
Aging and Adiposity—Focus on Biological Females at Midlife and Beyond


Abstract
:1. Introduction
2. Adiposity
3. Adipose Tissue—A Heterogeneous Organ
4. Quantifying Adiposity in Clinical Practice
5. Adipokines and Cytokines—Effectors of Adiposity Related Harm
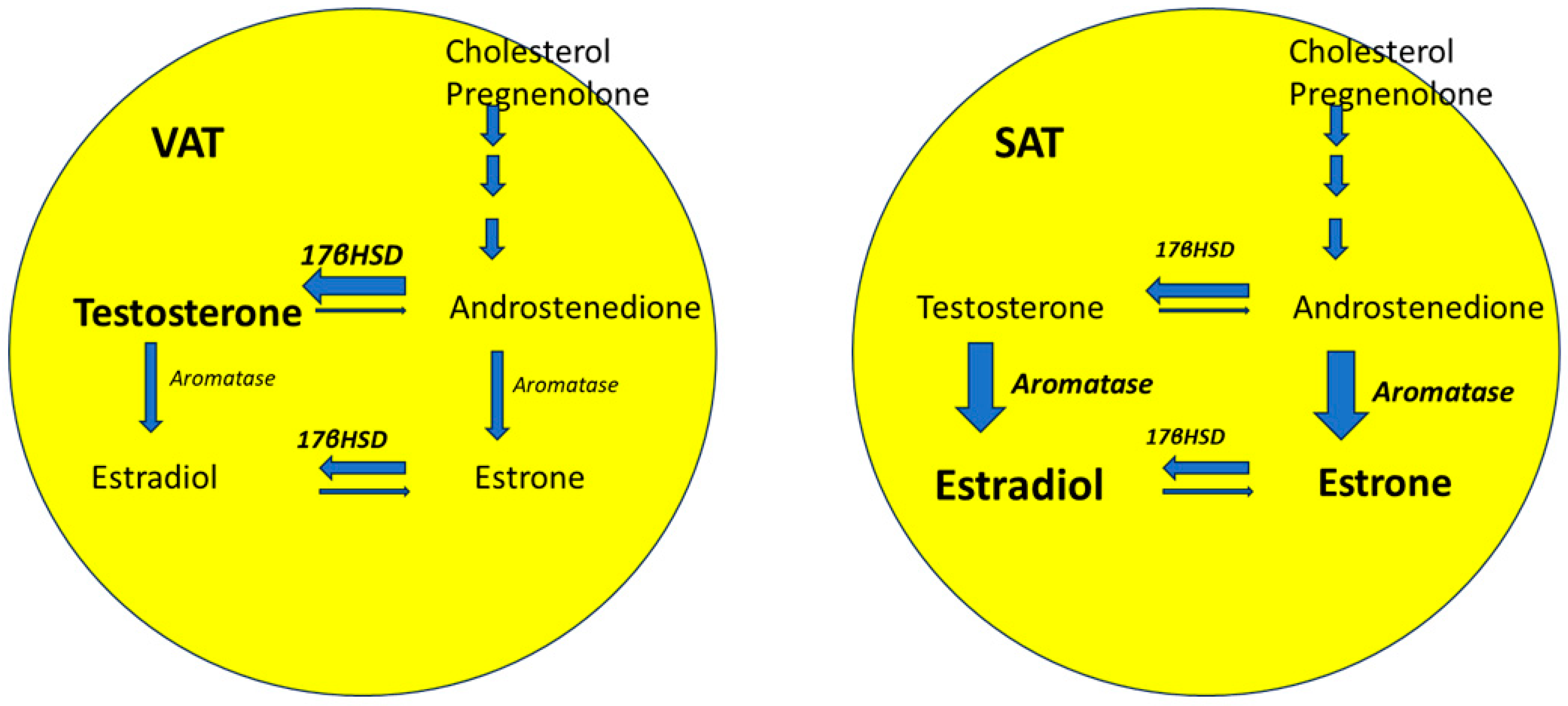
6. Sexual Dimorphism and Adipose Tissue
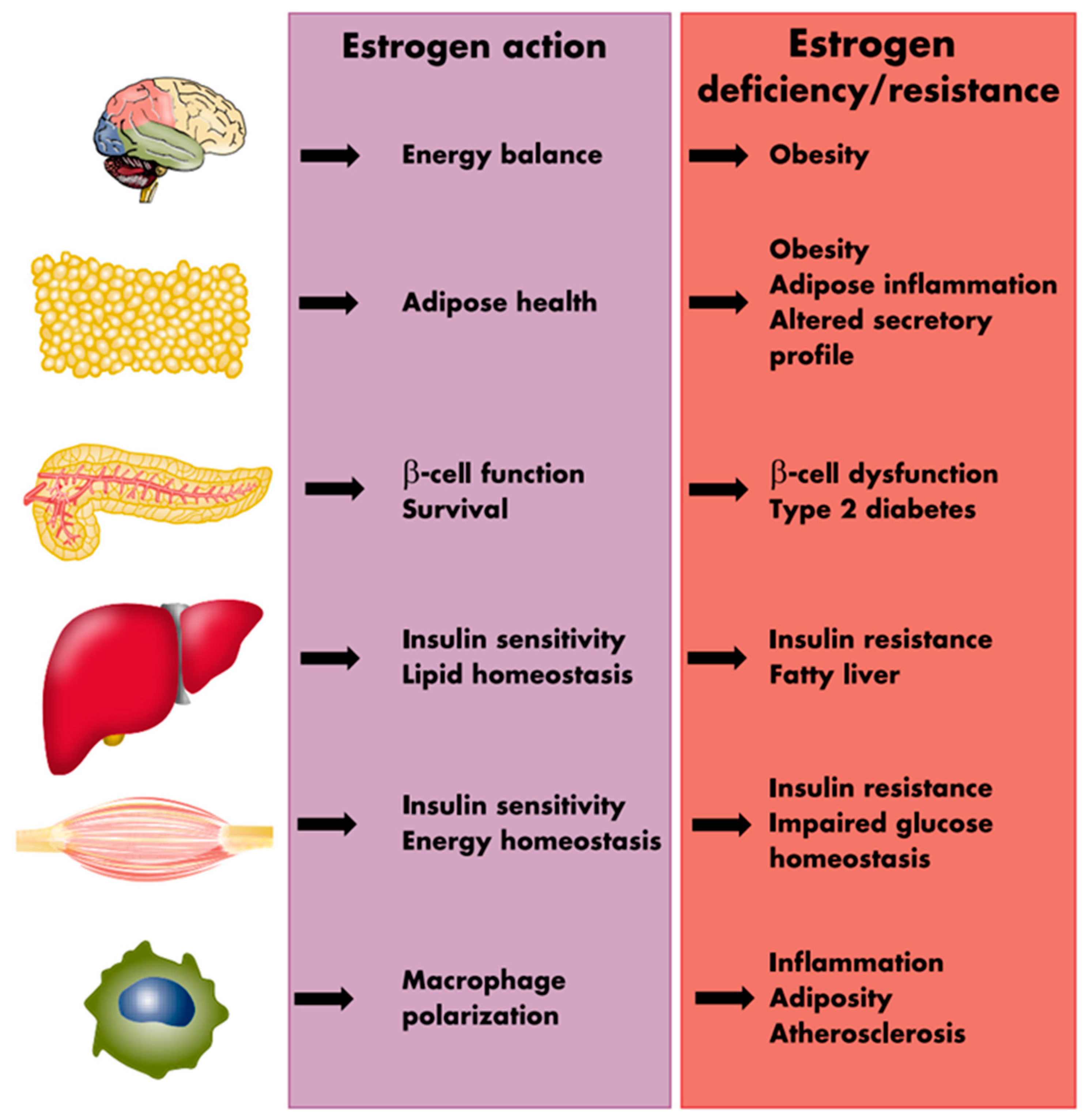
7. Female Reproductive Hormones and Adipose Tissue
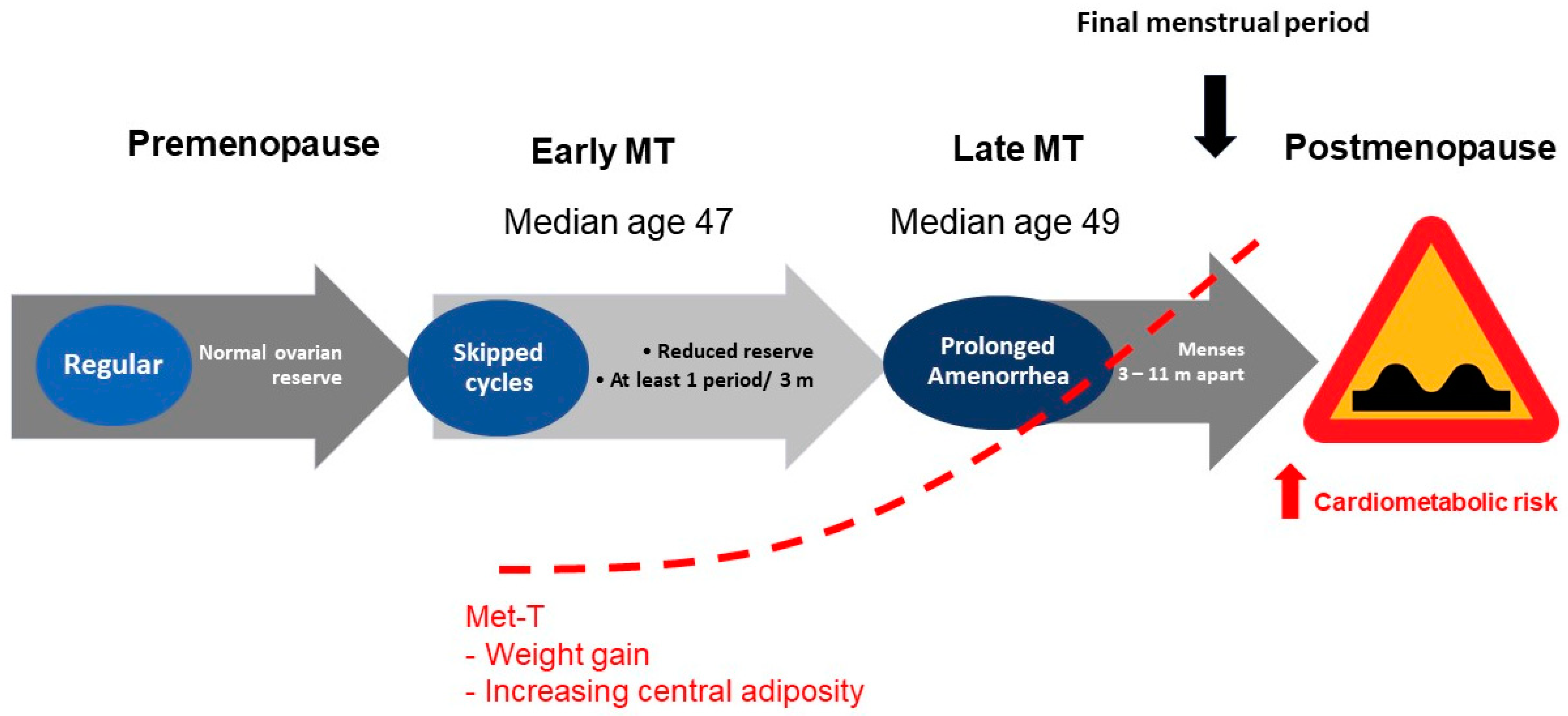
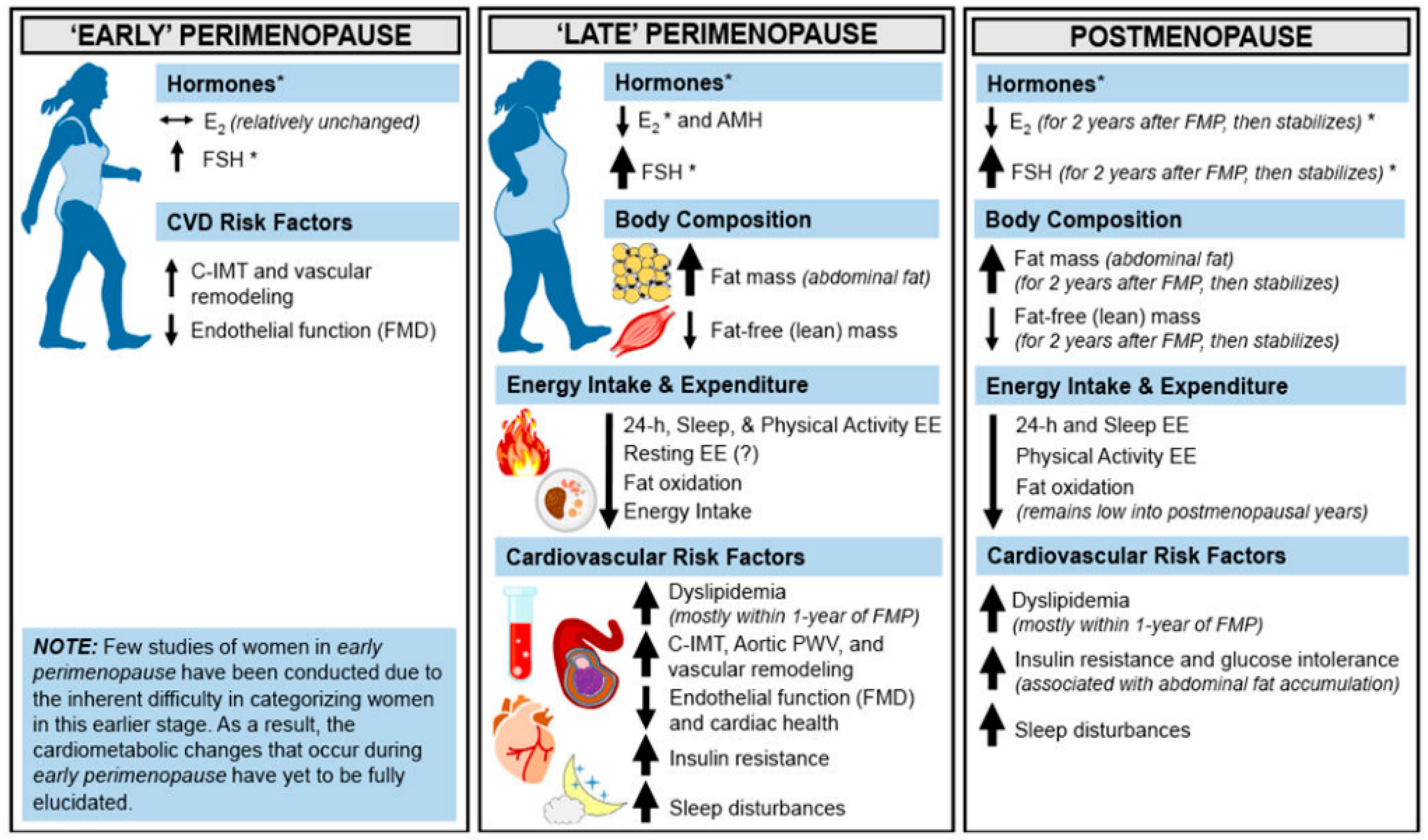
8. Endocrinology of Reproductive Aging and Adiposity
9. Factors Influencing Adiposity during Menopausal Transition
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Harlow, S.D.; Gass, M.; Hall, J.E.; Lobo, R.; Maki, P.; Rebar, R.W.; Sherman, S.; Sluss, P.M.; de Villiers, T.J. Executive summary of the Stages of Reproductive Aging Workshop + 10: Addressing the unfinished agenda of staging reproductive aging. Menopause 2012, 19, 387–395. [Google Scholar] [CrossRef]
- El Khoudary, S.R.; Aggarwal, B.; Beckie, T.M.; Hodis, H.N.; Johnson, A.E.; Langer, R.D.; Limacher, M.C.; Manson, J.E.; Stefanick, M.L.; Allison, M.A.; et al. Menopause Transition and Cardiovascular Disease Risk: Implications for Timing of Early Prevention: A Scientific Statement From the American Heart Association. Circulation 2020, 142, e506–e532. [Google Scholar] [CrossRef]
- Shifren, J.L.; Gass, M.L.S.; NAMS Recommendations for Clinical Care of Midlife Women Working Group. The North American Menopause Society Recommendations for Clinical Care of Midlife Women. Menopause 2014, 21, 1038–1062. [Google Scholar] [CrossRef] [PubMed]
- Flegal, K.M.; Kruszon-Moran, D.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Trends in Obesity Among Adults in the United States, 2005 to 2014. JAMA 2016, 315, 2284–2291. [Google Scholar] [CrossRef] [PubMed]
- Mayes, J.S.; Watson, G.H. Direct effects of sex steroid hormones on adipose tissues and obesity. Obes. Rev. 2004, 5, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G. The metabolic syndrome or the insulin resistance syndrome? Different names, different concepts, and different goals. Endocrinol. Metab. Clin. N. Am. 2004, 33, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.D.; Elliott, M.R.; Riosmena, F.; Cunningham, S.A. Beyond recent BMI: BMI exposure metrics and their relationship to health. SSM Popul. Health 2020, 11, 100547. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Himms-Hagen, J. Brown adipose tissue thermogenesis: Interdisciplinary studies. FASEB J. 1990, 4, 2890–2898. [Google Scholar] [CrossRef]
- Siiteri, P.K. Adipose tissue as a source of hormones. Am. J. Clin. Nutr. 1987, 45, 277–282. [Google Scholar] [CrossRef]
- Shen, W.; Wang, Z.; Punyanita, M.; Lei, J.; Sinav, A.; Kral, J.G.; Imielinska, C.; Ross, R.; Heymsfield, S.B. Adipose tissue quantification by imaging methods: A proposed classification. Obes. Res. 2003, 11, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef]
- Humińska-Lisowska, K.; Mieszkowski, J.; Kochanowicz, A.; Bojarczuk, A.; Niespodziński, B.; Brzezińska, P.; Stankiewicz, B.; Michałowska-Sawczyn, M.; Grzywacz, A.; Petr, M.; et al. Implications of Adipose Tissue Content for Changes in Serum Levels of Exercise-Induced Adipokines: A Quasi-Experimental Study. Int. J. Environ. Res. Public Health 2022, 19, 8782. [Google Scholar] [CrossRef] [PubMed]
- Chechi, K.; Carpentier, A.C.; Richard, D. Understanding the brown adipocyte as a contributor to energy homeostasis. Trends Endocrinol. Metab. 2013, 24, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Esteve Ràfols, M. Tejido adiposo: Heterogeneidad celular y diversidad funcional. Endocrinol. Y Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef]
- Townsend, K.; Tseng, Y.H. Brown adipose tissue: Recent insights into development, metabolic function and therapeutic potential. Adipocyte 2012, 1, 13–24. [Google Scholar] [CrossRef]
- Ravussin, E.; Galgani, J.E. The implication of brown adipose tissue for humans. Annu. Rev. Nutr. 2011, 31, 33–47. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Mancuso, P. The role of adipokines in chronic inflammation. Immunotargets Ther. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Hu, F.B. Measurements of Adiposity and Body Composition. In Obesity Epidemiology; Oxford University Press: Oxford, UK, 2008. [Google Scholar] [CrossRef]
- Rush, E.C.; Goedecke, J.H.; Jennings, C.; Micklesfield, L.; Dugas, L.; Lambert, E.V.; Plank, L.D. BMI, fat and muscle differences in urban women of five ethnicities from two countries. Int. J. Obes. 2007, 31, 1232–1239. [Google Scholar] [CrossRef]
- Yates, A.; Edman, J.; Aruguete, M. Ethnic differences in BMI and body/self-dissatisfaction among Whites, Asian subgroups, Pacific Islanders, and African-Americans. J. Adolesc. Health 2004, 34, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Deurenberg, P. Universal cut-off BMI points for obesity are not appropriate. Br. J. Nutr. 2001, 85, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Deurenberg, P.; Deurenberg-Yap, M. Differences in body-composition assumptions across ethnic groups: Practical consequences. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Deurenberg, P.; Yap, M.; van Staveren, W.A. Body mass index and percent body fat: A meta analysis among different ethnic groups. Int. J. Obes. 1998, 22, 1164–1171. [Google Scholar] [CrossRef]
- WHO expert consultation. Appropriate body-mass index for Asian populations and its implications for policy and intervention strategies. Lancet 2004, 363, 157–163. [Google Scholar] [CrossRef]
- WHO Consultation. Obesity: Preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ. Tech. Rep. Ser. 2000, 894, 1–253. [Google Scholar]
- World Health Organization Western Pacific Region. The Asia-Pacific Perspective: Redefining Obesity and Its Treatment [Internet]; World Health Organization: Geneva, Switzerland; Available online: http://www.wpro.who.int/nutrition/documents/Redefining_obesity/en/ (accessed on 10 November 2023).
- Nishida, C.; Ko, G.T.; Kumanyika, S. Body fat distribution and noncommunicable diseases in populations: Overview of the 2008 WHO Expert Consultation on Waist Circumference and Waist–Hip Ratio. Eur. J. Clin. Nutr. 2010, 64, 2–5. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S.; et al. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef]
- Frühbeck, G. Intracellular signalling pathways activated by leptin. Biochem. J. 2006, 393, 7–20. [Google Scholar] [CrossRef]
- Chan, J.L.; Heist, K.; DePaoli, A.M.; Veldhuis, J.D.; Mantzoros, C.S. The role of falling leptin levels in the neuroendocrine and metabolic adaptation to short-term starvation in healthy men. J. Clin. Investig. 2003, 111, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Considine, R.V.; Considine, E.L.; Williams, C.J.; Nyce, M.R.; Magosin, S.A.; Bauer, T.L.; Rosato, E.L.; Colberg, J.; Caro, J.F. Evidence against either a premature stop codon or the absence of obese gene mRNA in human obesity. J. Clin. Investig. 1995, 95, 2986–2988. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.S.; Paglia, D.; Kwan, A.Y.M.; Deitel, M. Increased obese mRNA expression in omental fat cells from massively obese humans. Nat. Med. 1995, 1, 953–956. [Google Scholar] [CrossRef]
- Considine, R.V.; Sinha, M.K.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Nyce, M.R.; Ohannesian, J.P.; Marco, C.C.; McKee, L.J.; Bauer, T.L.; et al. Serum Immunoreactive-Leptin Concentrations in Normal-Weight and Obese Humans. N. Engl. J. Med. 1996, 334, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.S.; Elahi, D. The effects of acute hyperglycemia and hyperinsulinemia on plasma leptin levels: Its relationships with body fat, visceral adiposity, and age in women. J. Clin. Endocrinol. Metab. 1996, 81, 4433–4438. [Google Scholar] [CrossRef] [PubMed]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef]
- Guerre-Millo, M. Adiponectin: An update. Diabetes Metab. 2008, 34, 12–18. [Google Scholar] [CrossRef]
- Yamauchi, T.; Nio, Y.; Maki, T.; Kobayashi, M.; Takazawa, T.; Iwabu, M.; Okada-Iwabu, M.; Kawamoto, S.; Kubota, N.; Kubota, T.; et al. Targeted disruption of AdipoR1 and AdipoR2 causes abrogation of adiponectin binding and metabolic actions. Nat. Med. 2007, 13, 332–339. [Google Scholar] [CrossRef]
- Weyer, C.; Funahashi, T.; Tanaka, S.; Hotta, K.; Matsuzawa, Y.; Pratley, R.E.; Tataranni, P.A. Hypoadiponectinemia in Obesity and Type 2 Diabetes: Close Association with Insulin Resistance and Hyperinsulinemia. J. Clin. Endocrinol. Metab. 2001, 86, 1930–1935. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Maeda, N.; Takahashi, M.; Funahashi, T.; Kihara, S.; Nishizawa, H.; Kishida, K.; Nagaretani, H.; Matsuda, M.; Komuro, R.; Ouchi, N.; et al. PPARγ Ligands Increase Expression and Plasma Concentrations of Adiponectin, an Adipose-Derived Protein. Diabetes 2001, 50, 2094–2099. [Google Scholar] [CrossRef]
- Lindsay, R.S.; Funahashi, T.; Hanson, R.L.; Matsuzawa, Y.; Tanaka, S.; Tataranni, P.A.; Knowler, W.C.; Krakoff, J. Adiponectin and development of type 2 diabetes in the Pima Indian population. Lancet 2002, 360, 57–58. [Google Scholar] [CrossRef]
- Chandran, M.; Phillips, S.A.; Ciaraldi, T.; Henry, R.R. Adiponectin: More Than Just Another Fat Cell Hormone? Diabetes Care 2003, 26, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Díez, J.J.; Iglesias, P. The role of the novel adipocyte-derived hormone adiponectin in human disease. Eur. J. Endocrinol. 2003, 148, 293–300. [Google Scholar] [CrossRef]
- Päth, G.n.; Bornstein, S.R.; Gurniak, M.; Chrousos, G.P.; Scherbaum, W.A.; Hauner, H. Human Breast Adipocytes Express Interleukin-6 (IL-6) and Its Receptor System: Increased IL-6 Production by β-Adrenergic Activation and Effects of IL-6 on Adipocyte Function. J. Clin. Endocrinol. Metab. 2001, 86, 2281–2288. [Google Scholar] [CrossRef]
- Wallenius, K.; Wallenius, V.; Sunter, D.; Dickson, S.L.; Jansson, J.-O. Intracerebroventricular interleukin-6 treatment decreases body fat in rats. Biochem. Biophys. Res. Commun. 2002, 293, 560–565. [Google Scholar] [CrossRef]
- Vgontzas, A.N.; Papanicolaou, D.A.; Bixler, E.O.; Kales, A.; Tyson, K.; Chrousos, G.P. Elevation of Plasma Cytokines in Disorders of Excessive Daytime Sleepiness: Role of Sleep Disturbance and Obesity. J. Clin. Endocrinol. Metab. 1997, 82, 1313–1316. [Google Scholar] [CrossRef]
- Straub, R.H.; Hense, H.W.; Andus, T.; Schölmerich, J.; Riegger, G.A.J.; Schunkert, H. Hormone Replacement Therapy and Interrelation between Serum Interleukin-6 and Body Mass Index in Postmenopausal Women: A Population-Based Study. J. Clin. Endocrinol. Metab. 2000, 85, 1340–1344. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Miles, P.D.G.; Ladd, C.M.; Ross, K.; Golub, T.R.; Olefsky, J.M.; Lodish, H.F. Profiling Gene Transcription In Vivo Reveals Adipose Tissue as an Immediate Target of Tumor Necrosis Factor-α: Implications for Insulin Resistance. Diabetes 2002, 51, 3176–3188. [Google Scholar] [CrossRef]
- Juhan-Vague, I.; Alessi, M.C.; Mavri, A.; Morange, P.E. Plasminogen activator inhibitor-1, inflammation, obesity, insulin resistance and vascular risk. J. Thromb. Haemost. 2003, 1, 1575–1579. [Google Scholar] [CrossRef] [PubMed]
- Mertens, I.; Gaal, L.F.V. Obesity, haemostasis and the fibrinolytic system. Obes. Rev. 2002, 3, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.R.; Lazar, M.A. Resistin: Molecular history and prognosis. J. Mol. Med. 2003, 81, 218–226. [Google Scholar] [CrossRef]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef]
- Wells, J.C.K. Sexual dimorphism of body composition. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 415–430. [Google Scholar] [CrossRef]
- Vague, J. Sexual differentiation; Factor determining forms of obesity. Presse Med. (1893) 1947, 55, 339. [Google Scholar]
- Lemieux, S.; Prud’homme, D.; Bouchard, C.; Tremblay, A.; Després, J.P. Sex differences in the relation of visceral adipose tissue accumulation to total body fatness. Am. J. Clin. Nutr. 1993, 58, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Allard, C.; Tremblay, A.; Talbot, J.; Bouchard, C. Evidence for a regional component of body fatness in the association with serum lipids in men and women. Metab. Clin. Exp. 1985, 34, 967–973. [Google Scholar] [CrossRef]
- González-García, I.; García-Clavé, E.; Cebrian-Serrano, A.; Le Thuc, O.; Contreras, R.E.; Xu, Y.; Gruber, T.; Schriever, S.C.; Legutko, B.; Lintelmann, J.; et al. Estradiol regulates leptin sensitivity to control feeding via hypothalamic Cited1. Cell Metab. 2023, 35, 438–455.e437. [Google Scholar] [CrossRef]
- Coll, A.P.; Farooqi, I.S.; O’Rahilly, S. The hormonal control of food intake. Cell 2007, 129, 251–262. [Google Scholar] [CrossRef]
- Price, T.M.; O’Brien, S.N. Determination of estrogen receptor messenger ribonucleic acid (mRNA) and cytochrome P450 aromatase mRNA levels in adipocytes and adipose stromal cells by competitive polymerase chain reaction amplification. J. Clin. Endocrinol. Metab. 1993, 77, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Nishikawa, Y.; Adachi, H.; Enomoto, T.; Ikegami, H.; Kurachi, H.; Nomura, T.; Miyake, A. Identification of estrogen receptor in human adipose tissue and adipocytes. J. Clin. Endocrinol. Metab. 1994, 78, 950–954. [Google Scholar] [CrossRef]
- Anwar, A.; McTernan, P.G.; Anderson, L.A.; Askaa, J.; Moody, C.G.; Barnett, A.H.; Eggo, M.C.; Kumar, S. Site-specific regulation of oestrogen receptor-α and -β by oestradiol in human adipose tissue. Diabetes Obes. Metab. 2001, 3, 338–349. [Google Scholar] [CrossRef]
- Joyner, J.M.; Hutley, L.J.; Cameron, D.P. Estrogen receptors in human preadipocytes. Endocrine 2001, 15, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef]
- Pallottini, V.; Bulzomi, P.; Galluzzo, P.; Martini, C.; Marino, M. Estrogen Regulation of Adipose Tissue Functions: Involvement of Estrogen Receptor Isoforms. Infect. Disord. Drug Targets 2008, 8, 52–60. [Google Scholar] [CrossRef]
- Marlatt, K.L.; Pitynski-Miller, D.R.; Gavin, K.M.; Moreau, K.L.; Melanson, E.L.; Santoro, N.; Kohrt, W.M. Body composition and cardiometabolic health across the menopause transition. Obesity 2022, 30, 14–27. [Google Scholar] [CrossRef]
- Zamboni, M.; Armellini, F.; Milani, M.P.; De Marchi, M.; Todesco, T.; Robbi, R.; Bergamo-Andreis, I.A.; Bosello, O. Body fat distribution in pre- and post-menopausal women: Metabolic and anthropometric variables and their inter-relationships. Int. J. Obes. Relat. Metab. Disord. 1992, 16, 495–504. [Google Scholar]
- Ozbey, N.; Sencer, E.; Molvalilar, S.; Orhan, Y. Body Fat Distribution and Cardiovascular Disease Risk Factors in Pre- and Postmenopausal Obese Women with Similar BMI. Endocr. J. 2002, 49, 503–509. [Google Scholar] [CrossRef]
- Svendsen, O.L.; Hassager, C.; Christiansen, C. Age- and menopause-associated variations in body composition and fat distribution in healthy women as measured by dual-energy x-ray absorptiometry. Metab. Clin. Exp. 1995, 44, 369–373. [Google Scholar] [CrossRef]
- Karvonen-Gutierrez, C.; Kim, C. Association of Mid-Life Changes in Body Size, Body Composition and Obesity Status with the Menopausal Transition. Healthcare 2016, 4, 42. [Google Scholar] [CrossRef]
- Sowers, M.R.; Eyvazzadeh, A.D.; McConnell, D.; Yosef, M.; Jannausch, M.L.; Zhang, D.; Harlow, S.; Randolph, J.F., Jr. Anti-mullerian hormone and inhibin B in the definition of ovarian aging and the menopause transition. J. Clin. Endocrinol. Metab. 2008, 93, 3478–3483. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.G.; Cahir, N.; Robertson, D.M.; Groome, N.P.; Dudley, E.; Green, A.; Dennerstein, L. Serum Inhibins A and B fall differentially as FSH rises in perimenopausal women. Clin. Endocrinol. 1998, 48, 809–813. [Google Scholar] [CrossRef]
- Burger, H.G.; Hale, G.E.; Robertson, D.M.; Dennerstein, L. A review of hormonal changes during the menopausal transition: Focus on findings from the Melbourne Women’s Midlife Health Project. Hum. Reprod. Update 2007, 13, 559–565. [Google Scholar] [CrossRef]
- Fogle, R.H.; Stanczyk, F.Z.; Zhang, X.; Paulson, R.J. Ovarian Androgen Production in Postmenopausal Women. J. Clin. Endocrinol. Metab. 2007, 92, 3040–3043. [Google Scholar] [CrossRef] [PubMed]
- Gershagen, S.; Doeberl, A.; Jeppsson, S.; Rannevik, G. Decreasing serum levels of sex hormone-binding globulin around the menopause and temporary relation to changing levels of ovarian steroids, as demonstrated in a longitudinal study. Fertil. Steril. 1989, 51, 616–621. [Google Scholar] [CrossRef]
- Brzozowska, M.; Lewiński, A. Changes of androgens levels in menopausal women. Prz. Menopauzalny 2020, 19, 151–154. [Google Scholar] [CrossRef]
- Guthrie, J.R.; Dennerstein, L.; Taffe, J.R.; Lehert, P.; Burger, H.G. The menopausal transition: A 9-year prospective population-based study. The Melbourne Women’s Midlife Health Project. Climacteric 2004, 7, 375–389. [Google Scholar] [CrossRef]
- Zaman, A.; Rothman, M.S. Postmenopausal Hyperandrogenism: Evaluation and Treatment Strategies. Endocrinol. Metab. Clin. North Am. 2021, 50, 97–111. [Google Scholar] [CrossRef]
- Brodowska, A.; Brodowski, J.; Laszczyńska, M.; Słuczanowska-Głąbowska, S.; Rumianowski, B.; Rotter, I.; Starczewski, A.; Ratajczak, M.Z. Immunoexpression of aromatase cytochrome P450 and 17β-hydroxysteroid dehydrogenase in women’s ovaries after menopause. J. Ovarian Res. 2014, 7, 52. [Google Scholar] [CrossRef]
- Meseguer, A.; Puche, C.; Cabero, A. Sex Steroid Biosynthesis in White Adipose Tissue. Horm. Metab. Res. 2002, 34, 731–736. [Google Scholar] [CrossRef]
- Bélanger, C.; Luu-The, V.; Dupont, P.; Tchernof, A. Adipose Tissue Intracrinology: Potential Importance of Local Androgen/Estrogen Metabolism in the Regulation of Adiposity. Horm. Metab. Res. 2002, 34, 737–745. [Google Scholar] [CrossRef]
- Gambacciani, M.; Ciaponi, M.; Cappagli, B.; Piaggesi, L.; De Simone, L.; Orlandi, R.; Genazzani, A.R. Body Weight, Body Fat Distribution, and Hormonal Replacement Therapy in Early Postmenopausal Women. J. Clin. Endocrinol. Metab. 1997, 82, 414–417. [Google Scholar] [CrossRef]
- Haarbo, J.; Hansen, B.F.; Christiansen, C. Hormone replacement therapy prevents coronary artery disease in ovariectomized cholesterol-fed rabbits. APMIS 1991, 99, 721–727. [Google Scholar] [CrossRef]
- Costa, G.B.C.; Carneiro, G.; Umeda, L.; Pardini, D.; Zanella, M.T. Influence of Menopausal Hormone Therapy on Body Composition and Metabolic Parameters. Biores. Open Access 2020, 9, 80–85. [Google Scholar] [CrossRef]
- Salpeter, S.R.; Walsh, J.M.E.; Ormiston, T.M.; Greyber, E.; Buckley, N.S.; Salpeter, E.E. Meta-analysis: Effect of hormone-replacement therapy on components of the metabolic syndrome in postmenopausal women. Diabetes Obes. Metab. 2006, 8, 538–554. [Google Scholar] [CrossRef]
- Bondarev, D.; Finni, T.; Kokko, K.; Kujala, U.M.; Aukee, P.; Kovanen, V.; Laakkonen, E.K.; Sipilä, S. Physical Performance During the Menopausal Transition and the Role of Physical Activity. J. Gerontol. Ser. A 2021, 76, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Sternfeld, B.; Dugan, S. Physical activity and health during the menopausal transition. Obs. Gynecol. Clin. North Am. 2011, 38, 537–566. [Google Scholar] [CrossRef] [PubMed]
- Juppi, H.K.; Sipilä, S.; Fachada, V.; Hyvärinen, M.; Cronin, N.; Aukee, P.; Karppinen, J.E.; Selänne, H.; Kujala, U.M.; Kovanen, V.; et al. Total and regional body adiposity increases during menopause-evidence from a follow-up study. Aging Cell 2022, 21, e13621. [Google Scholar] [CrossRef] [PubMed]
- Cespedes Feliciano, E.M.; Tinker, L.; Manson, J.E.; Allison, M.; Rohan, T.; Zaslavsky, O.; Waring, M.E.; Asao, K.; Garcia, L.; Rosal, M.; et al. Change in Dietary Patterns and Change in Waist Circumference and DXA Trunk Fat Among Postmenopausal Women. Obesity 2016, 24, 2176–2184. [Google Scholar] [CrossRef] [PubMed]
- Rask-Andersen, M.; Karlsson, T.; Ek, W.E.; Johansson, Å. Genome-wide association study of body fat distribution identifies adiposity loci and sex-specific genetic effects. Nat. Commun. 2019, 10, 339. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADIPOCYTE | WHITE | BEIGE | BROWN |
|---|---|---|---|
| CELL TYPE |  |  |  |
| APPEARANCE | Unilocular lipid droplets White (WAT) | Multilocular Beige | Multilocular lipid droplets Brown (BAT) |
| FUNCTION | Energy reservoir, makes up bulk of stored adipose tissue. During high energy needs, surplus energy stored via glucose uptake and lipogenesis. Release fatty acids via lipolysis during low energy supply/increased expenditure | Bifunctional. Low basal UCP-1 Suitable for energy storage. In response to cold or sympathetic stimulation, white adipocytes increase expression of UCP-1 and release heat, “browning” | High basal UCP-1 Generates heat via uncoupling of UCP-1 in mitochondria: non shivering thermogenesis |
| UCP-1 EXPRESSION | Absent | Low | High |
| MITOCHONDRIAL DENSITY | Low | Medium | High |
| LOCATION | In the connective tissue beneath the skin (SAT) and in abdominal cavity (VAT) | WAT locations derived from white fat by “browning” | Between shoulder blades, neck, along spinal cord, and collar bone |
| Measure | Calculation | Strength | Limitation |
|---|---|---|---|
| Body Mass Index (BMI) [20] |
|
|
|
| Waist Circumference (WC) [20] |
|
|
|
| Waist-to-Hip Ratio (WHR) [20] |
|
|
|
| Skinfold Thickness (SFT) [20] | Requires a special caliper to measure site specific thickness of a “pinch” of skin and the fat beneath Sites that can be assessed include the trunk, thighs, front and back of the upper arm, and under the shoulder blade. Equations are used to predict BF% based on SFT. | Simple Safe Portable Reusable caliper Fast and easy (except in individuals with a BMI of 35 or higher) | Lesser accuracy and reproducibility of measurements in comparison to other methods Difficult to measure in individuals of BMI ≥ 35 kg/m2 |
| Bioelectric Impedance (BIA) [20] |
|
|
|
| Dual Energy X-ray Absorptiometry (DXA) [20] |
|
|
|
| Computerized Tomography (CT) [20] |
|
|
|
| Magnetic Resonance Imaging (MRI) [20] |
|
|
|
| Categories | Non-Asians | Asians |
|---|---|---|
| BMI | ||
| Underweight | <18.5 kg/m2 | <18.5 kg/m2 |
| Ideal weight | 18.5–24.9 kg/m2 | 18.5–23 kg/m2 |
| Overweight | 25–29.9 kg/m2 | 23–27.5 kg/m2 |
| Obesity | ≥30 kg/m2 | >27.5 kg/m2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, A.; Lathief, S.; Charoenngam, N.; Pal, L. Aging and Adiposity—Focus on Biological Females at Midlife and Beyond. Int. J. Mol. Sci. 2024, 25, 2972. https://doi.org/10.3390/ijms25052972
Rehman A, Lathief S, Charoenngam N, Pal L. Aging and Adiposity—Focus on Biological Females at Midlife and Beyond. International Journal of Molecular Sciences. 2024; 25(5):2972. https://doi.org/10.3390/ijms25052972
Chicago/Turabian StyleRehman, Amna, Sanam Lathief, Nipith Charoenngam, and Lubna Pal. 2024. "Aging and Adiposity—Focus on Biological Females at Midlife and Beyond" International Journal of Molecular Sciences 25, no. 5: 2972. https://doi.org/10.3390/ijms25052972
APA StyleRehman, A., Lathief, S., Charoenngam, N., & Pal, L. (2024). Aging and Adiposity—Focus on Biological Females at Midlife and Beyond. International Journal of Molecular Sciences, 25(5), 2972. https://doi.org/10.3390/ijms25052972