
MoNOT3 Subunit Has Important Roles in Infection-Related Development and Stress Responses in Magnaporthe oryzae

 , and
, and
Abstract
:1. Introduction
2. Results
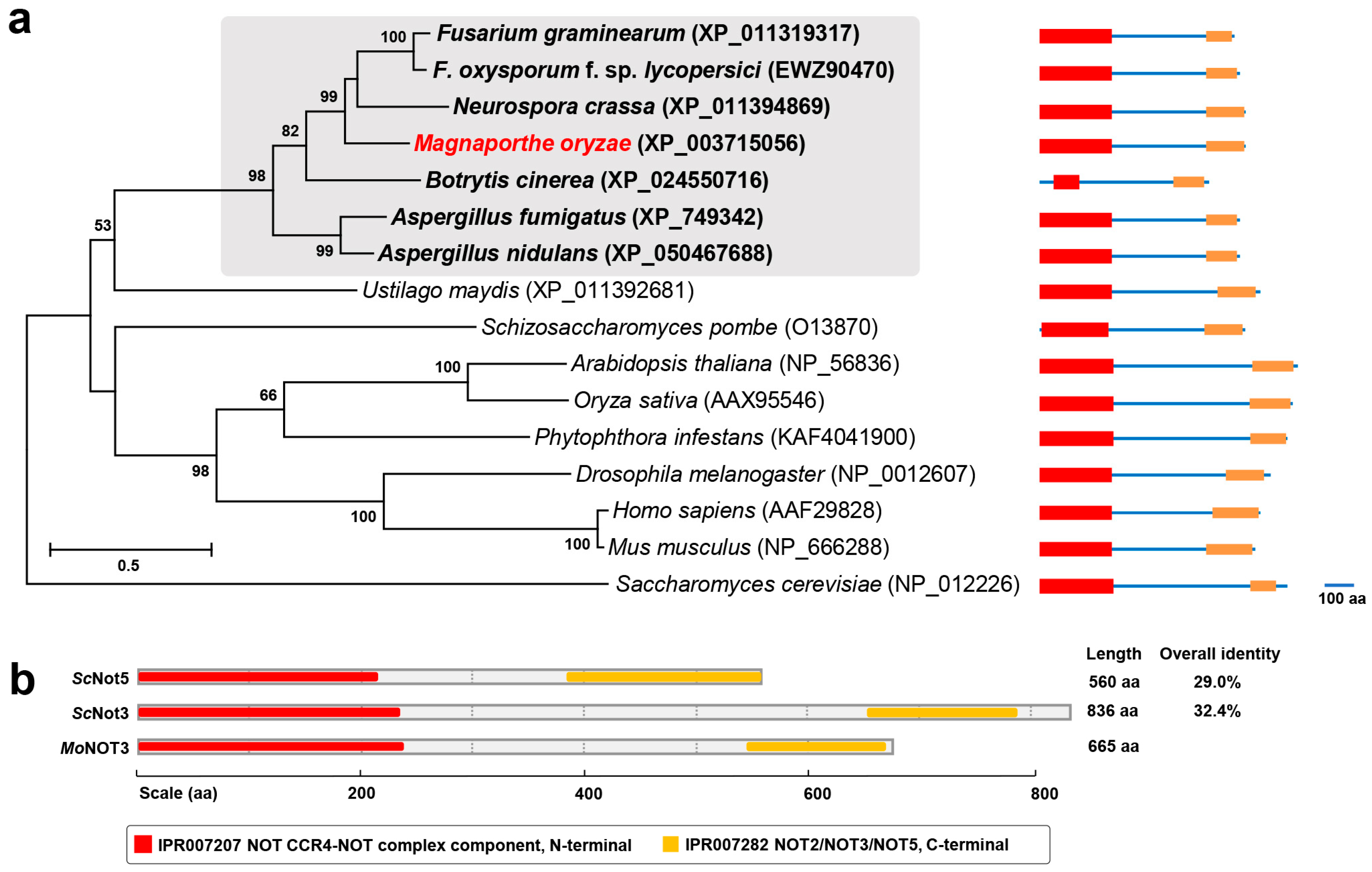
2.1. Molecular Characterisation of MoNOT3
2.2. Targeted Replacement of MoNOT3
2.3. MoNOT3 Is Required for Vegetative Growth and Conidiogenesis
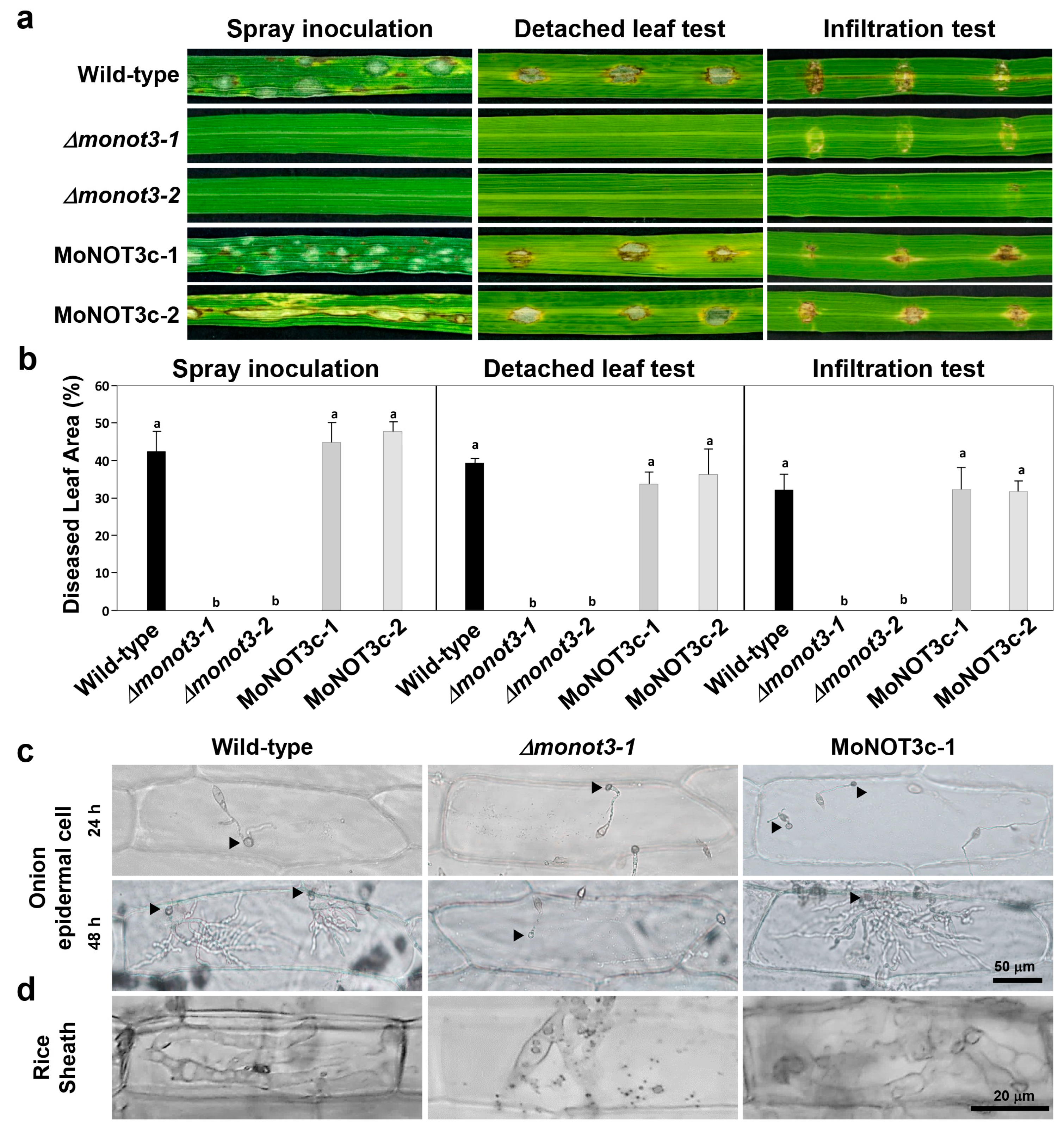
2.4. MoNOT3 Is Required for Conidial Germination and Appressorium Formation
2.5. MoNOT3 Is Required for Penetration and Virulence in M. oryzae
2.6. MoNOT3 Is Important for Stress Tolerance
2.7. Analysis of MoNOT3 Gene Expression Patterns
3. Discussion
4. Materials and Methods
4.1. Fungal Isolate and Culture Conditions
4.2. Computational and Phylogenetic Analyses
4.3. Vector Construction and Fungal Transformation
4.4. Nucleic Acid Manipulation and Southern Blot Analysis
4.5. Conidiation, Conidial Morphology, Conidial Germination, and Appressorium Formation
4.6. Plant Infection Assays
4.7. RNA Isolation, Quantitative Real-Time (qRT)-PCR, and Gene Expression Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iyer, V.; Struhl, K. Mechanism of differential utilization of the his3 TR and TC TATA elements. Mol. Cell. Biol. 1995, 15, 7059–7066. [Google Scholar] [CrossRef]
- Collart, M.A.; Panasenko, O.O. The Ccr4–not complex. Gene 2012, 492, 42–53. [Google Scholar] [CrossRef]
- Collart, M.A.; Struhl, K. NOT1 (CDC39), NOT2 (CDC36), NOT3, and NOT4 encode a global-negative regulator of transcription that differentially affects TATA-element utilization. Genes Dev. 1994, 8, 525–537. [Google Scholar] [CrossRef]
- Miller, J.E.; Reese, J.C. Ccr4-Not complex: The control freak of eukaryotic cells. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 315–333. [Google Scholar] [CrossRef]
- Malvar, T.; Biron, R.W.; Kaback, D.B.; Denis, C.L. The CCR4 protein from Saccharomyces cerevisiae contains a leucine-rich repeat region which is required for its control of ADH2 gene expression. Genetics 1992, 132, 951–962. [Google Scholar] [CrossRef]
- Collart, M.A. The Ccr4-Not complex is a key regulator of eukaryotic gene expression. Wiley Interdiscip. Rev. RNA 2016, 7, 438–454. [Google Scholar] [CrossRef]
- Lau, N.-C.; Kolkman, A.; van Schaik, F.M.A.; Mulder, K.W.; Pijnappel, W.W.M.P.; Heck, A.J.R.; Timmers, H.T.M. Human Ccr4-Not complexes!contain!variable deadenylase subunits. Biochem. J. 2009, 422, 443–453. [Google Scholar] [CrossRef]
- Collart, M.A.; Panasenko, O.O.; Nikolaev, S.I. The Not3/5 subunit of the CCR4-Not complex: A central regulator of gene expression that integrates signals between the cytoplasm and the nucleus in eukaryotic cells. Cell. Signal. 2013, 25, 743–751. [Google Scholar] [CrossRef]
- Cheng, S.; Clancy, C.J.; Checkley, M.A.; Handfield, M.; Hillman, J.D.; Progulske-Fox, A.; Lewin, A.S.; Fidel, P.L.; Nguyen, M.H. Identification of Candida albicans genes induced during thrush offers insight into pathogenesis. Mol. Microbiol. 2003, 48, 1275–1288. [Google Scholar] [CrossRef]
- Panepinto, J.C.; Heinz, E.; Traven, A. The cellular roles of CCR4-NOT in model and pathogenic fungi–implications for fungal virulence. Front. Genet. 2013, 4, 302. [Google Scholar] [CrossRef]
- Dai, Y.; Cao, Z.; Huang, L.; Liu, S.; Shen, Z.; Wang, Y.; Wang, H.; Zhang, H.; Li, D.; Song, F. Ccr4-Not complex subunit Not2 plays critical roles in vegetative growth, conidiation and virulence in watermelon Fusarium wilt pathogen Fusarium oxysporum f. sp. niveum. Front. Microbiol. 2016, 7, 1449. [Google Scholar] [CrossRef]
- Bui, D.-C.; Son, H.; Shin, J.Y.; Kim, J.-C.; Kim, H.; Choi, G.J.; Lee, Y.-W. The FgNot3 subunit of the Ccr4-Not complex regulates vegetative growth, sporulation, and virulence in Fusarium graminearum. PLoS ONE 2016, 11, e0147481. [Google Scholar] [CrossRef]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.-R.; Pan, H. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef]
- Talbot, N.J. Having a blast: Exploring the pathogenicity of Magnaporthe grisea. Trends Microbiol. 1995, 3, 9–16. [Google Scholar] [CrossRef]
- Thines, E.; Weber, R.W.; Talbot, N.J. MAP kinase and protein kinase A–dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 2000, 12, 1703–1718. [Google Scholar]
- Mitchell, T.K.; Dean, R.A. The cAMP-dependent protein kinase catalytic subunit is required for appressorium formation and pathogenesis by the rice blast pathogen Magnaporthe grisea. Plant Cell 1995, 7, 1869–1878. [Google Scholar] [CrossRef]
- Xu, J.R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef]
- Choi, J.; Kim, Y.; Kim, S.; Park, J.; Lee, Y.-H. MoCRZ1, a gene encoding a calcineurin-responsive transcription factor, regulates fungal growth and pathogenicity of Magnaporthe oryzae. Fungal Genet. Biol. 2009, 46, 243–254. [Google Scholar] [CrossRef]
- Kim, S.; Hu, J.; Oh, Y.; Park, J.; Choi, J.; Lee, Y.H.; Dean, R.A.; Mitchell, T.K. Combining ChIP-chip and expression profiling to model the MoCRZ1 mediated circuit for Ca/calcineurin signaling in the rice blast fungus. PLoS Pathog. 2010, 6, e1000909. [Google Scholar] [CrossRef]
- Li, G.; Zhou, X.; Xu, J.-R. Genetic control of infection-related development in Magnaporthe oryzae. Curr. Opin. Microbiol. 2012, 15, 678–684. [Google Scholar] [CrossRef]
- Ebbole, D.J. Magnaporthe as a model for understanding host-pathogen interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456. [Google Scholar] [CrossRef]
- Odenbach, D.; Breth, B.; Thines, E.; Weber, R.W.; Anke, H.; Foster, A.J. The transcription factor Con7p is a central regulator of infection-related morphogenesis in the rice blast fungus Magnaporthe grisea. Mol. Microbiol. 2007, 64, 293–307. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Sun, J.; Kang, Z.; Ding, S.; Xu, J.-R.; Peng, Y.-L. A novel protein Com1 is required for normal conidium morphology and full virulence in Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2010, 23, 112–123. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, J.; Xie, X.; Liu, S.; Jiang, Y.; Pei, M.; Wu, Q.; Qi, P.; Du, L.; Peng, B.; et al. Characterization of two infection-induced transcription factors of Magnaporthe oryzae reveals their roles in regulating early infection and effector expression. Mol. Plant Pathol. 2022, 23, 1200–1213. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.Y.; Kim, K.S.; Rho, H.S.; Chi, M.H.; Choi, J.; Park, J.; Kong, S.; Park, J.; Goh, J.; et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae. PLoS Genet. 2009, 5, e1000757. [Google Scholar] [CrossRef]
- Matheis, S.; Yemelin, A.; Scheps, D.; Andresen, K.; Jacob, S.; Thines, E.; Foster, A.J. Functions of the Magnaporthe oryzae Flb3p and Flb4p transcription factors in the regulation of conidiation. Microbiol. Res. 2017, 196, 106–117. [Google Scholar] [CrossRef]
- Park, S.-Y.; Choi, J.; Lim, S.-E.; Lee, G.-W.; Park, J.; Kim, Y.; Kong, S.; Kim, S.R.; Rho, H.-S.; Jeon, J. Global expression profiling of transcription factor genes provides new insights into pathogenicity and stress responses in the rice blast fungus. PLoS Pathog. 2013, 9, e1003350. [Google Scholar] [CrossRef]
- Chi, M.-H.; Park, S.-Y. Identification of differentially expressed genes by cDNA-AFLP in Magnaporthe oryzae. Res. Plant Dis. 2019, 25, 205–212. [Google Scholar]
- Oberholzer, U.; Collart, M. Characterization of NOT5 that encodes a new component of the Not protein complex. Gene 1998, 207, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Franck, W.L.; Han, S.-O.; Shows, A.; Gokce, E.; Muddiman, D.C.; Dean, R.A. Polyubiquitin is required for growth, development and pathogenicity in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2012, 7, e42868. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, M.; Liu, X.; Yin, Z.; Sun, Y.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. Heat-shock proteins MoSsb1, MoSsz1, and MoZuo1 attenuate MoMkk1-mediated cell-wall integrity signaling and are important for growth and pathogenicity of Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2018, 31, 1211–1221. [Google Scholar] [CrossRef]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar]
- Choi, J.; Cheong, K.; Jung, K.; Jeon, J.; Lee, G.-W.; Kang, S.; Kim, S.; Lee, Y.-W.; Lee, Y.-H. CFGP 2.0: A versatile web-based platform for supporting comparative and evolutionary genomics of fungi and Oomycetes. Nucleic Acids Res. 2013, 41, D714–D719. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Gritz, L.; Davies, J. Plasmid-encoded hygromycin B resistance: The sequence of hygromycin B phosphotransferase gene and its expression in Escherichia coli and Saccharomyces cerevisiae. Gene 1983, 25, 179–188. [Google Scholar] [CrossRef]
- Leung, H.; Lehtinen, U.; Karjalainen, R.; Skinner, D.; Tooley, P.; Leong, S.; Ellingboe, A. Transformation of the rice blast fungus Magnaporthe grisea to hygromycin B resistance. Curr. Genet. 1990, 17, 409–411. [Google Scholar] [CrossRef]
- Haldane, D.J.; Robart, E. A comparison of calcofluor white, potassium hydroxide, and culture for the laboratory diagnosis of superficial fungal infection. Diagn. Microbiol. Infect. Dis. 1990, 13, 337–339. [Google Scholar] [CrossRef]
- Yi, M.; Chi, M.H.; Khang, C.H.; Park, S.Y.; Kang, S.; Valent, B.; Lee, Y.H. The ER chaperone LHS1 is involved in asexual development and rice infection by the blast fungus Magnaporthe oryzae. Plant Cell 2009, 21, 681–695. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Hyphal Growth A (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| CM | CaCl2 (0.2 M) | NaCl (0.5 M) | MnCl2 (0.02 M) | SDS (0.01%) | Congo Red (200 ppm) | CFW (2 mM) | H2O2 (2 mM) | 32 °C | |
| Wild-type | 69.3 ± 0.4 aB | 50.9 ± 1.9 a | 44.4 ± 1.7 a | 53.0 ± 1.7 a | 30.6 ± 0.3 a | 45.4 ± 0.8 a | 68.3 ± 0.2 a | 58.9 ± 1.5 a | 65.3 ± 0.2 a |
| Δmonot3-1 | 34.3 ± 0.3 b | 26.0 ± 0.4 b | 22.2 ± 0.5 b | 27.7 ± 0.8 b | 11.3 ± 0.6 b | 12.2 ± 0.2 b | 33.2 ± 0.4 b | 29.8 ± 1.0 b | 17.4 ± 0.4 b |
| Δmonot3-2 | 32.7 ± 0.6 b | 25.9 ± 0.4 b | 23.8 ± 0.8 b | 28.1 ± 0.2 b | 14.0 ± 0.5 b | 11.4 ± 0.6 b | 33.7 ± 0.5 b | 30.1 ± 0.1 b | 17.3 ± 0.3 b |
| MoNOT3 c-1 | 68.3 ± 0.6 a | 49.9 ± 1.3 a | 44.2 ± 1.3 a | 52.8 ± 1.1 a | 32.1 ± 0.6 a | 42.3 ± 1.9 a | 67.2 ± 0.4 a | 59.1 ± 0.2 a | 65.2 ± 0.3 a |
| MoNOT3 c-2 | 67.7 ± 0.2 a | 46.7 ± 0.8 a | 43.0 ± 1.4 a | 50.8 ± 1.1 a | 30.0 ± 1.0 a | 45.7 ± 1.2 a | 68.0 ± 0.5 a | 58.9 ± 1.4 a | 65.3 ± 0.3 a |
| Target | Name | Sequence (5′-3′) |
|---|---|---|
| Knockout | MoNOT3_5UF | ATATACAGGAGGCGGGGTCAGAGT |
| construct | MoNOT3_5UR | CCTCCACTAGCTCCAGCCAAGCCGGCTAGTTGTTGTTTCGGATGTCT |
| MoNOT3_3DF | GTTGGTGTCGATGTCAGCTCCGGAGGAGTGAATACGGCCAATAGC | |
| MoNOT3_3DR | GCACAGAGCCTAACATCAAACCCC | |
| MoNOT3_5UF_nested | GACACCATGAAACCACGCACTCTA | |
| MoNOT3_3DR_nested | GGACCAACAAGCTCCTCTCA | |
| hph gene | HygB_F | TCAGCTTCGATGTAGGAGGG |
| HygB_R | TTCTACACAGCCATCGGTCC | |
| Screening of | MoNOT3_ORF_F | TACATACGCCACTACCAACCATTA |
| transformant | MoNOT3_ORF_R | GATGCTGGAAGGTGGTAGATAGAT |
| qRT-PCR | Mo β-tubulin_F | ACAACTTCGTCTTCGGTCAG |
| Mo β-tubulin_R | GTGATCTGGAAACCCTGGAG | |
| MoNOT3_qRT_F | CAAATGGAGAGGTTCAAAGCG | |
| MoNOT3_qRT_R | TGTCGATCATGTTCCCAAGG | |
| COS1_qRT_F | TGCACCACGATCCCAGAGA | |
| COS1_qRT_R | GCGATGTTGTGCCGTTGTTCC | |
| COM1_qRT_F | GCCAGAGGTCCGCTATCAAA | |
| COM1_qRT_R | CGGGATCTCGTCACTGGATT | |
| CON7_qRT_F | TAAGGAGATCCGCAAAGAGT | |
| CON7_qRT_R | TAGCGTTGTAGTCGGGGAGT | |
| HOX2_qRT_F | TGGGGTTCTGCAGCCATGTT | |
| HOX2_qRT_R | GTCCCGTGGTGTTACGTTCTGG | |
| Polyubiqutin_qRT-F | CAACGCCTTATTTTCGCTGG | |
| Polyubiqutin_qRT-R | TCTTGCCCGTCAAAGTCTTG | |
| Ubiqutin_qRT-F | GCAAGTTCAACTGCGACAAG | |
| Ubiqutin_qRT-R | TCCACACTTTCTCTTCCTGC | |
| Ubiqutin40_qRT-F | AGTTGGCTGTGCTCAAGTAC | |
| Ubiqutin40_qRT-R | GTAGGTCAGGTGGCAACG | |
| Skp1-qRT-F | TGGGATCAGAAGTTCATGCAG | |
| Skp1-qRT-R | ATATCAAGGTAGTTGCTCGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Jo, M.; An, S.; Lee, Y.; Choi, E.D.; Jeong, M.-H.; Kim, K.-T.; Park, S.-Y. MoNOT3 Subunit Has Important Roles in Infection-Related Development and Stress Responses in Magnaporthe oryzae. Int. J. Mol. Sci. 2024, 25, 3290. https://doi.org/10.3390/ijms25063290
Kim Y, Jo M, An S, Lee Y, Choi ED, Jeong M-H, Kim K-T, Park S-Y. MoNOT3 Subunit Has Important Roles in Infection-Related Development and Stress Responses in Magnaporthe oryzae. International Journal of Molecular Sciences. 2024; 25(6):3290. https://doi.org/10.3390/ijms25063290
Chicago/Turabian StyleKim, Youngmin, Miju Jo, Sunmin An, Yerim Lee, Eu Ddeum Choi, Min-Hye Jeong, Ki-Tae Kim, and Sook-Young Park. 2024. "MoNOT3 Subunit Has Important Roles in Infection-Related Development and Stress Responses in Magnaporthe oryzae" International Journal of Molecular Sciences 25, no. 6: 3290. https://doi.org/10.3390/ijms25063290
APA StyleKim, Y., Jo, M., An, S., Lee, Y., Choi, E. D., Jeong, M. -H., Kim, K. -T., & Park, S. -Y. (2024). MoNOT3 Subunit Has Important Roles in Infection-Related Development and Stress Responses in Magnaporthe oryzae. International Journal of Molecular Sciences, 25(6), 3290. https://doi.org/10.3390/ijms25063290