
Mitotic Spindle Positioning (MISP) Facilitates Colorectal Cancer Progression by Forming a Complex with Opa Interacting Protein 5 (OIP5) and Activating the JAK2-STAT3 Signaling Pathway
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MISP Expression Is Elevated in Patients with CRC
2.2. MISP Enhances Cell Proliferation in Human CRC Cell Lines
2.3. Impact of MISP on Inflammation in a Mouse Model of Colitis-Induced Colon Tumors
2.4. MISP Facilitates the Growth of Colitis-Induced Colorectal Tumors
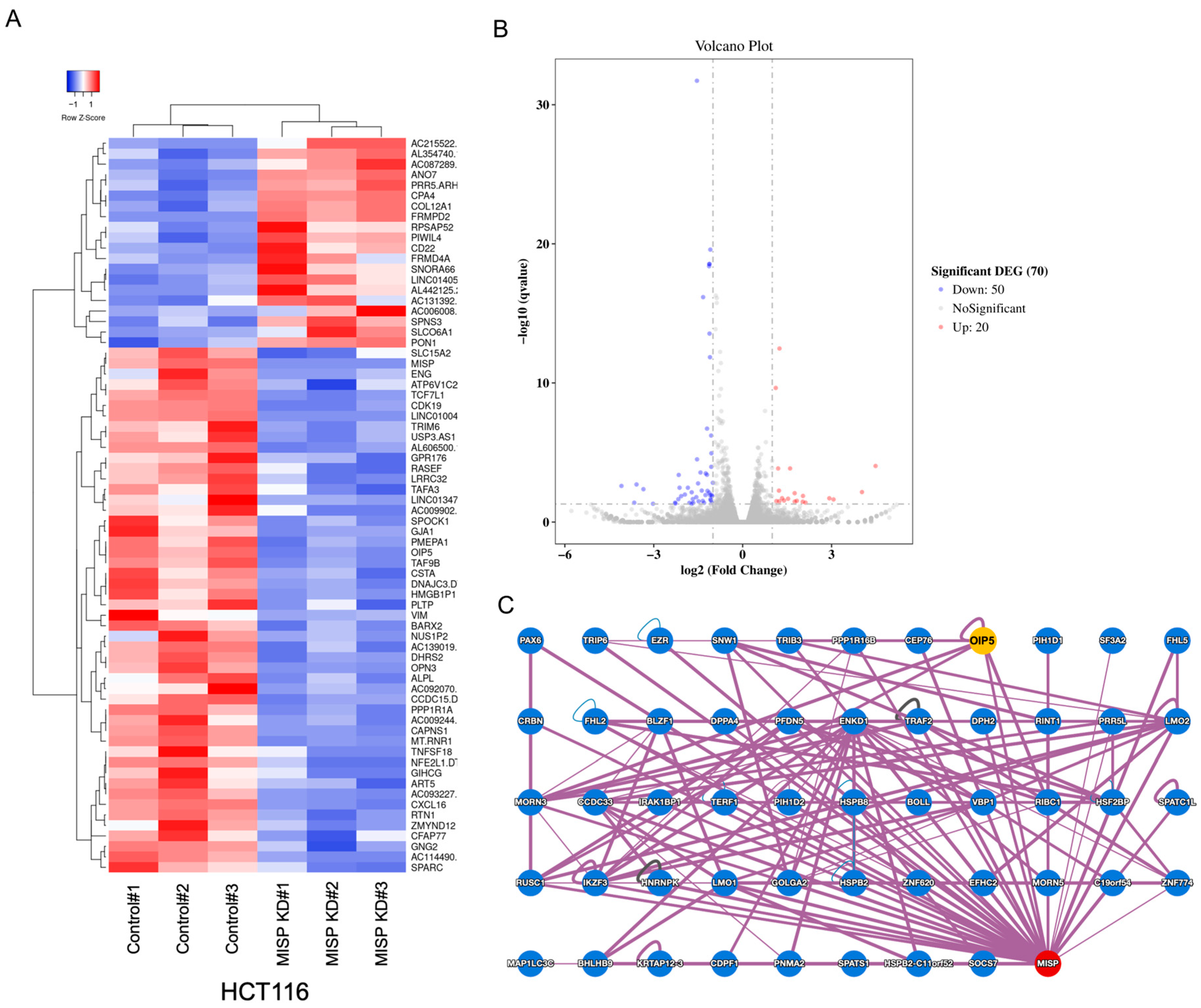
2.5. Investigating Genes Associated with MISP in CRC
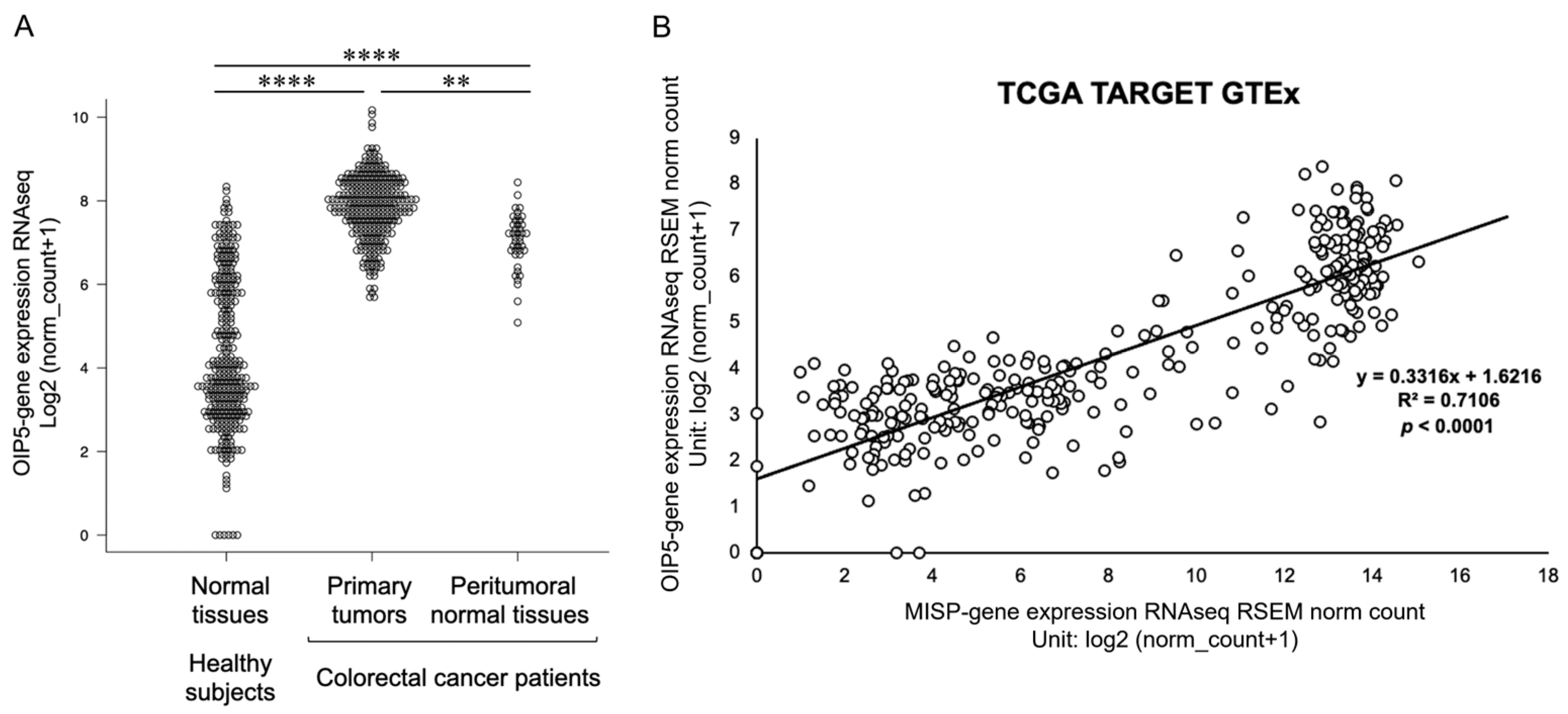
2.6. Elevated Expression of OIP5 in Colorectal Cancer Patients and Its Correlation with MISP Expression in the Colon
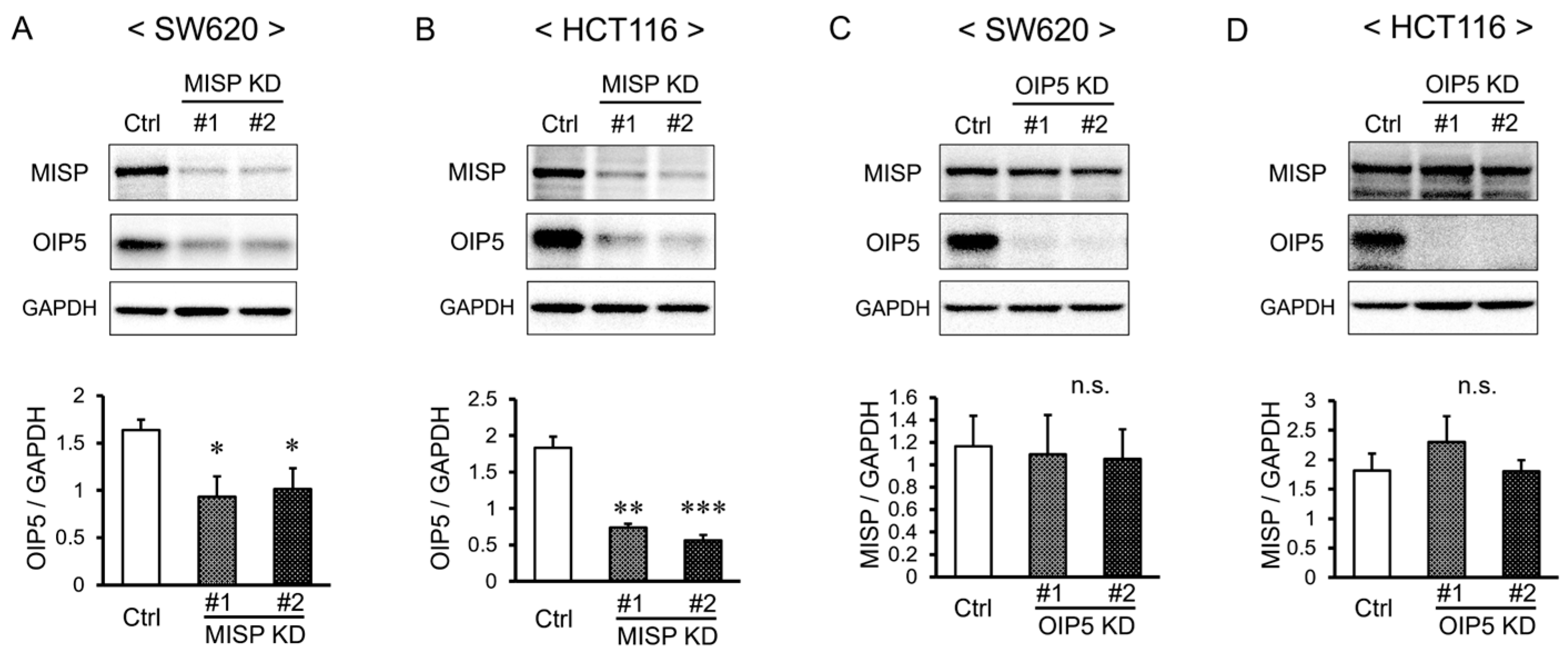
2.7. MISP Influences OIP5 Expression through Complex Formation, While OIP5 Does Not Alter MISP Expression
2.8. Reduction in MISP Expression Decreases the Levels of Phosphorylated STAT3 in Colorectal Cancer Cells
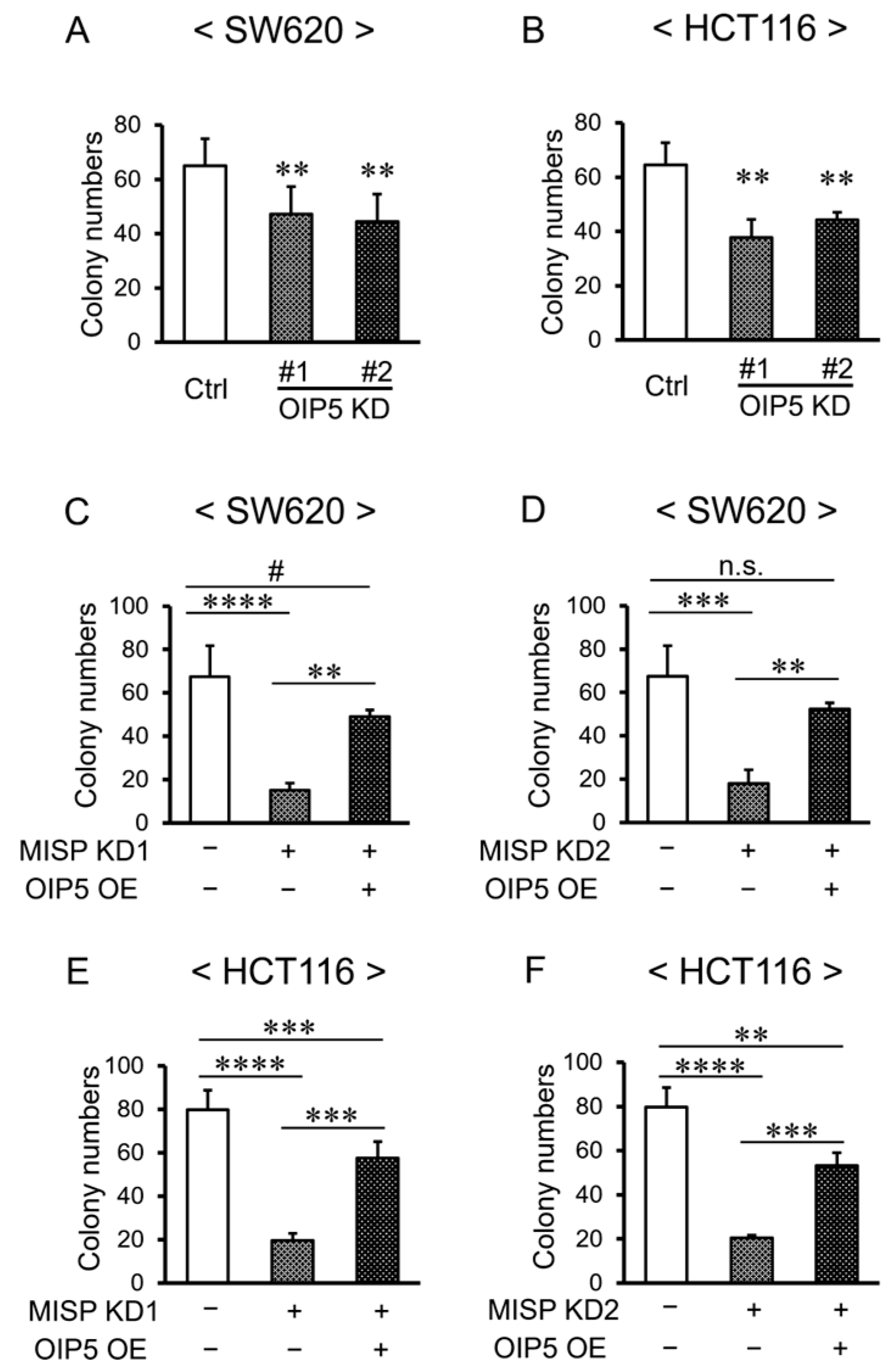
2.9. Rescue of Reduced Cell Proliferation through OIP5 Overexpression in the Context of Decreased MISP Expression
3. Discussion
4. Materials and Methods
4.1. Gene Expression Analysis Utilizing the Cancer Genome Database
4.2. Cell Culture
4.3. Transfection of siRNA into Human CRC Cell Lines
4.4. Colony Formation Assay
4.5. Ethical Statement
4.6. Animals
4.7. Colitis-Induced Colon Tumor Mouse Model
4.8. Exploration of MISP-Related Factors in Colorectal Tumorigenesis
4.9. Gene Expression Correlation Analysis
4.10. Immunofluorescence Staining
4.11. Western Blot Analysis
4.12. Construction of the MISP Expression Plasmid
4.13. Investigation of the Interaction between MISP and OIP5 through Immunoprecipitation
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.W.; Harpaz, N.; Itzkowitz, S.H.; Parsons, R.E. Molecular Mechanisms in Colitis-Associated Colorectal Cancer. Oncogenesis 2023, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Cho, J.H. Inflammatory Bowel Disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Mehandru, S.; Colombel, J.-F.; Peyrin-Biroulet, L. Crohn’s Disease. Lancet 2017, 389, 1741–1755. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.-F. Ulcerative Colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Gargalionis, A.N.; Papavassiliou, K.A.; Papavassiliou, A.G. Targeting STAT3 Signaling Pathway in Colorectal Cancer. Biomedicines 2021, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Lang, X.; Li, X. The Role of IL-6/JAK2/STAT3 Signaling Pathway in Cancers. Front. Oncol. 2022, 12, 1023177. [Google Scholar] [CrossRef]
- Leystra, A.A.; Clapper, M.L. Gut Microbiota Influences Experimental Outcomes in Mouse Models of Colorectal Cancer. Genes 2019, 10, 900. [Google Scholar] [CrossRef]
- Yu, L.C.-H.; Wei, S.-C.; Ni, Y.-H. Impact of Microbiota in Colorectal Carcinogenesis: Lessons from Experimental Models. Intest. Res. 2018, 16, 346–357. [Google Scholar] [CrossRef]
- Wu, M.; Li, J.; An, Y.; Li, P.; Xiong, W.; Li, J.; Yan, D.; Wang, M.; Zhong, G. Chitooligosaccharides Prevents the Development of Colitis-Associated Colorectal Cancer by Modulating the Intestinal Microbiota and Mycobiota. Front. Microbiol. 2019, 10, 2101. [Google Scholar] [CrossRef] [PubMed]
- Haberman, Y.; Karns, R.; Dexheimer, P.J.; Schirmer, M.; Somekh, J.; Jurickova, I.; Braun, T.; Novak, E.; Bauman, L.; Collins, M.H.; et al. Ulcerative Colitis Mucosal Transcriptomes Reveal Mitochondriopathy and Personalized Mechanisms Underlying Disease Severity and Treatment Response. Nat. Commun. 2019, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Kumeta, M.; Gilmore, J.L.; Umeshima, H.; Ishikawa, M.; Kitajiri, S.; Horigome, T.; Kengaku, M.; Takeyasu, K. Caprice/MISP Is a Novel F-Actin Bundling Protein Critical for Actin-Based Cytoskeletal Reorganizations. Genes Cells 2014, 19, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Kirsch, M.; Anderhub, S.; Zentgraf, H.; Krämer, A. The Novel Actin/Focal Adhesion-Associated Protein MISP Is Involved in Mitotic Spindle Positioning in Human Cells. Cell Cycle 2013, 12, 1457–1471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Settele, F.; Kotak, S.; Sanchez-Pulido, L.; Ehret, L.; Ponting, C.P.; Gönczy, P.; Hoffmann, I. MISP Is a Novel Plk1 Substrate Required for Proper Spindle Orientation and Mitotic Progression. J. Cell Biol. 2013, 200, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.A.; Arnaiz, C.; Krystofiak, E.S.; Zanic, M.; Tyska, M.J. Mitotic Spindle Positioning (MISP) Is an Actin Bundler That Selectively Stabilizes the Rootlets of Epithelial Microvilli. Cell Rep. 2022, 39, 110692. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, A.; Torre, D.; Keenan, A.B.; Jagodnik, K.M.; Lee, H.J.; Wang, L.; Silverstein, M.C.; Ma’ayan, A. Massive mining of publicly available RNA-seq data from human and mouse. Nat. Commun. 2018, 9, 1366. [Google Scholar] [CrossRef]
- Hiura, K.; Maruyama, T.; Watanabe, M.; Nakano, K.; Okamura, T.; Sasaki, H.; Sasaki, N. Mitotic Spindle Positioning Protein (MISP) Deficiency Exacerbates Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. J. Vet. Med. Sci. 2022, 85, 167–174. [Google Scholar] [CrossRef]
- Vodicska, B.; Cerikan, B.; Schiebel, E.; Hoffmann, I. MISP Regulates the IQGAP1/Cdc42 Complex to Collectively Orchestrate Spindle Orientation and Mitotic Progression. Sci. Rep. 2018, 8, 6330. [Google Scholar] [CrossRef]
- Casper, J.; Zweig, A.S.; Villarreal, C.; Tyner, C.; Speir, M.L.; Rosenbloom, K.R.; Raney, B.J.; Lee, C.M.; Lee, B.T.; Karolchik, D.; et al. The UCSC Genome Browser Database: 2018 Update. Nucleic Acids Res. 2018, 46, D762–D769. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and Interpreting Cancer Genomics Data via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Zweig, A.S.; Karolchik, D.; Kuhn, R.M.; Haussler, D.; Kent, W.J. UCSC Genome Browser Tutorial. Genomics 2008, 92, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic Study of Dextran Sulfate Sodium Experimental Murine Colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Ghia, J.-E.; Blennerhassett, P.; Collins, S.M. Impaired Parasympathetic Function Increases Susceptibility to Inflammatory Bowel Disease in a Mouse Model of Depression. J. Clin. Investig. 2008, 118, 2209–2218. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for Inferring Cellular Functions from Protein Sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG Mapping Tools for Uncovering Hidden Features in Biological Data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]
- Luck, K.; Kim, D.-K.; Lambourne, L.; Spirohn, K.; Begg, B.E.; Bian, W.; Brignall, R.; Cafarelli, T.; Campos-Laborie, F.J.; Charloteaux, B.; et al. A Reference Map of the Human Binary Protein Interactome. Nature 2020, 580, 402–408. [Google Scholar] [CrossRef]
- Xu, H.; Wang, J.; Liu, J.; Huang, J. Opa Interacting Protein 5 Promotes Proliferation and Migration of Trophoblast Cells via Activating STAT3 Pathway. Reprod. Biol. 2022, 22, 100639. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, L.; Jin, Y.; Wang, Z.; Li, T.; Xu, H.; Wang, Q.; Wang, L. Up-Regulated MISP Is Associated With Poor Prognosis and Immune Infiltration in Pancreatic Ductal Adenocarcinoma. Front. Oncol. 2022, 12, 827051. [Google Scholar] [CrossRef]
- Jiminez, J.A.; Uwiera, T.C.; Douglas Inglis, G.; Uwiera, R.R.E. Animal Models to Study Acute and Chronic Intestinal Inflammation in Mammals. Gut Pathog. 2015, 7, 29. [Google Scholar] [CrossRef]
- Neufert, C.; Becker, C.; Neurath, M.F. An Inducible Mouse Model of Colon Carcinogenesis for the Analysis of Sporadic and Inflammation-Driven Tumor Progression. Nat. Protoc. 2007, 2, 1998–2004. [Google Scholar] [CrossRef]
- Chun, H.-K.; Chung, K.-S.; Kim, H.C.; Kang, J.-E.; Kang, M.A.; Kim, J.-T.; Choi, E.H.; Jung, K.-E.; Kim, M.-H.; Song, E.Y.; et al. OIP5 Is a Highly Expressed Potential Therapeutic Target for Colorectal and Gastric Cancers. BMB Rep. 2010, 43, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiao, F.; Li, W.; Hu, P.; Xu, R.; Li, J.; Li, G.; Zhu, C. Overexpression of Opa Interacting Protein 5 Increases the Progression of Liver Cancer via BMPR2/JUN/CHEK1/RAC1 Dysregulation. Oncol. Rep. 2019, 41, 2075–2088. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Li, Y.; Song, E.; Li, M.; Qiu, S.; Dong, W.; Yuan, R. OIP5 Is a Novel Prognostic Biomarker in Clear Cell Renal Cell Cancer Correlating With Immune Infiltrates. Front. Immunol. 2022, 13, 805552. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Z.; Lin, F.; Wang, Z.; Gao, Q.; Xie, H.; Xiao, H.; Zhou, Y.; Zhang, F.; Ma, Y.; et al. OIP5 Promotes Growth, Metastasis and Chemoresistance to Cisplatin in Bladder Cancer Cells. J. Cancer 2018, 9, 4684–4695. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Feng, W.; Cai, H.; Mei, Y.; Jiang, Y.; Chen, T.; Xu, K.; Feng, D. Loss of the Opa Interacting Protein 5 Inhibits Breast Cancer Proliferation through MiR-139-5p/NOTCH1 Pathway. Gene 2017, 603, 1–8. [Google Scholar] [CrossRef]
- Chen, L.; Wang, R.; Gao, L.; Shi, W. Opa-Interacting Protein 5 Expression in Human Glioma Tissues Is Essential to the Biological Function of U251 Human Malignant Glioma Cells. Cancer Control. 2020, 27, 107327482096891. [Google Scholar] [CrossRef]
- Naetar, N.; Hutter, S.; Dorner, D.; Dechat, T.; Korbei, B.; Gotzmann, J.; Beug, H.; Foisner, R. LAP2α-Binding Protein LINT-25 Is a Novel Chromatin-Associated Protein Involved in Cell Cycle Exit. J. Cell Sci. 2007, 120, 737–747. [Google Scholar] [CrossRef]
- Fujita, Y.; Hayashi, T.; Kiyomitsu, T.; Toyoda, Y.; Kokubu, A.; Obuse, C.; Yanagida, M. Priming of Centromere for CENP-A Recruitment by Human HMis18α, HMis18β, and M18BP1. Dev. Cell 2007, 12, 17–30. [Google Scholar] [CrossRef]
- Xia, X.-H.; Xiao, C.-J.; Shan, H. Facilitation of Liver Cancer SMCC7721 Cell Aging by Sirtuin 4 via Inhibiting JAK2/STAT3 Signal Pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1248–1253. [Google Scholar] [PubMed]
- Yu, H.; Pardoll, D.; Jove, R. STATs in Cancer Inflammation and Immunity: A Leading Role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Xiong, H.; Zhang, Z.-G.; Tian, X.-Q.; Sun, D.-F.; Liang, Q.-C.; Zhang, Y.-J.; Lu, R.; Chen, Y.-X.; Fang, J.-Y. Inhibition of JAK1, 2/STAT3 Signaling Induces Apoptosis, Cell Cycle Arrest, and Reduces Tumor Cell Invasion in Colorectal Cancer Cells. Neoplasia 2008, 10, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulos, P.; Zizi-Sermpetzoglou, A.; Rizos, S.; Kostakis, A.; Nikiteas, N.; Papavassiliou, A.G. TGF-Beta Signalling in Colon Carcinogenesis. Cancer Lett. 2012, 314, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. Transcriptional Mechanisms of WNT5A Based on NF-ΚB, Hedgehog, TGFβ, and Notch Signaling Cascades. Int. J. Mol. Med. 2009, 23, 763–769. [Google Scholar] [CrossRef]
- Dissanayake, S.K.; Wade, M.; Johnson, C.E.; O’Connell, M.P.; Leotlela, P.D.; French, A.D.; Shah, K.V.; Hewitt, K.J.; Rosenthal, D.T.; Indig, F.E.; et al. The Wnt5A/Protein Kinase C Pathway Mediates Motility in Melanoma Cells via the Inhibition of Metastasis Suppressors and Initiation of an Epithelial to Mesenchymal Transition. J. Biol. Chem. 2007, 282, 17259–17271. [Google Scholar] [CrossRef]
- Yu, M.R.; Kim, H.J.; Park, H.R. Fusobacterium Nucleatum Accelerates the Progression of Colitis-Associated Colorectal Cancer by Promoting EMT. Cancers 2020, 12, 2728. [Google Scholar] [CrossRef]
- Bhowmick, N.A.; Ghiassi, M.; Bakin, A.; Aakre, M.; Lundquist, C.A.; Engel, M.E.; Arteaga, C.L.; Moses, H.L. Transforming Growth Factor-Β1 Mediates Epithelial to Mesenchymal Transdifferentiation through a RhoA-Dependent Mechanism. Mol. Biol. Cell 2001, 12, 27–36. [Google Scholar] [CrossRef]
- Spaderna, S.; Schmalhofer, O.; Hlubek, F.; Berx, G.; Eger, A.; Merkel, S.; Jung, A.; Kirchner, T.; Brabletz, T. A Transient, EMT-Linked Loss of Basement Membranes Indicates Metastasis and Poor Survival in Colorectal Cancer. Gastroenterology 2006, 131, 830–840. [Google Scholar] [CrossRef]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient Selection for High-Expression Transfectants with a Novel Eukaryotic Vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the Freely Available Easy-to-Use Software ‘EZR’ for Medical Statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiura, K.; Watanabe, M.; Hirose, N.; Nakano, K.; Okamura, T.; Sasaki, H.; Sasaki, N. Mitotic Spindle Positioning (MISP) Facilitates Colorectal Cancer Progression by Forming a Complex with Opa Interacting Protein 5 (OIP5) and Activating the JAK2-STAT3 Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 3061. https://doi.org/10.3390/ijms25053061
Hiura K, Watanabe M, Hirose N, Nakano K, Okamura T, Sasaki H, Sasaki N. Mitotic Spindle Positioning (MISP) Facilitates Colorectal Cancer Progression by Forming a Complex with Opa Interacting Protein 5 (OIP5) and Activating the JAK2-STAT3 Signaling Pathway. International Journal of Molecular Sciences. 2024; 25(5):3061. https://doi.org/10.3390/ijms25053061
Chicago/Turabian StyleHiura, Koki, Masaki Watanabe, Naoki Hirose, Kenta Nakano, Tadashi Okamura, Hayato Sasaki, and Nobuya Sasaki. 2024. "Mitotic Spindle Positioning (MISP) Facilitates Colorectal Cancer Progression by Forming a Complex with Opa Interacting Protein 5 (OIP5) and Activating the JAK2-STAT3 Signaling Pathway" International Journal of Molecular Sciences 25, no. 5: 3061. https://doi.org/10.3390/ijms25053061
APA StyleHiura, K., Watanabe, M., Hirose, N., Nakano, K., Okamura, T., Sasaki, H., & Sasaki, N. (2024). Mitotic Spindle Positioning (MISP) Facilitates Colorectal Cancer Progression by Forming a Complex with Opa Interacting Protein 5 (OIP5) and Activating the JAK2-STAT3 Signaling Pathway. International Journal of Molecular Sciences, 25(5), 3061. https://doi.org/10.3390/ijms25053061