
Multi-Omics Analysis of the Effects of Soil Amendment on Rapeseed (Brassica napus L.) Photosynthesis under Drip Irrigation with Brackish Water

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
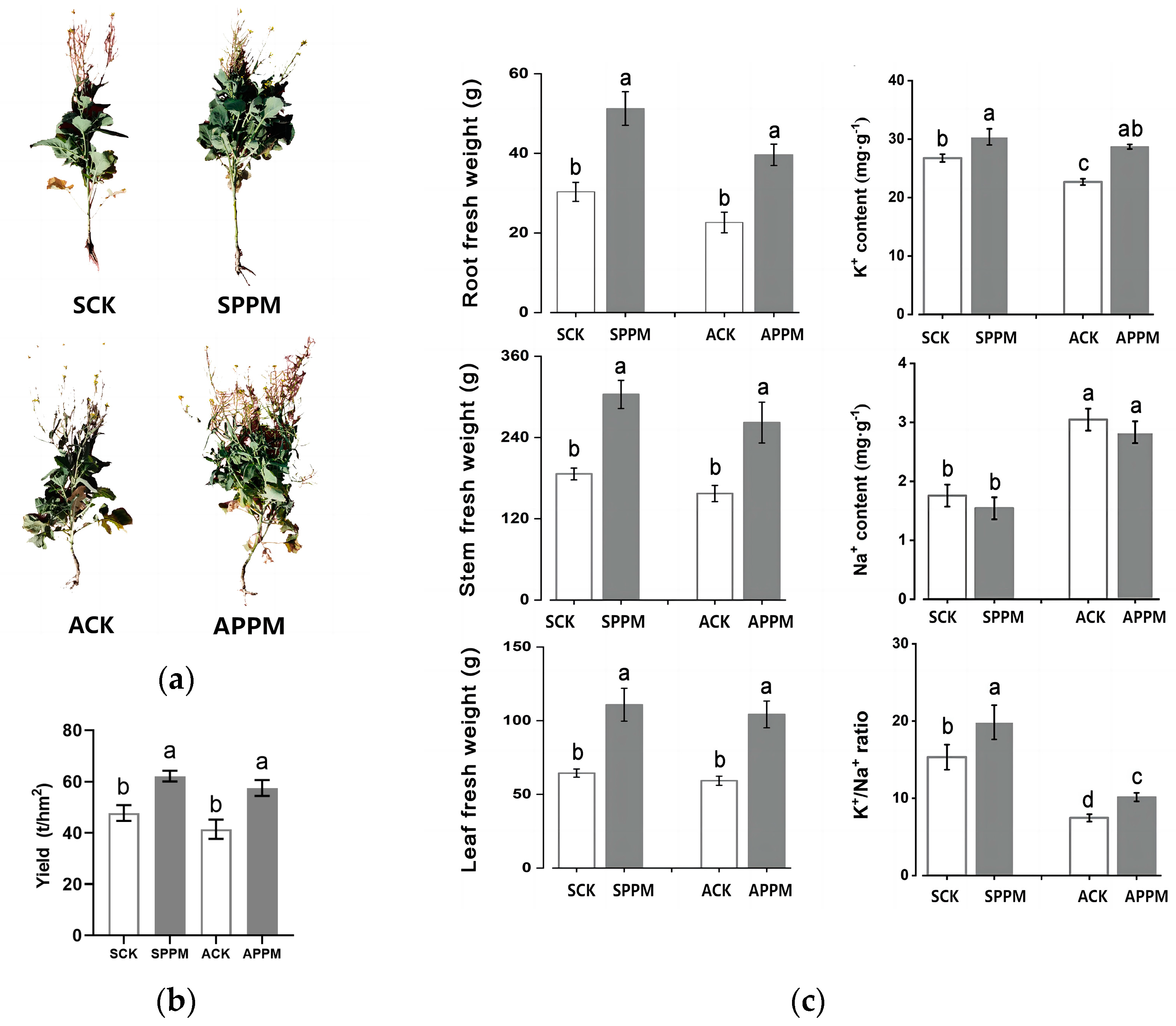
2.1. Effects of Application of PPM on Biomass and Leaf Physiological Parameters of B. napus
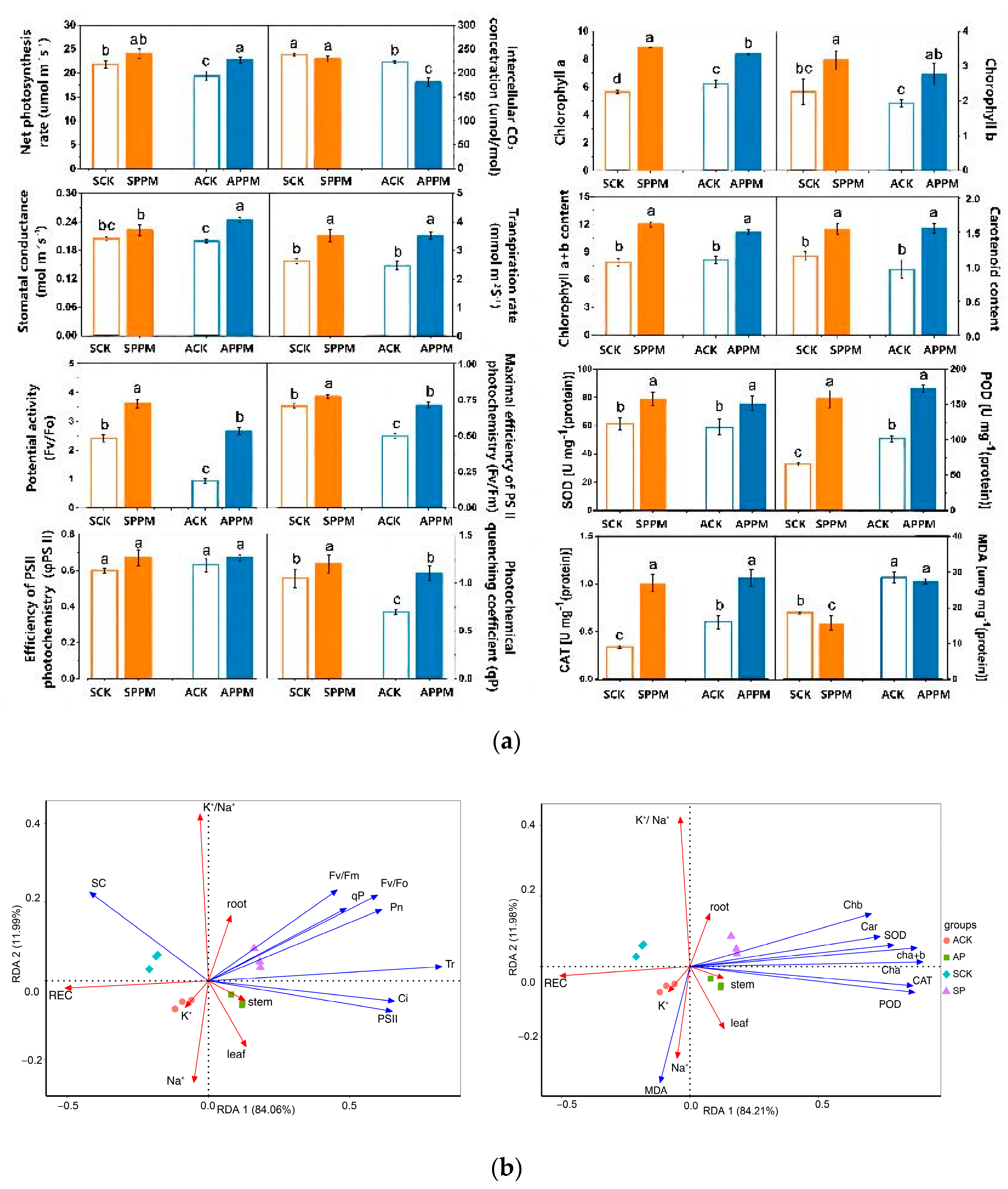
2.2. Effects of PPM Application on Photosynthetic Parameters of B. napus Leaves
2.3. Effects of PPM Application on Antioxidant Enzyme Activity
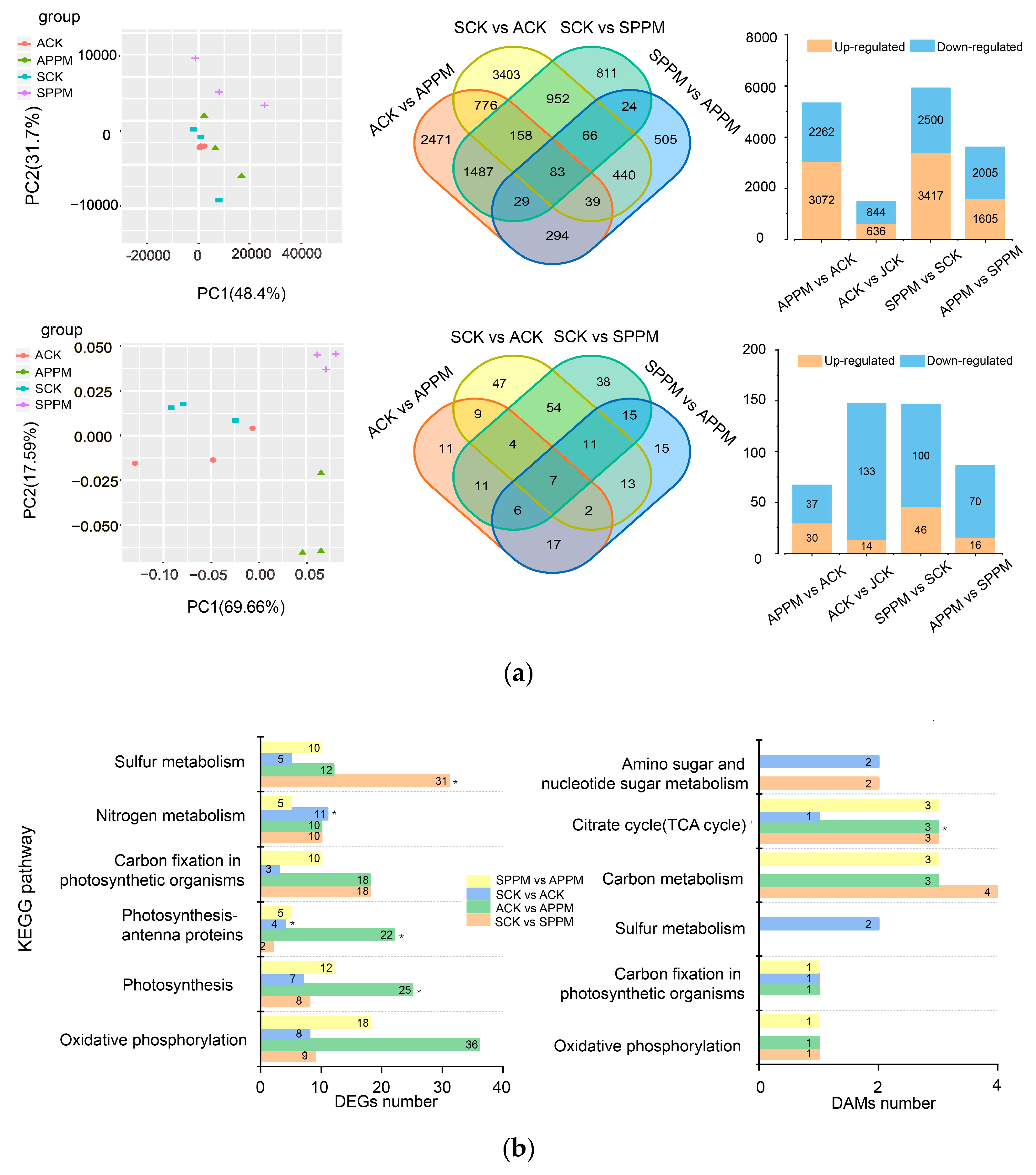
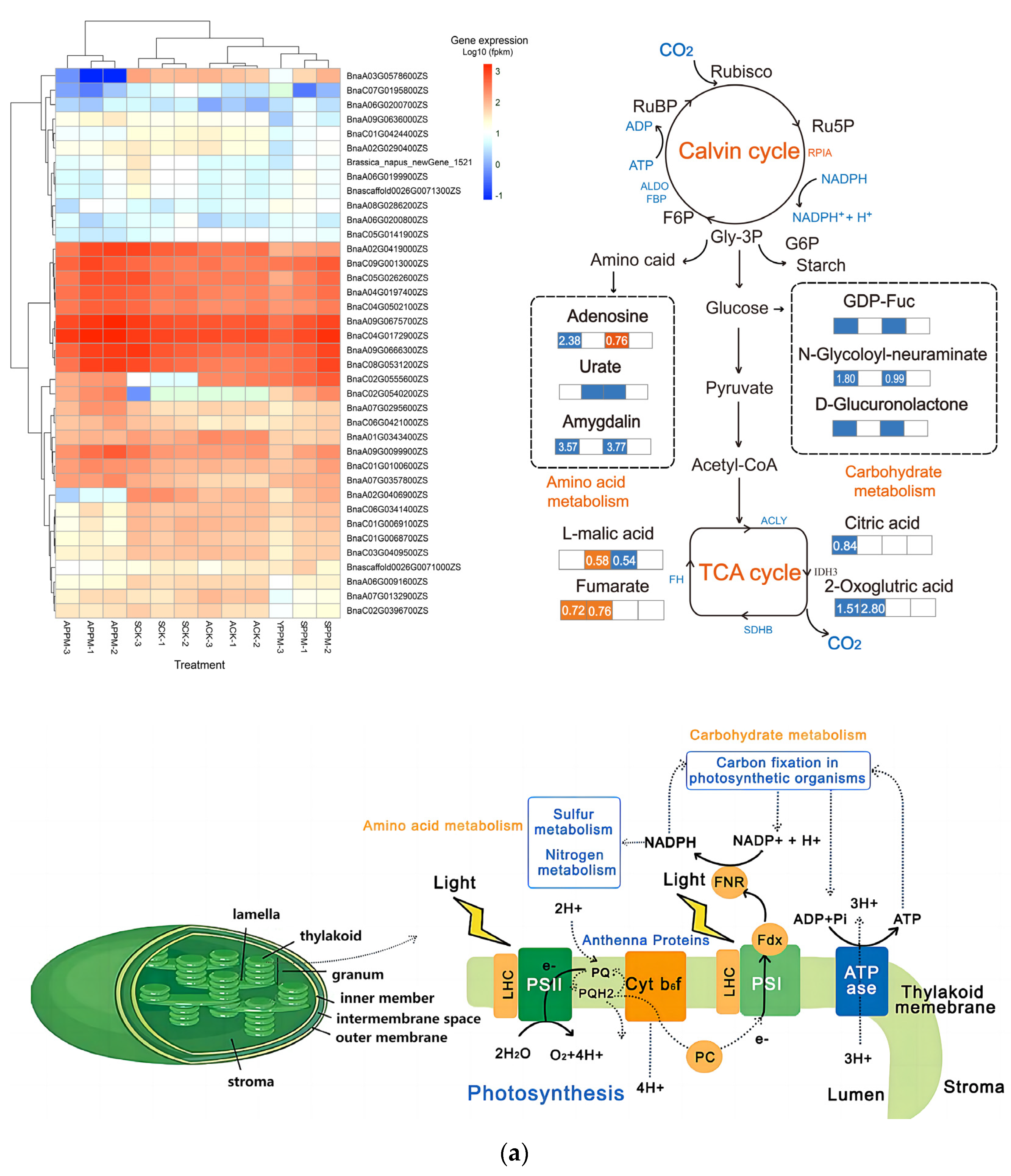
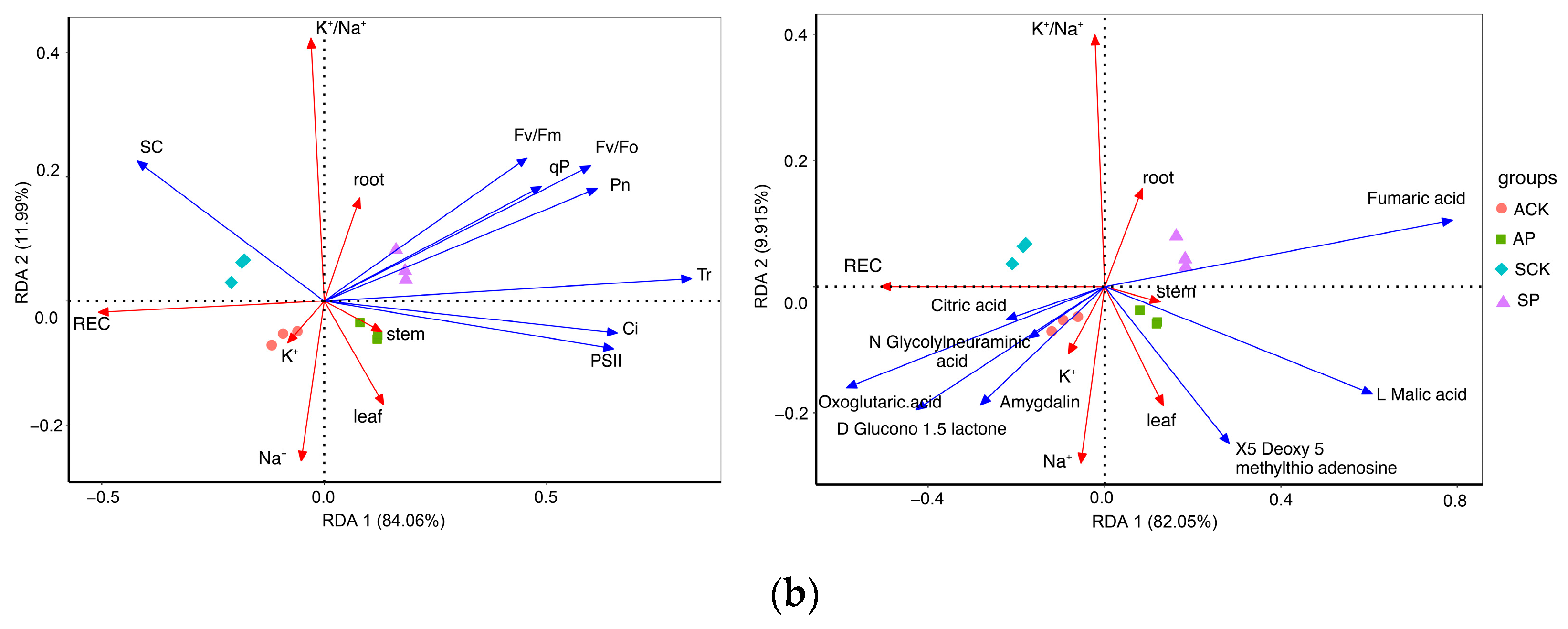
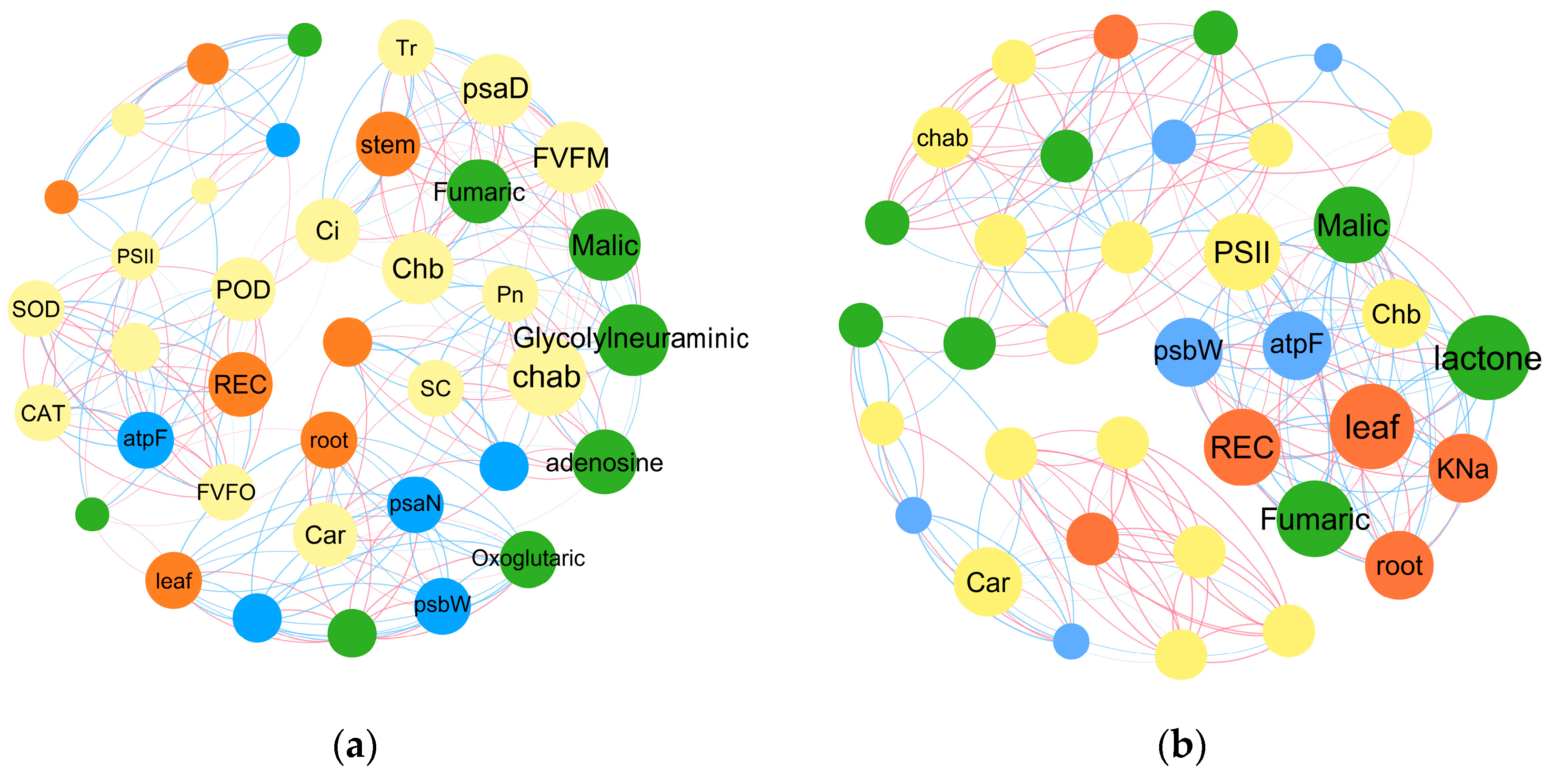
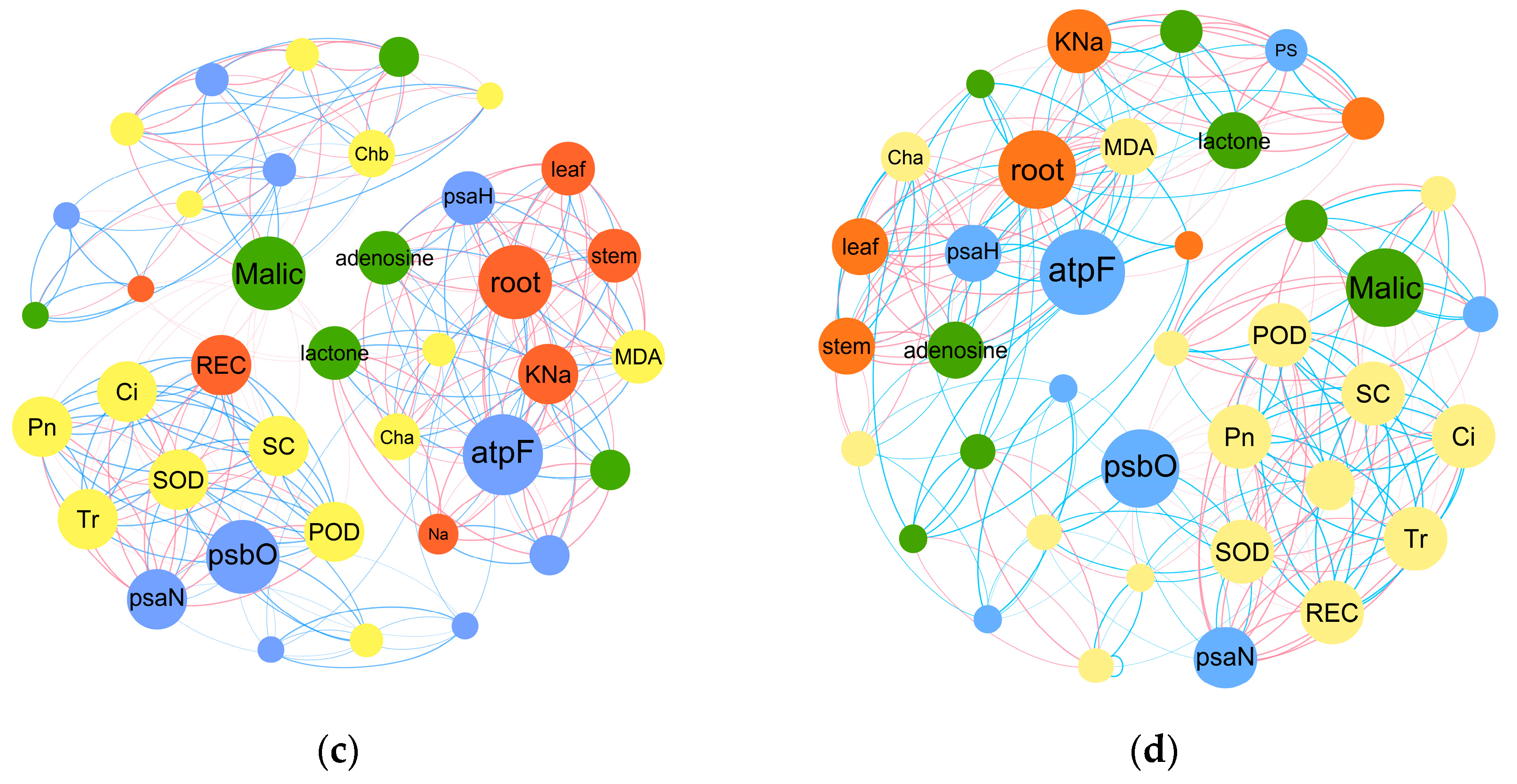
2.4. Transcriptomic and Metabonomic Analysis
2.4.1. Determination of Photosynthetic Parameters, Chlorophyll Fluorescence Parameters and Plant Fresh Weight
2.4.2. PPM’s Impacts on Energy Metabolism and Carbohydrate Metabolism Pathways in B. napus
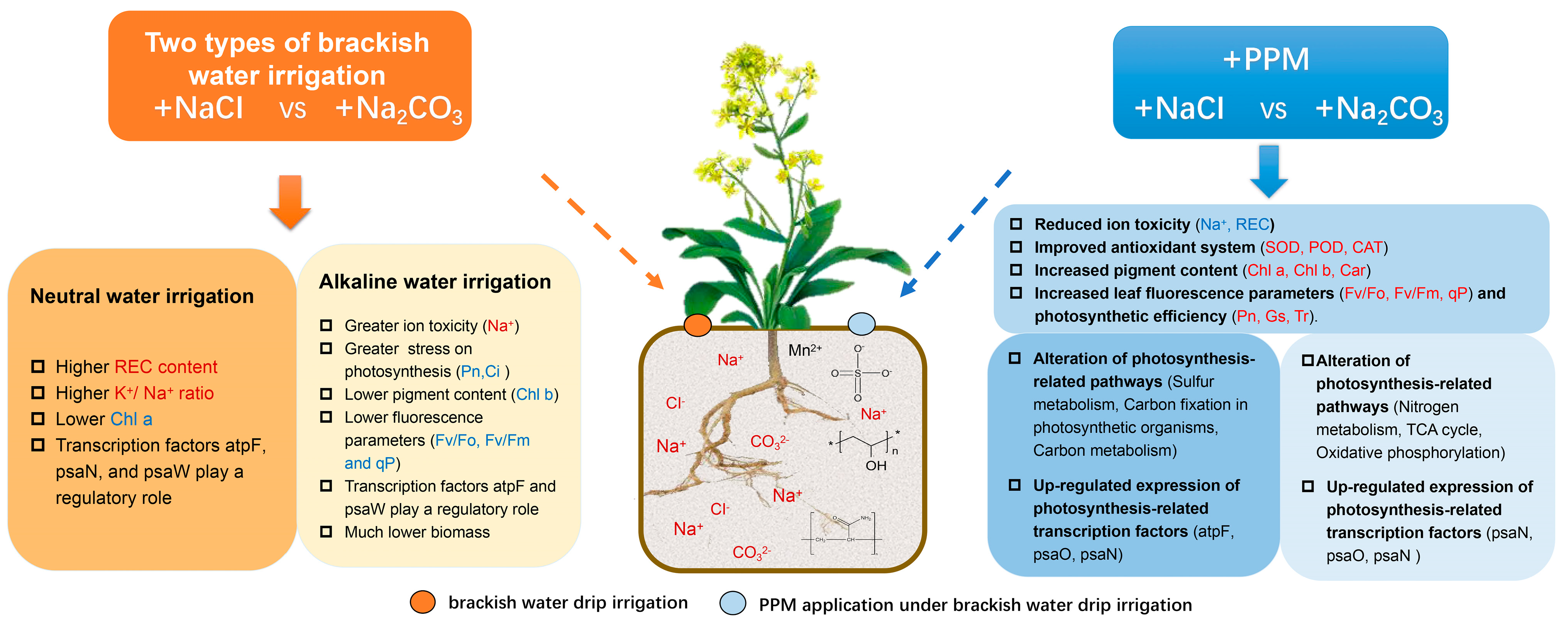
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Materials
4.2. Experimental Design
4.3. Measurement Methods
4.3.1. Determination of Photosynthetic Parameters, Chlorophyll Fluorescence Parameters and Plant Fresh Weight
4.3.2. Determination of Chlorophyll and Carotenoids in Plant Leaves
4.3.3. Determination of Leaf Antioxidant Enzyme Activity and Malondialdehyde (MDA) Content
4.3.4. Determination of Na+, K+ Content and Relative Electrical Conductivity in Leaves
4.3.5. Transcriptomic and Metabolomic Assays
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PPM | a soil amendment composed of polyacrylamide, polyvinyl alcohol, and manganese sulfate |
| Pn | leaf net photosynthetic rate |
| Gs | stomatal conductance |
| Tr | transpiration rate |
| Ci | intracellular CO2 concentration |
| Fv | maximum fluorescence after dark adaptation |
| F0 | minimum fluorescence value after dark adaptation |
| F0′ | minimum fluorescence under light |
| Fm | variable fluorescence |
| Fm′ | maximum fluorescence after light adaptation |
| Fs | steady-state fluorescence after light adaptation |
| Fv/Fm | maximum photochemical efficiency of photosystem II |
| Fv/F0 | potential activity of photosystem II photochemistry |
| qP | photochemical quenching coefficient |
| Chl a | chlorophyll a |
| Chl b | chlorophyll b |
| Car | carotenoids |
| SOD | superoxide dismutase |
| POD | peroxidase |
| CAT | catalase |
| MDA | malondialdehyde |
| REC | relative electrical conductivity |
| DEGs | differentially expressed genes |
| DAMs | differentially accumulated metabolites |
| LC/MS | liquid chromatography-mass spectrometry |
| PCA | principal component analysis |
| RDA | redundancy analysis |
| FPKM | fragments per kilobase of exon per million fragments mapped |
References
- Hussain, M.I.; Farooq, M.; Muscolo, A.; Rehman, A. Crop diversification and saline water irrigation as potential strategies to save freshwater resources and reclamation of marginal soils—A review. Environ. Sci. Pollut. Res. Int. 2020, 27, 28695–28729. [Google Scholar] [CrossRef] [PubMed]
- Leogrande, R.; Vitti, C.; Lopedota, O.; Ventrella, D.; Montemurro, F. Effects of irrigation volume and saline water on maize yield and soil in Southern Italy. Irrig. Drain. 2016, 65, 243–253. [Google Scholar] [CrossRef]
- Ahmad, H.; Hayat, S.; Ali, M.; Liu, T.; Cheng, Z.H. The combination of arbuscular mycorrhizal fungi inoculation (Glomus versiforme) and 28-homobrassinolide spraying intervals improves growth by enhancing photosynthesis, nutrient absorption, and antioxidant system in cucumber (Cucumis sativus L.) under salinity. Evol. Ecol. 2018, 8, 5724–5740. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Zheng, B.S. Photosynthetic response of plants under different abiotic stresses: A review. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.P.; Ma, C.L.; Wang, P.P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Omoto, E.; Taniguchi, M.; Miyake, H. Effects of salinity stress on the structure of bundle sheath and mesophyll chloroplasts in NAD-malic enzyme and PCK type C4 plants. Plant Prod. Sci. 2010, 13, 169–176. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef]
- Chojak-Koźniewska, J.; Kuźniak, E.; Linkiewicz, A.; Sowa, S. Primary carbon metabolism-related changes in cucumber exposed to single and sequential treatments with salt stress and bacterial infection. Plant Physiol. Biochem. 2018, 123, 160–169. [Google Scholar] [CrossRef]
- Li, N.; Zhang, Z.; Gao, S.; Lv, Y.; Chen, Z.J.; Cao, B.L.; Xu, K. Different responses of two Chinese cabbage (Brassica rapa L. ssp. pekinensis) cultivars in photosynthetic characteristics and chloroplast ultrastructure to salt and alkali stress. Planta 2021, 254, 102. [Google Scholar] [CrossRef]
- Zhao, B.; Nan, X.; Xu, H.; Zhang, T.; Ma, F.F. Sulfate sorption on rape (Brassica campestris L.) straw biochar, loess soil and a biochar-soil mixture. J. Environ. Manag. 2017, 201, 309–314. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Wang, J. Seasonality of soil respiration under gypsum and straw amendments in an arid saline-alkali soil. J. Environ. Manag. 2021, 1, 111494. [Google Scholar] [CrossRef]
- Lu, P.; Bainard, L.D.; Ma, B.; Liu, J. Bio-fertilizer and rotten straw amendments alter the rhizosphere bacterial community and increase oat productivity in a saline-alkaline environment. Sci. Rep. 2020, 16, 19896. [Google Scholar] [CrossRef]
- An, M.; Wang, X.; Chang, D.; Wang, S.; Wang, K.Y. Application of compound material alleviates saline and alkaline stress in cotton leaves through regulation of the transcriptome. BMC Plant Biol. 2020, 20, 462. [Google Scholar] [CrossRef]
- Sojka, R.E.; Bjorneberg, D.L.; Entry, J.A.; Lentz, R.D.; Orts, W.J. Polyacrylamides in agriculture and environmental land management. Adv. Agron. 2007, 92, 75–162. [Google Scholar]
- Boya, X.; Dettam, L.R.; Derrick, S.; Taylor, P.; Richard, H.; Andrew, Z.L.; Manish, K. Polyacrylamide degradation and its implications in environmental systems. Npj Clean Water 2018, 1, 17–25. [Google Scholar]
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin/Heidelberg, Germany, 2007; Volume 5, 1. 52. [Google Scholar]
- Kolvenbach, B.A.; Fournier, S.; Mu, Q. Polyvinyl alcohol biodegradation in wastewater treatment plants: A review. Water Res. 2018, 139, 118–128. [Google Scholar]
- Hutton, M.; Eggett, D.; Van, H.S. Manganese for plant growth and development: A review. Agronomy 2020, 2, 279–284. [Google Scholar]
- An, M.; Wei, C.; Wang, K.; Fan, H.; Wang, X.L.; Chang, D.D. Effects of polymer modifiers on the bacterial communities in cadmium-contaminated alkaline soil. Appl. Soil Ecol. 2021, 157, 103777. [Google Scholar] [CrossRef]
- Wang, X.; An, M.; Wang, K.; Cheng, K. Effects of organic polymer compound material on k+ and na+ distribution and physiological characteristics of cotton under saline and alkaline stresses. Front. Plant Sci. 2021, 12, 636536. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Nishiyama, Y.; Miyairi, S.; Yamamoto, H.; Inagaki, N.; Kanesaki, Y.; Murata, N. Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in synechocystis. Plant Physiol. 2002, 130, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.N.; Tanveer, M.; Abbas, A.; Fahad, S.; Baloch, M.S.; Ahmad, M.I.; Saud, S.; Song, Y.H. Targeting salt stress coping mechanisms for stress tolerance in Brassica: A research perspective. Plant Physiol. Biochem. 2021, 158, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mak, M.; Babla, M.; Wang, F.; Chen, G.; Veljanoski, F.; Wang, G.; Shabala, S.; Zhou, M.; Chen, Z.H. Linking stomatal traits and expression of slow anion channel genesHvSLAH1 and HvSLAC1 with grain yield for increasing salinity tolerance in barley. Front. Plant Sci. 2014, 5, 634. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Feng, B.L. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 2021, 11, 615942. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Mishra, A.K.; Kumar, K. Molecular insights into the role of plant transporters in salt stress response. Plant Physiol. 2021, 173, 1481–1494. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Dai, S.Y.; Yong, L.T.; Zou, B.H.; Wang, N.; Dong, Y.Y.; Liu, W.C.; Wang, F.W.; Yang, H.Y.; Li, X.W. A Soybean Sucrose Non-Fermenting Protein Kinase 1 Gene, GmSNF1, Positively Regulates Plant Response to Salt and Salt-Alkali Stress in Transgenic Plants. Int. J. Mol. Sci. 2023, 24, 12482. [Google Scholar] [CrossRef]
- Smith, J.D.; Johnson, R.L. Saline water irrigation: Effects of soil characteristics and irrigation management on crop yield and quality. Agric. Water Manag. 2019, 221, 301–312. [Google Scholar]
- Farooq, M.; Hussain, M.; Usman, M.; Farooq, S.; Alghamdi, S.S.; Siddique, K.H.M. Impact of abiotic stresses on grain composition and quality in food legumes. J. Agric. Food Chem. 2018, 66, 8887–8897. [Google Scholar] [CrossRef]
- Kopriva, S.; Chu, C. Are we ready to improve phosphorus homeostasis in rice? Exp. Bot. 2018, 69, 3515–3522. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Asis, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Wang, X.; Gao, X.; Liang, H.Y.; Liu, B.X.; Huang, X.X. Effects of salt stress on the photosynthetic physiology and mineral ion absorption and distribution in white willow (Salix alba L.). PLoS ONE 2021, 16, e0260086. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; Gomaa, A.M.; Hasanuzzaman, M. Exogenous melatonin enhances the reactive oxygen species metabolism, antioxidant defense-related gene expression, and photosynthetic capacity of Phaseolus vulgaris L. to confer salt stress tolerance. Plant Physiol. 2021, 173, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Chang, D.; Hong, D.; Fan, H.; Wang, K.Y. Metabolic regulation in soil microbial succession and niche differentiation by the polymer amendment under cadmium stress. J. Hazard. Mater. 2021, 416, 126094. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Hassan, M.A.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of salinity and drought on growth, ionic relations, compatible solutes and activation of antioxidant systems in oleander (Nerium oleander L.). PLoS ONE 2017, 12, e0185017. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, F.; Klähn, S.; Hagemann, M. Salt-regulated accumulation of the compatible solutes sucrose and glucosylglycerol in cyanobacteria and its biotechnological potential. Front. Microbiol. 2019, 10, 2139. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, W.; Dai, F. Study on the physiological response differences of Pinus yunnanensis seedlings under different drought conditions. J. Fujian For. Sci. Technol. 2012, 39, 218–222. [Google Scholar]
- Bricker, T.M.; Frankel, L.K. Auxiliary functions of the PsbO, PsbP and PsbQ proteins of higher plant Photosystem II: A critical analysis. Photochem. Photobiol. 2011, 104, 65–178. [Google Scholar] [CrossRef]
- Wang, W.; Pang, J.; Zhang, F.; Sun, L.P.; Yang, L.; Zhao, Y.Z.; Yang, Y.; Wang, Y.J.; Siddique, K.H.M. Integrated transcriptomics and metabolomics analysis to characterize alkali stress responses in canola (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 605–620. [Google Scholar] [CrossRef]
- Zhao, R.; An, L.; Song, D.; Li, M.; Qiao, L.; Liu, N.; Sun, H. Detection of chlorophyll fluorescence parameters of potato leaves based on continuous wavelet transform and spectral analysis. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 259, 119768. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar]
- Martin, W.F.; Bryant, D.A.; Beatty, J.T. A physiological perspective on the origin and evolution of photosynthesis. FEMS Microbiol. Rev. 2018, 42, 205–231. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jiang, Y.; Zhao, H.; Guo, D.; He, L.Z.; Liu, F.G.; Zhou, Q.; Nandwani, D.; Hui, D.F.; Yu, J.Z. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. ssp. Chinensis) in a hydroponic system. PLoS ONE 2018, 13, e0202090. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 99–200. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Song, H.; Li, J.; Chen, D.; Tu, M.Y.; Jiang, G.L.; Yu, G.Q.; Zhou, Z.Q. Comparative transcriptome analysis reveals gene expression differences between two peach cultivars under saline-alkaline stress. Hereditas 2020, 157, 9. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Fan, H.; Yang, L.; Wang, S.; Hong, D.; Cui, W.; Wang, T.; Wei, C.; Sun, Y.; Wang, K.; et al. Multi-Omics Analysis of the Effects of Soil Amendment on Rapeseed (Brassica napus L.) Photosynthesis under Drip Irrigation with Brackish Water. Int. J. Mol. Sci. 2024, 25, 2521. https://doi.org/10.3390/ijms25052521
Li Z, Fan H, Yang L, Wang S, Hong D, Cui W, Wang T, Wei C, Sun Y, Wang K, et al. Multi-Omics Analysis of the Effects of Soil Amendment on Rapeseed (Brassica napus L.) Photosynthesis under Drip Irrigation with Brackish Water. International Journal of Molecular Sciences. 2024; 25(5):2521. https://doi.org/10.3390/ijms25052521
Chicago/Turabian StyleLi, Ziwei, Hua Fan, Le Yang, Shuai Wang, Dashuang Hong, Wenli Cui, Tong Wang, Chunying Wei, Yan Sun, Kaiyong Wang, and et al. 2024. "Multi-Omics Analysis of the Effects of Soil Amendment on Rapeseed (Brassica napus L.) Photosynthesis under Drip Irrigation with Brackish Water" International Journal of Molecular Sciences 25, no. 5: 2521. https://doi.org/10.3390/ijms25052521
APA StyleLi, Z., Fan, H., Yang, L., Wang, S., Hong, D., Cui, W., Wang, T., Wei, C., Sun, Y., Wang, K., & Liu, Y. (2024). Multi-Omics Analysis of the Effects of Soil Amendment on Rapeseed (Brassica napus L.) Photosynthesis under Drip Irrigation with Brackish Water. International Journal of Molecular Sciences, 25(5), 2521. https://doi.org/10.3390/ijms25052521