
Overexpression of Human sFLT1 in the Spongiotrophoblast Is Sufficient to Induce Placental Dysfunction and Fetal Growth Restriction in Transgenic Mice
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
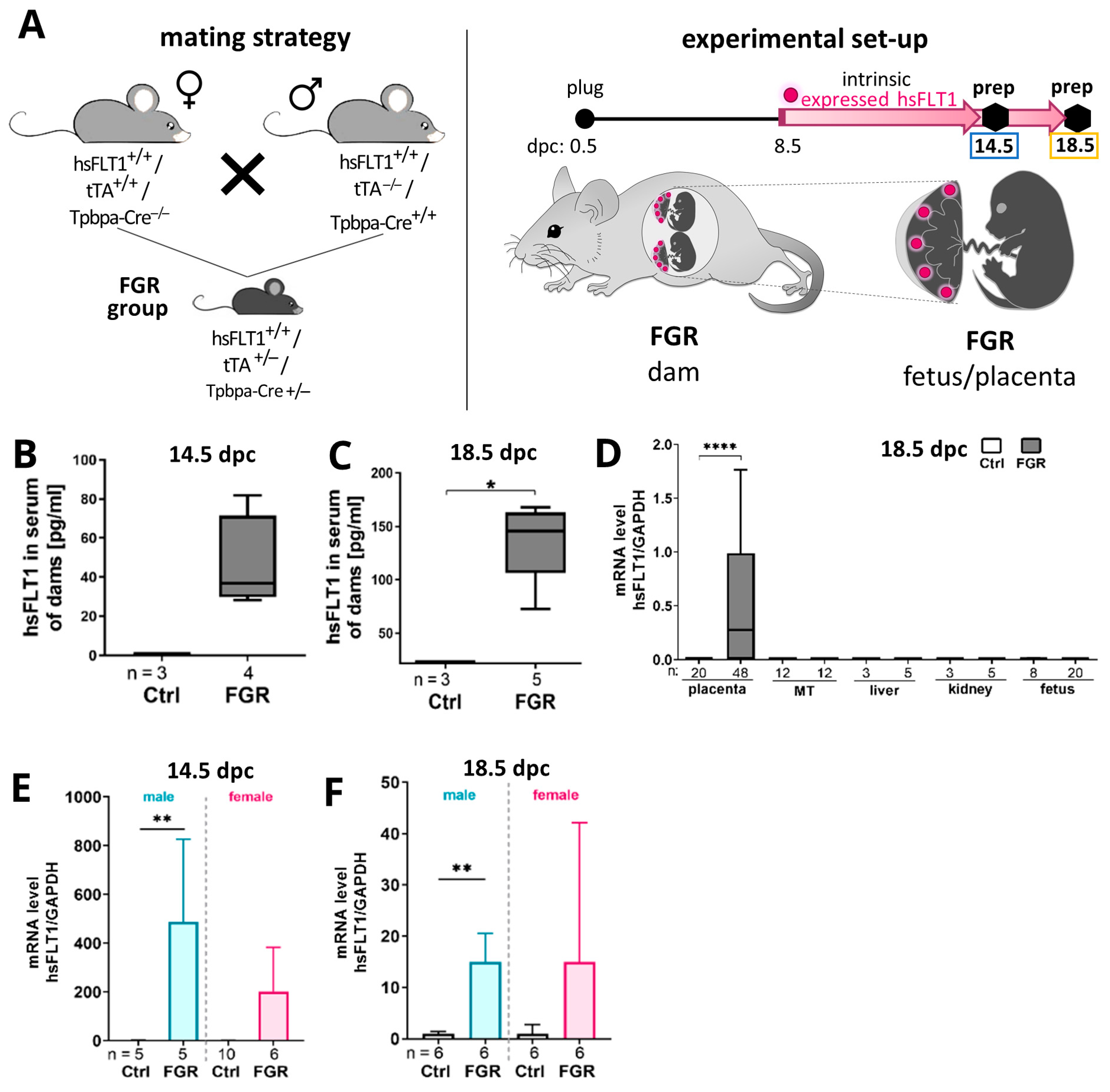
2.1. Human Soluble Fms-like Tyrosine Kinase Receptor-1 (hsFLT1) Is Exclusively Expressed in Spongiotrophoblast Cells in Transgenic Mice
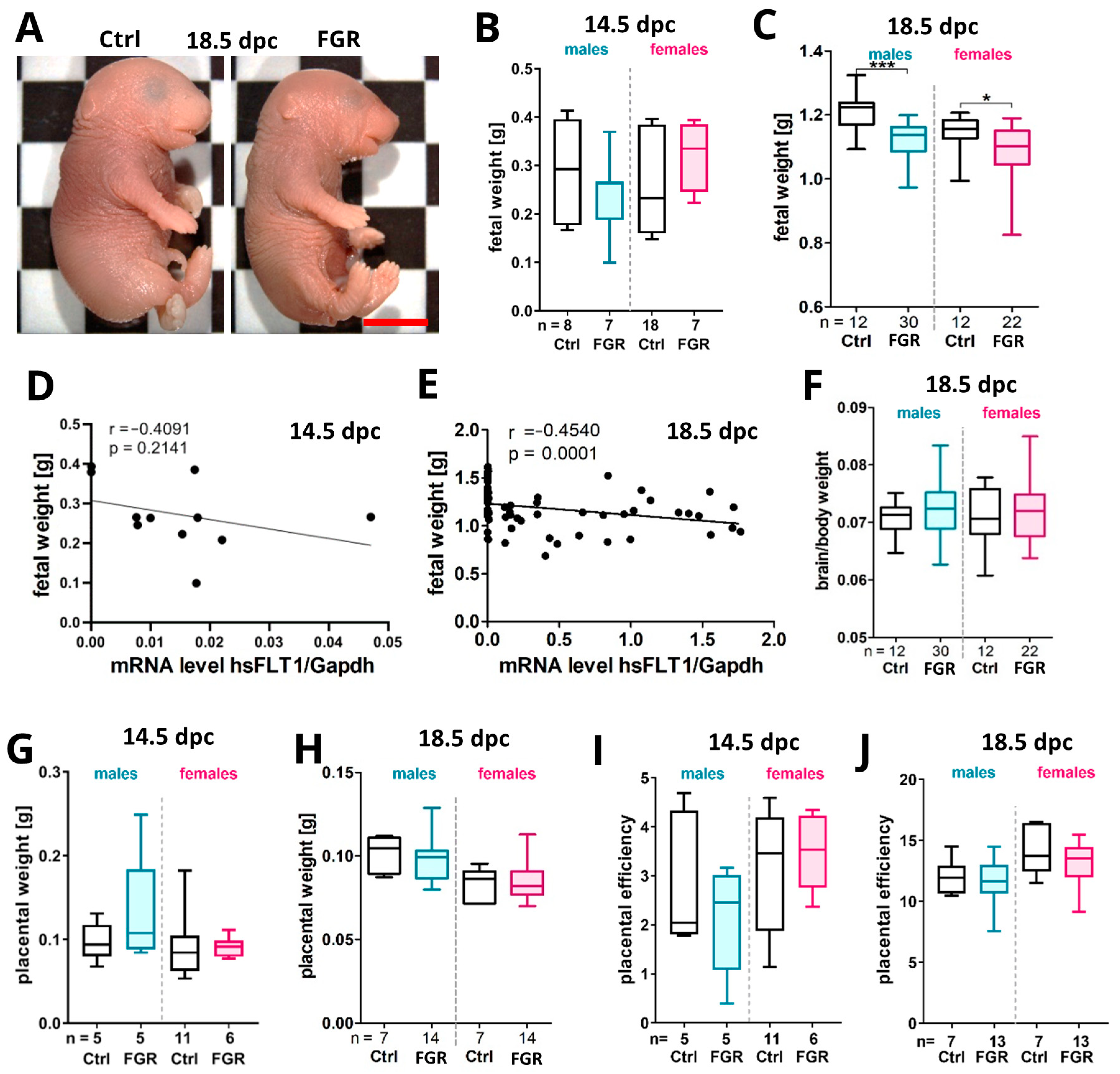
2.2. Placenta-Specific hsFLT1 Overexpression Resulted in Growth-Restricted Fetuses
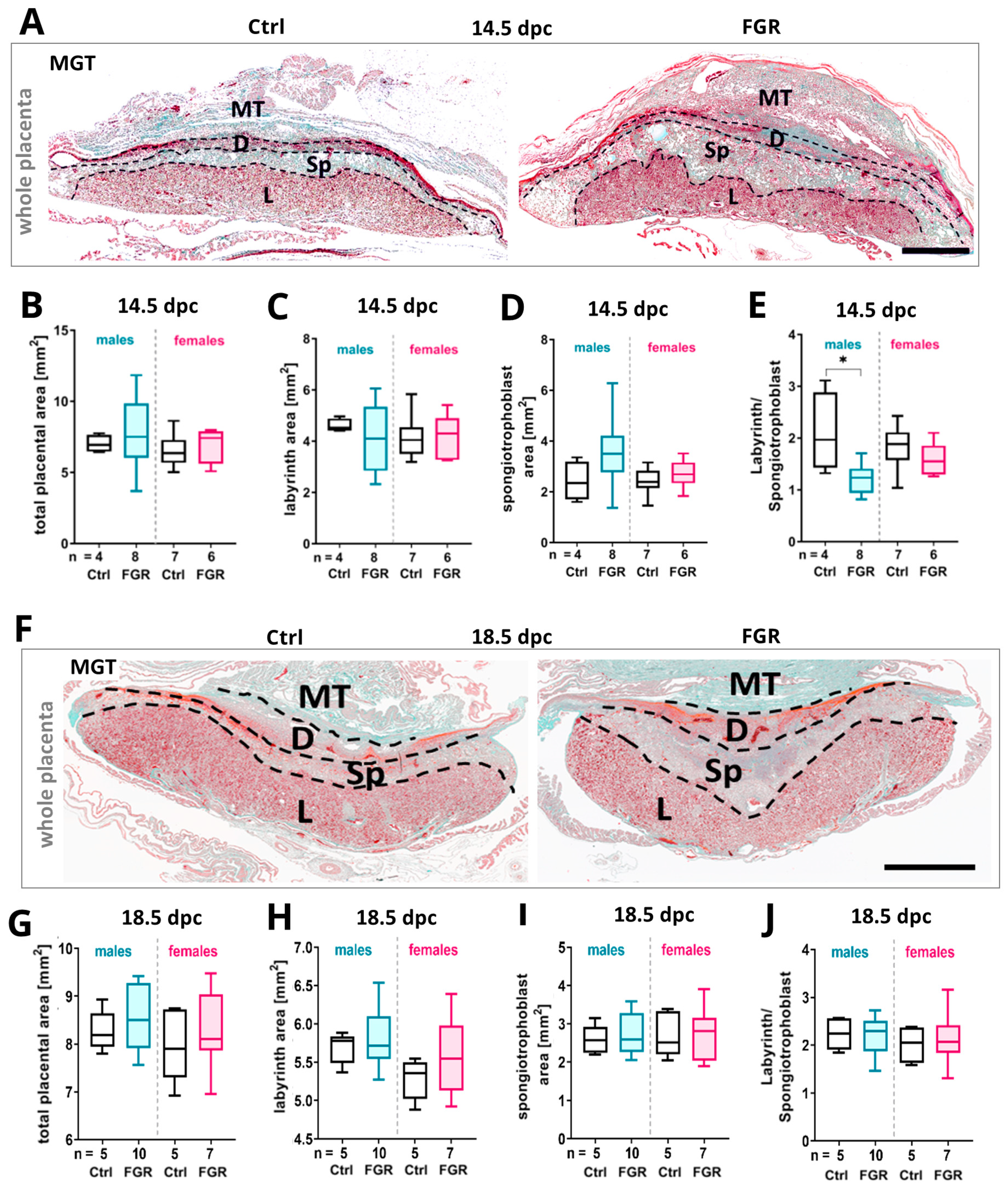
2.3. Expression of Placental hsFLT1 Caused an Increase in Size of the Placental Spongiotrophoblast
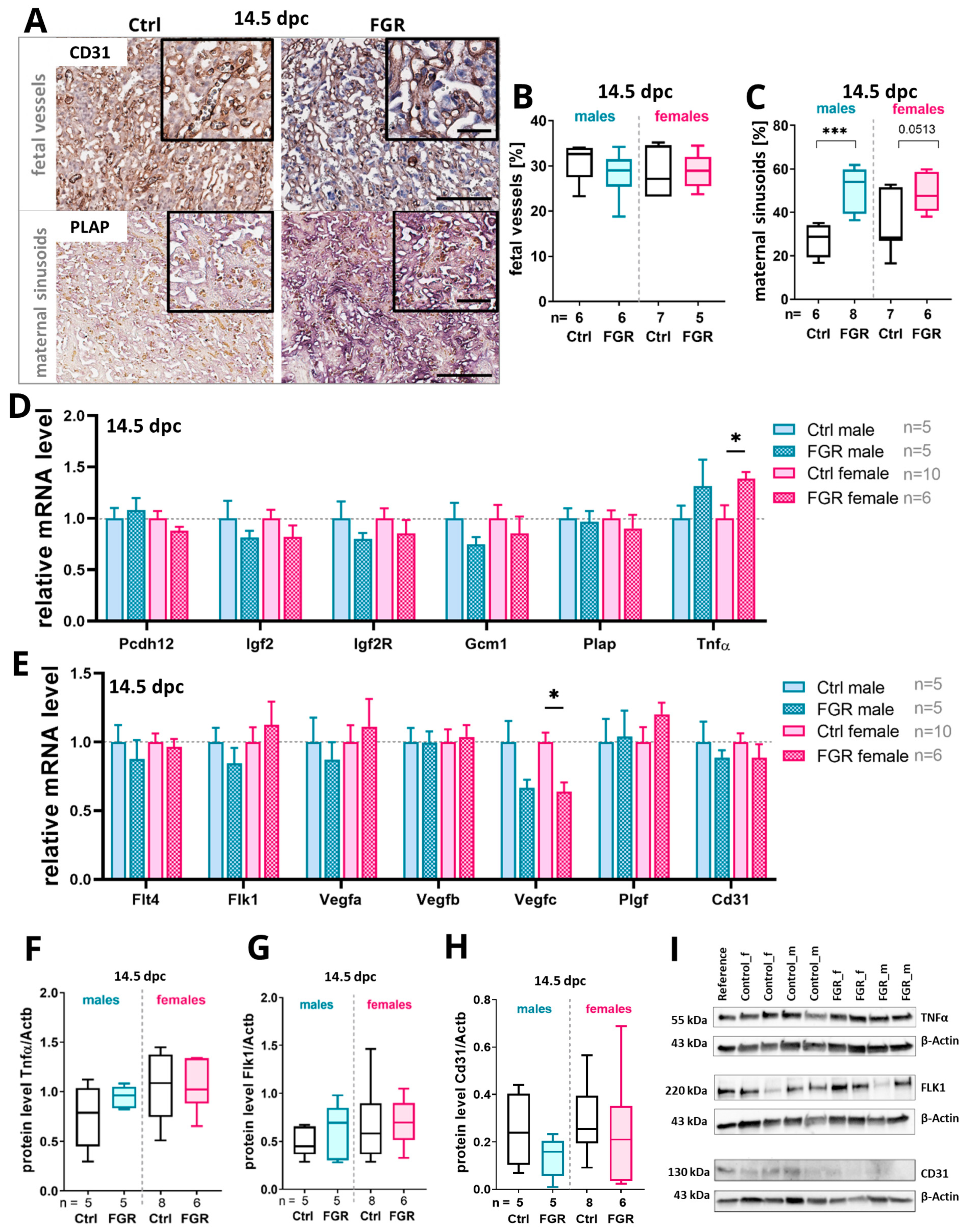
2.4. Elevated Placental hsFLT1 Levels Led to Increased Maternal Vascularization in the Labyrinth at 14.5 dpc
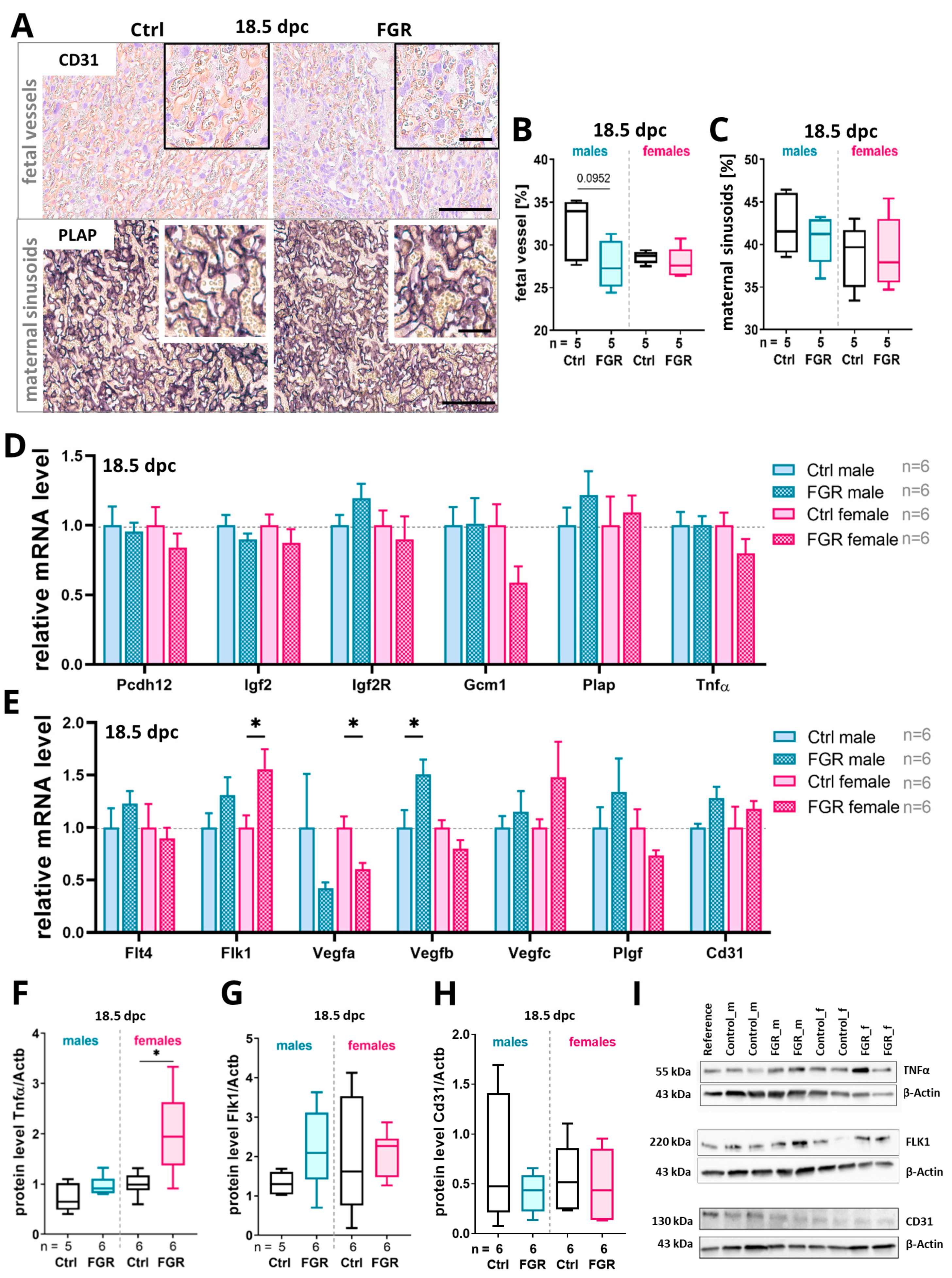
2.5. Placenta-Derived hsFLT1 Led to a Reduced Amount of Fetal Labyrinthine Vessels and to an Accumulation of TNFα at 18.5 dpc
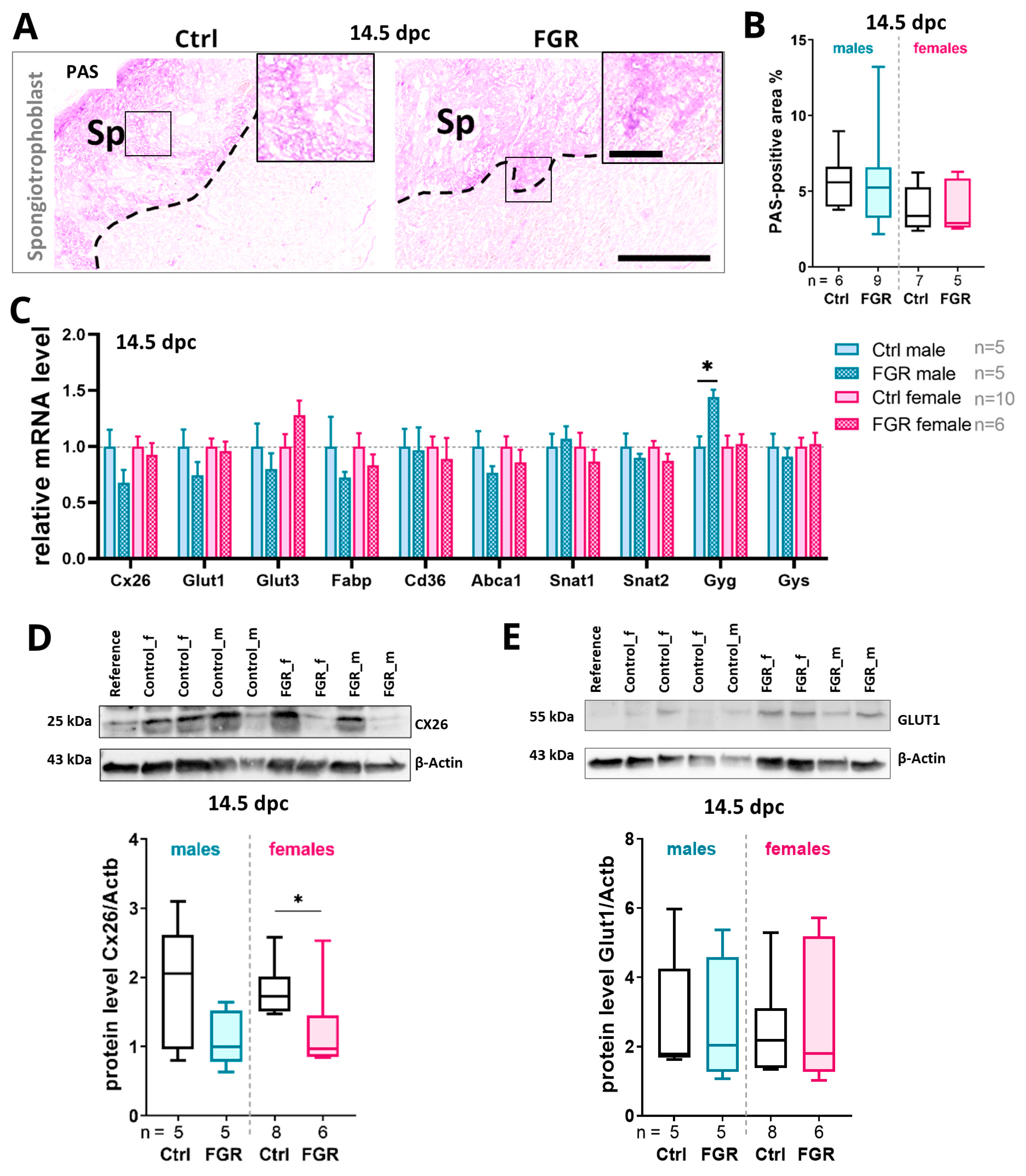
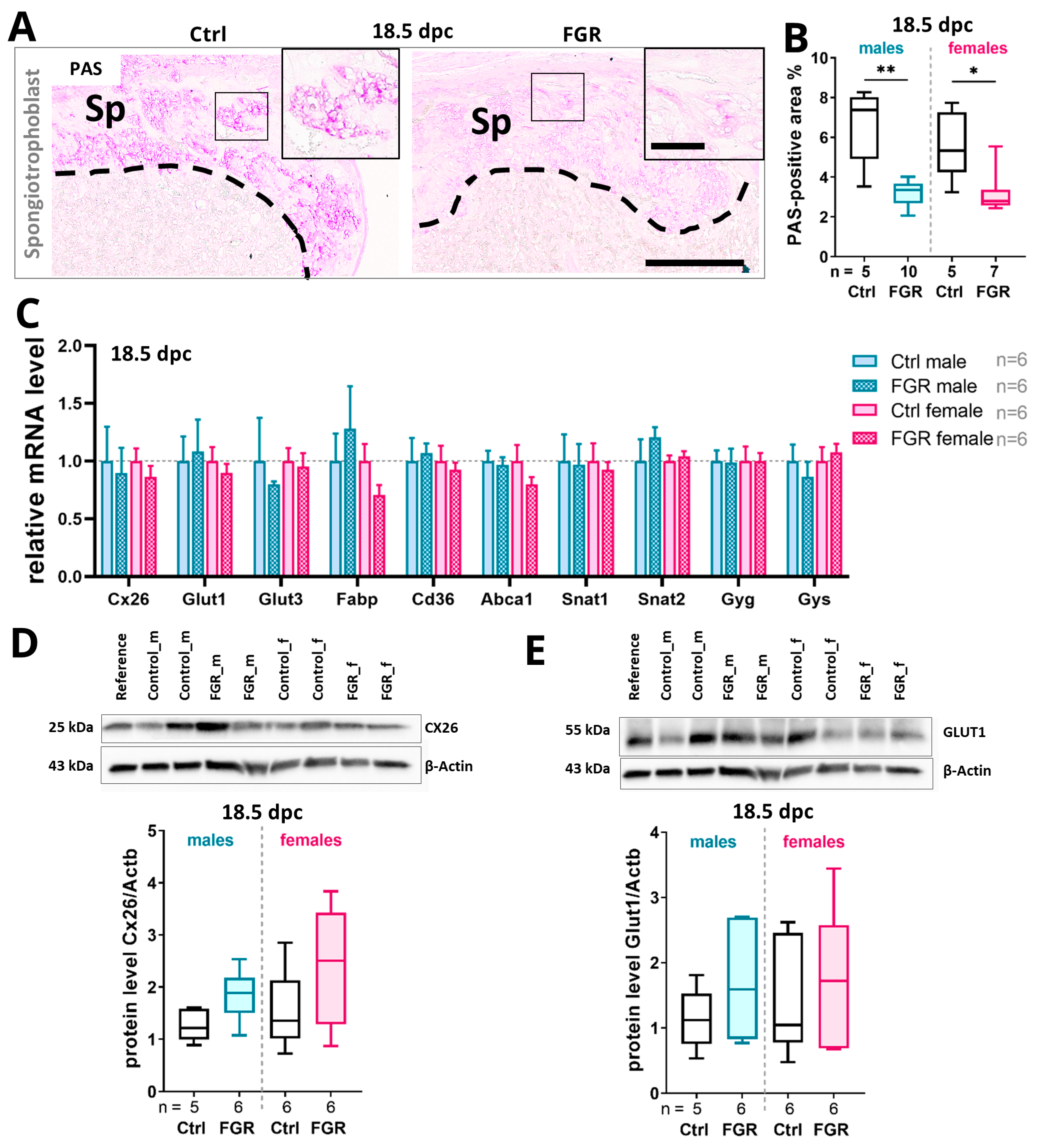
2.6. Placental hsFLT1 Levels Reduced the Expression of Placental Nutrient Transporters and Glycogen Cells in the Spongiotrophoblast
3. Discussion
3.1. The Placental Spongiotrophoblast Compartment Is Strongly Affected by Placenta-Specific hsFLT1 Expression at 14.5 dpc
3.2. Placenta-Specific hsFLT1 Expression Leads to Reduced Expression of Glucose Transporters and Reduced Placental Glycogen Storage
3.3. Placenta-Derived hsFLT1 Leads to Increased Hypoxia, Inflammation, and Disturbed Vascularization at the Feto-Maternal Interface Especially in Males
4. Materials and Methods
4.1. Generation of hsFLT1/tTA/Tpbpa-Cre Mice and Experimental Setting
4.2. Tissue Preparation
4.3. Genomic DNA Isolation, Genotyping, and Sex Determination
4.4. RNA Extraction, cDNA Synthesis, and Quantitative PCR
4.5. Analysis of hsFLT1 Serum Levels
4.6. Histological and Morphometrical Analysis
4.7. Staining with Masson–Goldner Trichrome (MGT)
4.8. Staining with Periodic Acid Schiff Reagent (PAS)
4.9. Staining for Placental Alkaline Phosphatase (PLAP)
4.10. Immunohistochemical Analysis
4.11. Immunoblot Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerdeira, A.S.; Kandzija, N.; Pargmae, P.; Cooke, W.; James, T.; Redman, C.; Vatish, M. Circulating soluble fms-like tyrosine kinase-1 is placentally derived in normal pregnancy: First in vivo evidence. Pregnancy Hypertens. 2019, 16, 145–147. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, T.M.; Walker, S.P.; Hannan, N.J.; Tong, S.; Kaitu’u-Lino, T.J. Clinical tools and biomarkers to predict preeclampsia. EBioMedicine 2022, 75, 103780. [Google Scholar] [CrossRef] [PubMed]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, R.M.; Wynia, K.; Ganzevoort, W. Consensus definition of fetal growth restriction: A Delphi procedure. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Borowicz, P.P.; Caton, J.S.; Crouse, M.S.; Dahlen, C.R.; Ward, A.K. Developmental Programming of Fetal Growth and Development. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 229–247. [Google Scholar] [CrossRef]
- Kumasawa, K.; Ikawa, M.; Kidoya, H.; Hasuwa, H.; Saito-Fujita, T.; Morioka, Y.; Takakura, N.; Kimura, T.; Okabe, M. Pravastatin induces placental growth factor (PGF) and ameliorates preeclampsia in a mouse model. Proc. Natl. Acad. Sci. USA 2011, 108, 1451–1455. [Google Scholar] [CrossRef]
- Clark, D.E.; Smith, S.K.; He, Y.; Day, K.A.; Licence, D.R.; Corps, A.N.; Lammoglia, R.; Charnock-Jones, D.S. A vascular endothelial growth factor antagonist is produced by the human placenta and released into the maternal circulation. Biol. Reprod. 1998, 59, 1540–1548. [Google Scholar] [CrossRef]
- Sakowicz, A.; Bralewska, M.; Kamola, P.; Pietrucha, T. Reliability of Rodent and Rabbit Models in Preeclampsia Research. Int. J. Mol. Sci. 2022, 23, 14344. [Google Scholar] [CrossRef] [PubMed]
- Kuhnel, E.; Kleff, V.; Stojanovska, V.; Kaiser, S.; Waldschutz, R.; Herse, F.; Plosch, T.; Winterhager, E.; Gellhaus, A. Placental-Specific Overexpression of sFlt-1 Alters Trophoblast Differentiation and Nutrient Transporter Expression in an IUGR Mouse Model. J. Cell Biochem. 2017, 118, 1316–1329. [Google Scholar] [CrossRef]
- Sato, E.; Tsunokuni, Y.; Kaneko, M.; Saigusa, D.; Saito, R.; Shimma, S.; Sekimoto, A.; Kawana, Y.; Oe, Y.; Ito, S.; et al. Metabolomics of a mouse model of preeclampsia induced by overexpressing soluble fms-like tyrosine kinase 1. Biochem. Biophys. Res. Commun. 2020, 527, 1064–1071. [Google Scholar] [CrossRef]
- Zhou, Y.; McMaster, M.; Woo, K.; Janatpour, M.; Perry, J.; Karpanen, T.; Alitalo, K.; Damsky, C.; Fisher, S.J. Vascular endothelial growth factor ligands and receptors that regulate human cytotrophoblast survival are dysregulated in severe preeclampsia and hemolysis, elevated liver enzymes, and low platelets syndrome. Am. J. Pathol. 2002, 160, 1405–1423. [Google Scholar] [CrossRef]
- Wei, X.W.; Zhang, Y.C.; Wu, F.; Tian, F.J.; Lin, Y. The role of extravillous trophoblasts and uterine NK cells in vascular remodeling during pregnancy. Front. Immunol. 2022, 13, 951482. [Google Scholar] [CrossRef] [PubMed]
- Admati, I.; Skarbianskis, N.; Hochgerner, H.; Ophir, O.; Weiner, Z.; Yagel, S.; Solt, I.; Zeisel, A. Two distinct molecular faces of preeclampsia revealed by single-cell transcriptomics. Med 2023, 4, 687–709.e7. [Google Scholar] [CrossRef] [PubMed]
- Yagel, S.; Cohen, S.M.; Admati, I.; Skarbianskis, N.; Solt, I.; Zeisel, A.; Beharier, O.; Goldman-Wohl, D. Expert review: Preeclampsia Type I and Type II. Am. J. Obstet. Gynecol. MFM 2023, 5, 101203. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, R.; Kuhnel, E.; Dicke, N.; Verkaik-Schakel, R.N.; Plosch, T.; Schorle, H.; Stojanovska, V.; Herse, F.; Koninger, A.; Kimmig, R.; et al. Human sFLT1 Leads to Severe Changes in Placental Differentiation and Vascularization in a Transgenic hsFLT1/rtTA FGR Mouse Model. Front. Endocrinol. 2019, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, R.; Heupel, J.; Herse, F.; Matin, M.; Hagmann, H.; Bendix, I.; Kraker, K.; Dechend, R.; Winterhager, E.; Kimmig, R.; et al. Circulating Maternal sFLT1 (Soluble fms-Like Tyrosine Kinase-1) Is Sufficient to Impair Spiral Arterial Remodeling in a Preeclampsia Mouse Model. Hypertension 2021, 78, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, R.; Burk, L.V.; Serdar, M.; Kimmig, R.; Bendix, I.; Gellhaus, A. Systemic Maternal Human sFLT1 Overexpression Leads to an Impaired Foetal Brain Development of Growth-Restricted Foetuses upon Experimental Preeclampsia. Oxid. Med. Cell Longev. 2022, 2022, 3024032. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.G.; Fortier, A.L.; Cross, J.C. Diverse subtypes and developmental origins of trophoblast giant cells in the mouse placenta. Dev. Biol. 2007, 304, 567–578. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Tal, R. The role of hypoxia and hypoxia-inducible factor-1alpha in preeclampsia pathogenesis. Biol. Reprod. 2012, 87, 134. [Google Scholar] [CrossRef]
- Yang, Y.; Jin, H.; Qiu, Y.; Liu, Y.; Wen, L.; Fu, Y.; Qi, H.; Baker, P.N.; Tong, C. Reactive Oxygen Species are Essential for Placental Angiogenesis During Early Gestation. Oxid. Med. Cell Longev. 2022, 2022, 4290922. [Google Scholar] [CrossRef]
- Karumanchi, S.A. Angiogenic Factors in Preeclampsia: From Diagnosis to Therapy. Hypertension 2016, 67, 1072–1079. [Google Scholar] [CrossRef]
- Orzack, S.H.; Stubblefield, J.W.; Akmaev, V.R.; Colls, P.; Munne, S.; Scholl, T.; Steinsaltz, D.; Zuckerman, J.E. The human sex ratio from conception to birth. Proc. Natl. Acad. Sci. USA 2015, 112, E2102–E2111. [Google Scholar] [CrossRef]
- Verburg, P.E.; Tucker, G.; Scheil, W.; Erwich, J.J.; Dekker, G.A.; Roberts, C.T. Sexual Dimorphism in Adverse Pregnancy Outcomes—A Retrospective Australian Population Study 1981–2011. PLoS ONE 2016, 11, e0158807. [Google Scholar] [CrossRef]
- Alur, P. Sex Differences in Nutrition, Growth, and Metabolism in Preterm Infants. Front. Pediatr. 2019, 7, 22. [Google Scholar] [CrossRef]
- Kedziora, S.M.; Obermayer, B.; Sugulle, M.; Herse, F.; Kraker, K.; Haase, N.; Langmia, I.M.; Muller, D.N.; Staff, A.C.; Beule, D.; et al. Placental Transcriptome Profiling in Subtypes of Diabetic Pregnancies Is Strongly Confounded by Fetal Sex. Int. J. Mol. Sci. 2022, 23, 15388. [Google Scholar] [CrossRef]
- Surmon, L.; Young, S.; Bobek, G.; Chiu, C.L.; Xu, B.; Makris, A.; Lind, J.M.; Hennessy, A. OS055. Sex-dependent differences in expression of FLT-1 variants andJMJD6 in mouse placenta. Pregnancy Hypertens. 2012, 2, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Stojanovska, V.; Dijkstra, D.J.; Vogtmann, R.; Gellhaus, A.; Scherjon, S.A.; Plosch, T. A double-hit pre-eclampsia model results in sex-specific growth restriction patterns. Dis. Models Mech. 2019, 12, dmm035980. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.; Perez-Garcia, V.; Hemberger, M. Regulation of Placental Development and Its Impact on Fetal Growth-New Insights From Mouse Models. Front. Endocrinol. 2018, 9, 570. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, T.A.; Sparrow, D.B.; Scott, A.N.; Withington, S.L.; Preis, J.I.; Michalicek, J.; Clements, M.; Tsang, T.E.; Shioda, T.; Beddington, R.S.; et al. Cited1 is required in trophoblasts for placental development and for embryo growth and survival. Mol. Cell Biol. 2004, 24, 228–244. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.T.; Soloveva, V.; Tzeng, S.J.; Lowe, L.A.; Pfendler, K.C.; Iannaccone, P.M.; Kuehn, M.R.; Linzer, D.I. Nodal regulates trophoblast differentiation and placental development. Dev. Biol. 2001, 236, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Tunster, S.J.; Creeth, H.D.J.; John, R.M. The imprinted Phlda2 gene modulates a major endocrine compartment of the placenta to regulate placental demands for maternal resources. Dev. Biol. 2016, 409, 251–260. [Google Scholar] [CrossRef]
- Li, Y.; Behringer, R.R. Esx1 is an X-chromosome-imprinted regulator of placental development and fetal growth. Nat. Genet. 1998, 20, 309–311. [Google Scholar] [CrossRef]
- Rampon, C.; Bouillot, S.; Climescu-Haulica, A.; Prandini, M.H.; Cand, F.; Vandenbrouck, Y.; Huber, P. Protocadherin 12 deficiency alters morphogenesis and transcriptional profile of the placenta. Physiol. Genom. 2008, 34, 193–204. [Google Scholar] [CrossRef]
- Shin, B.C.; Suzuki, T.; Matsuzaki, T.; Tanaka, S.; Kuraoka, A.; Shibata, Y.; Takata, K. Immunolocalization of GLUT1 and connexin 26 in the rat placenta. Cell Tissue Res. 1996, 285, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, H.D.; Jung, D.; Butzler, C.; Temme, A.; Traub, O.; Winterhager, E.; Willecke, K. Transplacental uptake of glucose is decreased in embryonic lethal connexin26-deficient mice. J. Cell Biol. 1998, 140, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Redline, R.W.; Chernicky, C.L.; Tan, H.Q.; Ilan, J.; Ilan, J. Differential expression of insulin-like growth factor-II in specific regions of the late (post day 9.5) murine placenta. Mol. Reprod. Dev. 1993, 36, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Sibley, C.P.; Coan, P.M.; Ferguson-Smith, A.C.; Dean, W.; Hughes, J.; Smith, P.; Reik, W.; Burton, G.J.; Fowden, A.L.; Constancia, M. Placental-specific insulin-like growth factor 2 (Igf2) regulates the diffusional exchange characteristics of the mouse placenta. Proc. Natl. Acad. Sci. USA 2004, 101, 8204–8208. [Google Scholar] [CrossRef] [PubMed]
- Reuss, B.; Hellmann, P.; Dahl, E.; Traub, O.; Butterweck, A.; Grummer, R.; Winterhager, E. Connexins and E-cadherin are differentially expressed during trophoblast invasion and placenta differentiation in the rat. Dev. Dyn. 1996, 205, 172–182. [Google Scholar] [CrossRef]
- Carter, A.M. Placental Gas Exchange and the Oxygen Supply to the Fetus. Compr. Physiol. 2015, 5, 1381–1403. [Google Scholar]
- Granger, J.P.; Alexander, B.T.; Llinas, M.T.; Bennett, W.A.; Khalil, R.A. Pathophysiology of preeclampsia: Linking placental ischemia/hypoxia with microvascular dysfunction. Microcirculation 2002, 9, 147–160. [Google Scholar] [CrossRef]
- Colson, A.; Sonveaux, P.; Debieve, F.; Sferruzzi-Perri, A.N. Adaptations of the human placenta to hypoxia: Opportunities for interventions in fetal growth restriction. Hum. Reprod. Update 2021, 27, 531–569. [Google Scholar] [CrossRef]
- Sasagawa, T.; Nagamatsu, T.; Yanagisawa, M.; Fujii, T.; Shibuya, M. Hypoxia-inducible factor-1beta is essential for upregulation of the hypoxia-induced FLT1 gene in placental trophoblasts. Mol. Hum. Reprod. 2021, 27, gaab065. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Yang, Y.P.; Hu, X.L.; Yelavarthi, K.K.; Fishback, J.L.; Hunt, J.S. Tumor necrosis factor alpha mRNA and protein are present in human placental and uterine cells at early and late stages of gestation. Am. J. Pathol. 1991, 139, 327–335. [Google Scholar] [PubMed]
- Raghupathy, R.; Al-Azemi, M.; Azizieh, F. Intrauterine growth restriction: Cytokine profiles of trophoblast antigen-stimulated maternal lymphocytes. Clin. Dev. Immunol. 2012, 2012, 734865. [Google Scholar] [CrossRef] [PubMed]
- Fakhr, Y.; Koshti, S.; Habibyan, Y.B.; Webster, K.; Hemmings, D.G. Tumor Necrosis Factor-alpha Induces a Preeclamptic-like Phenotype in Placental Villi via Sphingosine Kinase 1 Activation. Int. J. Mol. Sci. 2022, 23, 3750. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Jain, A.K.; Mittal, P.; Kohli, M.; Jawanjal, P.; Rath, G. Association of pro- and anti-inflammatory cytokines in preeclampsia. J. Clin. Lab. Anal. 2019, 33, e22834. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Walsh, S.W. TNF alpha concentrations and mRNA expression are increased in preeclamptic placentas. J. Reprod. Immunol. 1996, 32, 157–169. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogtmann, R.; Riedel, A.; Sassmannshausen, I.; Langer, S.; Kühnel-Terjung, E.; Kimmig, R.; Schorle, H.; Winterhager, E.; Gellhaus, A. Overexpression of Human sFLT1 in the Spongiotrophoblast Is Sufficient to Induce Placental Dysfunction and Fetal Growth Restriction in Transgenic Mice. Int. J. Mol. Sci. 2024, 25, 2040. https://doi.org/10.3390/ijms25042040
Vogtmann R, Riedel A, Sassmannshausen I, Langer S, Kühnel-Terjung E, Kimmig R, Schorle H, Winterhager E, Gellhaus A. Overexpression of Human sFLT1 in the Spongiotrophoblast Is Sufficient to Induce Placental Dysfunction and Fetal Growth Restriction in Transgenic Mice. International Journal of Molecular Sciences. 2024; 25(4):2040. https://doi.org/10.3390/ijms25042040
Chicago/Turabian StyleVogtmann, Rebekka, Alina Riedel, Ivanka Sassmannshausen, Sarah Langer, Elisabeth Kühnel-Terjung, Rainer Kimmig, Hubert Schorle, Elke Winterhager, and Alexandra Gellhaus. 2024. "Overexpression of Human sFLT1 in the Spongiotrophoblast Is Sufficient to Induce Placental Dysfunction and Fetal Growth Restriction in Transgenic Mice" International Journal of Molecular Sciences 25, no. 4: 2040. https://doi.org/10.3390/ijms25042040
APA StyleVogtmann, R., Riedel, A., Sassmannshausen, I., Langer, S., Kühnel-Terjung, E., Kimmig, R., Schorle, H., Winterhager, E., & Gellhaus, A. (2024). Overexpression of Human sFLT1 in the Spongiotrophoblast Is Sufficient to Induce Placental Dysfunction and Fetal Growth Restriction in Transgenic Mice. International Journal of Molecular Sciences, 25(4), 2040. https://doi.org/10.3390/ijms25042040