
Aerobic Exercise Improves Radiation Therapy Efficacy in Non-Small Cell Lung Cancer: Preclinical Study Using a Xenograft Mouse Model

 , ,
, ,
Abstract
:1. Introduction
2. Results
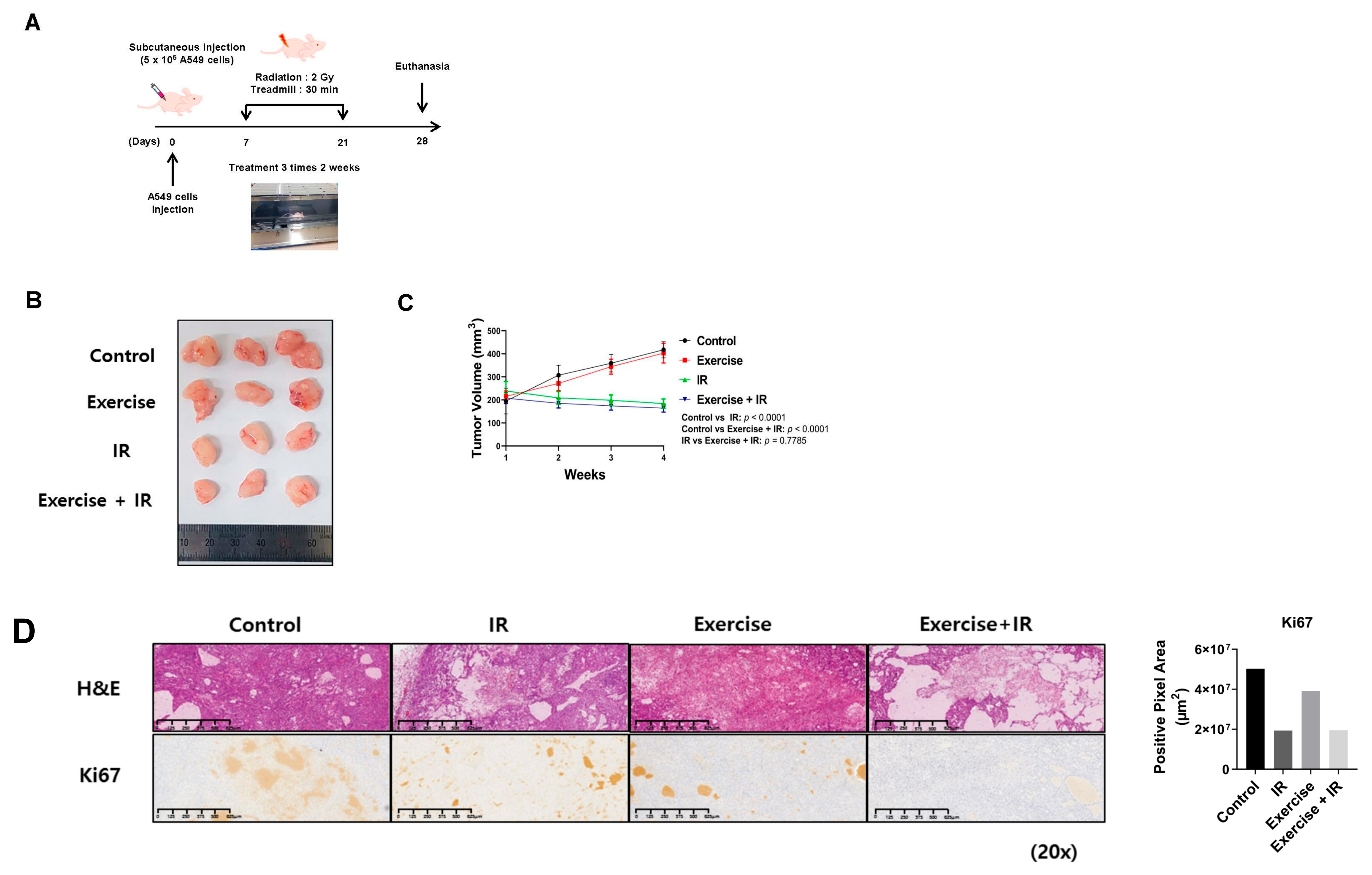
2.1. Combined Aerobic Exercise and Radiation Therapy Reduces NSCLC Cell Growth
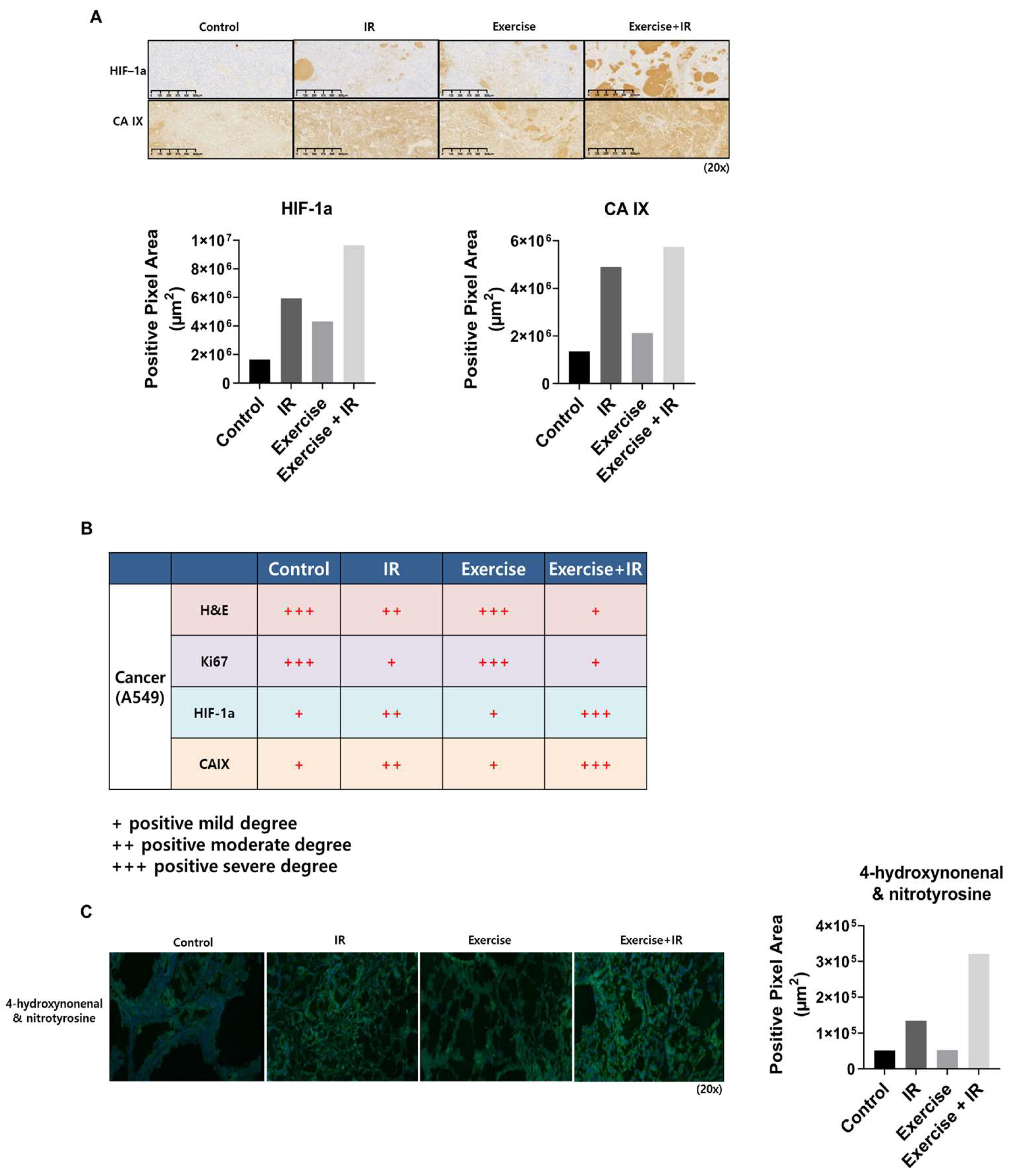
2.2. Aerobic Exercise Combined with Radiation Induces Changes in Tumor Cell Oxygen Concentration
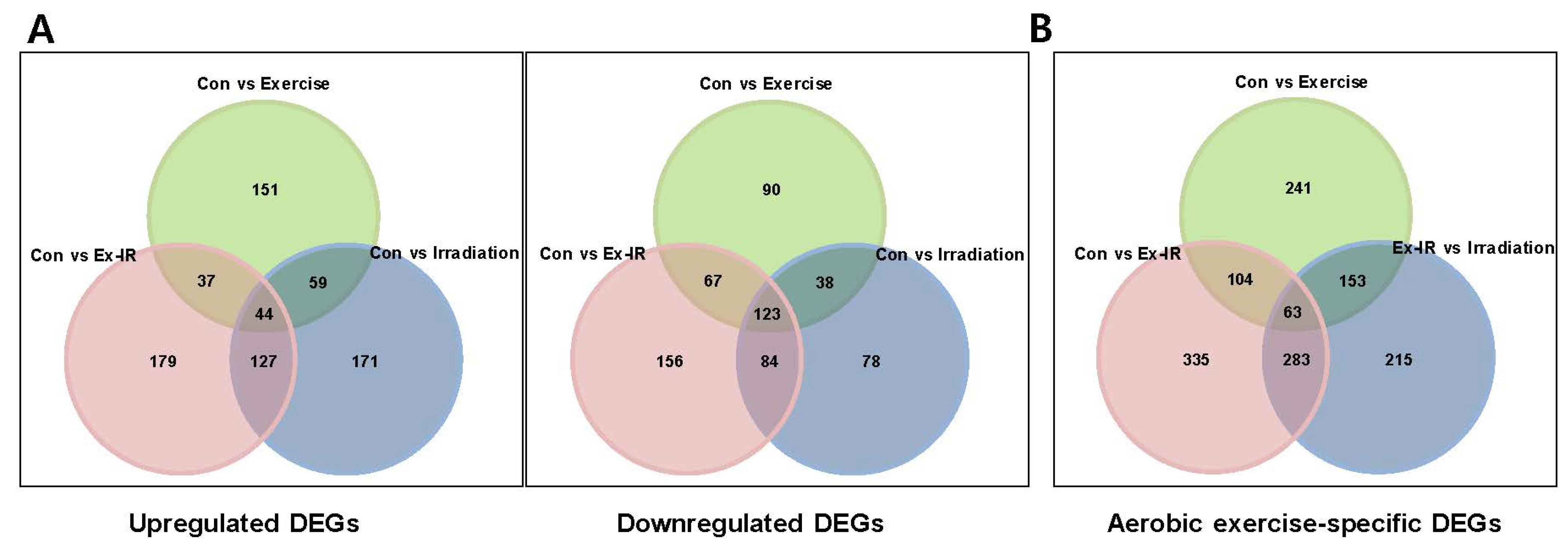
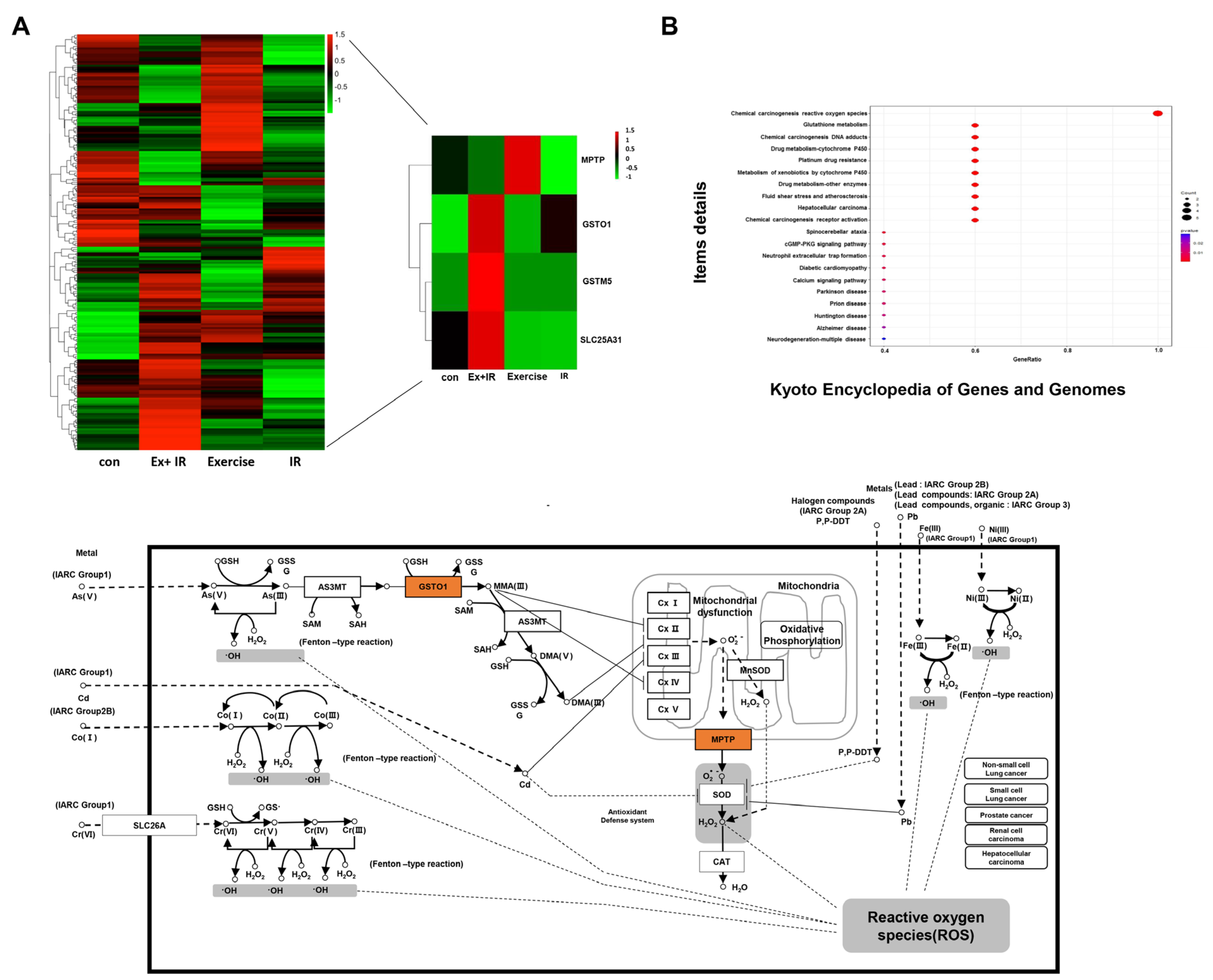
2.3. Aerobic Exercise Combined with Radiation Alters Gene Expression
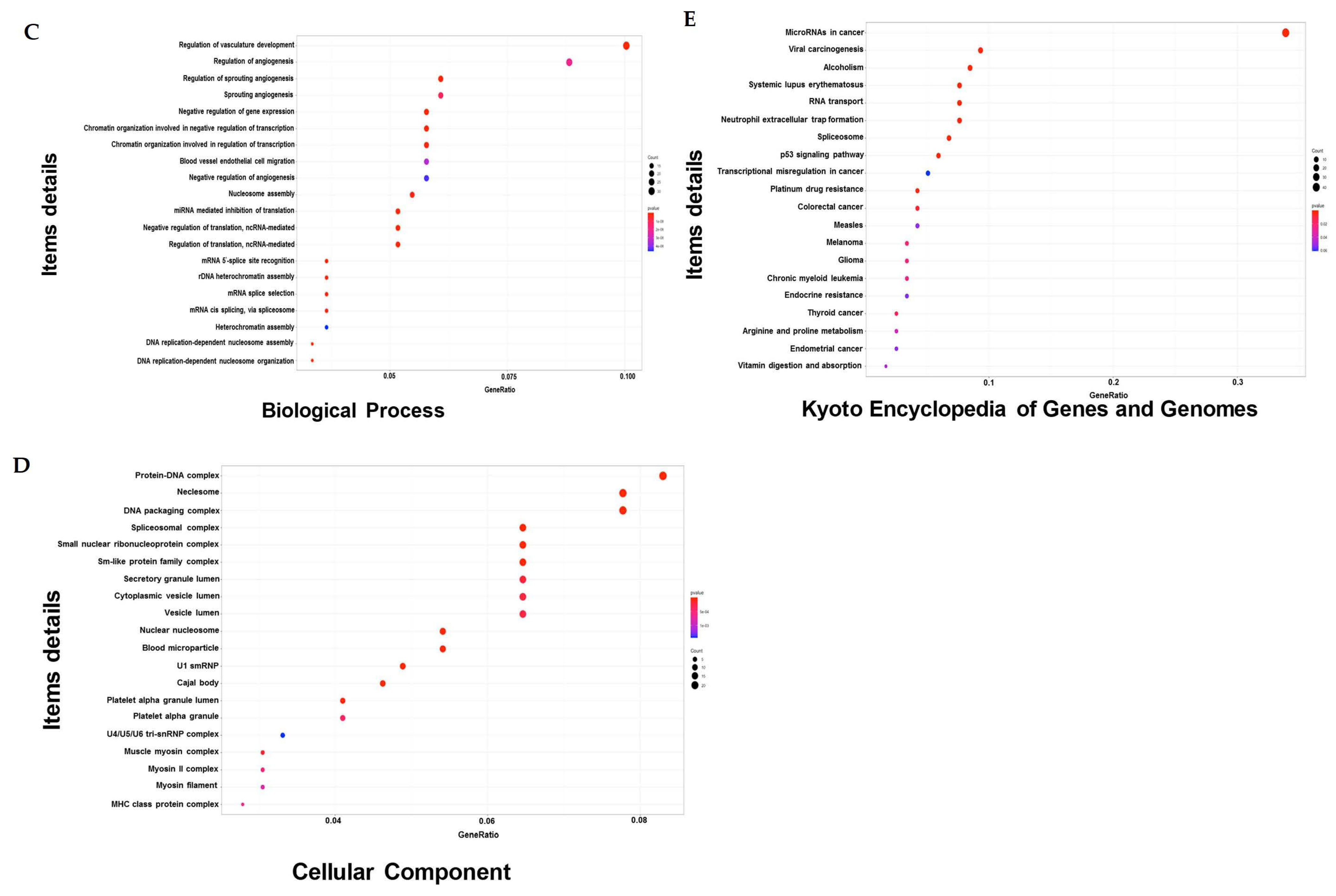
2.4. DEGs That Alter the Oxidation Levels of NSCLC Cells during Combined Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Models
4.3. Histological Analysis
4.4. Animal Radiation and Aerobic Exercise Treatment Procedure
4.5. Ribonucleic Acid (RNA) Isolation, Gene Expression Profiling, and Data Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.O.; Flanigan, J. Novel methods for measuring global cancer burden: Implications for global cancer control. JAMA Oncol. 2015, 1, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global epidemiology of lung cancer. Ann. Glob. Health 2019, 85, 8. [Google Scholar] [CrossRef] [PubMed]
- Thun, M.J.; Carter, B.D.; Feskanich, D.; Freedman, N.D.; Prentice, R.; Lopez, A.D.; Hartge, P.; Gapstur, S.M. 50-year trends in smoking-related mortality in the United States. N. Engl. J. Med. 2013, 368, 351–364. [Google Scholar] [CrossRef]
- Thun, M.J.; Lopez, A.D.; Hartge, P. Smoking-related mortality in the United States: The authors reply. N. Engl. J. Med. 2013, 368, 1753. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Introduction to the 2015 World Health Organization classification of tumors of the lung, pleura, thymus, and heart. J. Thorac. Oncol. 2015, 10, 1240–1242. [Google Scholar] [CrossRef]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef]
- Chi, A.; Chen, H.; Wen, S.; Yan, H.; Liao, Z. Comparison of particle beam therapy and stereotactic body radiotherapy for early stage non-small cell lung cancer: A systematic review and hypothesis-generating meta-analysis. Radiother. Oncol. 2017, 123, 346–354. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, Z.; Zhang, C.; Wang, Y.; Zhang, H.; Gan, Z.; Guo, Z.; Wang, X. Mitochondrion-targeted platinum complexes suppressing lung cancer through multiple pathways involving energy metabolism. Chem. Sci. 2019, 10, 3089–3095. [Google Scholar] [CrossRef]
- Olusanya, T.O.; Haj Ahmad, R.R.; Ibegbu, D.M.; Smith, J.R.; Elkordy, A.A. Liposomal drug delivery systems and anticancer drugs. Molecules 2018, 23, 907. [Google Scholar] [CrossRef]
- Taratula, O.; Garbuzenko, O.B.; Chen, A.M.; Minko, T. Innovative strategy for treatment of lung cancer: Targeted nanotechnology-based inhalation co-delivery of anticancer drugs and siRNA. J. Drug Target. 2011, 19, 900–914. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Paganetti, H.; Grassberger, C. Prediction of treatment response for combined chemo-and radiation therapy for non-small cell lung cancer patients using a bio-mathematical model. Sci. Rep. 2017, 7, 13542. [Google Scholar] [CrossRef] [PubMed]
- Le Chevalier, T.; Arriagada, R.; Quoix, E.; Ruffle, P.; Martin, M.; Tarayre, M.; Marie-José, L.-T.; Douillard, J.-Y.; Laplanche, A. Radiotherapy alone versus combined chemotherapy and radiotherapy in nonresectable non-small-cell lung cancer: First analysis of a randomized trial in 353 patients. J. Natl. Cancer Inst. 1991, 83, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Segawa, Y.; Kiura, K.; Takigawa, N.; Kamei, H.; Harita, S.; Hiraki, S.; Watanabe, Y.; Sugimoto, K.; Shibayama, T.; Yonei, T. Phase III trial comparing docetaxel and cisplatin combination chemotherapy with mitomycin, vindesine, and cisplatin combination chemotherapy with concurrent thoracic radiotherapy in locally advanced non–small-cell lung cancer: OLCSG 0007. J. Clin. Oncol. 2010, 28, 3299–3306. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.M.; Desouky, S.; Marzouk, M.; Sayed, A.A. Origanum majorana attenuates nephrotoxicity of cisplatin anticancer drug through ameliorating oxidative stress. Nutrients 2016, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Wang, F.-X.; Jia, K.-K.; Kong, L.-D. Natural product interventions for chemotherapy and radiotherapy-induced side effects. Front. Pharmacol. 2018, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.-J.; Kim, J.; Jeong, H.; Bok, S.; Kim, Y.-E.; Ahn, G.-O. Tumor hypoxia and reoxygenation: The yin and yang for radiotherapy. Radiat. Oncol. J. 2016, 34, 239. [Google Scholar] [CrossRef]
- Gray, L.H.; Conger, A.D.; Ebert, M.; Hornsey, S.; Scott, O. The concentration of oxygen dissolved in tissues at the time of irradiation as a factor in radiotherapy. Br. J. Radiol. 1953, 26, 638–648. [Google Scholar] [CrossRef]
- Gray, L. Radiobiologic basis of oxygen as a modifying factor in radiation therapy. Am. J. Roentgenol. Radium Ther. Nucl. Med. 1961, 84. [Google Scholar]
- Thomlinson, R.H.; Gray, L.H. The histological structure of some human lung cancers and the possible implications for radiotherapy. Br. J. Cancer 1955, 9, 539. [Google Scholar] [CrossRef]
- Overgaard, J. Hypoxic radiosensitization: Adored and ignored. J. Clin. Oncol. 2007, 25, 4066–4074. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, S.; Dobrucki, I.T.; Kim, E.Y.; Marrison, S.T.; Vu, V.T. Hypoxia and radiation therapy: Past history, ongoing research, and future promise. Curr. Mol. Med. 2009, 9, 442–458. [Google Scholar] [CrossRef] [PubMed]
- Suit, H.D. Radiation biology: A basis for radiotherapy. In Textbook of Radiotherapy; Lea and Febiger Philadelphia: Philadelphia, PA, USA, 1973; pp. 75–121. [Google Scholar]
- Kuperman, V.; Lubich, L. Effect of reoxygenation on hypofractionated radiotherapy of prostate cancer. Med. Phys. 2020, 47, 5383–5391. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Betzer, O.; Tao, D.; Sadan, T.; Popovtzer, R.; Liu, Z. Oxygen nanoshuttles for tumor oxygenation and enhanced cancer treatment. CCS Chem. 2019, 1, 239–250. [Google Scholar]
- Feng, L.; Dong, Z.; Liang, C.; Chen, M.; Tao, D.; Cheng, L.; Yang, K.; Liu, Z. Iridium nanocrystals encapsulated liposomes as near-infrared light controllable nanozymes for enhanced cancer radiotherapy. Biomaterials 2018, 181, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Song, X.; Liang, C.; Yi, X.; Song, G.; Chao, Y.; Yang, Y.; Yang, K.; Feng, L.; Liu, Z. Catalase-loaded cisplatin-prodrug-constructed liposomes to overcome tumor hypoxia for enhanced chemo-radiotherapy of cancer. Biomaterials 2017, 138, 13–21. [Google Scholar] [CrossRef]
- Jones, L.W.; Viglianti, B.L.; Tashjian, J.A.; Kothadia, S.M.; Keir, S.T.; Freedland, S.J.; Potter, M.Q.; Moon, E.J.; Schroeder, T.; Herndon, J.E., 2nd; et al. Effect of aerobic exercise on tumor physiology in an animal model of human breast cancer. J. Appl. Physiol. 2010, 108, 343–348. [Google Scholar] [CrossRef]
- Jee, H.; Park, E.; Hur, K.; Kang, M.; Kim, Y. High-Intensity Aerobic Exercise Suppresses Cancer Growth by Regulating Skeletal Muscle-Derived Oncogenes and Tumor Suppressors. Front. Mol. Biosci. 2022, 9, 818470. [Google Scholar] [CrossRef]
- Jia, N.; Zhou, Y.; Dong, X.; Ding, M. The antitumor mechanisms of aerobic exercise: A review of recent preclinical studies. Cancer Med. 2021, 10, 6365–6373. [Google Scholar] [CrossRef]
- Eroglu, H.; Okyaz, B.; Türkçapar, Ü. The Effect of Acute Aerobic Exercise on Arterial Blood Oxygen Saturation of Athletes. J. Educ. Train. Stud. 2018, 6, 74–79. [Google Scholar] [CrossRef]
- Coli, C.; Sari, G.M.; Rejeki, P.S. Acute Moderate Intensity Exercise Decreases Oxygen Saturation in Obese Women. STRADA J. Ilm. Kesehat. 2020, 9, 310–315. [Google Scholar] [CrossRef]
- Neto, G.R.; Sousa, M.S.; Costa e Silva, G.V.; Gil, A.L.; Salles, B.F.; Novaes, J.S. Acute resistance exercise with blood flow restriction effects on heart rate, double product, oxygen saturation and perceived exertion. Clin. Physiol. Funct. Imaging 2016, 36, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.; Choy, H.; Bradley, J.; Rosenzweig, K.E.; Bogart, J.; Curran, W.J., Jr.; Gore, E.; Langer, C.; Louie, A.V.; Lutz, S.; et al. Definitive radiation therapy in locally advanced non-small cell lung cancer: Executive summary of an American Society for Radiation Oncology (ASTRO) evidence-based clinical practice guideline. Pract. Radiat. Oncol. 2015, 5, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Nishimura, Y.; Hanaoka, K.; Nakamatsu, K.; Doi, H.; Uehara, T.; Komanishi, M.; Ishii, K.; Kaida, H.; Hosono, M. Visualization of tumor hypoxia and re-oxygenation after stereotactic body radiation therapy in early peripheral lung cancer: A prospective study. Radiother. Oncol. 2023, 180, 109491. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Feng, L.; Liu, J.; Zhu, W.; Dong, Z.; Wu, Y.; Liu, Z. Intelligent albumin–MnO2 nanoparticles as pH-/H2O2-responsive dissociable nanocarriers to modulate tumor hypoxia for effective combination therapy. Adv. Mater. 2016, 28, 7129–7136. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Feng, L.; Liang, C.; Yang, K.; Liu, Z. Ultrasound triggered tumor oxygenation with oxygen-shuttle nanoperfluorocarbon to overcome hypoxia-associated resistance in cancer therapies. Nano Lett. 2016, 16, 6145–6153. [Google Scholar] [CrossRef]
- Sprod, L.K.; Palesh, O.G.; Janelsins, M.C.; Peppone, L.J.; Heckler, C.E.; Adams, M.J.; Morrow, G.R.; Mustian, K.M. Exercise, sleep quality, and mediators of sleep in breast and prostate cancer patients receiving radiation therapy. Community Oncol. 2010, 7, 463. [Google Scholar] [CrossRef]
- Ezema, C.I.; Omeh, E.; Onyeso, O.K.K.; Anyachukwu, C.C.; Nwankwo, M.J.; Amaeze, A.; Ugwulebor, J.U.; Nna, E.O.; Ohotu, E.O.; Ugwuanyi, I. The effect of an aerobic exercise programme on blood glucose level, cardiovascular parameters, peripheral oxygen saturation, and body mass index among Southern Nigerians with type 2 diabetes mellitus, undergoing concurrent sulfonylurea and metformin treatment. Malays. J. Med. Sci. 2019, 26, 88. [Google Scholar]
- Borghi-Silva, A.; Arena, R.; Castello, V.; Simoes, R.P.; Martins, L.E.B.; Catai, A.M.; Costa, D. Aerobic exercise training improves autonomic nervous control in patients with COPD. Respir. Med. 2009, 103, 1503–1510. [Google Scholar] [CrossRef]
- Kapur, G.; Windsor, P.; Mc Cowan, C. The effect of aerobic exercise on treatment-related acute toxicity in men receiving radical external beam radiotherapy for localised prostate cancer. Eur. J. Cancer Care 2010, 19, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, A.; Tarnawski, A.S. Critical role of hypoxia sensor-HIF-1α in VEGF gene activation. Implications for angiogenesis and tissue injury healing. Curr. Med. Chem. 2012, 19, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hagen, T. Oxygen versus reactive oxygen in the regulation of HIF-1: The balance tips. Biochem. Res. Int. 2012, 2012, 436981. [Google Scholar] [CrossRef] [PubMed]
- Chia, S.K.; Wykoff, C.C.; Watson, P.H.; Han, C.; Leek, R.D.; Pastorek, J.; Gatter, K.C.; Ratcliffe, P.; Harris, A.L. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J. Clin. Oncol. 2001, 19, 3660–3668. [Google Scholar] [CrossRef] [PubMed]
- Tafreshi, N.K.; Lloyd, M.C.; Proemsey, J.B.; Bui, M.M.; Kim, J.; Gillies, R.J.; Morse, D.L. Evaluation of CAIX and CAXII expression in breast cancer at varied O 2 levels: CAIX is the superior surrogate imaging biomarker of tumor hypoxia. Mol. Imaging Biol. 2016, 18, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Ilie, M.; Hofman, V.; Zangari, J.; Chiche, J.; Mouroux, J.; Mazure, N.M.; Pouysségur, J.; Brest, P.; Hofman, P. Response of CAIX and CAXII to in vitro re-oxygenation and clinical significance of the combined expression in NSCLC patients. Lung Cancer 2013, 82, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Matés, J.M.; Sánchez-Jiménez, F.M. Role of reactive oxygen species in apoptosis: Implications for cancer therapy. Int. J. Biochem. Cell Biol. 2000, 32, 157–170. [Google Scholar] [CrossRef]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef]
- Kuschel, A.; Simon, P.; Tug, S. Functional regulation of HIF-1α under normoxia—Is there more than post-translational regulation? J. Cell. Physiol. 2012, 227, 514–524. [Google Scholar] [CrossRef]
- Manea, A. NADPH oxidase-derived reactive oxygen species: Involvement in vascular physiology and pathology. Cell Tissue Res. 2010, 342, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, T.; Pissas, G.; Filippidis, G.; Liakopoulos, V.; Stefanidis, I. Reoxygenation induces reactive oxygen species production and ferroptosis in renal tubular epithelial cells by activating aryl hydrocarbon receptor. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Weng, W.; Xu, M.; Wang, Q.; Tan, C.; Sun, H.; Wang, L.; Huang, D.; Du, X.; Sheng, W. miR-106b-5p inhibits the invasion and metastasis of colorectal cancer by targeting CTSA. OncoTargets Ther. 2018, 3835–3845. [Google Scholar] [CrossRef]
- Engel, L.S.; Taioli, E.; Pfeiffer, R.; Garcia-Closas, M.; Marcus, P.M.; Lan, Q.; Boffetta, P.; Vineis, P.; Autrup, H.; Bell, D.A. Pooled analysis and meta-analysis of glutathione S-transferase M1 and bladder cancer: A HuGE review. Am. J. Epidemiol. 2002, 156, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.O.; Kieran, N.E.; Howell, K.; Burne, M.J.; Varadarajan, R.; Dhakshinamoorthy, S.; Porter, A.G.; O’Farrelly, C.; Rabb, H.; Taylor, C.T. Reoxygenation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia-reperfusion injury. FASEB J. 2006, 20, 2624–2626. [Google Scholar] [CrossRef] [PubMed]
- Jou, Y.-C.; Wang, S.-C.; Dia, Y.-C.; Wang, S.-T.; Yu, M.-H.; Yang, H.-Y.; Chen, L.-C.; Shen, C.-H.; Liu, Y.-W. Anti-cancer effects and tumor marker role of glutathione S-transferase Mu 5 in human bladder cancer. Int. J. Mol. Sci. 2021, 22, 3056. [Google Scholar] [CrossRef] [PubMed]
- Pankratz, V.S.; Sun, Z.; Aakre, J.; Li, Y.; Johnson, C.; Garces, Y.I.; Aubry, M.C.; Molina, J.R.; Wigle, D.A.; Yang, P. Systematic evaluation of genetic variants in three biological pathways on patient survival in low-stage non-small cell lung cancer. J. Thorac. Oncol. 2011, 6, 1488–1495. [Google Scholar] [CrossRef]
- Halestrap, A.P. What is the mitochondrial permeability transition pore? J. Mol. Cell. Cardiol. 2009, 46, 821–831. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef]
- Husainy, M.A. The Role of Protein Kinase B and Mitochondrial Permeability Transition Pore in Ischaemic Preconditioning of Myocardium. Ph.D. Thesis, University of Leicester, Leicester, UK, 2012. [Google Scholar]
- Waseem, M.; Wang, B.-D. Promising Strategy of mPTP Modulation in Cancer Therapy: An Emerging Progress and Future Insight. Int. J. Mol. Sci. 2023, 24, 5564. [Google Scholar] [CrossRef]
- Battaglia, A.M.; Chirillo, R.; Aversa, I.; Sacco, A.; Costanzo, F.; Biamonte, F. Ferroptosis and cancer: Mitochondria meet the “iron maiden” cell death. Cells 2020, 9, 1505. [Google Scholar] [CrossRef] [PubMed]
- Bround, M.J.; Bers, D.M.; Molkentin, J.D. A 20/20 view of ANT function in mitochondrial biology and necrotic cell death. J. Mol. Cell. Cardiol. 2020, 144, A3–A13. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M. Energetic aspects of transport of ADP and ATP through the mitochondrial membrane. Energy Transform. Biol. Syst. 2008, 105–124. [Google Scholar]
- Dolce, V.; Scarcia, P.; Iacopetta, D.; Palmieri, F. A fourth ADP/ATP carrier isoform in man: Identification, bacterial expression, functional characterization and tissue distribution. FEBS Lett. 2005, 579, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Trisolini, L.; Laera, L.; Favia, M.; Muscella, A.; Castegna, A.; Pesce, V.; Guerra, L.; De Grassi, A.; Volpicella, M.; Pierri, C.L. Differential expression of adp/atp carriers as a biomarker of metabolic remodeling and survival in kidney cancers. Biomolecules 2020, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Laliberte, R.E.; Perregaux, D.G.; Hoth, L.R.; Rosner, P.J.; Jordan, C.K.; Peese, K.M.; Eggler, J.F.; Dombroski, M.A.; Geoghegan, K.F.; Gabel, C.A. Glutathione S-Transferase Omega 1-1 Is a Target of Cytokine Release Inhibitory Drugs and May Be Responsible for Their Effect on Interleukin-1β Posttranslational Processing. J. Biol. Chem. 2003, 278, 16567–16578. [Google Scholar] [CrossRef] [PubMed]
- Menon, D.; Coll, R.; Board, P.G. Glutathione transferase omega 1 is required for the lipopolysaccharide-stimulated induction of NADPH oxidase 1 and the production of reactive oxygen species in macrophages. Free Radic. Biol. Med. 2014, 73, 318–327. [Google Scholar] [CrossRef]
- Xu, Y.; Bankhead, A., III; Tian, X.; Tang, J.; Ljungman, M.; Neamati, N. Deletion of glutathione S-transferase omega 1 activates type I interferon genes and downregulates tissue factor. Cancer Res. 2020, 80, 3692–3705. [Google Scholar] [CrossRef]
- Wu, X.; Gao, S.; Lian, Y. Effects of continuous aerobic exercise on lung function and quality of life with asthma: A systematic review and meta-analysis. J. Thorac. Dis. 2020, 12, 4781. [Google Scholar] [CrossRef]
- Armstrong, M.; Vogiatzis, I. Personalized exercise training in chronic lung diseases. Respirology 2019, 24, 854–862. [Google Scholar] [CrossRef]
- Ashcraft, K.A.; Peace, R.M.; Betof, A.S.; Dewhirst, M.W.; Jones, L.W. Efficacy and mechanisms of aerobic exercise on cancer initiation, progression, and metastasis: A critical systematic review of in vivo preclinical data. Cancer Res. 2016, 76, 4032–4050. [Google Scholar] [CrossRef]
- Cho, J.; Shin, M.-K.; Kim, D.; Lee, I.; Kim, S.; Kang, H. Treadmill running reverses cognitive declines due to Alzheimer disease. Med. Sci. Sports Exerc. 2015, 47, 1814–1824. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Fold Change (log2) | Up/DownRegulated | |
|---|---|---|---|
| 100422859 | Mir3136 | −14.56455009 | Downregulated |
| 100189415 | Trf-Gaa1-3 | −13.88255615 | Downregulated |
| 113218489 | miR10398 | 13.86087595 | Upregulated |
| 406910 | miR125a | −13.83876398 | Downregulated |
| 407011 | miR23b | −13.66512849 | Downregulated |
| 102466194 | miR6773 | −13.64055237 | Downregulated |
| 113218510 | miR10523 | −13.60208128 | Downregulated |
| 102466741 | miR6813 | −13.45770358 | Downregulated |
| 692195 | Snord75 | −13.35817707 | Downregulated |
| 26809 | Snord42a | −13.31087593 | Downregulated |
| 442907 | miR339 | −13.29544742 | Downregulated |
| 100616294 | miR4740 | −13.28779461 | Downregulated |
| 100033413 | Snord116-1 | −13.25012801 | Downregulated |
| 100189284 | Trg-Gcc1-5 | −13.11534573 | Downregulated |
| 100500904 | miR3942 | −13.08187484 | Downregulated |
| 100188995 | Trk-Ctt6-1 | −13.07527287 | Downregulated |
| 102465842 | miR7856 | −13.04270882 | Downregulated |
| 406900 | miR106b | 13.01436327 | Upregulated |
| 102465429 | miR6511b1 | 12.96253071 | Upregulated |
| 102466743 | miR6822 | −12.91934167 | Downregulated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, S.; Jeon, J.; Park, G.; Do, H.-K.; Kang, J.; Ahn, K.J.; Ma, S.Y.; Choi, Y.M.; Kim, D.; Youn, B.; et al. Aerobic Exercise Improves Radiation Therapy Efficacy in Non-Small Cell Lung Cancer: Preclinical Study Using a Xenograft Mouse Model. Int. J. Mol. Sci. 2024, 25, 2757. https://doi.org/10.3390/ijms25052757
Jo S, Jeon J, Park G, Do H-K, Kang J, Ahn KJ, Ma SY, Choi YM, Kim D, Youn B, et al. Aerobic Exercise Improves Radiation Therapy Efficacy in Non-Small Cell Lung Cancer: Preclinical Study Using a Xenograft Mouse Model. International Journal of Molecular Sciences. 2024; 25(5):2757. https://doi.org/10.3390/ijms25052757
Chicago/Turabian StyleJo, Sunmi, Jaewan Jeon, Geumju Park, Hwan-Kwon Do, JiHoon Kang, Ki Jung Ahn, Sun Young Ma, Young Min Choi, Donghyun Kim, BuHyun Youn, and et al. 2024. "Aerobic Exercise Improves Radiation Therapy Efficacy in Non-Small Cell Lung Cancer: Preclinical Study Using a Xenograft Mouse Model" International Journal of Molecular Sciences 25, no. 5: 2757. https://doi.org/10.3390/ijms25052757
APA StyleJo, S., Jeon, J., Park, G., Do, H. -K., Kang, J., Ahn, K. J., Ma, S. Y., Choi, Y. M., Kim, D., Youn, B., & Ki, Y. (2024). Aerobic Exercise Improves Radiation Therapy Efficacy in Non-Small Cell Lung Cancer: Preclinical Study Using a Xenograft Mouse Model. International Journal of Molecular Sciences, 25(5), 2757. https://doi.org/10.3390/ijms25052757