
Prolonged Heat Stress during Winter Diapause Alters the Expression of Stress-Response Genes in Ostrinia nubilalis (Hbn.)

 ,
,  , , , and
, , , and 
Abstract
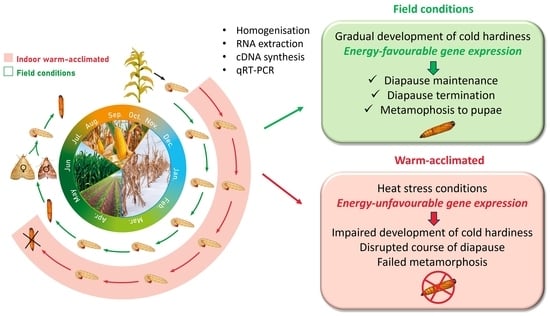
:
1. Introduction
2. Results
2.1. ADS-Related Genes
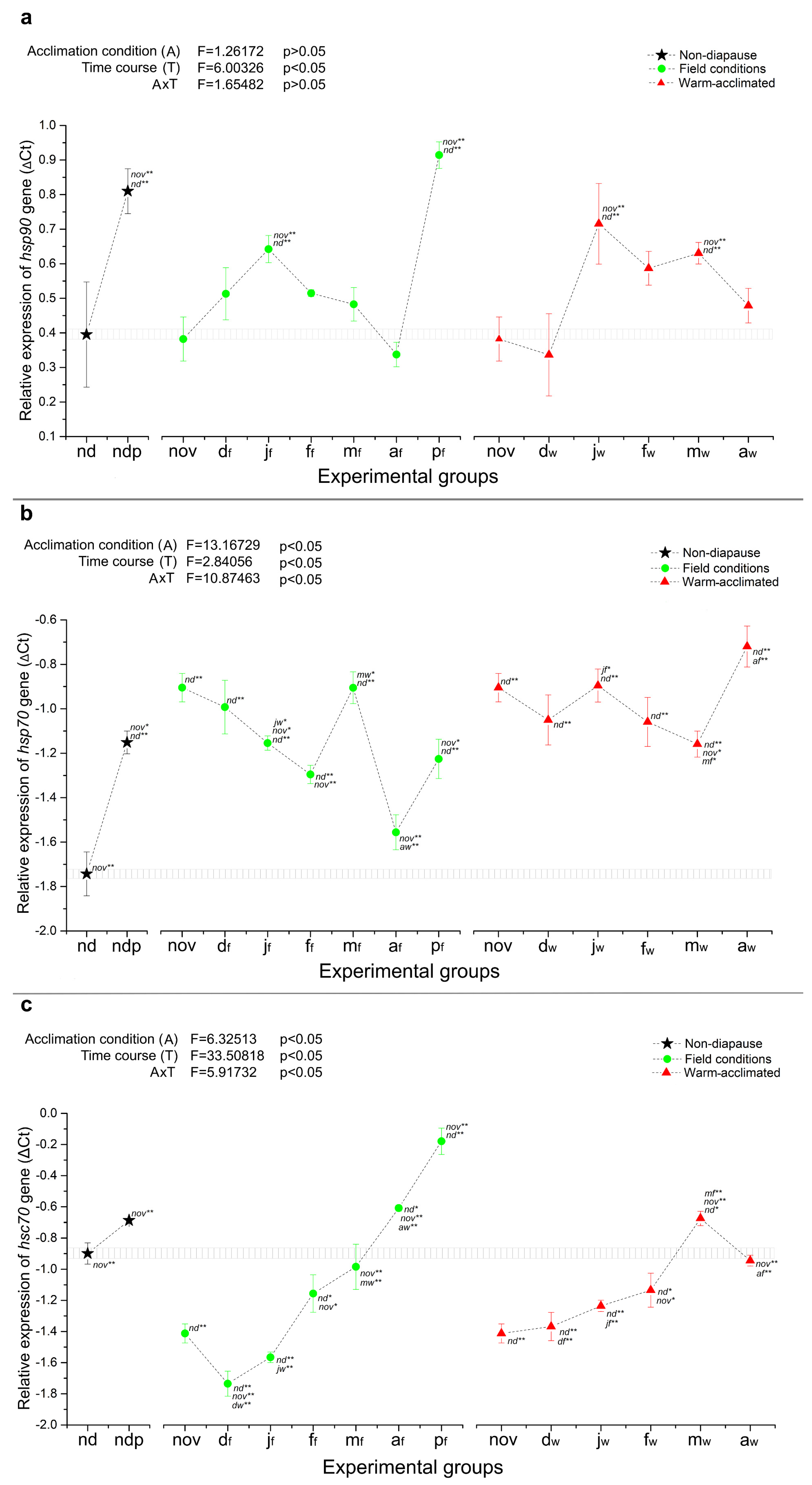
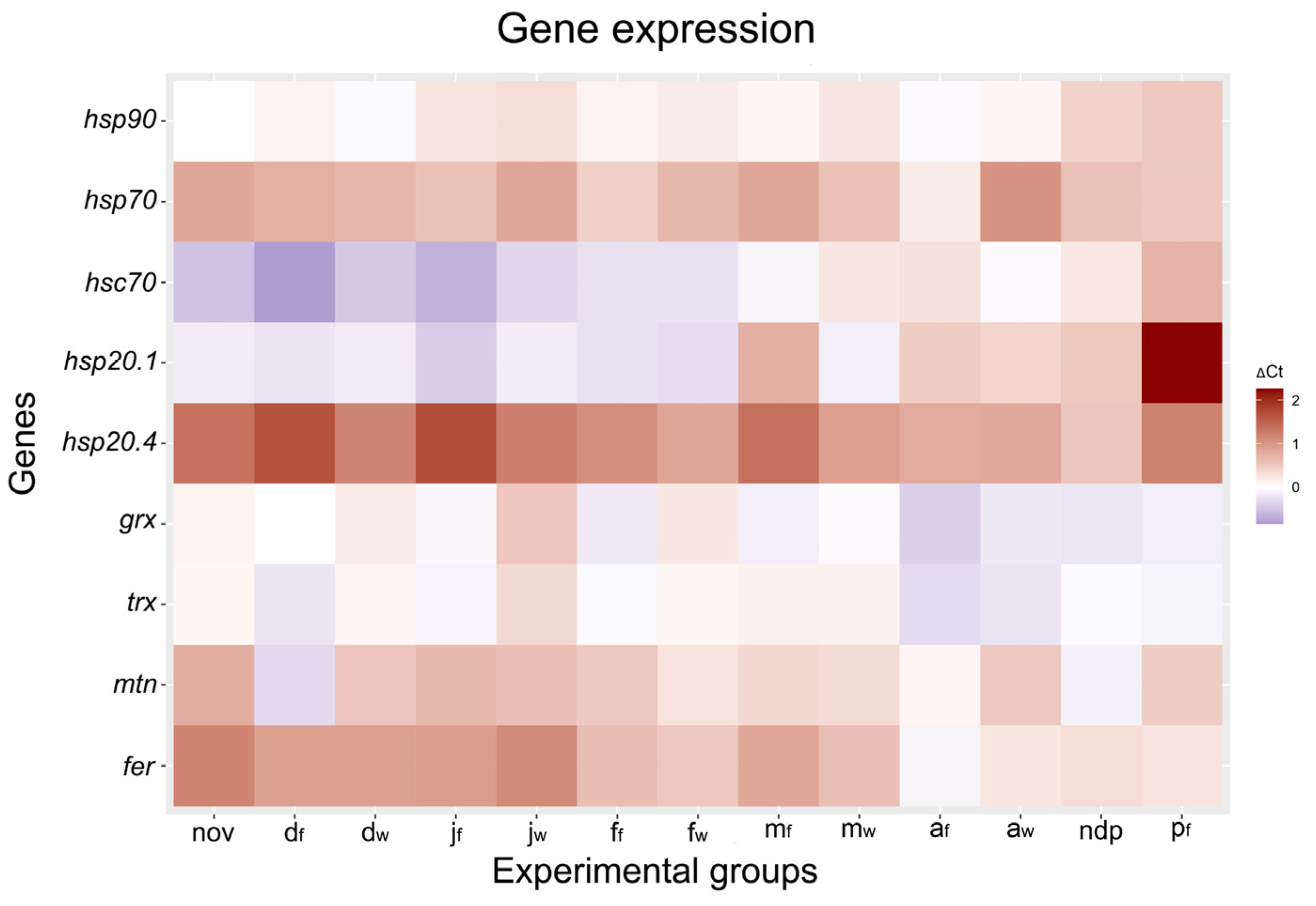
2.2. Heat Shock Protein Genes
2.3. Principal Component Analysis (PCA)
3. Discussion
3.1. Expression of ADS-Related Genes
3.1.1. Thioredoxin and Glutaredoxin
3.1.2. Metallothionein and Ferritin
3.2. Expression of HSP Genes
3.2.1. ATP-Dependent HSPs—HSC70, HSP70 and HSP90
3.2.2. ATP-Independent HSPs—HSP20.1 and HSP20.4
4. Materials and Methods
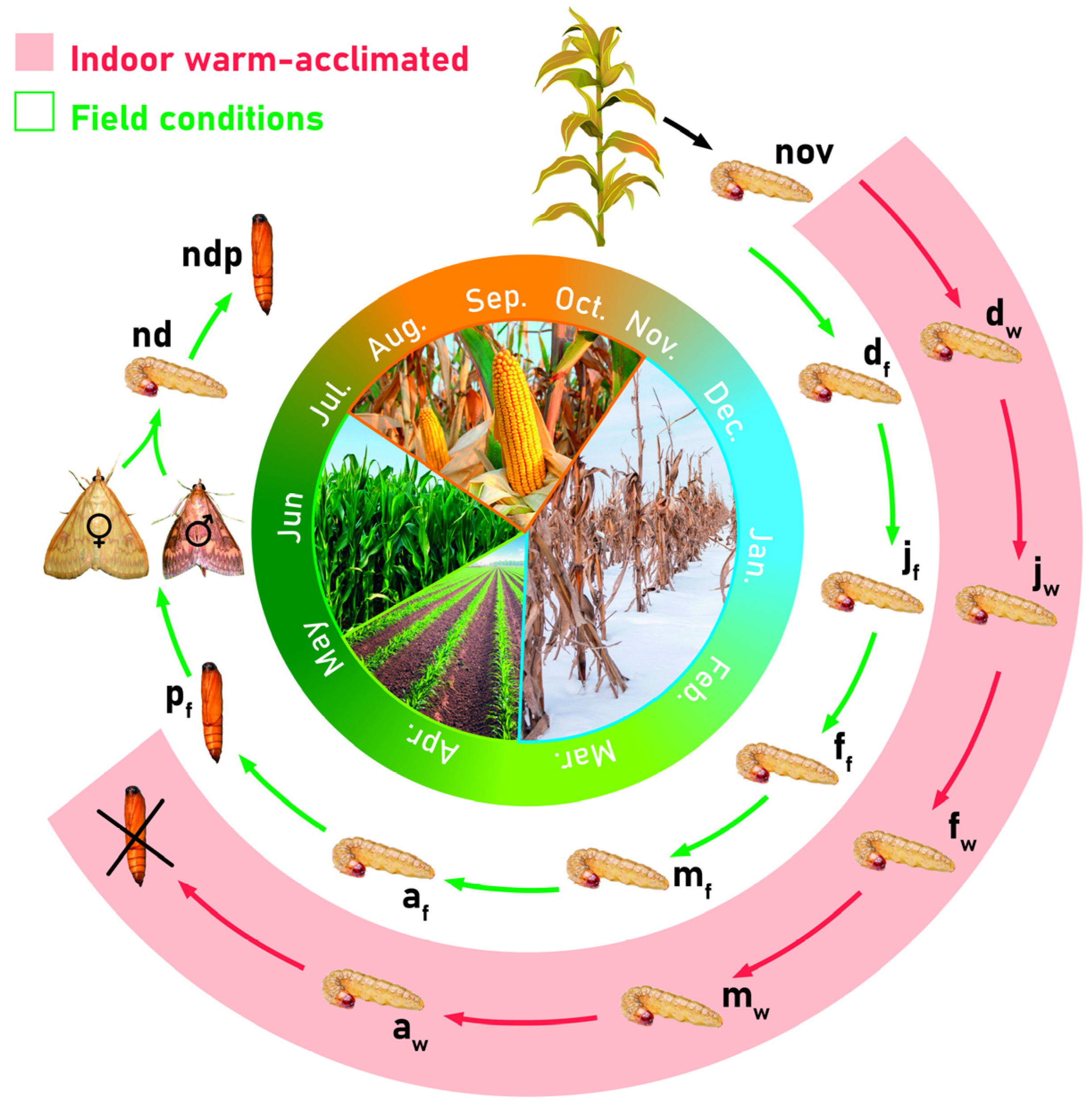
4.1. Experimental Design
4.2. Total RNA Extraction, cDNA Synthesis and qRT-PCR
4.3. Relative Expression and Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; p. 184. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef]
- Beck, S.D. Thermoperiodism. In Insects at Low Temperature; Lee, R.E., Jr., Denlinger, D.L., Eds.; Springer: Boston, MA, USA, 1991; pp. 199–228. [Google Scholar] [CrossRef]
- Beck, S.D. Photoperiodic induction of diapause in an insect. Biol. Bull. 1962, 122, 1–12. [Google Scholar] [CrossRef]
- Grubor-Lajsic, G.; Block, W.; Worland, R. Comparison of the cold hardiness of two larval Lepidoptera (Noctuidae). Physiol. Entomol. 1992, 17, 148–152. [Google Scholar] [CrossRef]
- Tomcala, A.; Tollarová, M.; Overgaard, J.; Simek, P.; Kostal, V. Seasonal acquisition of chill tolerance and restructuring of membrane glycerophospholipids in an overwintering insect: Triggering by low temperature, desiccation and diapause progression. J. Exp. Biol. 2006, 209, 4102–4114. [Google Scholar] [CrossRef] [PubMed]
- McWatters, H.G.; Saunders, D.S. Maternal temperature has different effects on the photoperiodic response and duration of larval diapause in blow fly (Calliphora vicina) strains collected at two latitudes. Physiol. Entomol. 1998, 23, 369–375. [Google Scholar] [CrossRef]
- Mutchmor, J.A.; Beckel, W.E. Some factors affecting diapause in the European corn borer, Ostrinia nubilalis (Hbn.) (Lepidoptera: Pyralidae). Can. J. Zool. 1959, 37, 161–168. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Kagkou, E.A. Effect of thermoperiod on diapause induction of Sesamia nonagrioides (Lepidoptera-Noctuidae). Environ. Entomol. 2000, 29, 489–494. [Google Scholar] [CrossRef]
- Kovac, H.; Käfer, H.; Petrocelli, I.; Amstrup, A.B.; Stabentheiner, A. The impact of climate on the energetics of overwintering paper wasp gynes (Polistes dominula and Polistes gallicus). Insects 2023, 14, 849. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- MacRae, T.H. Gene expression, metabolic regulation and stress tolerance during diapause. Cell. Mol. Life Sci. 2010, 67, 2405–2424. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Insect cold hardiness: Metabolic, gene, and protein adaptation. Can. J. Zool. 2012, 90, 456–475. [Google Scholar] [CrossRef]
- Carpene, E.; Andreani, G.; Isani, G. Metallothionein functions and structural characteristics. J. Trace Elem. Med. Biol. 2007, 21, 35–39. [Google Scholar] [CrossRef]
- Formigari, A.; Alberton, P.; Cantale, V.; Nadal, V.D.; Feltrin, M.; Ferronato, S.; Santon, A.; Schiavon, L.; Irato, P. Relationship between metal transcription Factor-1 and Zinc in resistance to metals producing free radicals. Curr. Chem. Biol. 2008, 2, 256–266. [Google Scholar] [CrossRef]
- Egli, D.; Domènech, J.; Selvaraj, A.; Balamurugan, K.; Hua, H.; Capdevila, M.; Georgiev, O.; Schaffner, W.; Atrian, S. The four members of the Drosophila metallothionein family exhibit distinct yet overlapping roles in heavy metal homeostasis and detoxification. Genes Cells 2006, 11, 647–658. [Google Scholar] [CrossRef]
- Atanesyan, L.; Günther, V.; Celniker, S.E.; Georgiev, O.; Schaffner, W. Characterization of MtnE, the fifth metallothionein member in Drosophila. JBIC J. Biol. Inorg. Chem. 2011, 16, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef]
- Makarow, M.; Sistonen, L. A point worth stressing. Stress-inducible Cellular Responses. Trends Cell Biol. 1997, 7, 376–377. [Google Scholar] [CrossRef]
- Kregel, K.C. Invited review: Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186. [Google Scholar] [CrossRef]
- Zhang, G.; Storey, J.M.; Storey, K.B. Chaperone proteins and winter survival by a freeze tolerant insect. J. Insect Physiol. 2011, 57, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Yocum, G.D.; Childers, A.K.; Rinehart, J.P.; Rajamohan, A.; Pitts-Singer, T.L.; Greenlee, K.J.; Bowsher, J.H. Environmental history impacts gene expression during diapause development in the alfalfa leafcutting bee, Megachile rotundata. J. Exp. Biol. 2018, 221, jeb173443. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.G.; Yoshida, K.M.; Kimura, M.T. Accumulation of Hsp70 mRNA under environmental stresses in diapausing and nondiapausing adults of Drosophila triauraria. J. Insect Physiol. 1998, 44, 1009–1015. [Google Scholar] [CrossRef]
- Nordin, J.H.; Cui, Z.; Yin, C.M. Cold-induced glycerol accumulation by Ostrinia nubilalis larvae is developmentally regulated. J. Insect Physiol. 1984, 30, 563–566. [Google Scholar] [CrossRef]
- Grubor-Lajsic, G.; Block, W.; Palanacki, V.; Glumac, S. Cold hardiness parameters of overwintering diapause larvae of Ostrinia nubilalis in Vojvodina, Yugoslavia. Cryo-Lett. 1991, 12, 177–182. [Google Scholar]
- Stanic, B.; Jovanovic-Galovic, A.; Blagojevic, D.P.; Grubor-Lajsic, G.; Worland, R.; Spasic, M.B. Cold hardiness in Ostrinia nubilalis (Lepidoptera: Pyralidae): Glycerol content, hexose monophosphate shunt activity, and antioxidative defense system. Eur. J. Entomol. 2004, 101, 459–466. [Google Scholar] [CrossRef]
- Vukašinović, E.L.; Pond, D.W.; Worland, M.R.; Kojić, D.; Purać, J.; Blagojević, D.P.; Grubor-Lajšić, G. Diapause induces changes in the composition and biophysical properties of lipids in larvae of the European corn borer, Ostrinia nubilalis (Lepidoptera: Crambidae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 165, 219–225. [Google Scholar] [CrossRef]
- Vukašinović, E.L.; Pond, D.W.; Worland, M.R.; Kojić, D.; Purać, J.; Popović, Ž.D.; Grubor-Lajšić, G. Diapause induces remodeling of the fatty acid composition of membrane and storage lipids in overwintering larvae of Ostrinia nubilalis, Hubn. (Lepidoptera: Crambidae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 184, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Popović, Ž.D.; Subotić, A.; Nikolić, T.V.; Radojičić, R.; Blagojević, D.P.; Grubor-Lajšić, G.; Koštál, V. Expression of stress-related genes in diapause of European corn borer (Ostrinia nubilalis Hbn.). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 186, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Uzelac, I.; Avramov, M.; Čelić, T.; Vukašinović, E.; Gošić-Dondo, S.; Purać, J.; Kojić, D.; Blagojević, D.; Popović, Ž.D. Effect of cold acclimation on selected metabolic enzymes during diapause in the European corn borer Ostrinia nubilalis (Hbn.). Sci. Rep. 2020, 10, 9085. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, K.S.; Kim, B.Y.; Choo, Y.M.; Sohn, H.D.; Jin, B.R. Thioredoxin from the silkworm, Bombyx mori: cDNA sequence, expression, and functional characterization. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 574–581. [Google Scholar] [CrossRef]
- Choi, Y.S.; Lee, K.S.; Yoon, H.J.; Kim, I. A bumblebee thioredoxin-like protein gene that is up-regulated by a temperature stimulus and lipopolysaccharide injection. Eur. J. Entomol. 2006, 103, 291. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Z.; Nian, X.; Wu, F.; Shen, Z.; Zhang, B.; Zhang, Q.; Liu, X. Sequence analysis, expression profiles and function of thioredoxin 2 and thioredoxin reductase 1 in resistance to nucleopolyhedrovirus in Helicoverpa armigera. Sci. Rep. 2015, 5, 15531. [Google Scholar] [CrossRef]
- Shen, Z.J.; Liu, Y.J.; Gao, X.H.; Liu, X.M.; Zhang, S.D.; Li, Z.; Zhang, Q.W.; Liu, X.X. Molecular identification of two thioredoxin genes from Grapholita molesta and their function in resistance to emamectin benzoate. Front. Physiol. 2018, 9, 1421. [Google Scholar] [CrossRef]
- Zhang, S.D.; Shen, Z.J.; Liu, X.M.; Li, Z.; Zhang, Q.W.; Liu, X.X. Molecular identification of three novel glutaredoxin genes that play important roles in antioxidant defense in Helicoverpa armigera. Insect Biochem. Mol. Biol. 2016, 75, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Kojic, D.; Spasojevic, I.; Mojovic, M.; Blagojevic, D.; Worland, M.R.; Grubor-Lajsic, G.; Spasic, M.B. Potential role of hydrogen peroxide and melanin in the cold hardiness of Ostrinia nubilalis (Lepidoptera: Pyralidae). Eur. J. Entomol. 2009, 106, 451. [Google Scholar] [CrossRef]
- Kojić, D.; Purać, J.; Popović, Ž.D.; Pamer, E.; Grubor-Lajšić, G. Importance of the body water management for winter cold survival of the European corn borer Ostrinia nubilalis Hübn. (Lepidoptera: Pyralidae). Biotechnol. Biotechnol. Equip. 2010, 24 (Suppl. S1), 648–654. [Google Scholar] [CrossRef]
- Popović, Ž.D.; Maier, V.; Avramov, M.; Uzelac, I.; Gošić-Dondo, S.; Blagojević, D.; Koštál, V. Acclimations to cold and warm conditions differently affect the energy metabolism of diapausing larvae of the European corn borer Ostrinia nubilalis (Hbn.). Front. Physiol. 2021, 12, 2126. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Zhang, Y.; Wang, T.; Luo, M.; Li, C. Molecular characterization of glutaredoxin 2 from Ostrinia furnacalis. Integr. Zool. 2013, 8, 30–38. [Google Scholar] [CrossRef]
- Yao, P.; Chen, X.; Yan, Y.; Liu, F.; Zhang, Y.; Guo, X.; Xu, B. Glutaredoxin 1, glutaredoxin 2, thioredoxin 1, and thioredoxin peroxidase 3 play important roles in antioxidant defense in Apis cerana cerana. Free Radic. Biol. Med. 2014, 68, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, F.; Shen, Z.; Moural, T.W.; Liu, L.; Li, Z.; Liu, X.; Xu, H. Glutaredoxins and thioredoxin peroxidase involved in defense of emamectin benzoate induced oxidative stress in Grapholita molesta. Pestic. Biochem. Physiol. 2021, 176, 104881. [Google Scholar] [CrossRef]
- Yao, P.; Hao, L.; Wang, F.; Chen, X.; Yan, Y.; Guo, X.; Xu, B. Molecular cloning, expression and antioxidant characterisation of a typical thioredoxin gene (AccTrx2) in Apis cerana cerana. Gene 2013, 527, 33–41. [Google Scholar] [CrossRef]
- Cornette, R.; Kikawada, T. The induction of anhydrobiosis in the sleeping chironomid: Current status of our knowledge. IUBMB Life 2011, 63, 419–429. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Molecular biology of freezing tolerance. Compr. Physiol. 2013, 3, 1283–1308. [Google Scholar] [CrossRef]
- Nesmelov, A.A.; Devatiyarov, R.M.; Voronina, T.A.; Kondratyeva, S.A.; Cherkasov, A.V.; Cornette, R.; Kikawada, T.; Shagimardanova, E.I. New antioxidant genes from an anhydrobiotic insect: Unique structural features in functional motifs of thioredoxins. BioNanoScience 2016, 6, 568–570. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Robich, R.M.; Denlinger, D.L. Isolation of diapause-regulated genes from the flesh fly, Sarcophaga crassipalpis by suppressive subtractive hybridization. J. Insect Physiol. 2010, 56, 603–609. [Google Scholar] [CrossRef]
- Storey, K.B. Strategies for exploration of freeze responsive gene expression: Advances in vertebrate freeze tolerance. Cryobiology 2004, 48, 134–145. [Google Scholar] [CrossRef]
- Vukašinović, E.L.; Čelić, T.V.; Kojić, D.; Franeta, F.; Milić, S.; Ninkov, J.; Blagojević, D.; Purać, J. The effect of long term exposure to cadmium on Ostrinia nubilalis growth, development, survival rate and oxidative status. Chemosphere 2020, 243, 125375. [Google Scholar] [CrossRef]
- Purać, J.; Čelić, T.V.; Vukašinović, E.L.; Đorđievski, S.; Milić, S.; Ninkov, J.; Kojić, D. Identification of a metallothionein gene and the role of biological thiols in stress induced by short-term Cd exposure in Ostrinia nubilalis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 250, 109148. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, Z.; Gao, Z.; Wen, Y.; Wang, W.; Liu, W.; Wang, L.; Zhu, F. Identification of three metallothioneins in the black soldier fly and their functions in Cd accumulation and detoxification. Environ. Pollut. 2021, 286, 117146. [Google Scholar] [CrossRef]
- Tarrant, A.M.; Baumgartner, M.F.; Verslycke, T.; Johnson, C.L. Differential gene expression in diapausing and active Calanus finmarchicus (Copepoda). Mar. Ecol. Prog. Ser. 2008, 355, 193–207. [Google Scholar] [CrossRef]
- Wolschin, F.; Gadau, J. Deciphering proteomic signatures of early diapause in Nasonia. PLoS ONE 2009, 4, e6394. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Villeneuve, T.S.; Garant, K.A.; Amons, R.; MacRae, T.H. Functional characterization of artemin, a ferritin homolog synthesized in Artemia embryos during encystment and diapause. FEBS J. 2007, 274, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Xu, W.H. Identification of gene expression changes associated with the initiation of diapause in the brain of the cotton bollworm, Helicoverpa armigera. BMC Genom. 2011, 12, 1–14. [Google Scholar] [CrossRef]
- Larade, K.; Storey, K.B. Accumulation and translation of ferritin heavy chain transcripts following anoxia exposure in a marine invertebrate. J. Exp. Biol. 2004, 207, 1353–1360. [Google Scholar] [CrossRef]
- Clark, M.S.; Thorne, M.A.; Purać, J.; Burns, G.; Hillyard, G.; Popović, Ž.D.; Gubor-Lajšić, G.; Worland, M.R. Surviving the cold: Molecular analyses of insect cryoprotective dehydration in the Arctic springtail Megaphorura arctica (Tullberg). BMC Genom. 2009, 10, 1–19. [Google Scholar] [CrossRef]
- Schokraie, E.; Warnken, U.; Hotz-Wagenblatt, A.; Grohme, M.A.; Hengherr, S.; Förster, F.; Schill, R.O.; Frohme, M.; Dandekar, T.; Schnölzer, M. Comparative proteome analysis of Milnesium tardigradum in early embryonic state versus adults in active and anhydrobiotic state. PLoS ONE 2012, 7, e45682. [Google Scholar] [CrossRef]
- Danks, H.V. Dehydration in dormant insects. J. Insect Physiol. 2000, 46, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L. Diapause. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2009; pp. 267–271. [Google Scholar] [CrossRef]
- Popović, Ž.D. Molecular and Biochemical Basis of Diapause of European Corn Borer Ostrinia nubilalis (Hbn.) (Lepidoptera: Pyralidae). Ph.D. Dissertation, University of Belgrade, Belgrade, Serbia, 2014. [Google Scholar]
- Hamer, D.H. Metallothionein. Annu. Rev. Biochem. 1986, 55, 913–951. [Google Scholar] [CrossRef]
- Luo, M.; Finet, C.; Cong, H.; Wei, H.Y.; Chung, H. The evolution of insect metallothioneins. Proc. R. Soc. B Biol. Sci. 2020, 287, 20202189. [Google Scholar] [CrossRef]
- Pham, D.Q.; Winzerling, J.J. Insect ferritins: Typical or atypical? Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 824–833. [Google Scholar] [CrossRef]
- Etges, W.J.; De Oliveira, C.C.; Rajpurohit, S.; Gibbs, A.G. Effects of temperature on transcriptome and cuticular hydrocarbon expression in ecologically differentiated populations of desert Drosophila. Ecol. Evol. 2017, 7, 619–637. [Google Scholar] [CrossRef]
- Mahadav, A.; Kontsedalov, S.; Czosnek, H.; Ghanim, M. Thermotolerance and gene expression following heat stress in the whitefly Bemisia tabaci B and Q biotypes. Insect Biochem. Mol. Biol. 2009, 39, 668–676. [Google Scholar] [CrossRef]
- Tungjitwitayakul, J.; Tatun, N.; Singtripop, T.; Sakurai, S. Characteristic expression of three heat shock-responsive genes during larval diapause in the bamboo borer Omphisa fuscidentalis. Zool. Sci. 2008, 25, 321–333. [Google Scholar] [CrossRef]
- Sonoda, S.; Fukumoto, K.; Izumi, Y.; Yoshida, H.; Tsumuki, H. Cloning of heat shock protein genes (hsp90 and hsc70) and their expression during larval diapause and cold tolerance acquisition in the rice stem borer, Chilo suppressalis Walker. Arch. Insect Biochem. Physiol. 2006, 63, 36–47. [Google Scholar] [CrossRef]
- Gkouvitsas, T.; Kontogiannatos, D.; Kourti, A. Cognate Hsp70 gene is induced during deep larval diapause in the moth Sesamia nonagrioides. Insect Mol. Biol. 2009, 18, 253–264. [Google Scholar] [CrossRef]
- Gkouvitsas, T.; Kontogiannatos, D.; Kourti, A. Expression of the Hsp83 gene in response to diapause and thermal stress in the moth Sesamia nonagrioides. Insect Mol. Biol. 2009, 18, 759–768. [Google Scholar] [CrossRef]
- Gkouvitsas, T.; Kontogiannatos, D.; Kourti, A. Differential expression of two small Hsps during diapause in the corn stalk borer Sesamia nonagrioides (Lef.). J. Insect Physiol. 2008, 54, 1503–1510. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Yocum, G.D.; Denlinger, D.L. Developmental upregulation of inducible hsp70 transcripts, but not the cognate form, during pupal diapause in the flesh fly, Sarcophaga crassipalpis. Insect Biochem. Mol. Biol. 2000, 30, 515–521. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Li, A.; Yocum, G.D.; Robich, R.M.; Hayward, S.A.; Denlinger, D.L. Up-regulation of heat shock proteins is essential for cold survival during insect diapause. Proc. Natl. Acad. Sci. USA 2007, 104, 11130–11137. [Google Scholar] [CrossRef]
- Yocum, G.D.; Kemp, W.P.; Bosch, J.; Knoblett, J.N. Temporal variation in overwintering gene expression and respiration in the solitary bee Megachile rotundata. J. Insect Physiol. 2005, 51, 621–629. [Google Scholar] [CrossRef]
- Zhang, Q.; Denlinger, D.L. Molecular characterization of heat shock protein 90, 70 and 70 cognate cDNAs and their expression patterns during thermal stress and pupal diapause in the corn earworm. J. Insect Physiol. 2010, 56, 138–150. [Google Scholar] [CrossRef]
- Cheng, W.; Li, D.; Wang, Y.; Liu, Y.; Zhu-Salzman, K. Cloning of heat shock protein genes (hsp70, hsc70 and hsp90) and their expression in response to larval diapause and thermal stress in the wheat blossom midge, Sitodiplosis mosellana. J. Insect Physiol. 2016, 95, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.K.; Lee, K.Y. Molecular characterization of heat shock protein 70 cognate cDNA and its upregulation after diapause termination in Lycorma delicatula eggs. J. Asia-Pac. Entomol. 2015, 18, 709–714. [Google Scholar] [CrossRef]
- Li, A.; Denlinger, D.L. Pupal cuticle protein is abundant during early adult diapause in the mosquito Culex pipiens. J. Med. Entomol. 2009, 46, 1382–1386. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Shim, J.K.; Kim, D.W.; Kwon, Y.J.; Lee, K.Y. Tissue-specific variation of heat shock protein gene expression in relation to diapause in the bumblebee Bombus terrestris. Entomol. Res. 2008, 38, 10–16. [Google Scholar] [CrossRef]
- Clare, D.K.; Saibil, H.R. ATP-driven molecular chaperone machines. Biopolymers 2013, 99, 846–859. [Google Scholar] [CrossRef]
- Thomas, S.R.; Lengyel, J.A. Ecdysteroid-regulated heat-shock gene expression during Drosophila melanogaster development. Dev. Biol. 1986, 115, 434–438. [Google Scholar] [CrossRef]
- Goto, S.G.; Kimura, M.T. Heat-shock-responsive genes are not involved in the adult diapause of Drosophila triauraria. Gene 2004, 326, 117–122. [Google Scholar] [CrossRef]
- Tachibana, S.I.; Numata, H.; Goto, S.G. Gene expression of heat-shock proteins (Hsp23, Hsp70 and Hsp90) during and after larval diapause in the blow fly Lucilia sericata. J. Insect Physiol. 2005, 51, 641–647. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Denlinger, D.L. Heat-shock protein 90 is down-regulated during pupal diapause in the flesh fly, Sarcophaga crassipalpis, but remains responsive to thermal stress. Insect Mol. Biol. 2000, 9, 641–645. [Google Scholar] [CrossRef]
- Hayward, S.A.L.; Pavlides, S.C.; Tammariello, S.P.; Rinehart, J.P.; Denlinger, D.L. Temporal expression patterns of diapause-associated genes in flesh fly pupae from the onset of diapause through post-diapause quiescence. J. Insect Physiol. 2005, 51, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, W.; Wang, X.; Niu, C.; Lei, C. Analysis of pupal head proteome and its alteration in diapausing pupae of Helicoverpa armigera. J. Insect Physiol. 2010, 56, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Kayukawa, T.; Monteiro, A.; Ishikawa, Y. The expression of the HSP90 gene in response to winter and summer diapauses and thermal-stress in the onion maggot, Delia antiqua. Insect Mol. Biol. 2005, 14, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhang, S.; Wang, H.; Liang, L.; Liu, Z.; Wang, Y.; Baohua, X.; Zhao, H. Differential expression characterisation of the heat shock proteins DnaJB6, DnaJshv, DnaJB13, and DnaJB14 in Apis cerana cerana under various stress conditions. Front. Ecol. Evol. 2022, 10, 873791. [Google Scholar] [CrossRef]
- Yocum, G.D.; Joplin, K.H.; Denlinger, D.L. Expression of heat shock proteins in response to high and low temperature extremes in diapausing pharate larvae of the gypsy moth, Lymantria dispar. Arch. Insect Biochem. Physiol. 1991, 18, 239–249. [Google Scholar] [CrossRef]
- Wu, Y.K.; Zou, C.; Fu, D.M.; Zhang, W.N.; Xiao, H.J. Molecular characterization of three Hsp90 from Pieris and expression patterns in response to cold and thermal stress in summer and winter diapause of Pieris melete. Insect Sci. 2018, 25, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Fremdt, H.; Amendt, J.; Zehner, R. Diapause-specific gene expression in Calliphora vicina (Diptera: Calliphoridae)—A useful diagnostic tool for forensic entomology. Int. J. Leg. Med. 2014, 128, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Kayukawa, T.; Monteiro, A.; Ishikawa, Y. Cloning and characterization of the HSP70 gene, and its expression in response to diapauses and thermal stress in the onion maggot, Delia antiqua. J. Biochem. Mol. Biol. 2006, 39, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xu, X.L.; Li, Y.P.; Wu, J.X. Characterization of heat shock protein 90, 70 and their transcriptional expression patterns on high temperature in adult of Grapholita molesta (Busck). Insect Sci. 2014, 21, 439–448. [Google Scholar] [CrossRef]
- Luo, S.; Ahola, V.; Shu, C.; Xu, C.; Wang, R. Heat shock protein 70 gene family in the Glanville fritillary butterfly and their response to thermal stress. Gene 2015, 556, 132–141. [Google Scholar] [CrossRef]
- Yocum, G.D. Differential expression of two HSP70 transcripts in response to cold shock, thermoperiod, and adult diapause in the Colorado potato beetle. J. Insect Physiol. 2001, 47, 1139–1145. [Google Scholar] [CrossRef]
- Lebenzon, J.E.; Torson, A.S.; Sinclair, B.J. Diapause differentially modulates the transcriptomes of fat body and flight muscle in the Colorado potato beetle. Comp. Biochem. Physiol. D Genom. Proteom. 2021, 40, 100906. [Google Scholar] [CrossRef]
- Yocum, G.D.; Joplin, K.H.; Denlinger, D.L. Upregulation of a 23 kDa small heat shock protein transcript during pupal diapause in the flesh fly, Sarcophaga crassipalpis. Insect Biochem. Mol. Biol. 1998, 28, 677–682. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, Q.; Zhang, G.; Zhu-Salzman, K.; Cheng, W. Characterization of two small heat shock protein genes (Hsp17.4 and Hs20.3) from Sitodiplosis mosellana, and their expression regulation during diapause. Insects 2021, 12, 119. [Google Scholar] [CrossRef]
- Miano, F.N.; Jiang, T.; Zhang, J.; Zhang, W.N.; Peng, Y.; Xiao, H.J. Identification and up-regulation of three small heat shock proteins in summer and winter diapause in response to temperature stress in Pieris melete. Int. J. Biol. Macromol. 2022, 209, 1144–1154. [Google Scholar] [CrossRef]
- Atungulu, E.; Tanaka, H.; Fujita, K.; Yamamoto, K.I.; Sakata, M.; Sato, E.; Hara, M.; Yamashita, T.; Suzuki, K. A double chaperone function of the sHsp genes against heat-based environmental adversity in the soil-dwelling leaf beetles. J. Insect Biotechnol. Sericol. 2006, 75, 15–22. [Google Scholar] [CrossRef]
- Sakano, D.; Li, B.; Xia, Q.; Yamamoto, K.; Fujii, H.; Aso, Y. Genes encoding small heat shock proteins of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2006, 70, 2443–2450. [Google Scholar] [CrossRef]
- Saravanakumar, R.; Ponnuvel, K.; Qadri, S. Expression of metabolic enzyme genes and heat-shock protein genes during embryonic development in diapause and non-diapause egg of multivoltine silkworm Bombyx mori. Biologia 2008, 63, 737–744. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ganger, M.T.; Dietz, G.D.; Ewing, S.J. A common base method for analysis of qPCR data and the application of simple blocking in qPCR experiments. BMC Bioinform. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Amount (g) | Component | Amount (g) |
|---|---|---|---|
| Wheat kernels | 160.0 | Citric acid | 5.0 |
| Barley kernels | 25.0 | Vitamin C | 5.0 |
| Brewer’s yeast | 25.0 | Methylparaben | 1.7 |
| Agar | 20.0 | Acetic acid | 1.4 |
| Formaldehyde 37% | 6.0 | Vitamin B complex | 0.07 |
| Component | Volume (μL) |
|---|---|
| 2X Power SYBR™ Green PCR Master Mix | 7.0 |
| F primer (10 μM) | 0.7 |
| R primer (10 μM) | 0.7 |
| DEPC-treated H2O | 0.6 |
| cDNA mixture (~12.5 ng/μL) | 5.0 |
| Total per reaction | 14.0 |
| Gene | Primer Sequence (5′-3′) | Tm (°C) | Accession No. |
|---|---|---|---|
| Actin (reference) | F: CAGAAGGAAATCACAGCTCTAGCC R: ATCGTACTCCTGTTTCGAGATCCA | 63.33 62.95 | EL928709.1 |
| Glutaredoxin (grx) | F: TCGGCAAGGTCAAACAACCA R: GCCACCTCCTACACAGTTCC | 57.50 57.60 | EL930102.1 |
| Thioredoxin (trx) | F: GGGTTTTGATGACACTGACGC R: ACGCTTCTACGGTGACAACA | 56.70 56.50 | EL929289.1 |
| Ferritin (fer) | F: GGCGCTCACTTCTCTAAGGATACTG R: GATTAGTGAGGTGACGTCAGAGGTG | 63.82 63.46 | EL929400.1 |
| Metallothionein (mtn) | F: AAAGAGACACAGCTCCTCCAAATTC R: ATTGAGACACAGCCACTTCATCTTC | 63.31 63.04 | EL929052.1 |
| Heat shock protein 90 (hsp90) | F: CAAGATCGTTCTTCACATCAAGGAG R: CGTCCTCTTTCTTCTCTTCTTCAGC | 63.43 63.31 | EL929806.1 |
| Heat shock protein 70 (hsp70) | F: GCACAGGCCGCAGCAAGAAC R: AGGGCTTGTCGCACGCTGAA | 65.14 65.12 | XM_028309302 |
| Heat shock protein cognate 70 (hsc70) | F: GAGGCGGAAGATTACAAGAAACAAA R: GAGATCACATTTTGCTTCAATCACG | 63.78 64.08 | EL928755.1 |
| Heat shock protein 20.1 (hsp20.1) | F: CAGCGCTAAAGAATGAAAGGTCTGT R: TAGGTATCTCTCATTTCGCCTGTCC | 64.11 63.90 | AB568468.1 |
| Heat shock protein 20.4 (hsp20.4) | F: CGAAGAAAGTATCAGACGTGTCCAA R: TAAATGCAACGCATCACGAGATTAC | 63.76 64.27 | AB568467.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uzelac, I.; Avramov, M.; Knežić, T.; Tatić, V.; Gošić-Dondo, S.; Popović, Ž.D. Prolonged Heat Stress during Winter Diapause Alters the Expression of Stress-Response Genes in Ostrinia nubilalis (Hbn.). Int. J. Mol. Sci. 2024, 25, 3100. https://doi.org/10.3390/ijms25063100
Uzelac I, Avramov M, Knežić T, Tatić V, Gošić-Dondo S, Popović ŽD. Prolonged Heat Stress during Winter Diapause Alters the Expression of Stress-Response Genes in Ostrinia nubilalis (Hbn.). International Journal of Molecular Sciences. 2024; 25(6):3100. https://doi.org/10.3390/ijms25063100
Chicago/Turabian StyleUzelac, Iva, Miloš Avramov, Teodora Knežić, Vanja Tatić, Snežana Gošić-Dondo, and Željko D. Popović. 2024. "Prolonged Heat Stress during Winter Diapause Alters the Expression of Stress-Response Genes in Ostrinia nubilalis (Hbn.)" International Journal of Molecular Sciences 25, no. 6: 3100. https://doi.org/10.3390/ijms25063100
APA StyleUzelac, I., Avramov, M., Knežić, T., Tatić, V., Gošić-Dondo, S., & Popović, Ž. D. (2024). Prolonged Heat Stress during Winter Diapause Alters the Expression of Stress-Response Genes in Ostrinia nubilalis (Hbn.). International Journal of Molecular Sciences, 25(6), 3100. https://doi.org/10.3390/ijms25063100