
The Potential Role of Probiotics, Especially Butyrate Producers, in the Management of Gastrointestinal Mucositis Induced by Oncologic Chemo-Radiotherapy


 ,
, 

Abstract
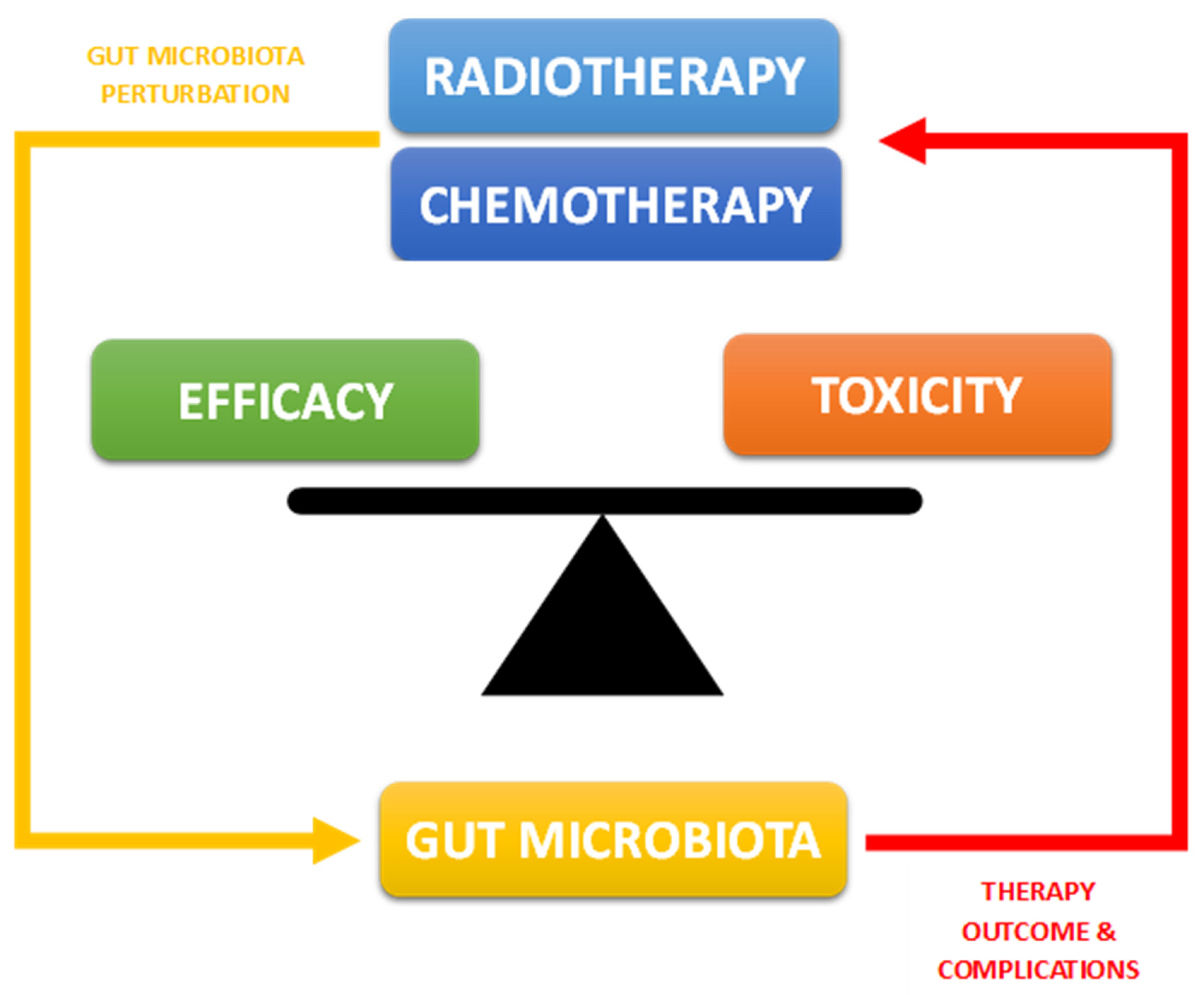
:1. Introduction
2. Alterations in the Intestinal Microbiota during Chemo-Radiotherapy
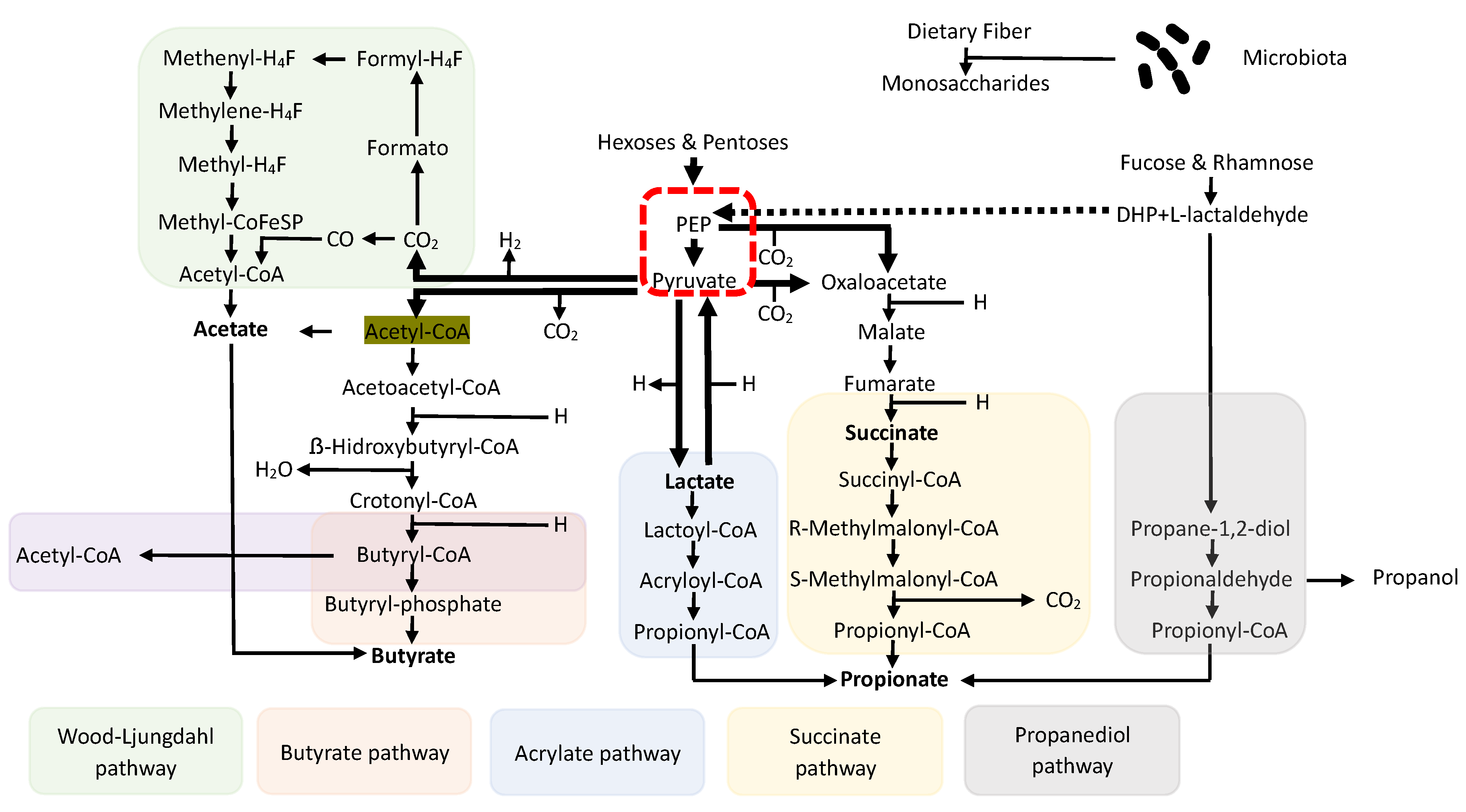
3. Short-Chain Fatty Acids (SCFAs)
4. Oncological-Treatment-Induced Mucositis and Its Correlation with SCFAs
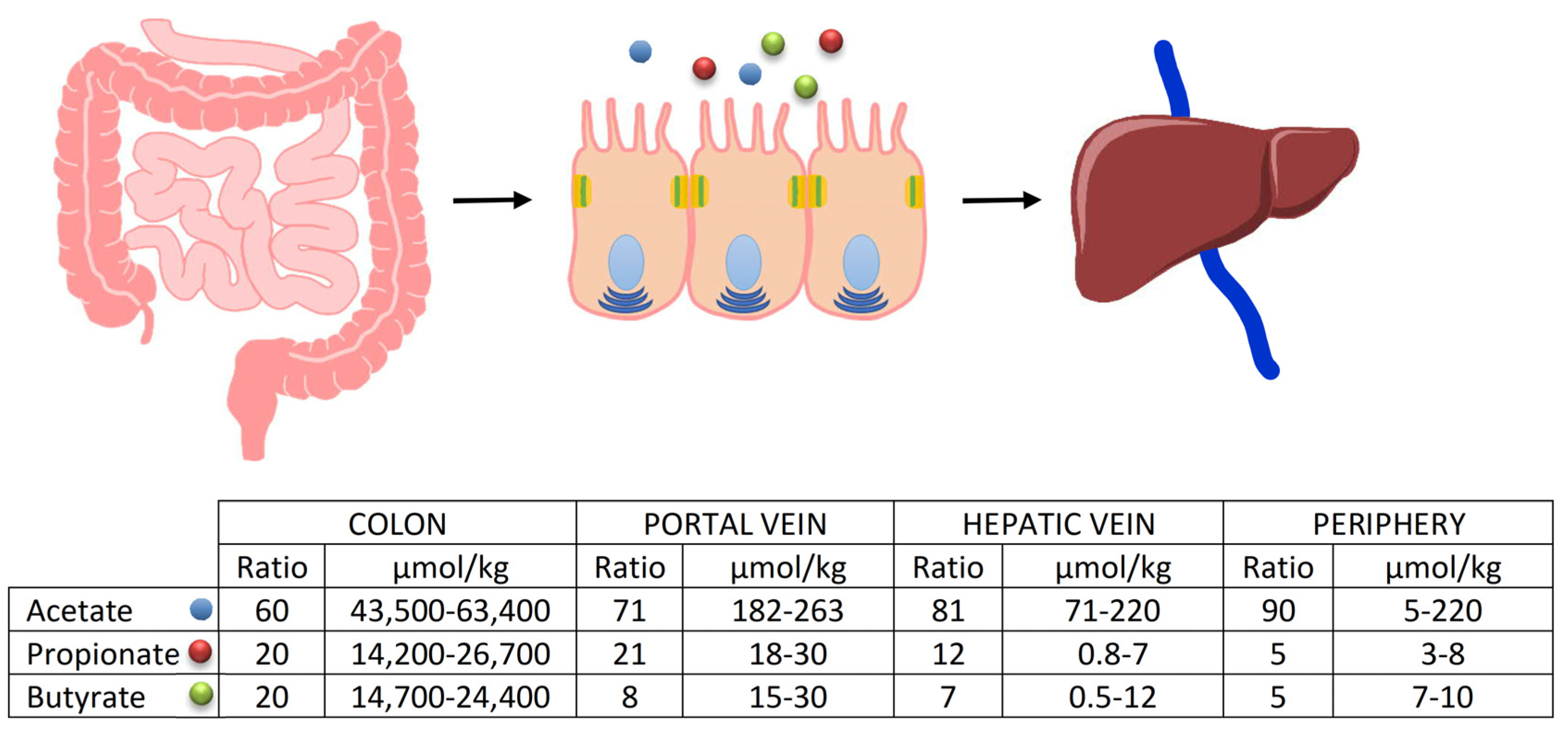
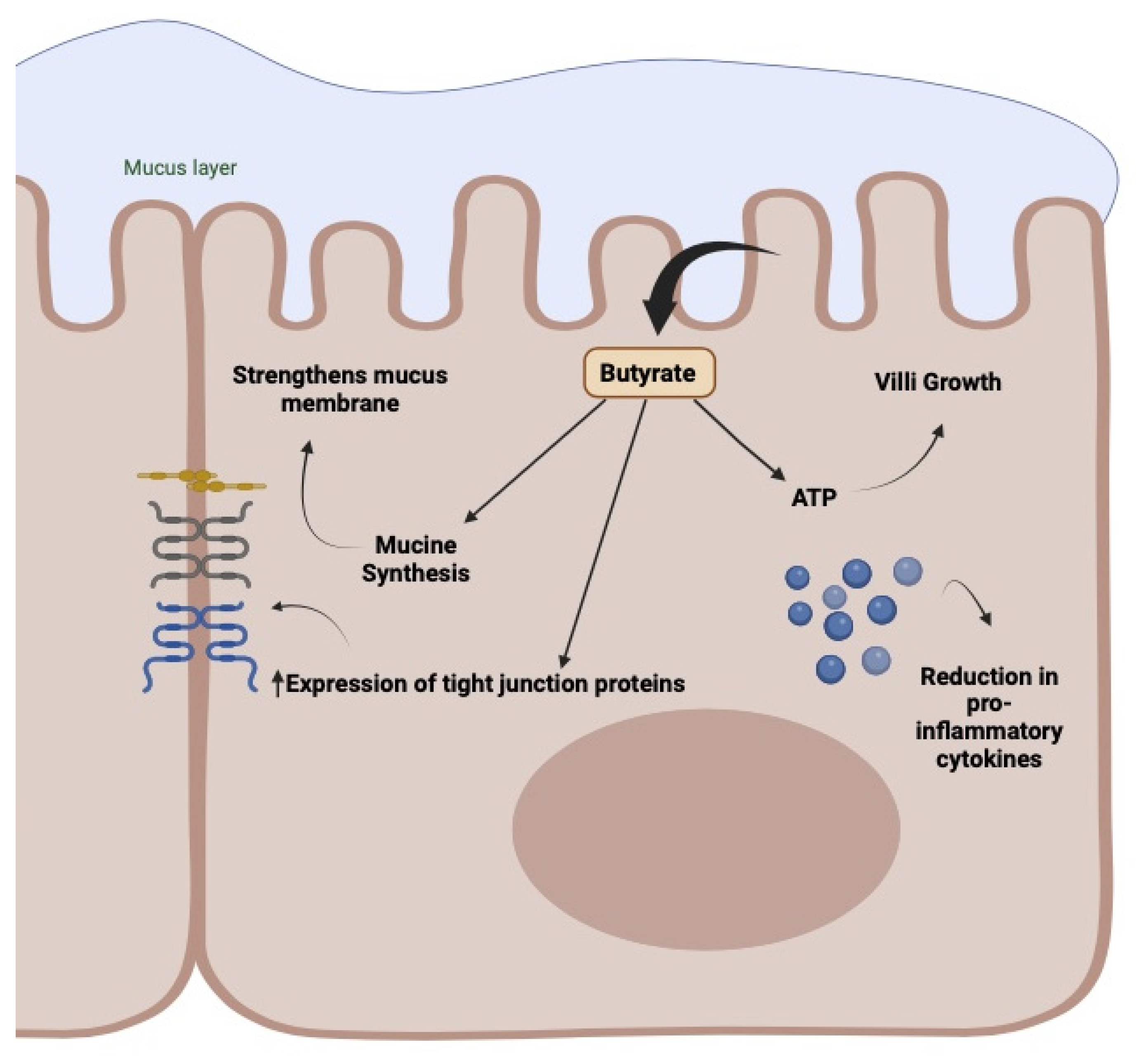
5. The Increase in Circulating Butyrate and Its Impact on the Treatment of Gastrointestinal Disorders
6. Clostridium butyricum CBM 588
6.1. Clostridium butyricum CBM 588’s Effects in Animal Models
6.2. Clostridium butyricum CBM 588’s Effects in Clinical Evaluations
6.3. Clostridium butyricum CBM 588’s Effects on Digestive Disease Models
7. The Potential Oncological Applications of Clostridium butyricum CBM 588
8. Limitations and Future Developments
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rahman, M.M.; Islam, M.R.; Shohag, S.; Ahasan, M.T.; Sarkar, N.; Khan, H.; Hasan, A.M.; Cavalu, S.; Rauf, A. Microbiome in Cancer: Role in Carcinogenesis and Impact in Therapeutic Strategies. Biomed. Pharmacother. 2022, 149, 112898. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, G. Microbiota: A Key Orchestrator of Cancer Therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef]
- Van Vliet, M.J.; Harmsen, H.J.M.; De Bont, E.S.J.M.; Tissing, W.J.E. The Role of Intestinal Microbiota in the Development and Severity of Chemotherapy-Induced Mucositis. PLoS Pathog. 2010, 6, e1000879. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Wei, K.; He, J.; Ding, N.; Hua, J.; Zhou, T.; Niu, F.; Zhou, G.; Shi, T.; et al. Review: Effect of Gut Microbiota and Its Metabolite SCFAs on Radiation-Induced Intestinal Injury. Front. Cell. Infect. Microbiol. 2021, 11, 577236. [Google Scholar] [CrossRef]
- Touchefeu, Y.; Montassier, E.; Nieman, K.; Gastinne, T.; Potel, G.; Bruley Des Varannes, S.; Le Vacon, F.; De La Cochetière, M.F. Systematic Review: The Role of the Gut Microbiota in Chemotherapy– or Radiation–induced Gastrointestinal Mucositis–Current Evidence and Potential Clinical Applications. Aliment. Pharmacol. Ther. 2014, 40, 409–421. [Google Scholar] [CrossRef]
- Fernandes, A.; Oliveira, A.; Soares, R.; Barata, P. The Effects of Ionizing Radiation on Gut Microbiota, a Systematic Review. Nutrients 2021, 13, 3025. [Google Scholar] [CrossRef]
- Panebianco, C.; Andriulli, A.; Pazienza, V. Pharmacomicrobiomics: Exploiting the Drug-Microbiota Interactions in Anticancer Therapies. Microbiome 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, M.J.; Tissing, W.J.E.; Dun, C.A.J.; Meessen, N.E.L.; Kamps, W.A.; De Bont, E.S.J.M.; Harmsen, H.J.M. Chemotherapy Treatment in Pediatric Patients with Acute Myeloid Leukemia Receiving Antimicrobial Prophylaxis Leads to a Relative Increase of Colonization with Potentially Pathogenic Bacteria in the Gut. Clin. Infect. Dis. 2009, 49, 262–270. [Google Scholar] [CrossRef]
- Von Bültzingslöwen, I.; Adlerberth, I.; Wold, A.E.; Dahlén, G.; Jontell, M. Oral and Intestinal Microflora in 5–fluorouracil Treated Rats, Translocation to Cervical and Mesenteric Lymph Nodes and Effects of Probiotic Bacteria. Oral. Microbiol. Immunol. 2003, 18, 278–284. [Google Scholar] [CrossRef]
- Stringer, A.M.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Yeoh, A.S.J.; Hamilton, J.; Keefe, D.M.K. Gastrointestinal Microflora and Mucins May Play a Critical Role in the Development of 5-Fluorouracil-Induced Gastrointestinal Mucositis. Exp. Biol. Med. 2009, 234, 430–441. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, K.; Qu, J.; Wang, X. The Changes Induced by Cyclophosphamide in Intestinal Barrier and Microflora in Mice. Eur. J. Pharmacol. 2013, 714, 120–124. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X. Effects of Cyclophosphamide on Immune System and Gut Microbiota in Mice. Microbiol. Res. 2015, 171, 97–106. [Google Scholar] [CrossRef]
- Lin, X.B.; Dieleman, L.A.; Ketabi, A.; Bibova, I.; Sawyer, M.B.; Xue, H.; Field, C.J.; Baracos, V.E.; Gänzle, M.G. Irinotecan (CPT-11) Chemotherapy Alters Intestinal Microbiota in Tumour Bearing Rats. PLoS ONE 2012, 7, e39764. [Google Scholar] [CrossRef]
- Montassier, E.; Gastinne, T.; Vangay, P.; Al–Ghalith, G.A.; Bruley Des Varannes, S.; Massart, S.; Moreau, P.; Potel, G.; De La Cochetière, M.F.; Batard, E.; et al. Chemotherapy–driven Dysbiosis in the Intestinal Microbiome. Aliment. Pharmacol. Ther. 2015, 42, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Adamberg, K.; Jaagura, M.; Copetti, M.; Fontana, A.; Adamberg, S.; Kolk, K.; Vilu, R.; Andriulli, A.; Pazienza, V. Influence of Gemcitabine Chemotherapy on the Microbiota of Pancreatic Cancer Xenografted Mice. Cancer Chemother. Pharmacol. 2018, 81, 773–782. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic Distribution of Three Pathways for Propionate Production within the Human Gut Microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Huazano-García, A.; López, M.G. Agavins Reverse the Metabolic Disorders in Overweight Mice through the Increment of Short Chain Fatty Acids and Hormones. Food Funct. 2015, 6, 3720–3727. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; Van Harsselaar, J.; et al. Short Chain Fatty Acids in Human Gut and Metabolic Health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Sougiannis, A.T.; VanderVeen, B.N.; Davis, J.M.; Fan, D.; Murphy, E.A. Understanding Chemotherapy-Induced Intestinal Mucositis and Strategies to Improve Gut Resilience. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 320, G712–G719. [Google Scholar] [CrossRef]
- Dahlgren, D.; Sjöblom, M.; Hellström, P.M.; Lennernäs, H. Chemotherapeutics-Induced Intestinal Mucositis: Pathophysiology and Potential Treatment Strategies. Front. Pharmacol. 2021, 12, 681417. [Google Scholar] [CrossRef]
- Al-Qadami, G.; Van Sebille, Y.; Le, H.; Bowen, J. Gut Microbiota: Implications for Radiotherapy Response and Radiotherapy-Induced Mucositis. Expert. Rev. Gastroenterol. Hepatol. 2019, 13, 485–496. [Google Scholar] [CrossRef]
- Sonis, S.T. Pathobiology of Mucositis. Semin. Oncol. Nurs. 2004, 20, 11–15. [Google Scholar] [CrossRef]
- Bertuccioli, A.; Ninfali, P. The Mediterranean Diet in the Era of Globalization: The Need to Support Knowledge of Healthy Dietary Factors in the New Socio-Economical Framework. Mediterr. J. Nutr. Metab. 2014, 7, 75–86. [Google Scholar] [CrossRef]
- Gaudier, E.; Jarry, A.; Blottière, H.M.; De Coppet, P.; Buisine, M.P.; Aubert, J.P.; Laboisse, C.; Cherbut, C.; Hoebler, C. Butyrate Specifically Modulates MUC Gene Expression in Intestinal Epithelial Goblet Cells Deprived of Glucose. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1168–G1174. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzmanr, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, Receptor for Niacin and the Commensal Metabolite Butyrate, Suppresses Colonic Inflammation and Carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Miknevicius, P.; Zulpaite, R.; Leber, B.; Strupas, K.; Stiegler, P.; Schemmer, P. The Impact of Probiotics on Intestinal Mucositis during Chemotherapy for Colorectal Cancer: A Comprehensive Review of Animal Studies. Int. J. Mol. Sci. 2021, 22, 9347. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Sakata, S.; Imamura, K.; Iyama, S.; Jodai, T.; Saruwatari, K.; Hamada, S.; Akaike, K.; Anai, M.; Fukusima, K.; et al. Association of Clostridium butyricum Therapy Using the Live Bacterial Product CBM588 with the Survival of Patients with Lung Cancer Receiving Chemoimmunotherapy Combinations. Cancers 2023, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Vinelli, V.; Biscotti, P.; Martini, D.; Del Bo’, C.; Marino, M.; Meroño, T.; Nikoloudaki, O.; Calabrese, F.M.; Turroni, S.; Taverniti, V.; et al. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients 2022, 14, 2559. [Google Scholar] [CrossRef]
- Huang, F.; Li, S.; Chen, W.; Han, Y.; Yao, Y.; Yang, L.; Li, Q.; Xiao, Q.; Wei, J.; Liu, Z.; et al. Postoperative Probiotics Administration Attenuates Gastrointestinal Complications and Gut Microbiota Dysbiosis Caused by Chemotherapy in Colorectal Cancer Patients. Nutrients 2023, 15, 356. [Google Scholar] [CrossRef]
- Hentges, D.J. (Ed.) Human Intestinal Microflora in Health and Disease; Academic Press: New York, NY, USA, 1983; ISBN 978-0-12-341280-5. [Google Scholar]
- Seki, H.; Shiohara, M.; Matsumura, T.; Miyagawa, N.; Tanaka, M.; Komiyama, A.; Kurata, S. Prevention of Antibiotic–associated Diarrhea in Children by Clostridium butyricum MIYAIRI. Pediatr. Int. 2003, 45, 86–90. [Google Scholar] [CrossRef]
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O.; Eid, J. Butyrate-Producing Human Gut Symbiont, Clostridium butyricum, and Its Role in Health and Disease. Gut Microbes 2021, 13, 1907272. [Google Scholar] [CrossRef]
- Ariyoshi, T.; Hagihara, M.; Takahashi, M.; Mikamo, H. Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines 2022, 10, 483. [Google Scholar] [CrossRef]
- Sun, Y.-Y.; Li, M.; Li, Y.-Y.; Li, L.-X.; Zhai, W.-Z.; Wang, P.; Yang, X.-X.; Gu, X.; Song, L.-J.; Li, Z.; et al. The Effect of Clostridium butyricum on Symptoms and Fecal Microbiota in Diarrhea-Dominant Irritable Bowel Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Sci. Rep. 2018, 8, 2964. [Google Scholar] [CrossRef]
- Wang, A.; Ling, Z.; Yang, Z.; Kiela, P.R.; Wang, T.; Wang, C.; Cao, L.; Geng, F.; Shen, M.; Ran, X.; et al. Gut Microbial Dysbiosis May Predict Diarrhea and Fatigue in Patients Undergoing Pelvic Cancer Radiotherapy: A Pilot Study. PLoS ONE 2015, 10, e0126312. [Google Scholar] [CrossRef]
- Bertuccioli, A.; Cardinali, M.; Zonzini, G.; Neri, M.; Palazzi, C.M.; Gregoretti, A.; Cazzaniga, M.; Di Pierro, F. Clostridium butyricum Strain MIYAIRI 588 (CBM588) as a Precision Probiotic Therapy in the Ketogenic Diet: A Possible Application? Microbiol. Res. 2023, 14, 492–506. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The Regulation of Intestinal Mucin MUC2 Expression by Short-Chain Fatty Acids: Implications for Epithelial Protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulen, M.F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K.; et al. Interleukin-23-Independent IL-17 Production Regulates Intestinal Epithelial Permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Ariyoshi, T.; Hagihara, M.; Eguchi, S.; Fukuda, A.; Iwasaki, K.; Oka, K.; Takahashi, M.; Yamagishi, Y.; Mikamo, H. Clostridium butyricum MIYAIRI 588-Induced Protectin D1 Has an Anti-Inflammatory Effect on Antibiotic-Induced Intestinal Disorder. Front. Microbiol. 2020, 11, 587725. [Google Scholar] [CrossRef]
- Gobbetti, T.; Dalli, J.; Colas, R.A.; Federici Canova, D.; Aursnes, M.; Bonnet, D.; Alric, L.; Vergnolle, N.; Deraison, C.; Hansen, T.V.; et al. Protectin D1 n-3 DPA and Resolvin D5 n-3 DPA Are Effectors of Intestinal Protection. Proc. Natl. Acad. Sci. USA 2017, 114, 3963–3968. [Google Scholar] [CrossRef]
- Zhao, Q.; Yang, W.-R.; Wang, X.-H.; Li, G.-Q.; Xu, L.-Q.; Cui, X.; Liu, Y.; Zuo, X.-L. Clostridium butyricum Alleviates Intestinal Low-Grade Inflammation in TNBS-Induced Irritable Bowel Syndrome in Mice by Regulating Functional Status of Lamina Propria Dendritic Cells. World J. Gastroenterol. 2019, 25, 5469–5482. [Google Scholar] [CrossRef]
- Wang, F.-Y. Potential Protective Effects of Clostridium butyricum on Experimental Gastric Ulcers in Mice. World J. Gastroenterol. 2015, 21, 8340. [Google Scholar] [CrossRef]
- Imase, K.; Takahashi, M.; Tanaka, A.; Tokunaga, K.; Sugano, H.; Tanaka, M.; Ishida, H.; Kamiya, S.; Takahashi, S. Efficacy of Clostridium butyricum Preparation Concomitantly with Helicobacter Pylori Eradication Therapy in Relation to Changes in the Intestinal Microbiota. Microbiol. Immunol. 2008, 52, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Chen, J.; Xia, J.; Wang, B.; Liu, H.; Yang, L.; Wang, Y.; Ling, Z. Role of Probiotics in the Treatment of Minimal Hepatic Encephalopathy in Patients with HBV-Induced Liver Cirrhosis. J. Int. Med. Res. 2018, 46, 3596–3604. [Google Scholar] [CrossRef]
- Yasueda, A.; Mizushima, T.; Nezu, R.; Sumi, R.; Tanaka, M.; Nishimura, J.; Kai, Y.; Hirota, M.; Osawa, H.; Nakajima, K.; et al. The Effect of Clostridium butyricum MIYAIRI on the Prevention of Pouchitis and Alteration of the Microbiota Profile in Patients with Ulcerative Colitis. Surg. Today 2016, 46, 939–949. [Google Scholar] [CrossRef]
- Araki, Y.; Andoh, A.; Takizawa, J.; Takizawa, W.; Fujiyama, Y. Clostridium butyricum, a Probiotic Derivative, Suppresses Dextran Sulfate Sodium-Induced Experimental Colitis in Rats. Int. J. Mol. Med. 2004, 13, 577–580. [Google Scholar] [CrossRef]
- Hayashi, A.; Sato, T.; Kamada, N.; Mikami, Y.; Matsuoka, K.; Hisamatsu, T.; Hibi, T.; Roers, A.; Yagita, H.; Ohteki, T.; et al. A Single Strain of Clostridium butyricum Induces Intestinal IL-10-Producing Macrophages to Suppress Acute Experimental Colitis in Mice. Cell Host Microbe 2013, 13, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Sasaki, M.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T.; Kusunoki, M. Preventive Efficacy of Butyrate Enemas and Oral Administration of Clostridium butyricum M588 in Dextran Sodium Sulfate-Induced Colitis in Rats. J. Gastroenterol. 2000, 35, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Ikeda, T.; Sakata, S.; Saruwatari, K.; Sato, R.; Iyama, S.; Jodai, T.; Akaike, K.; Ishizuka, S.; Saeki, S.; et al. Association of Probiotic Clostridium butyricum Therapy with Survival and Response to Immune Checkpoint Blockade in Patients with Lung Cancer. Cancer Immunol. Res. 2020, 8, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Hasegawa, R.; Shibutani, K.; Mikami, Y.; Kawai, F.; Matsuo, T.; Uehara, Y.; Mori, N. Probiotic-Related Clostridium butyricum Bacteremia: A Case Report and Literature Review. Anaerobe 2023, 83, 102770. [Google Scholar] [CrossRef]
- Sato, T.; Kudo, D.; Kushimoto, S. Association between Nutrition Protocol with Clostridium butyricum MIYAIRI 588 and Reduced Incidence of Clostridioides Difficile Infection in Critically Ill Patients: A Single-Center, Before-and-After Study. Surg. Infect. 2022, 23, 483–488. [Google Scholar] [CrossRef]
- Kullar, R.; Goldstein, E.J.C.; Johnson, S.; McFarland, L.V. Lactobacillus Bacteremia and Probiotics: A Review. Microorganisms 2023, 11, 896. [Google Scholar] [CrossRef]
- Akbarali, H.I.; Muchhala, K.H.; Jessup, D.K.; Cheatham, S. Chemotherapy Induced Gastrointestinal Toxicities. In Advances in Cancer Research; Elsevier: Amsterdam, The Netherlands, 2022; Volume 155, pp. 131–166. ISBN 978-0-323-90087-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemotherapeutic Treatment | Microbiota Modifications | Refs |
|---|---|---|
| 5-Fluorouracil | Increase in Gram-negative anaerobes Increased translocation to mesenteric lymph nodes Increase in Clostridium spp., Staphylococcus spp., and Escherichia coli and decrease in Lactobacillus spp. and Bacteroides spp. | [9,10] |
| Cycles I and Il: high-dose cytarabine, daunorubicin, and etoposide; cycle Ill: amsacrine, high-dose cytarabine, and etoposide; cycle IV: mitoxantrone and high-dose cytarabine [9] | Lower total number and diversity of intestinal bacteria; decrease in Bacteroides spp., Clostridium cluster XIVa, Faecalibacterium prausnitzii, and Bifidobacterium spp.; increase in pathogenic enterococci and decrease in streptococci | [8] |
| Cyclophosphamide | Decrease in Clostridium cluster XIa, Roseburia, Lachnospiraceae, Coprococcus, lactobacilli, and enterococci Increased translocation of Gram-positive species to mesenteric lymph nodes and spleen Increased Escherichia coli, Pseudomonas, Enterobacteriaceae, and enterococci Increased Firmicutes/Bacteroidetes ratio Increased Actinobacteria, Bacteroidia, Alphaproteobacteria, Lachnospiraceae, Coriobacteriaceae, Lactobacillaceae, and Staphylococcaceae; decreased Bacteroidetes, Bacilli, Clostridia, Coriobacteria, Mollicutes, Prevotellaceae, 524-7, Alcaligenaceae, and Rhodospinillacene; disappeared Verrucomicrobia and Streptococcacede | [11,12,13] |
| Irinotecan | Increased Clostridium cluster XI (including Peptoclostridium difficile) and Enterobacteriaceae | [14] |
| High-dose carmustine, etoposide, aracytine, and melphalan | Increased Proteobacteria, decreased Firmicutes and Actinobacteria | [15] |
| Gemcitabine | Increased Proteobacteria, Verrucomicrobia, Akkermansia muciniphila, Escherichia coli, and Peptoclostridium difficile Decreased Firmicutes, Bacteroidetes, Bacteroidales, Lachnospiraceae, Ruminococcaceae, Bacteroides acidifaciens, and Lactobacillus animalis | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazzaniga, M.; Cardinali, M.; Di Pierro, F.; Zonzini, G.B.; Palazzi, C.M.; Gregoretti, A.; Zerbinati, N.; Guasti, L.; Bertuccioli, A. The Potential Role of Probiotics, Especially Butyrate Producers, in the Management of Gastrointestinal Mucositis Induced by Oncologic Chemo-Radiotherapy. Int. J. Mol. Sci. 2024, 25, 2306. https://doi.org/10.3390/ijms25042306
Cazzaniga M, Cardinali M, Di Pierro F, Zonzini GB, Palazzi CM, Gregoretti A, Zerbinati N, Guasti L, Bertuccioli A. The Potential Role of Probiotics, Especially Butyrate Producers, in the Management of Gastrointestinal Mucositis Induced by Oncologic Chemo-Radiotherapy. International Journal of Molecular Sciences. 2024; 25(4):2306. https://doi.org/10.3390/ijms25042306
Chicago/Turabian StyleCazzaniga, Massimiliano, Marco Cardinali, Francesco Di Pierro, Giordano Bruno Zonzini, Chiara Maria Palazzi, Aurora Gregoretti, Nicola Zerbinati, Luigina Guasti, and Alexander Bertuccioli. 2024. "The Potential Role of Probiotics, Especially Butyrate Producers, in the Management of Gastrointestinal Mucositis Induced by Oncologic Chemo-Radiotherapy" International Journal of Molecular Sciences 25, no. 4: 2306. https://doi.org/10.3390/ijms25042306
APA StyleCazzaniga, M., Cardinali, M., Di Pierro, F., Zonzini, G. B., Palazzi, C. M., Gregoretti, A., Zerbinati, N., Guasti, L., & Bertuccioli, A. (2024). The Potential Role of Probiotics, Especially Butyrate Producers, in the Management of Gastrointestinal Mucositis Induced by Oncologic Chemo-Radiotherapy. International Journal of Molecular Sciences, 25(4), 2306. https://doi.org/10.3390/ijms25042306