
Autism Spectrum Disorder: Brain Areas Involved, Neurobiological Mechanisms, Diagnoses and Therapies


{kind=link}
{kind=link}
Abstract
:1. Generalities about ASD
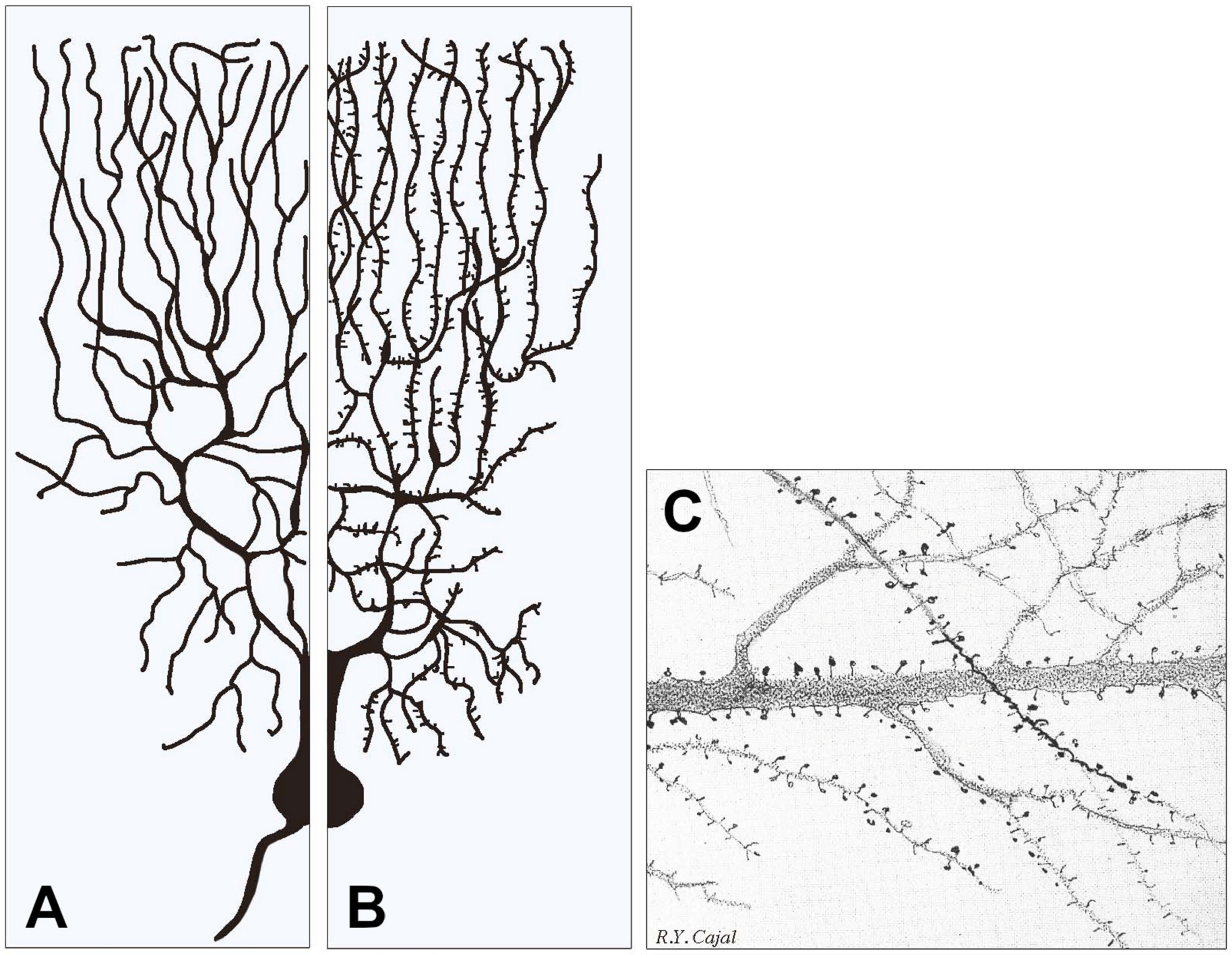
2. The ASD Structures in the Brain
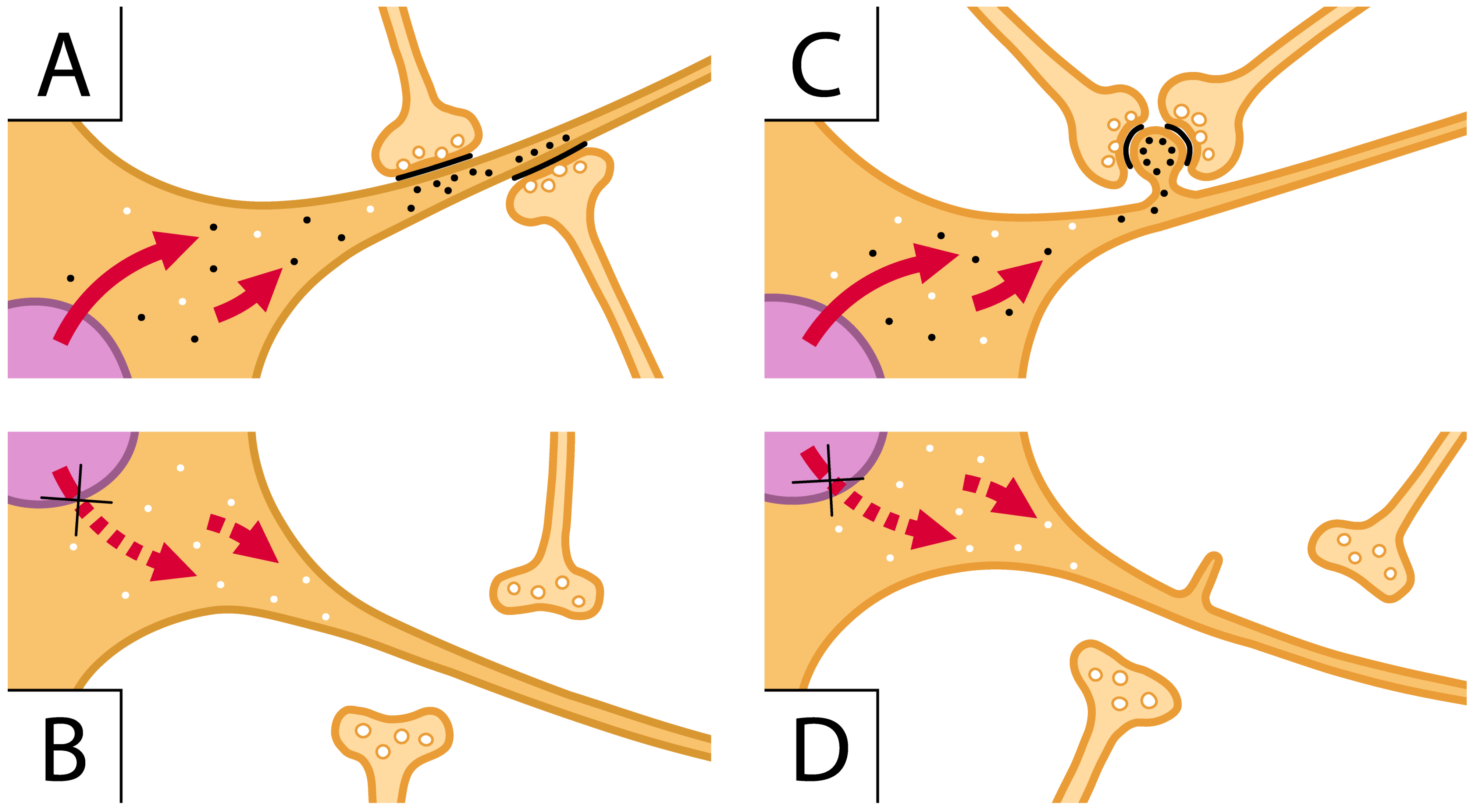
3. Mechanisms of ASD Gene Operations: Key Role of Synapses
4. Contribution of Glial Cells
5. Diagnoses
6. Therapies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADHD | attention deficit of hyperactive disorder |
| ARID1B | subunit of a protein complex active in chromatin remodeling |
| ASD | autism spectrum disorder |
| ID | intellectual disability |
| MRI | magnetic resonance imaging |
References
- Hirota, T.; King, B.H. Autism spectrum disorder: A review. JAMA 2023, 329, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Ford, T.J.L.; Jeon, B.T.; Lee, H.; Kim, W.Y. Dendritic spine and synapse pathology in chromatin. Front. Mol. Neurosci. 2022, 15, 1048713. [Google Scholar] [CrossRef] [PubMed]
- Forstein, S.E.; Rosen-Sheidly, B. Genetics of autism: Complex etiology for a heterogeneous disorder. Nat. Rev. Genet. 2001, 2, 943–955. [Google Scholar] [CrossRef]
- Fombonne, E.J. Epidemiological surveys of autism and other pervasive developmental disorders: An update. Autism Dev. Disord. 2003, 33, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Su Maw, S.; Haga, C. Effectiveness of cognitive, developmental, and behavioral interventions for autism spectrum disorder in preschool-aged children: A systematic review and meta-analysis. Heliyon 2018, 4, e00763. [Google Scholar] [CrossRef] [PubMed]
- Angelink van Entergem, J.A.; Deserno, M.K.; Geurts, H.M. Validation strategies for subtypes in psychiatry: A systematic review of research on autism spectrum disorder. Clin. Psychol. Rev. 2021, 87, 102033. [Google Scholar] [CrossRef]
- Meldolesi, J. Dendritic post-synapses in the brain: Role of flat and spine structures. Biomedicines 2022, 10, 185. [Google Scholar] [CrossRef]
- Maurice, V.; Russet, F.; Scocco, P.; McNicholas, F.; Santosh, P.; Sing, S.P.; Street, C.; Purper-Ouakil, D. Transition from child and adolescent mental health care to adult services for young people with attention-deficit/hyperactivity disorder (ADHD) or autism spectrum disorder (ADS) in Europe: Barriers and recommendations. Encephale 2022, 48, 555–559. [Google Scholar] [CrossRef]
- Boccaccio, F.M.; Platania, A.G.; Savia Guerrera, C.; Varrasi, S.; Privitera, R.C.; Caponnetto, P.; Porrone, C.; Castellano, S. Autism Spectrum Disorder: Recommended psychodiagnostic tools for early diagnosis. Health Psychol. Res. 2023, 11, 77357. [Google Scholar] [CrossRef]
- Park, G.; Jang, W.E.; Kim, S.; Gonzales, E.L.; Ji, J.; Choi, S.; Kim, Y.; Park, J.-H.; Mohammad, H.B.; Bang, G.; et al. Dysregulation of the Wnt/ β-catenin signaling pathway via Rnf146 up-regulation in a VPA-induced mouse model of autism spectrum disorder. Exp. Mol. Med. 2023, 55, 1783–1794. [Google Scholar] [CrossRef]
- Ohyama, S.; Kotani, T.; Iijima, Y.; Okuwaki, S.; Sunami, T.; Iwata, S.; Sakuma, T.; Ogata, Y.; Akazawa, T.; Shiga, Y.; et al. Incidence and potential risk factors of superior mesenteric artery syndrome after spinal corrective surgery in patients with adult spinal deformity. World Neurosurg. 2023, 5, e591–e598. [Google Scholar] [CrossRef] [PubMed]
- Mir, J.M.; Galetta, M.S.; Tretiakov, P.; Dave, P.; Lafage, V.; Lafage, R.; Schoenfeld, A.J.; Passies, P.G. Achievement and maintenance of optimal alignment following adult spinal deformity corrective surgery: A 5 years outcome analysis. World Neurosurg. 2023, 180, e523–e527. [Google Scholar] [CrossRef]
- Lee, B.J.; Bae, S.S.; Choi, H.Y.; Park, J.H.; Hyun, S.J.; Jo, D.J.; Cho, Y. Proximal junctional kyphosis or failure after adult spinal deformity surgery-review of risk factor and its prevention. Neurospine 2023, 20, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.; Mohammed, A.; Akioyamen, N.; Schwab, F.J.; Lafage, V. Preoperative planning for adult spinal deformity goals: Level selection and alignment goals. Neurosurg. Clin. N. Am. 2023, 34, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, Y.M.H.; Almarghany, A.A. Resolution of trifascicular heart block with effective closure of congenital atrial septal defect followed by later coronavirus disease 2019. J. Innov. Card. Rhythm. Manag. 2023, 14, 5533–5536. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, A.; Sakai, K.; Yamada, K.; Utagawa, K.; Hashimoto, J.; Morishita, S.; Matsukura, Y.; Oyaizu, T.; Hirai, T.; Inose, H.; et al. The lower osteotomy level is associated with decreased revision surgery due to mechanical complications after tree column osteotomy in patients with adult spinal deformity. Glob. Spine J. 2023, 18, 21925682231196449. [Google Scholar] [CrossRef] [PubMed]
- Faridi, F.; Seyedebrahimi, A.; Khorowabadi, R. Brain structural covariance network in Asperger syndrome differs from those in autism spectrum disorder and healthy controls. Basic Clin. Neurosci. 2022, 13, 815–838. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Molla, M. Asperger Syndrome; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Li, C.; Zhang, R.; Zhu, Y.; Li, T.; Quin, R.; Li, L.; Yuan, X.; Wang, L.; Wang, X. Gray matter asymmetry alterations in children and adolescents with comorbid autism spectrum disorder and attention-deficit/hyperactivity disorder. Eur. Child Adolesc. Psychiatry 2023, 1–12. [Google Scholar] [CrossRef]
- Kumar, M.; Hiremath, C.; Kumar Khokhar, S.; Bansai, E.; Vijay Sagar, K.J.; Patmanabha, H.; Grimaji, A.S.; Narayan, S.; Kishore, M.T.; Yamini, B.K.; et al. Altered cerebellar lobular volumes correlates with clinical deficits in siblings and children with ASD: Evidence from toddlers. J. Transl. Med. 2023, 21, 246. [Google Scholar] [CrossRef]
- Schumann, C.M.; Bloss, C.S.; Barnes, C.C.; Wideman, G.M.; Carper, R.A.; Aksoomoff, N.; Pierce, K.; Hagler, D.; Schork, N.; Lord, C.; et al. Longitudinal magnetic resonance imaging study of cortical development through early childhood in autism. J. Neurosci. 2010, 30, 4419–4427. [Google Scholar] [CrossRef]
- Akhavan, A.M.; Sharifi, A.; Pedram, M.M. Combination of rs-fMRI and sMRI data to discriminate autism spectrum disorders in young children using deep belief network. J. Digit. Imaging 2018, 31, 895–903. [Google Scholar] [CrossRef]
- Hazlett, H.C.; Gu, H.; Munsell, B.C.; Kim, S.H.; Styner, M.; Wolff, J.J.; Elison, J.D.; Swanson, M.R.; Zhu, H.; Botteron, K.N.; et al. Early brain development in infants at high risk for autism spectrum disorder. Nature 2017, 542, 348–351. [Google Scholar] [CrossRef]
- Ohta, H.; Nordahl, C.W.; Iosif, A.M.; Lee, A.; Rogers, S.; Amaral, D.G. Increased surface area, but not cortical thickness, in a subset of young boys with autism spectrum disorder. Autism Res. 2016, 9, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Andrews, D.S.; Lee, J.K.; Harvey, D.J.; Waizbard-Bartov, E.; Solomon, M.; Rogers, S.J.; Nordhal, C.W.; Amaral, D.J. A longitudinal study of white matter-development in relation to changes in autism severity across early childhood. Biol. Psychiatry 2021, 89, 424–432. [Google Scholar] [CrossRef]
- Sun, F.; Chen, Y.; Gao, Q.; Zhao, Z. Abnormal gray matter structure in children and adolescents with high-functioning autism spectrum disorder. Psychiatry Res. Neuroimaging 2022, 327, 111564. [Google Scholar] [CrossRef] [PubMed]
- Arutiunian, V.; Davydova, E.; Pereverzeva, D.; Sorokin, A.; Tyushkevich, S.; Mamokhina, U.; Danilina, K. Reduced grey matter volume of amygdala and hippocampus is associated with the severity of autistic symptoms and language abilities in school-aged children with autism spectrum disorder: An exploratory study. Brain Struct. Funct. 2023, 228, 1573–1579. [Google Scholar] [CrossRef]
- Kohli, J.S.; Kinnear, M.K.; Fong, C.H.; Fishman, I.; Carper, R.A.; Muller, R.A. Local cortical gyrification is increased in children with autism spectrum disorders, but decreases rapidly in adolescents. Cereb. Cortex. 2019, 29, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, B.; Velasco, S.; Kedaigle, A.J.; Pigoni, M.; Quadrato, G.; Deo, A.J.; Adikonis, X.; Uzquiano, A.; Sartore, R.; Yang, S.M.; et al. Autism genes converge on asynchronous development of shared neuron classes. Nature 2022, 602, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Lenon de Souza Santos, J.; Almeida Arauio, C.; Arauio Gurge Rocha, C.; Costa-Ferro, Z.S.; Solano de Freitas Souza, B. Modeling autism spectrum disorders with induced pluripotent stem cell-derived brain organoids. Biomolecules 2023, 13, 260. [Google Scholar] [CrossRef]
- Cai, C.; Yin, Z.; Liu, A.; Wang, H.; Zeng, S.; Wang, Z.; Qiu, H.; Li, S.; Zhu, X.; Wang, M. Identifying rare genetic variants of immune mediators as risk factors for autism spectrum disorder. Genes 2022, 13, 1098. [Google Scholar] [CrossRef]
- Lima Caldeira, G.; Peca, J.; Carvalho, A.L. New insights on synaptic dysfunction in neuropsychiatric disorders. Curr. Opin. Neurobiol. 2019, 57, 62–70. [Google Scholar] [CrossRef]
- Senarathne, U.D.; Indika, N.L.R.; Izela-Sanek, A.; Ciara, E.; Frye, R.E.; Chen, C.; Stepien, K.M. Biochemical, genetic and clinical diagnostic approaches to autism-associated inherited metabolic disorders. Genes 2023, 14, 803. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Cui, Y.; Yin, Z.; Hou, M.; Guo, P.; Wang, H.; Liu, N.; Cai, C.; Wang, M. Comprehensive systematic and meta-analysis of the association between common genetic variants and autism spectrum disorder. Gene 2023, 18, 147723. [Google Scholar] [CrossRef]
- Dong, D.; Li, J.; Ju, Y.; Xiao, C.; Li, K.; Shi, B.; Zheng, W.; Zhang, Y. Altered relationship between functional connectivity and fiber bundle structure in high functioning male adults with autism spectrum disorder. Brain Sci. 2023, 12, 1098. [Google Scholar] [CrossRef]
- Taketomi, T.; Yasuda, T.; Morita, R.; Kim, J.; Shigeta, Y.; Eroglu, C.; Harada, R.; Tsuruta, F. Autism associated mutations in Hevin/Sparcl1 induces endoplasmic reticulum stress through structural instability. Sci. Rep. 2022, 12, 11891. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, M.; Takayama, J.; Sakai, N.; Kazuno, A.A.; Ishiwata, M.; Ueda, J.; Hayama, T.; Fujji, K.; Samaya, T.; Kuriyama, S.; et al. Deep exome sequencing identifies enrichment of deleterious mosaic variants in neurodevelopmental disorder genes and mitochondrial tRNA regions in bipolar disorder. Mol. Psychiatry 2023, 28, 4294–4306. [Google Scholar] [CrossRef] [PubMed]
- Caracci, M.O.; Avila, M.E.; Espinosa-Cavieres, F.A.; Lopez, H.R.; Ugarte, G.D.; De Ferrari, G.V. Wnt/βcatenin-dependent transcription in autism spectrum disorder. Front. Mol. Neurosci. 2021, 14, 764756. [Google Scholar] [CrossRef]
- Ka, M.; Chopra, D.A.; Dravid, S.M.; Kim, W.Y. Essential roles for ARID1B in dendritic arborization and spine morphology of developing pyramidal neurons. J. Neurosci. 2016, 36, 2723–2742. [Google Scholar] [CrossRef]
- Moffat, J.J.; Jung, E.M.; Ka, M.; Smith, A.J.; Jeon, B.T.; Santen, G.W.; Kim, W.Y. The role of ARID1B, a BAF chromatin remodeling complex subunit, in neural development and behavior. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 30–38. [Google Scholar] [CrossRef]
- Moffat, J.J.; Jung, E.M.; Ka, M.; Jeon, B.T.; Lee, H.; Kim, W.Y. Differential roles of ARID1B in excitatory and inhibitory neural progenitors in the developing cortex. Sci. Rep. 2021, 11, 3856. [Google Scholar] [CrossRef]
- Kim, H.; Kim, D.; Cho, Y.; Kim, K.; Roh, J.D.; Kim, Y.; Yang, E.; Kim, S.S.; Ahn, S.; Kim, H.; et al. Early postnatal serotonin modulation prevents adult-stage deficits in Arid1b deficient mice through synaptic transcriptional reprogramming. Nat. Commun. 2022, 13, 5051. [Google Scholar] [CrossRef]
- Moffat, G.; Zhukovsky, P.; Coughan, G.; Voineskos, A.N. Unravelling the relationship between amyloid accumulation and brain network function in normal aging and very mild cognitive decline: A longitudinal analysis. Brain Commun. 2022, 4, fcac282. [Google Scholar] [CrossRef]
- Nelson, A.D.; Bender, K.J. Dendritic integration dysfunction in neurodevelopment disorders. Dev. Neurosci. 2021, 43, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Vyas, Y.; Garner, C.C.; Montgomery, J.M. Autism associated Shank3 mutations alter mGluR expression and mGluR-dependent NMDA receptor-dependent long-term depression. Synapse 2019, 73, 222097. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Psukus, J.D.; Fingeton, E.; Roche, K.W.; Herring, B.E. Autism spectrum disorder intellectual disability-associated mutations in trio disrupt neuroligin 1-mediated synaptogenesis. J. Neurosci. 2021, 41, 7768–7778. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.Y.; Tuan, L.H.; Chang, H.C.; Wang, Y.C.; Chen, C.H.; Shy, H.C.; Lee, L.J.; Shur-Fen Gau, S. Altered synaptic protein expression, aberrant spine morphology, and impaired spatial memory in Digap2 mutant mice, a genetic model of autism spectrum disorder. Cereb. Cortex 2022, 28, bhac379. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, H.; Hu, B.; Zhang, X.; Liu, A.; Cai, C.; Li, S.; Chen, C.; Wang, Z.; Yin, Z.; et al. Dendritic spine in autism genetics: Whole-exome sequencing identifying de novo variants of CTTNBP2 in a quad family affected by autism spectrum disorder. Children 2022, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Ruiz, C.; Rombach, D.; Cursano, S.; Gerlach-Arbeiter, S.; Schoen, M.; Bockmann, J.; Demestre, M.; Boeckers, T.M. Deletion of the autism-associated protein SHANK3 abolishes structural synaptic plasticity after brain trauma. Int. J. Mol. Sci. 2022, 23, 6081. [Google Scholar] [CrossRef]
- Bucher, M.; Niebling, S.; Han, Y.; Molodenskiy, D.; Hassani-Nia, F.; Kreinkamp, H.J.; Svergun, D.; Kim, E.; Kostiukova, A.S.; Kreutz, M.R. Autism associated SHANK3 missense point mutation impact conformational fluctuations and protein turnover at synapses. eLife 2021, 10, e66165. [Google Scholar] [CrossRef]
- Nishiyama, J. Plasticity of dendritic spines: Molecular function and dysfunction in neurodevelopmental disorders. Psychiatry Clin. Neurosci. 2019, 73, 541–550. [Google Scholar] [CrossRef]
- Lamanna, J.; Isotti, F.; Ferro, M.; Spedini, S.; Racchetti, R.; Musazzi, L.; Malgaroli, A. Occlusion of dopamine-dependent synaptic plasticity in the prefrontal cortex mediates the expression of depressive-like behavior and is medulaed by ketamine. Sci. Rep. 2022, 12, 11055. [Google Scholar] [CrossRef] [PubMed]
- Kumar Tripathi, M.; Kumar Oiha, S.; Kartawy, M.; Hamoundi, W.; Choudhary, A.; Stern, S.; Aran, A.; Amal, H. The NO answer for autism spectrum disorder. Adv. Sci. (Weinh) 2023, 10, e2205783. [Google Scholar] [CrossRef] [PubMed]
- Porokhovnik, L.N.; Pisarev, V.M.; Chumachenko, A.G.; Chudkova, J.M.; Ershova, E.S.; Veiko, N.N.; Gorbachevskaya, N.L.; Mamokhina, U.A.; Sorokin, A.B.; Basova, A.Y. Association of NEF2L2 Rs35652124 polymorphism with Nrf2 induction and genotoxic stress biomarkers in autism. Gene 2023, 14, 718. [Google Scholar] [CrossRef] [PubMed]
- Memis, I.; Mittal, R.; Furar, E.; White, I.; Eshraghi, A.A. Altered blood brain barrier permeability and oxidative stress in Cntnap2 knockout rat model. J. Clin. Med. 2022, 11, 2725. [Google Scholar] [CrossRef] [PubMed]
- Petroni, V.; Subashi, E.; Premoli, M.; Wohr, M.; Crusio, W.E.; Memaire, V.; Pietropaolo, S. Autistic-like behavioral effects of prenatal stress in juvenile Fmr1 mice: The relevance of sex differences and gene environment interactions. Sci. Rep. 2022, 12, 7269. [Google Scholar] [CrossRef]
- Kilpatrick, S.; Irwin, C.; Singh, K.K. Human pluripotent stem cells (hPSC) and organoid models of autism: Opportunities and limitations. Transl. Psychiatry 2023, 13, 217. [Google Scholar] [CrossRef]
- Vakilzadeh, G.; Falcone, C.; Dufour, B.; Hong, T.; Noctor, S.C.; Martinez-Cerdeno, V. Decreased number and increased activation state of astrocytes in gray and white matter of the prefrontal cortex in autism. Cereb. Cortex 2022, 32, 4902–4912. [Google Scholar] [CrossRef]
- Allen, M.; Huang, B.S.; Notaras, M.J.; Lodhi, A.; Barrio-Alonso, E.; Lituma, P.J.; Wolujewicz, P.; Wiztum, J.; Longo, F.; Chen, M.; et al. Astrocytes derived from ASD individuals alter behavior and destabilize neuronal activity through aberrant Ca2+ signaling. Mol. Psychiatry 2022, 27, 2470–2484. [Google Scholar] [CrossRef]
- Vakilzadeh, G.; Martinez-Cerdeno, V. Pathology and astrocytes in autism. Neuropsychiatr. Dis. Treat. 2023, 19, 841–850. [Google Scholar] [CrossRef]
- Guneykaya, D.; Uguru, B.; Logiacco, P.; Popp, O.; Feiks, M.A.; Meyer, N.; Wendt, S.; Semter, M.; Cherif, F.; Gauthier, C.; et al. Sex-specific microglia state in neuroligin-4 knock-out mouse model of autism spectrum disorder. Brain Behav. Immunol. 2023, 111, 61–75. [Google Scholar] [CrossRef]
- Luo, Y.; Lv, K.; Du, Z.; Zhang, D.; Chen, M.; Luo, J.; Wang, L.; Liu, T.; Gong, H.; Fan, X. Monocycline improves autism-related behaviors by modulating microglia polarization in mouse model autism. Int. Immunopharmacol. 2023, 122, 110594. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, J.; Li, Y. Microglia and astrocytes underlie neuroinflammation and synaptic susceptibility in autism spectrum disorder. Front. Neurosci. 2023, 17, 1125428. [Google Scholar] [CrossRef]
- Frasch, M.G.; Yoon, B.J.; Helbing, D.L.; Snir, G.; Antonelli, M.C.; Bauer, R. Autism spectrum disorder: A neuro-immunometabolic hypothesis of the developmental origins. Biology 2023, 12, 914. [Google Scholar] [CrossRef]
- Usui, N.; Kobayashi, H.; Shimida, S. Neuroinflammation and oxidative stress in the pathogenesis of autism spectrum disorder. Int. J. Mol. Sci. 2023, 24, 5487. [Google Scholar] [CrossRef]
- Costa, A.N.; Ferguson, B.J.; Hawkins, E.; Coman, A.; Schauer, J.; Ramirez-Cells, A.; Hecht, P.M.; Bruce, D.; Tilley, M.; Zelebizadeh, Z.; et al. Relationship between maternal antibodies in fetal brain and pre-natal stress exposure in autism spectrum disorder. Metabolites 2023, 13, 663. [Google Scholar] [CrossRef] [PubMed]
- Woo, T.; King, C.; Ahmed, N.I.; Cordes, M.; Nistala, S.; Will, M.J.; Bloomer, C.; Kibiryev, N.; Rivera, M.R.; Telebizadeh, Z.; et al. microRNA as a maternal marker for the prenatal stress-associated ASD, evidence from a murine model. J. Pers. Med. 2023, 13, 1412. [Google Scholar] [CrossRef] [PubMed]
- Harrison Elder, J.; Maun Kreider, C.; Brasher, S.N.; Ansell, M. Clinical impact of early diagnosis of autism on the prognosis and parent-child relationships. Psychol. Res. Behav. Manag. 2017, 10, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Klin, A. Translating advances in developmental social neuroscience into greater access to early diagnosis in autism spectrum disorder. Medicine 2023, 83 (Suppl. 2), 32–36. [Google Scholar]
- Costantino, J.N.; Abbacchi, A.M.; May, B.K.; Klaiman, C.; Zhang, Y.; Lowe, J.K.; Marrus, N.; Klin, A.; Geschwin, D.H. Prospects for leveling the playing field for black children with autism. J. Am. Acad. Child Adolesc. Psychiatry 2023, 62, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Klaiman, C.; Richardson, S.; Lambha, M.; Reid, M.; Hamnes, T.; Beacham, C.; Lewis, P.; Paredes, J.; Edwards, L.; et al. Development and replication of objective measurements of social visual engagement to aid in early diagnosis and assessment of autism. JAMA Netw. Open 2023, 6, e2330145. [Google Scholar] [CrossRef] [PubMed]
- Sobieski, M.; Sobieska, A.; Sekulowicz, M.; Bujnowska-Fedak, M.M. Tools for early screening of autism spectrum disorders in primary health care- a scoping review. BMC Prim. Care 2022, 23, 46. [Google Scholar] [CrossRef]
- Klaiman, C.; White, S.; Richardson, S.; McQueen, E.; Walum, H.; Aoki, C.; Smith, C.; Minjarez, M.; Bernier, R.; Pedapati, E.; et al. Expert clinician certainty in diagnosing autism spectrum disorder in 16-30 months old: A multi-site trial secondary analysis. J. Autism Dev. Disord. 2022, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Strate, P. Autism spectrum disorders in adulthood-symptoms diagnosis. Dtsch. Arztebl. Int. 2023, 120, 87–93. [Google Scholar] [CrossRef]
- Angel, L.; Alley, S.H.; Delaney, K.R.; Mohr, L. Presentation of depressive symptoms in autism spectrum disorders. West. J. Nurs. Res. 2023, 45, 854–861. [Google Scholar] [CrossRef]
- Moffat, J.J.; Smith, A.L.; Jung, A.L.; Ka, M.; Kim, W.Y. Neurobiology of ARID1B haploinsufficiency related to neurodevelopmental and psychiatric disorders. Mol. Psychiatry 2022, 27, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Grzadzinski, R.; Dick, C.; Lord, C.; Bishop, S. Parent-reported and clinician-observed autism spectrum disorder (ASD) symptoms in children with attention deficit/hyperactivity disorder (ADHD): Implications for practice. Mol. Autism 2016, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Jubran, E.; Kumar, D.; Assad, N.D.; Nguyen, H. Variations in anxiety and related in psychiatric comorbidity levels among youth with individual diagnoses of autism spectrum disorder or attention deficit hyperactivity disorder and those with both diagnoses. Cureus 2023, 15, e41759. [Google Scholar] [CrossRef] [PubMed]
- Mannion, A.; Leader, G. Relationship between child sleep problems in autism spectrum disorder and parent mental health and well-being. Sleep Med. 2023, 109, 4–10. [Google Scholar] [CrossRef]
- Mai, A.S.; Yau, C.E.; Tzeng, F.S.; Foo, Q.X.J.; Wang, D.Q.; Tan, E.-K. Linking autism spectrum disorders and parkinsonism: Clinical and genetic association. Ann. Clin. Transl. Neurol. 2023, 10, 484–496. [Google Scholar] [CrossRef]
- Frederiksen, S.D.; Wicki-Stordeur, L.E.; Swayne, L.A. Overlap in synaptic neurological conditions susceptibility pathways and the neuronal pannexin 1 interactome revealed by bioinformatics analyses. Channels 2023, 17, 2253102. [Google Scholar] [CrossRef]
- Das, M.; Zomorrodi, R.; Mirialli, M.; Lirkovski, M.; Blumbrg, D.M.; Rajj, T.K.; Desarkar, P. Machine learning approaches for electroencephalography and magnetoencephaligraphy analyses in autism spectrum disorder. Progr. Neuropsychopharmacol. Biol. Psychiatry 2023, 123, 110705. [Google Scholar] [CrossRef] [PubMed]
- Lamy, M.; Erickson, C.A. Pharmacological management of behavioral disturbances in children and adolescents with autism spectrum disorders. Curr. Probl. Pediatr. Adolesc. Health Care 2018, 48, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Aran, A.; Rand, D.C. Cannabinoid treatment for the symptoms of autism spectrum disorder. Expert Opin. Emerg. Drugs 2024, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rahim, F.; Toguzbaeva, K.; Quasim, N.H.; Dzhusupov, K.O.; Zuhmagaliuly, A.; Koozhamkul, R. Probiotics, prebiotics, and synbiotics for patients with autism spectrum disorder: A meta analysis and umbrella review. Front Nutr. 2023, 10, 1294089. [Google Scholar] [CrossRef] [PubMed]
- Wichers, R.H.; wan der Wouw, L.C.; Brouwer, M.E.; Lok, A.; Bockting, C.L.H. Psychotherapy for co-occurring symptoms of depression, anxiety and obsessive-compulsive disorder in children and adults with autism spectrum disorder: A systematic review and meta-analysis. Psychol. Med. 2023, 53, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.P.; Johnson, K.A.; Riesenberg, R.; Orejudos, A.; Riccobene, T.; Kalluri, H.V.; Malik, P.R.; Varughese, S. Carioprazine in pediatric patients with autism spectrum disorder: Results of a pharmacokinetic, safety and tolerability study. J. Child Adolesc. Psychopharmacol. 2023, 33, 232–242. [Google Scholar] [CrossRef]
- Valdovinos, M.G.; Epperson, C.; Johnson, C. A review of the use of psychotropic medication address challenging behaviour in neurodevelopmental disorders. Int. Rev. Neurobiol. 2023, 173, 43–65. [Google Scholar] [CrossRef]
- Arteaga-Henriquez, G.; Gisbeert, L.; Ramos Quiroga, J.A. Immunoregulatory and/or anti-inflammatory agents for the management of core and associated symptoms in individuals with autism spectrum disorder: A narrative review of randomized, placebo-controlled trials. CNS Drugs 2023, 37, 215–229. [Google Scholar] [CrossRef]
- Caldani, S.; Humeau, E.; Deorme, R.; Bucci, M.P. Inhibition functions can be improved in children with autism spectrum disorders: An eye-tracking study. Int. J. Dev. Neurosci. 2023, 83, 431–441. [Google Scholar] [CrossRef]
- Patel, M.; Atluri, M.L.; Gonzalez, M.A.; Sakhamuri, N.; Athiyaman, S.; Randhi, B.; Gutlpalli, S.D.; Pu, J.; Faldi, M.F.; Khan, S.; et al. Systematic review of mixed studies exploring the effects of probiotics on gut-microbiome to modulate therapy in children with autism spectrum disorder. Cureus 2022, 14, e3213. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, P.; Tao, C.; Lu, L.; Tang, C. Efficacy of non-pharmacological interventions targeting social functions in children and adults with autism spectrum disorder: A systematic review and meta-analysis. PLoS ONE 2023, 18, e0291720. [Google Scholar] [CrossRef]
- Hellings, J. Pharmacotherapy in autism spectrum disorder, including promising drugs warranting trials. World J. Psychiatry 2023, 13, 262–277. [Google Scholar] [CrossRef]
- van der Plas, E.; Mason, D.; Happé, F. Decision making in autism: A narrative review. Autism 2023, 27, 1532–1546. [Google Scholar] [CrossRef]
- Adaralegbe, N.J.; Okobi, O.E.; Omar, Z.T.O.; Segun, E.; Evbayekha, E.O.; Abolurin, A.; Egberuare, E.O.; Ezegbe, H.C.; Adegbosin, A.; Adedeji, A.G.; et al. Impact of adverse childhood experience on resilience and school success in individuals with autism spectrum disorder and attention-deficit hyperactivity disorder. Cureus 2022, 14, e31907. [Google Scholar] [CrossRef] [PubMed]
- Tayanloo-Beik, A.; Hamidpour, S.K.; Abedi, M.; Shojaei, H.; Tavirani, M.R.; Namzi, N.; Larijani, B.; Arjmand, D. Zebrafish modeling of autism spectrum disorders, current status and future prospective. Front. Psychiatry 2022, 13, 911770. [Google Scholar] [CrossRef] [PubMed]
- Etkin, R.G.; Juel, E.K.; Lebwitz, E.R.; Silverman, W. Does cognitive-behavioral therapy for youth anxiety disorders improve social functioning and peer relationships? Clin. Child Fam. Psychol. Rev. 2023, 26, 1052–1076. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Joshi, A.; Singh, R.; Dubey, K. Review of industrially recognized polymers and manufacturing processes for amorphous solid dispersion based formulations. Pharm. Dev. Technol. 2023, 28, 678–696. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamanna, J.; Meldolesi, J. Autism Spectrum Disorder: Brain Areas Involved, Neurobiological Mechanisms, Diagnoses and Therapies. Int. J. Mol. Sci. 2024, 25, 2423. https://doi.org/10.3390/ijms25042423
Lamanna J, Meldolesi J. Autism Spectrum Disorder: Brain Areas Involved, Neurobiological Mechanisms, Diagnoses and Therapies. International Journal of Molecular Sciences. 2024; 25(4):2423. https://doi.org/10.3390/ijms25042423
Chicago/Turabian StyleLamanna, Jacopo, and Jacopo Meldolesi. 2024. "Autism Spectrum Disorder: Brain Areas Involved, Neurobiological Mechanisms, Diagnoses and Therapies" International Journal of Molecular Sciences 25, no. 4: 2423. https://doi.org/10.3390/ijms25042423
APA StyleLamanna, J., & Meldolesi, J. (2024). Autism Spectrum Disorder: Brain Areas Involved, Neurobiological Mechanisms, Diagnoses and Therapies. International Journal of Molecular Sciences, 25(4), 2423. https://doi.org/10.3390/ijms25042423