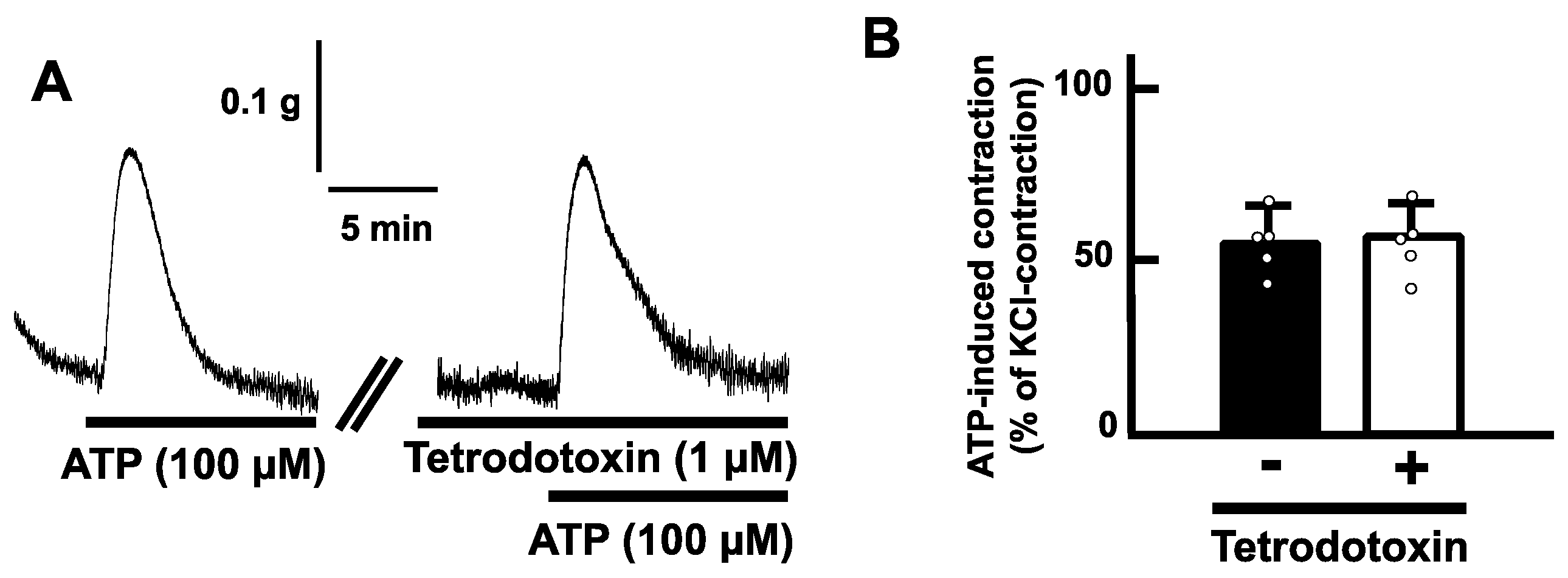
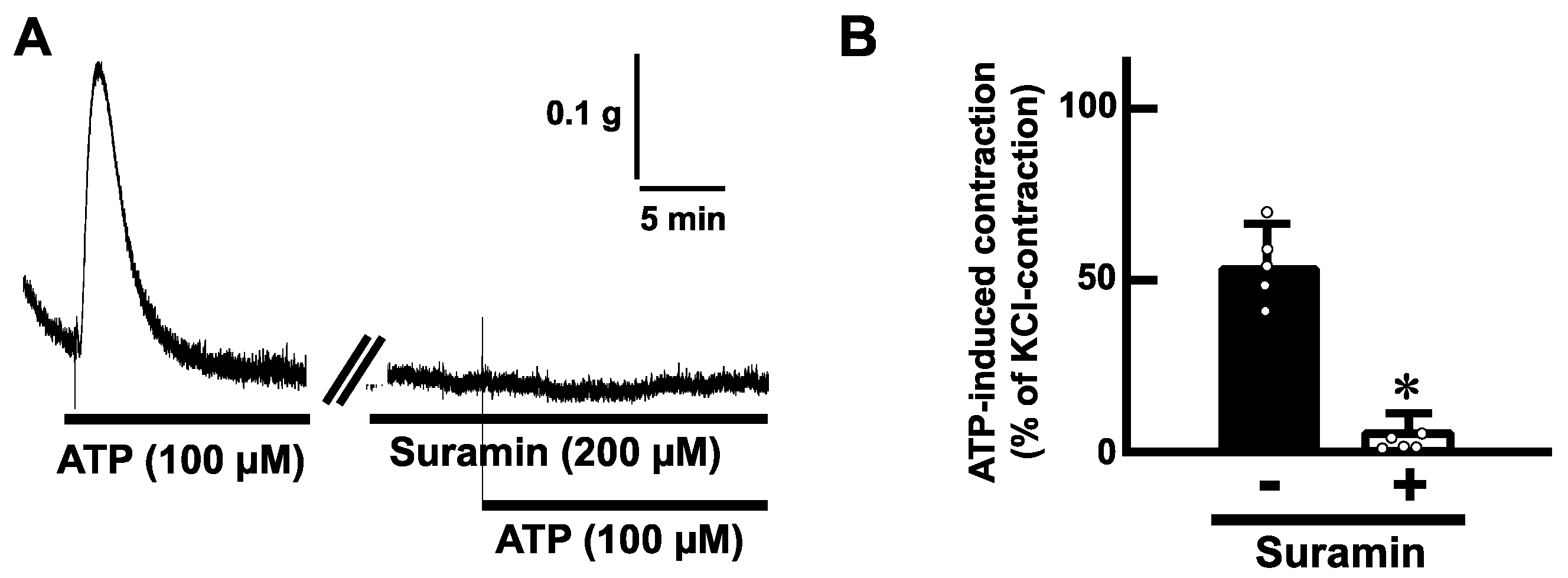
2.3. Effects of Purinoceptor Antagonists on ATP-Evoked Contractions in the Mouse Esophagus
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Esophageal Tissue Preparations
4.3. Recording of Mechanical Activity in Esophageal Segments
4.4. Solutions and Drugs
4.5. RNA Isolation and RT-PCR
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burnstock, G.; Ralevic, V. Purinergic signaling and blood vessels in health and disease. Pharmacol. Rev. 2013, 66, 102–192. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signalling in the gastrointestinal tract and related organs in health and disease. Purinergic Signal. 2014, 10, 3–50. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The Enteric Nervous System; Blackwell Publishing: Malden, MA, USA, 2006; pp. 103–131. [Google Scholar]
- Hansen, M.B. Neurohumoral control of gastrointestinal motility. Physiol. Res. 2003, 52, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M. G protein-coupled receptors in gastrointestinal physiology. IV. Neural regulation of gastrointestinal smooth muscle. Am. J. Physiol. 1998, 275, G1–G7. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Development and perspectives of the purinoceptor concept. J. Auton. Pharmacol. 1996, 16, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. The past, present and future of purine nucleotides as signalling molecules. Neuropharmacology 1997, 36, 1127–1139. [Google Scholar] [CrossRef]
- Jiménez, M.; Clavé, P.; Accarino, A.; Gallego, D. Purinergic neuromuscular transmission in the gastrointestinal tract; functional basis for future clinical and pharmacological studies. Br. J. Pharmacol. 2014, 171, 4360–4375. [Google Scholar] [CrossRef]
- Dreisig, K.; Kornum, B.R. A critical look at the function of the P2Y11 receptor. Purinergic Signal. 2016, 12, 427–437. [Google Scholar] [CrossRef]
- Giaroni, C.; Knight, G.E.; Zanetti, E.; Chiaravalli, A.M.; Lecchini, S.; Frigo, G.; Burnstock, G. Postnatal development of P2 receptors in the murine gastrointestinal tract. Neuropharmacology 2006, 50, 690–704. [Google Scholar] [CrossRef]
- Goyal, R.K.; Sullivan, M.P.; Chaudhury, A. Progress in understanding of inhibitory purinergic neuromuscular transmission in the gut. Neurogastroenterol. Motil. 2013, 25, 203–207. [Google Scholar] [CrossRef]
- Kestler, C.; Neuhuber, W.L.; Raab, M. Distribution of P2X3 receptor immunoreactivity in myenteric ganglia of the mouse esophagus. Histochem. Cell Biol. 2009, 131, 13–27. [Google Scholar] [CrossRef]
- Wang, Z.-J.; Neuhuber, W.L. Intraganglionic laminar endings in the rat esophagus contain purinergic P2X2 and P2X3 receptor immunoreactivity. Anat. Embryol. 2003, 207, 363–371. [Google Scholar] [CrossRef]
- Yuan, W.; Wang, Z.; Li, J.; Li, D.; Liu, D.; Bai, G.; Walsh, M.P.; Gui, Y.; Zheng, X.-L. Uridine adenosine tetraphosphate induces contraction of circular and longitudinal gastric smooth muscle by distinct signaling pathways. IUBMB Life 2013, 65, 623–632. [Google Scholar] [CrossRef]
- Zizzo, M.G.; Mulè, F.; Serio, R. Evidence that ATP or a related purine is an excitatory neurotransmitter in the longitudinal muscle of mouse distal colon. Br. J. Pharmacol. 2007, 151, 152–160. [Google Scholar] [CrossRef]
- Neuhuber, W.L.; Wörl, J. Enteric co-innervation of striated muscle in the esophagus: Still enigmatic? Histochem. Cell Biol. 2016, 146, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Wörl, J.; Neuhuber, W.L. Enteric co-innervation of motor endplates in the esophagus: State of the art ten years after. Histochem. Cell Biol. 2005, 123, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Bieger, D.; Neuhuber, W.L. Neural circuits and mediators regulating swallowing in the brainstem. GI Motil. Online 2006. [Google Scholar]
- Clouse, R.E.; Diamant, N.E. Motor Function of the Esophagus. In Physiology of the Gastrointestinal Tract, 4th ed.; Johnson, L.R., Ed.; Elsevier Academic Press: Burlington, MA, USA, 2006; pp. 913–926. [Google Scholar]
- Boudaka, A.; Wörl, J.; Shiina, T.; Shimizu, Y.; Takewaki, T.; Neuhuber, W.L. Galanin modulates vagally induced contractions in the mouse esophagus. Neurogastroenterol. Motil. 2009, 21, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Izumi, N.; Matsuyama, H.; Ko, M.; Shimizu, Y.; Takewaki, T. Role of intrinsic nitrergic neurones on vagally mediated striated muscle contractions in the hamster oesophagus. J. Physiol. 2003, 551, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Shiina, T.; Shimizu, Y.; Boudaka, A.; Wörl, J.; Takewaki, T. Tachykinins are involved in local reflex modulation of vagally mediated striated muscle contractions in the rat esophagus via tachykinin NK1 receptors. Neuroscience 2006, 139, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Shiina, T.; Shima, T.; Hirayama, H.; Kuramoto, H.; Takewaki, T.; Shimizu, Y. Contractile responses induced by physalaemin, an analogue of substance P, in the rat esophagus. Eur. J. Pharmacol. 2010, 628, 202–206. [Google Scholar] [CrossRef]
- Shiina, T.; Naitou, K.; Nakamori, H.; Suzuki, Y.; Horii, K.; Sano, Y.; Shimaoka, H.; Shimizu, Y. Serotonin-induced contractile responses of esophageal smooth muscle in the house musk shrew (Suncus murinus). Neurogastroenterol. Motil. 2016, 28, 1641–1648. [Google Scholar] [CrossRef]
- Bieger, D.; Triggle, C. Pharmacological properties of mechanical responses of the rat oesophageal muscularis mucosae to vagal and field stimulation. Br. J. Pharmacol. 1985, 84, 93–106. [Google Scholar] [PubMed]
- Christensen, J.; Percy, W.H. A pharmacological study of oesophageal muscularis mucosae from the cat, dog and American opossum (Didelphis virginiana). Br. J. Pharmacol. 1984, 83, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Percy, W.H.; Miller, A.J.; Brunz, J.T. Pharmacologic characteristics of rabbit esophageal muscularis mucosae in vitro. Dig. Dis. Sci. 1997, 42, 2537–2546. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.H.; Jung, H.; Cho, E.J.; Jeong, J.H.; Sohn, U.D. The Signaling Mechanism of Contraction Induced by ATP and UTP in Feline Esophageal Smooth Muscle Cells. Mol. Cells 2015, 38, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Storr, M.; Geisler, F.; Neuhuber, W.L.; Schusdziarra, V.; Allescher, H.D. Endomorphin-1 and -2, endogenous ligands for the μ-opioid receptor, inhibit striated and smooth muscle contraction in the rat oesophagus. Neurogastroenterol. Motil. 2000, 12, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Storr, M.; Geisler, F.; Neuhuber, W.L.; Schusdziarra, V.; Allescher, H.D. Characterization of vagal input to the rat esophageal muscle. Auton. Neurosci. Basic Clin. 2001, 91, 1–9. [Google Scholar] [CrossRef]
- Boudaka, A.; Tominaga, M. Physiological and Pathological Significance of Esophageal TRP Channels: Special Focus on TRPV4 in Esophageal Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 4550. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H.; Boudaka, A.; Sugiyama, T.; Moriyama, Y.; Tominaga, M. Transient receptor potential vanilloid 4 (TRPV4)-dependent calcium influx and ATP release in mouse oesophageal keratinocytes. J. Physiol. 2011, 589, 3471–3482. [Google Scholar] [CrossRef]
- Riquelme, M.A.; Cea, L.A.; Vega, J.L.; Boric, M.P.; Monyer, H.; Bennett, M.V.; Frank, M.; Willecke, K.; Sáez, J.C. The ATP required for potentiation of skeletal muscle contraction is released via pannexin hemichannels. Neuropharmacology 2013, 75, 594–603. [Google Scholar] [CrossRef]
- Furness, J.B. The enteric nervous system: Normal functions and enteric neuropathies. Neurogastroenterol. Motil. 2008, 20 (Suppl. S1), 32–38. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.K. Longitudinal muscle of the esophagus: Its role in esophageal health and disease. Curr. Opin. Gastroenterol. 2013, 29, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Horii, K.; Shiina, T.; Naitou, K.; Nakamori, H.; Horii, Y.; Shimaoka, H.; Shimizu, Y. Characterization of peristaltic motility in the striated muscle portion of the esophagus using a novel in vivo method in rats. Neurogastroenterol. Motil. 2019, 31, e13518. [Google Scholar] [CrossRef] [PubMed]
- Uneyama, H.; Uneyama, C.; Ebihara, S.; Akaike, N. Suramin and reactive blue 2 are antagonists for a newly identified purinoceptor on rat megakaryocyte. Br. J. Pharmacol. 1994, 111, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Fallon, M.; Bobbin, R.P. ATP antagonists cibacron blue, basilen blue and suramin alter sound-evoked responses of the cochlea and auditory nerve. Hear. Res. 1994, 78, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Hayato, R.; Ohtubo, Y.; Yoshii, K. Functional expression of ionotropic purinergic receptors on mouse taste bud cells. J. Physiol. 2007, 584, 473–488. [Google Scholar] [CrossRef]
- Hao, M.M.; Bergner, A.J.; Hirst, C.S.; Stamp, L.A.; Casagranda, F.; Bornstein, J.C.; Boesmans, W.; Vanden Berghe, P.; Young, H.M. Spontaneous calcium waves in the developing enteric nervous system. Dev. Biol. 2017, 428, 74–87. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Predicted Size (bp) |
|---|---|---|
| P2X1 | Forward CATTGTGCAGAGAACCCAGAA Reverse ATGTCCTCCGCATACTTGAAC | 776 |
| P2X2 | Forward ACGTTCATGAACAAAAACAAG Reverse TCAAAGTTGGGCCAAACCTTTGG | 360 |
| P2X3 | Forward AAGAGTGGGCAGTTACAAGGG Reverse GAAAACCCACCCCACAAAGT | 576 |
| P2X4 | Forward GAGAATGACGCTGGTGTGCC Reverse TTGGTGAGTGTGCGTTGCTC | 437 |
| P2X5 | Forward TAGTTAATGGCAAGGCGGGA Reverse AGCTCTGGCTACGTCTTCAC | 409 |
| P2X6 | Forward TACGTACTAACAGACGCA Reverse ATATCAGGGTTCTTTGGG | 254 |
| P2X7 | Forward TGTTTCCTTTGGCTGCTCCT Reverse CGCTCACCAAAGCAAAGCTAAT | 239 |
| P2Y1 | Forward TGGCGTGGTGTACCCTCTCAAGTC Reverse CGGGACAGTCTCCTTCTGAATGTA | 558 |
| P2Y2 | Forward CTCACGCGCACCCTCTACTA Reverse TCGGGTGCACTGCCTTTCTT | 549 |
| P2Y4 | Forward CTTTGGCTTTCCCTTCTTGA Reverse GTCCGCCCACCTGCTGAT | 427 |
| P2Y6 | Forward GCCCTGTGCTGGAGACCTTC Reverse CATGGCCCCAGTGACAAACA | 226 |
| P2Y12 | Forward CAGTGCAAGGGGTGGCATCT Reverse TGGCACACCAAGGTTCTCAG | 618 |
| P2Y13 | Forward GAAGAGAGGCACATGCAACA Reverse TTACTAATGCCAGGCCAACC | 345 |
| P2Y14 | Forward CAGTGCATGGAGCTCAAAAA Reverse GCAGCCGAGAGTAGCAGAGT | 347 |
| β-actin | Forward TGACCCTGAAGTACCCCATTG Reverse TCAGGATCTTCATGAGGTAG | 387 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, Y.; Shimizu, Y.; Shiina, T. ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice. Int. J. Mol. Sci. 2024, 25, 1985. https://doi.org/10.3390/ijms25041985
Suzuki Y, Shimizu Y, Shiina T. ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice. International Journal of Molecular Sciences. 2024; 25(4):1985. https://doi.org/10.3390/ijms25041985
Chicago/Turabian StyleSuzuki, Yuji, Yasutake Shimizu, and Takahiko Shiina. 2024. "ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice" International Journal of Molecular Sciences 25, no. 4: 1985. https://doi.org/10.3390/ijms25041985
APA StyleSuzuki, Y., Shimizu, Y., & Shiina, T. (2024). ATP-Induced Contractile Response of Esophageal Smooth Muscle in Mice. International Journal of Molecular Sciences, 25(4), 1985. https://doi.org/10.3390/ijms25041985