
Integrated Transcriptomic and Proteomic Study of the Mechanism of Action of the Novel Small-Molecule Positive Allosteric Modulator 1 in Targeting PAC1-R for the Treatment of D-Gal-Induced Aging Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SPAM1 Prevents D-Gal-Mediated Reduction in the Number of Mouse Hippocampal Neurons
2.2. SPAM1 Ameliorated RGC-5 Cell Senescence
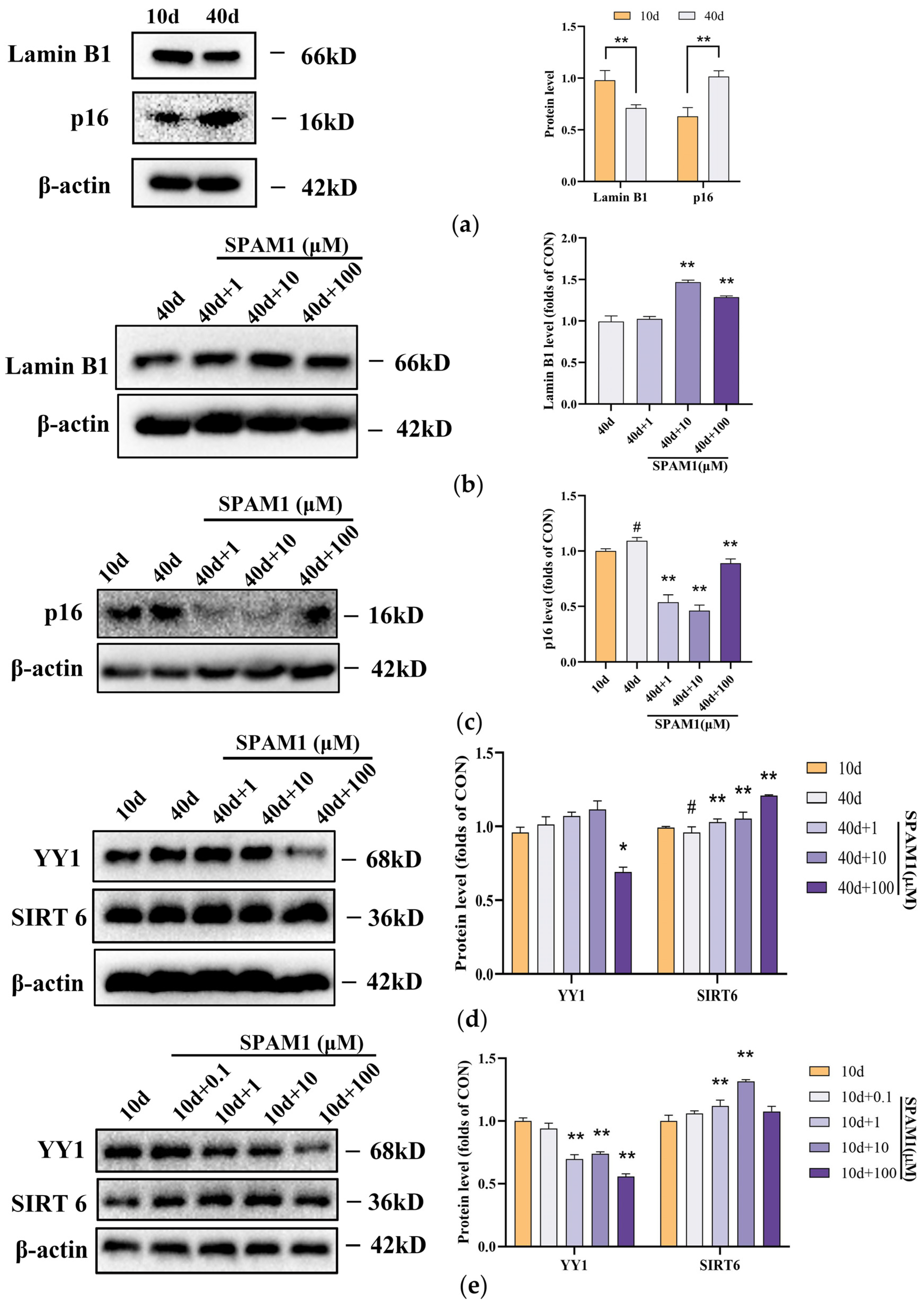
2.3. SPAM1 Treatment Up-Regulated the Expression of SIRT6 and Lamin B1 and Down-Regulated the Expression of YY1 and p16 in RGC-5 Cells and Mouse Brains
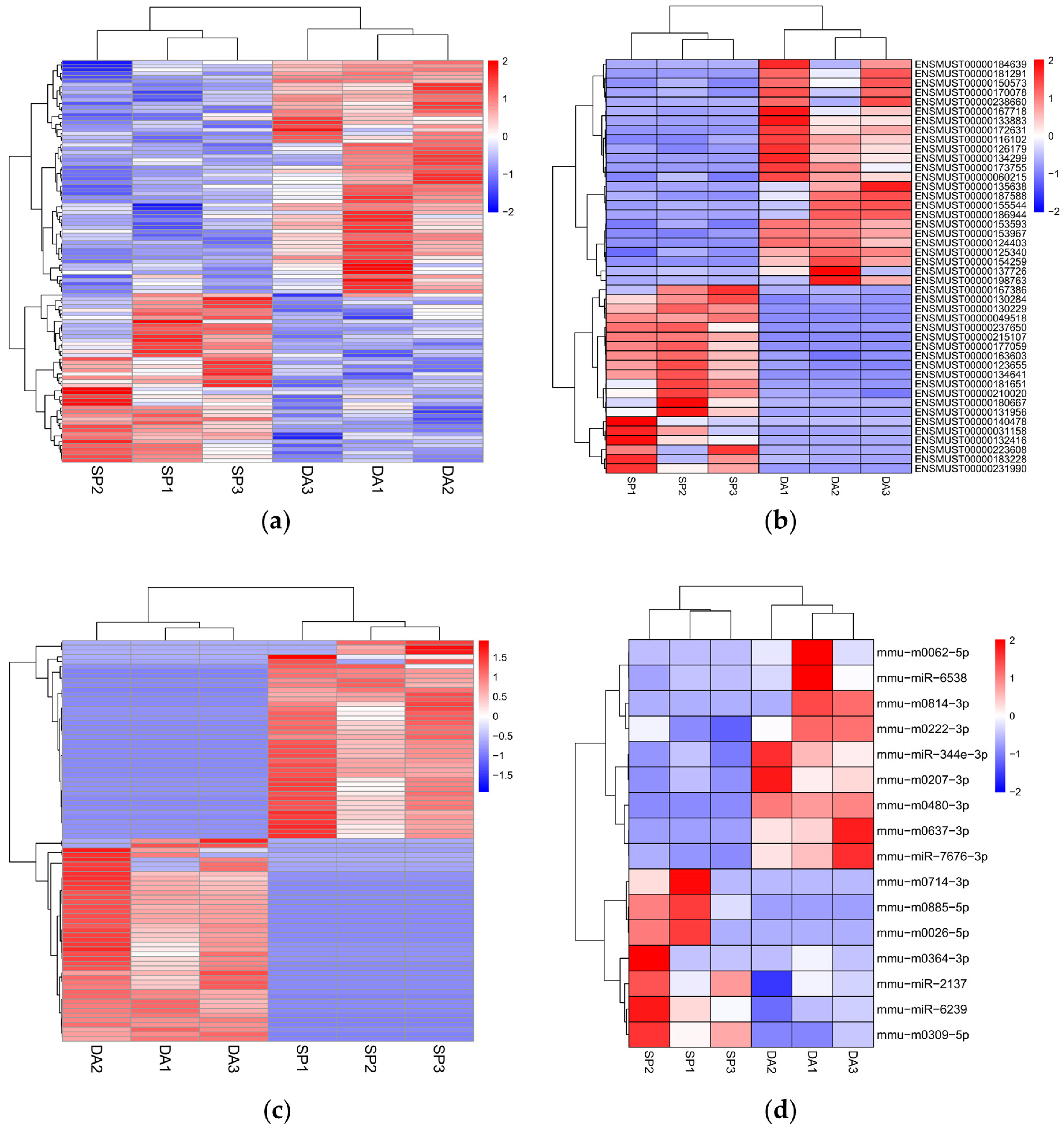
2.4. Analysis of Differentially Expressed mRNAs, lncRNAs, circRNAs, and miRNAs
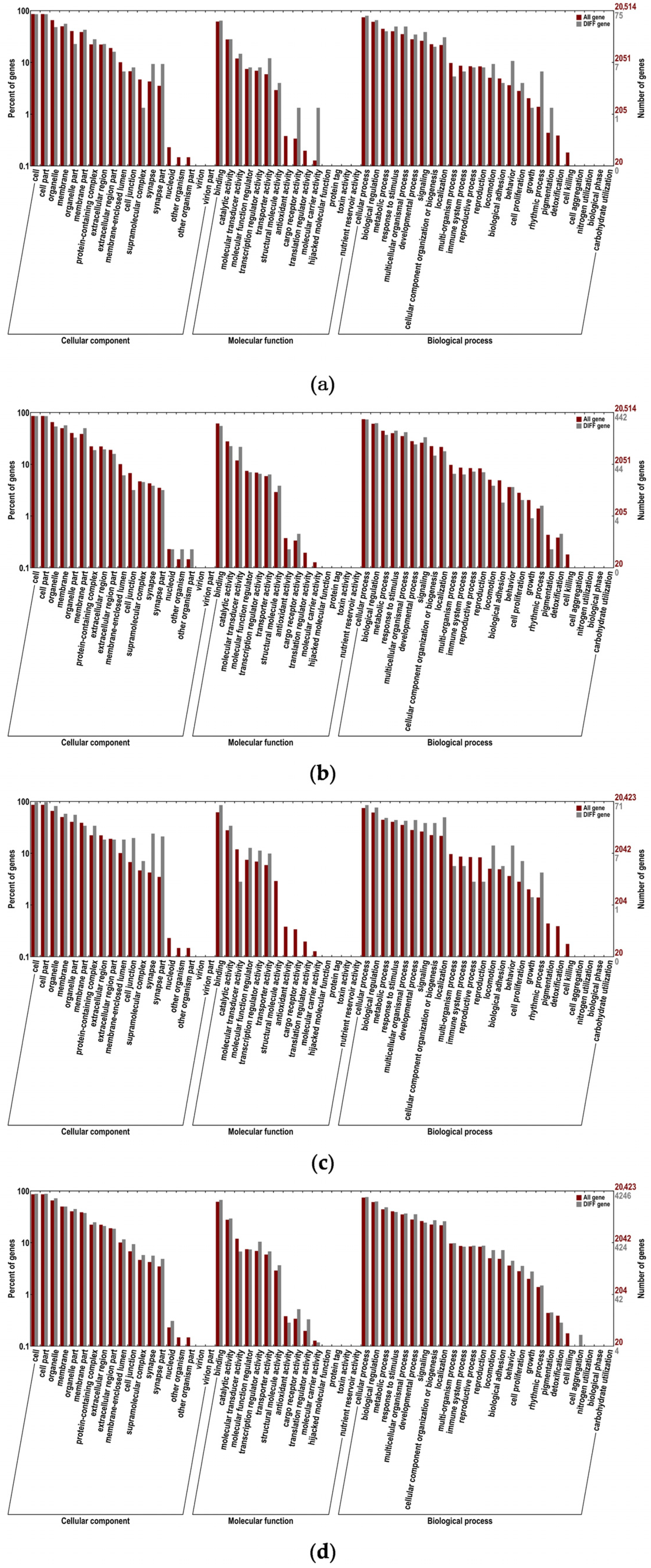
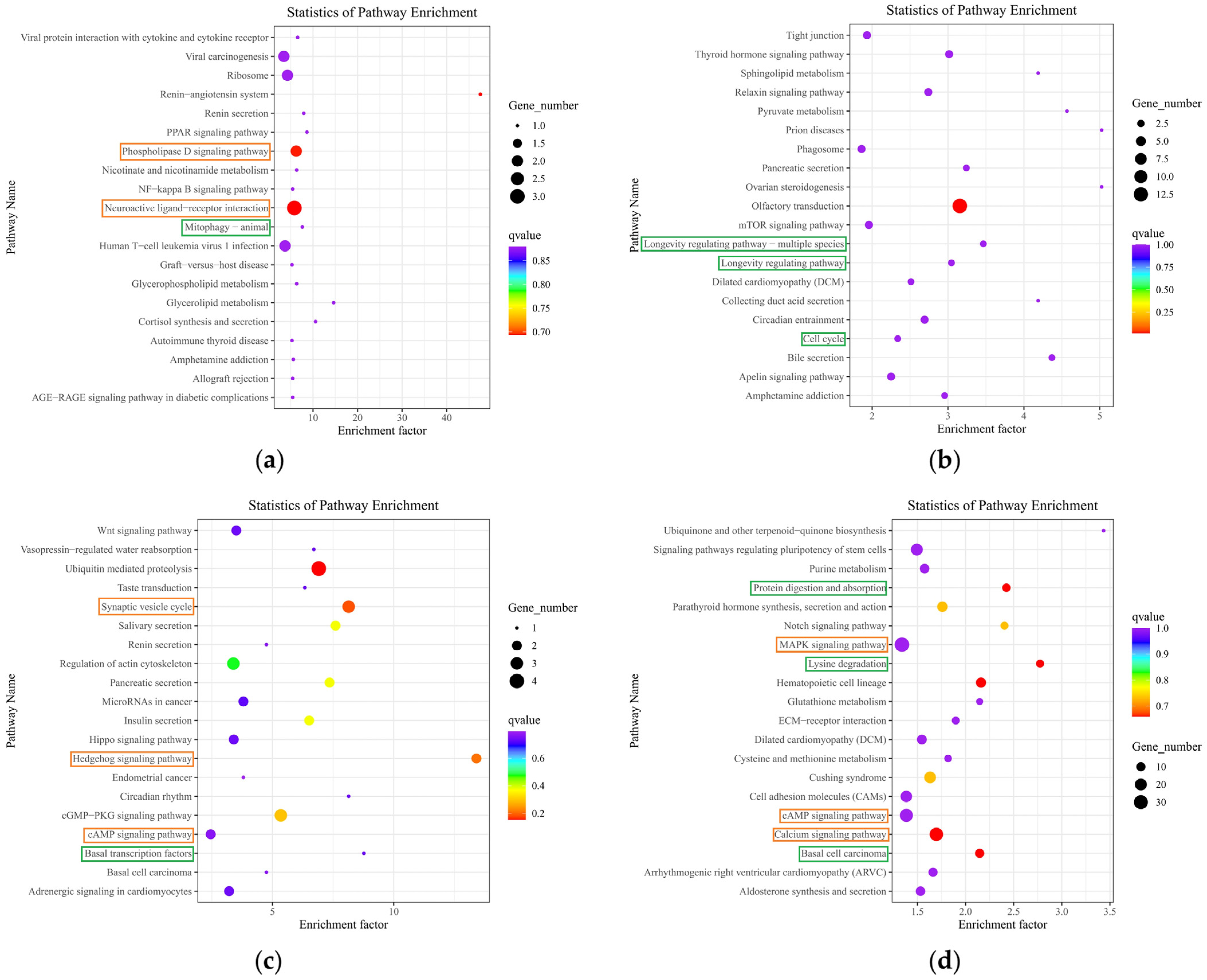
2.5. Functional Annotation and Enrichment Analysis of Differentially Expressed mRNAs
2.6. Functional Annotation and Enrichment Analysis of Differentially Expressed LncRNA Target Genes
2.7. Functional Annotation and Enrichment Analysis of Host Genes That Differentially Express circRNAs
2.8. Functional Annotation and Enrichment Analysis of Differentially Expressed miRNA Target Genes
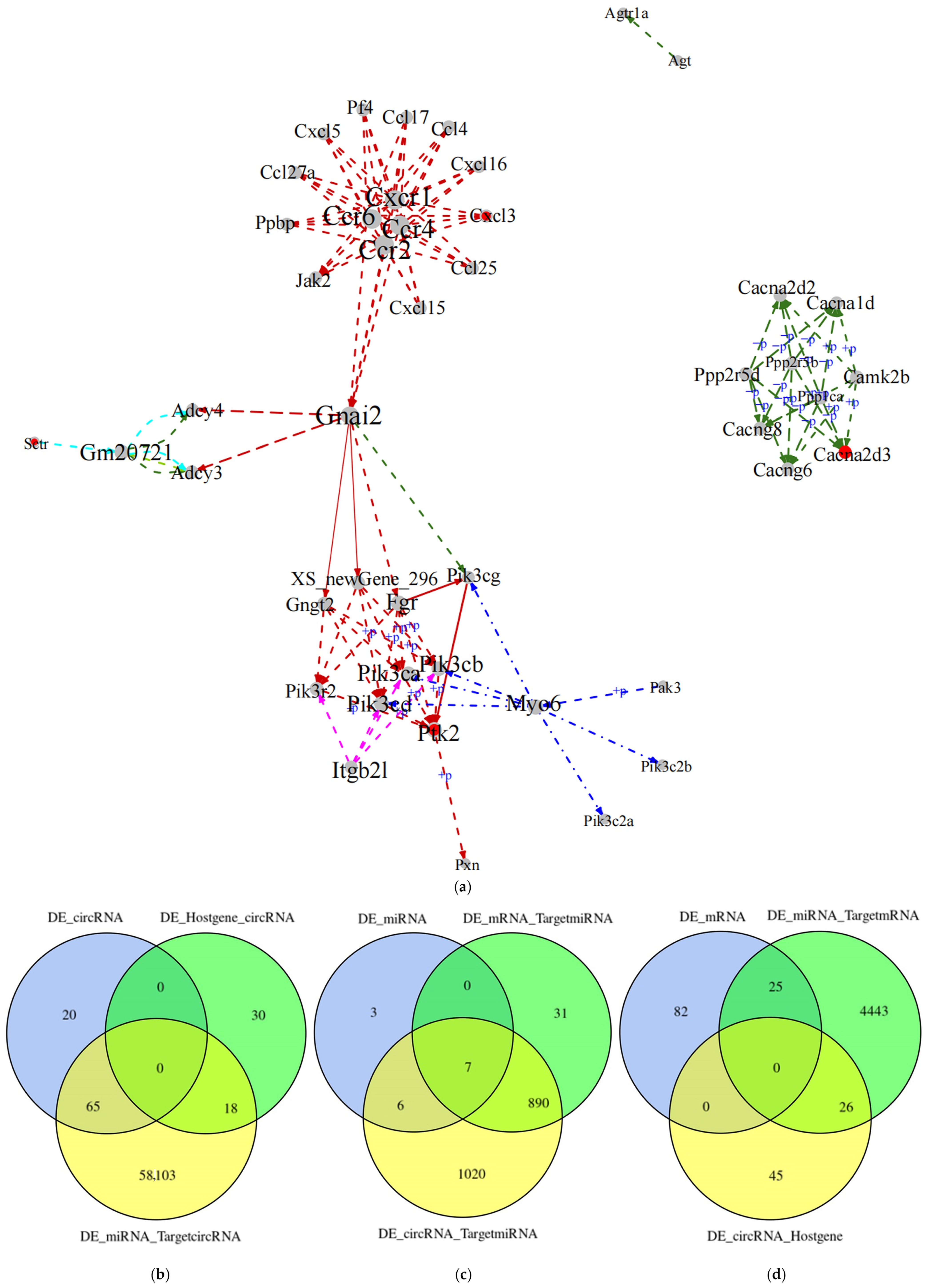
2.9. Whole-Transcriptome Association Analysis
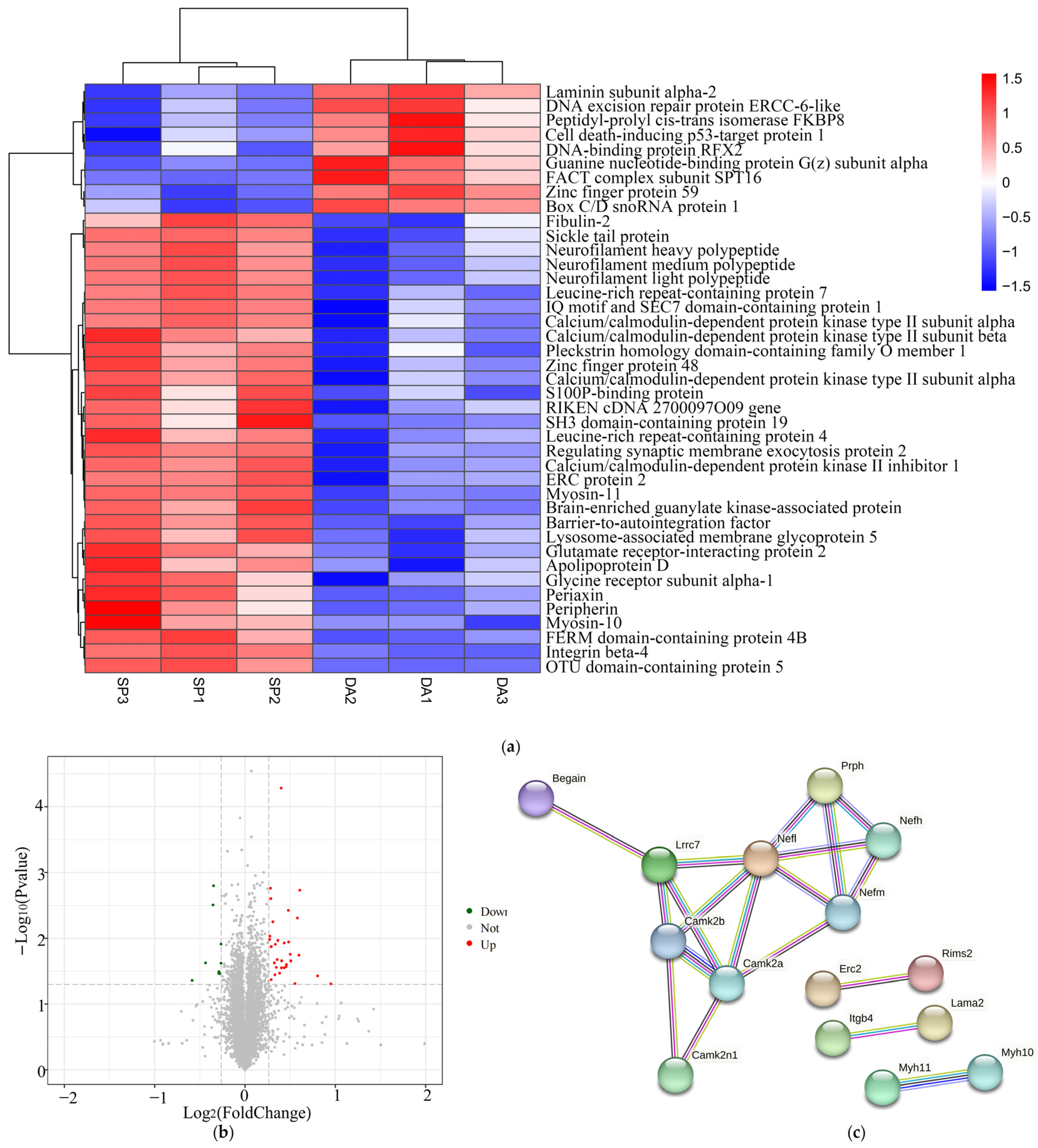
2.10. Analysis of Differentially Expressed Proteins
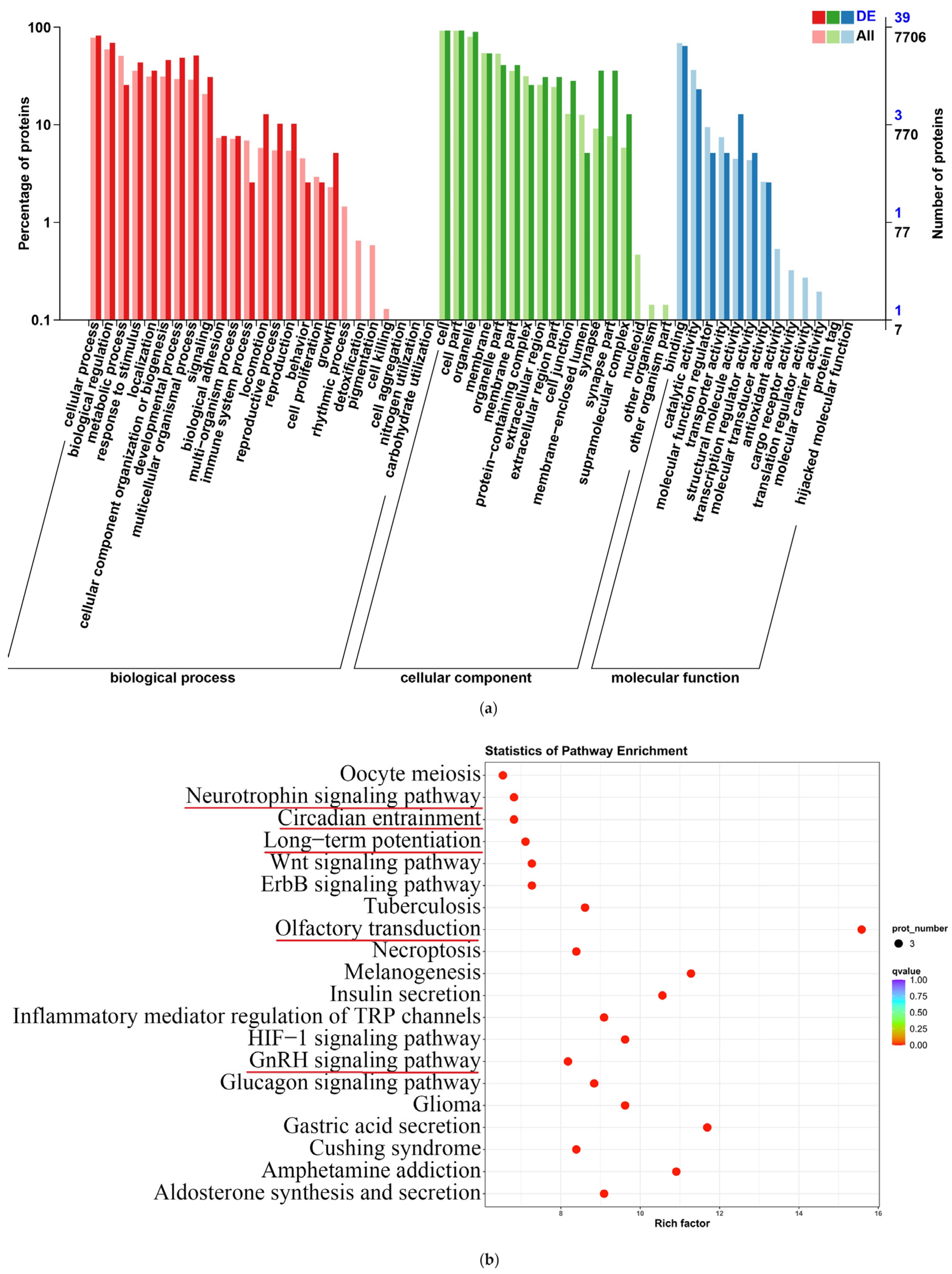
2.11. Functional Annotation and Enrichment Analysis of Differentially Expressed Proteins
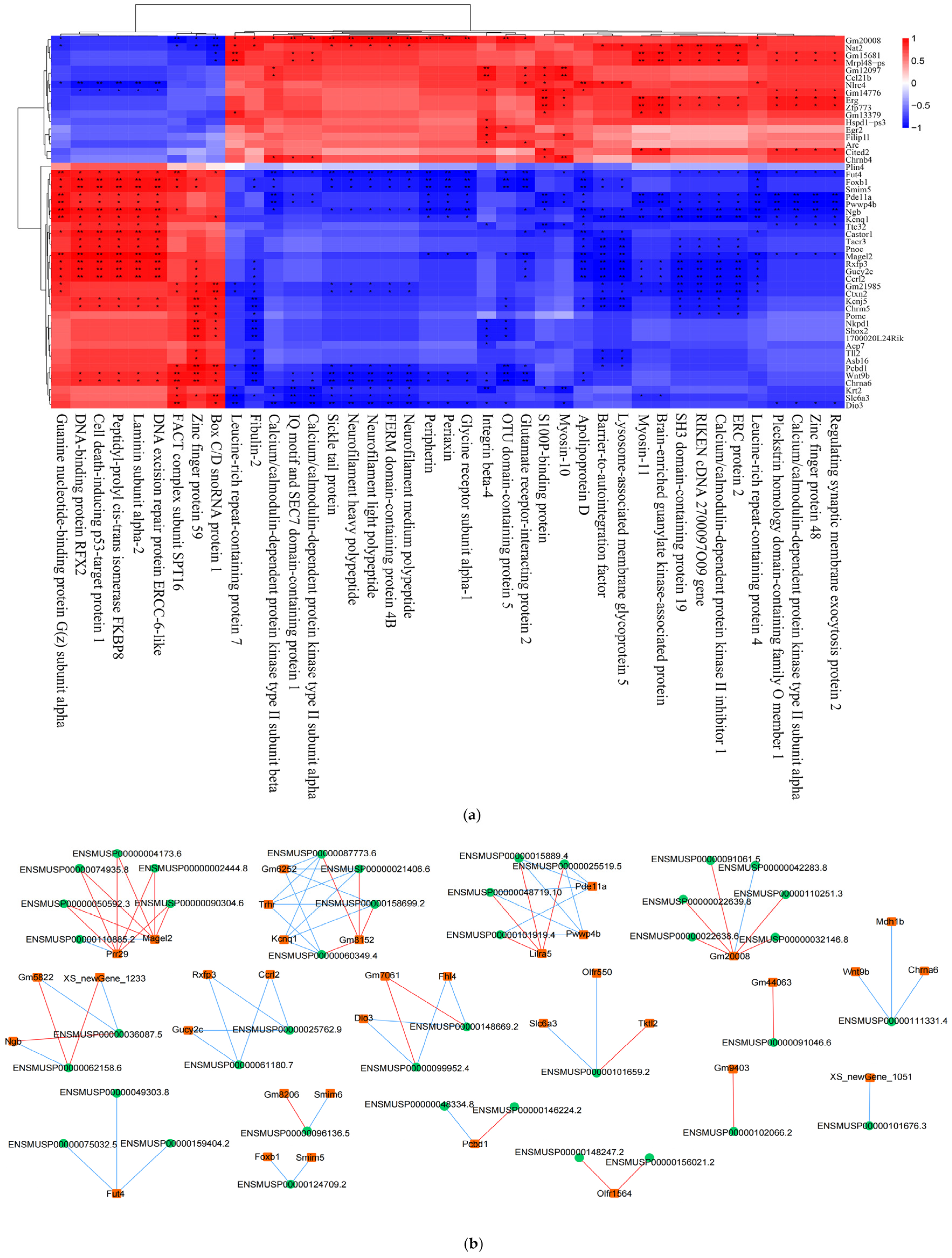
2.12. Combined Whole-Transcriptome and Proteome Analysis
2.13. Validation of Differentially Expressed Proteins
3. Discussion
4. Materials and Methods
4.1. Materials and Cell Lines
4.2. Grouping and Treatment of Animals
4.3. Tissue Preparation
4.4. Hematoxylin–Eosin (HE) Staining
4.5. Senescence-Associated β-Galactosidase Staining
4.6. Western Blotting Assays
4.7. Immunohistochemistry
4.8. Whole-Transcriptome Sequencing
4.9. Proteomic Analysis
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dautzenberg, F.M.; Mevenkamp, G.; Wille, S.; Hauger, R.L. N-terminal splice variants of the type I PACAP receptor: Isolation, characterization and ligand binding/selectivity determinants. J. Neuroendocrinol. 1999, 11, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef] [PubMed]
- Shioda, S.; Ohtaki, H.; Nakamachi, T.; Dohi, K.; Watanabe, J.; Nakajo, S.; Arata, S.; Kitamura, S.; Okuda, H.; Takenoya, F.; et al. Pleiotropic functions of PACAP in the CNS: Neuroprotection and neurodevelopment. Ann. N. Y. Acad. Sci. 2006, 1070, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Douiri, S.; Bahdoudi, S.; Hamdi, Y.; Cubi, R.; Basille, M.; Fournier, A.; Vaudry, H.; Tonon, M.C.; Amri, M.; Vaudry, D.; et al. Involvement of endogenous antioxidant systems in the protective activity of pituitary adenylate cyclase-activating polypeptide against hydrogen peroxide-induced oxidative damages in cultured rat astrocytes. J. Neurochem. 2016, 137, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Jiang, X.; Ji, R.; Meng, L.; Liu, F.; Chen, X.; Xin, Y. Therapeutic potential of PACAP for neurodegenerative diseases. Cell Mol. Biol. Lett. 2015, 20, 265–278. [Google Scholar] [CrossRef]
- Yu, R.; Zheng, L.; Cui, Y.; Zhang, H.; Ye, H. Doxycycline exerted neuroprotective activity by enhancing the activation of neuropeptide GPCR PAC1. Neuropharmacology 2016, 103, 1–15. [Google Scholar] [CrossRef]
- Song, S.; Wang, L.; Li, J.; Huang, X.; Yu, R. The allosteric modulation effects of doxycycline, minocycline, and their derivatives on the neuropeptide receptor PAC1-R. Acta Biochim. Biophys. Sin. 2019, 51, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Chen, S.; Tao, Z.; Zhang, H.; Yu, R. A novel small positive allosteric modulator of neuropeptide receptor PAC1-R exerts neuroprotective effects in MPTP mouse Parkinson’s disease model. Acta Biochim. Biophys. Sin. 2022, 54, 1349–1364. [Google Scholar] [CrossRef]
- Fan, G.; Chen, S.; Liang, L.; Zhang, H.; Yu, R. Novel Small Molecule Positive Allosteric Modulator SPAM1 Triggers the Nuclear Translocation of PAC1-R to Exert Neuroprotective Effects through Neuron-Restrictive Silencer Factor. Int. J. Mol. Sci. 2022, 23, 15996. [Google Scholar] [CrossRef]
- Matsuno, H.; Tsuchimine, S.; Fukuzato, N.; O’Hashi, K.; Kunugi, H.; Sohya, K. Sirtuin 6 is a regulator of dendrite morphogenesis in rat hippocampal neurons. Neurochem. Int. 2021, 145, 104959. [Google Scholar] [CrossRef]
- Garcia-Venzor, A.; Toiber, D. SIRT6 Through the Brain Evolution, Development, and Aging. Front. Aging Neurosci. 2021, 13, 747989. [Google Scholar] [CrossRef]
- Kaluski, S.; Portillo, M.; Besnard, A.; Stein, D.; Einav, M.; Zhong, L.; Ueberham, U.; Arendt, T.; Mostoslavsky, R.; Sahay, A.; et al. Neuroprotective Functions for the Histone Deacetylase SIRT6. Cell Rep. 2017, 18, 3052–3062. [Google Scholar] [CrossRef]
- Shi, Y.; Seto, E.; Chang, L.S.; Shenk, T. Transcriptional repression by YY1, a human GLI-Kruppel-related protein, and relief of repression by adenovirus E1A protein. Cell 1991, 67, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.; Mizrahi, A.; Golova, A.; Saretzky, A.; Venzor, A.G.; Slobodnik, Z.; Kaluski, S.; Einav, M.; Khrameeva, E.; Toiber, D. Aging and pathological aging signatures of the brain: Through the focusing lens of SIRT6. Aging 2021, 13, 6420–6441. [Google Scholar] [CrossRef]
- Pajarillo, E.; Nyarko-Danquah, I.; Digman, A.; Vied, C.; Son, D.S.; Lee, J.; Aschner, M.; Lee, E. Astrocytic Yin Yang 1 is critical for murine brain development and protection against apoptosis, oxidative stress, and inflammation. Glia 2023, 71, 450–466. [Google Scholar] [CrossRef]
- Koufi, F.D.; Neri, I.; Ramazzotti, G.; Rusciano, I.; Mongiorgi, S.; Marvi, M.V.; Fazio, A.; Shin, M.; Kosodo, Y.; Cani, I.; et al. Lamin B1 as a key modulator of the developing and aging brain. Front. Cell Neurosci. 2023, 17, 1263310. [Google Scholar] [CrossRef] [PubMed]
- Bin Imtiaz, M.K.; Jaeger, B.N.; Bottes, S.; Machado, R.A.C.; Vidmar, M.; Moore, D.L.; Jessberger, S. Declining lamin B1 expression mediates age-dependent decreases of hippocampal stem cell activity. Cell Stem Cell 2021, 28, 967–977.e8. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Sun, D.; Mei, L.; Xiong, W.C. Dorsal Dentate Gyrus, a Key Regulator for Mood and Psychiatric Disorders. Biol. Psychiatry 2023, 93, 1071–1080. [Google Scholar] [CrossRef]
- Ahmad, R.; Lahuna, O.; Sidibe, A.; Daulat, A.; Zhang, Q.; Luka, M.; Guillaume, J.L.; Gallet, S.; Guillonneau, F.; Hamroune, J.; et al. GPR50-Ctail cleavage and nuclear translocation: A new signal transduction mode for G protein-coupled receptors. Cell Mol. Life Sci. 2020, 77, 5189–5205. [Google Scholar] [CrossRef]
- Fan, G.; Tao, Z.; Chen, S.; Zhang, H.; Yu, R. Positive allosteric regulation of PAC1-R up-regulates PAC1-R and its specific ligand PACAP. Acta Biochim. Biophys. Sin. 2022, 54, 657–672. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Lin, Z.; Ouyang, Z.; Tao, Z.; Fan, G. Blue light induces the nuclear translocation of neuropeptide receptor PAC1-R associated with the up-regulation of PAC1-R its own in reactive oxygen species associated way. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129884. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; de Molliens, M.P.; Schneebeli, S.T.; Brewer, M.; Song, G.; Chatenet, D.; Braas, K.M.; May, V.; Li, J. Targeting the PAC1 Receptor for Neurological and Metabolic Disorders. Curr. Top. Med. Chem. 2019, 19, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.C.; Liu, J.H.; Wu, R.Y. Establishment of the mimetic aging effect in mice caused by D-galactose. Biogerontology 2003, 4, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Hainmueller, T.; Bartos, M. Dentate gyrus circuits for encoding, retrieval and discrimination of episodic memories. Nat. Rev. Neurosci. 2020, 21, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Mostoslavsky, R.; Chua, K.F.; Lombard, D.B.; Pang, W.W.; Fischer, M.R.; Gellon, L.; Liu, P.; Mostoslavsky, G.; Franco, S.; Murphy, M.M.; et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell 2006, 124, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, D.; Eremenko, E.; Stein, D.; Kaluski, S.; Jasinska, W.; Cosentino, C.; Martinez-Pastor, B.; Brotman, Y.; Mostoslavsky, R.; Khrameeva, E.; et al. SIRT6 is a key regulator of mitochondrial function in the brain. Cell Death Dis. 2023, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jin, K.; Jiao, J.; Liu, R.; Mou, T.; Chen, B.; Zhang, Z.; Jiang, C.; Zhao, H.; Wang, Z.; et al. YY1 (Yin-Yang 1), a transcription factor regulating systemic inflammation, is involved in cognitive impairment of depression. Psychiatry Clin. Neurosci. 2023, 77, 149–159. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Jabir, M.S.; Albuhadily, A.K.; Al-Gareeb, A.I.; Rafeeq, M.F. The link between metabolic syndrome and Alzheimer disease: A mutual relationship and long rigorous investigation. Ageing Res. Rev. 2023, 91, 102084. [Google Scholar] [CrossRef]
- Loera-Valencia, R.; Eroli, F.; Garcia-Ptacek, S.; Maioli, S. Brain Renin–Angiotensin System as Novel and Potential Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 10139. [Google Scholar] [CrossRef]
- Bottero, V.; Powers, D.; Yalamanchi, A.; Quinn, J.P.; Potashkin, J.A. Key Disease Mechanisms Linked to Alzheimer’s Disease in the Entorhinal Cortex. Int. J. Mol. Sci. 2021, 22, 3915. [Google Scholar] [CrossRef]
- Roy, A.; Derakhshan, F.; Wilson, R.J. Stress peptide PACAP engages multiple signaling pathways within the carotid body to initiate excitatory responses in respiratory and sympathetic chemosensory afferents. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R1070–R1084. [Google Scholar] [CrossRef]
- Hansel, D.E.; May, V.; Eipper, B.A.; Ronnett, G.V. Pituitary adenylyl cyclase-activating peptides and alpha-amidation in olfactory neurogenesis and neuronal survival in vitro. J. Neurosci. 2001, 21, 4625–4636. [Google Scholar] [CrossRef]
- Imai, N. Molecular and Cellular Neurobiology of Circadian and Circannual Rhythms in Migraine: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 10092. [Google Scholar] [CrossRef]
- Holland, P.R.; Barloese, M.; Fahrenkrug, J. PACAP in hypothalamic regulation of sleep and circadian rhythm: Importance for headache. J. Headache Pain 2018, 19, 20. [Google Scholar] [CrossRef]
- Vitiello, M.; Zullo, A.; Servillo, L.; Mancini, F.P.; Borriello, A.; Giovane, A.; Della Ragione, F.; D’Onofrio, N.; Balestrieri, M.L. Multiple pathways of SIRT6 at the crossroads in the control of longevity, cancer, and cardiovascular diseases. Ageing Res. Rev. 2017, 35, 301–311. [Google Scholar] [CrossRef]
- Patrick, M.B.; Omar, N.; Werner, C.T.; Mitra, S.; Jarome, T.J. The ubiquitin-proteasome system and learning-dependent synaptic plasticity—A 10 year update. Neurosci. Biobehav. Rev. 2023, 152, 105280. [Google Scholar] [CrossRef]
- Hegde, A.N.; Smith, S.G.; Duke, L.M.; Pourquoi, A.; Vaz, S. Perturbations of Ubiquitin-Proteasome-Mediated Proteolysis in Aging and Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 324. [Google Scholar] [CrossRef]
- Cohen, J.R.; Resnick, D.Z.; Niewiadomski, P.; Dong, H.; Liau, L.M.; Waschek, J.A. Pituitary adenylyl cyclase activating polypeptide inhibits gli1 gene expression and proliferation in primary medulloblastoma derived tumorsphere cultures. BMC Cancer 2010, 10, 676. [Google Scholar] [CrossRef]
- Waschek, J.A.; Dicicco-Bloom, E.; Nicot, A.; Lelievre, V. Hedgehog signaling: New targets for GPCRs coupled to cAMP and protein kinase A. Ann. N. Y. Acad. Sci. 2006, 1070, 120–128. [Google Scholar] [CrossRef]
- Zheng, R.; Ghirlando, R.; Lee, M.S.; Mizuuchi, K.; Krause, M.; Craigie, R. Barrier-to-autointegration factor (BAF) bridges DNA in a discrete, higher-order nucleoprotein complex. Proc. Natl. Acad. Sci. 2000, 97, 8997–9002. [Google Scholar] [CrossRef]
- Burgess, J.T.; Cheong, C.M.; Suraweera, A.; Sobanski, T.; Beard, S.; Dave, K.; Rose, M.; Boucher, D.; Croft, L.V.; Adams, M.N.; et al. Barrier-to-autointegration-factor (Banf1) modulates DNA double-strand break repair pathway choice via regulation of DNA-dependent kinase (DNA-PK) activity. Nucleic Acids Res. 2021, 49, 3294–3307. [Google Scholar] [CrossRef]
- Halfmann, C.T.; Sears, R.M.; Katiyar, A.; Busselman, B.W.; Aman, L.K.; Zhang, Q.; O’Bryan, C.S.; Angelini, T.E.; Lele, T.P.; Roux, K.J. Repair of nuclear ruptures requires barrier-to-autointegration factor. J. Cell Biol. 2019, 218, 2136–2149. [Google Scholar] [CrossRef]
- Puente, X.S.; Quesada, V.; Osorio, F.G.; Cabanillas, R.; Cadinanos, J.; Fraile, J.M.; Ordonez, G.R.; Puente, D.A.; Gutierrez-Fernandez, A.; Fanjul-Fernandez, M.; et al. Exome sequencing and functional analysis identifies BANF1 mutation as the cause of a hereditary progeroid syndrome. Am. J. Hum. Genet. 2011, 88, 650–656. [Google Scholar] [CrossRef]
- Yasuda, R.; Hayashi, Y.; Hell, J.W. CaMKII: A central molecular organizer of synaptic plasticity, learning and memory. Nat. Rev. Neurosci. 2022, 23, 666–682. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.V.; Veeranna; Nixon, R.A. Neurofilaments and Neurofilament Proteins in Health and Disease. Cold Spring Harb Perspect. Biol. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Tzeng, S.F.; Tsai, M.J.; Hung, S.C.; Cheng, H. Neuronal morphological change of size-sieved stem cells induced by neurotrophic stimuli. Neurosci. Lett. 2004, 367, 23–28. [Google Scholar] [CrossRef]
- Heraud, C.; Hilairet, S.; Muller, J.M.; Leterrier, J.F.; Chadeneau, C. Neuritogenesis induced by vasoactive intestinal peptide, pituitary adenylate cyclase-activating polypeptide, and peptide histidine methionine in SH-SY5y cells is associated with regulated expression of cytoskeleton mRNAs and proteins. J. Neurosci. Res. 2004, 75, 320–329. [Google Scholar] [CrossRef]
- Kosacka, J.; Schroder, T.; Bechmann, I.; Kloting, N.; Nowicki, M.; Mittag, A.; Gericke, M.; Spanel-Borowski, K.; Bluher, M. PACAP up-regulates the expression of apolipoprotein D in 3T3-L1 adipocytes. DRG/3T3-L1 co-cultures study. Neurosci. Res. 2011, 69, 8–16. [Google Scholar] [CrossRef]
- Wu, M.; Huang, H.; Chen, Q.; Li, D.; Zheng, Z.; Xiong, W.; Zhou, Y.; Li, X.; Zhou, M.; Lu, J.; et al. Leucine-rich repeat C4 protein is involved in nervous tissue development and neurite outgrowth, and induction of glioma cell differentiation. Acta Biochim. Biophys. Sin. 2007, 39, 731–738. [Google Scholar] [CrossRef]
- Halvorson, L.M. PACAP modulates GnRH signaling in gonadotropes. Mol. Cell Endocrinol. 2014, 385, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, L.; Chen, S.; Su, W.; Zhang, H.; Yu, R. Integrated Transcriptomic and Proteomic Study of the Mechanism of Action of the Novel Small-Molecule Positive Allosteric Modulator 1 in Targeting PAC1-R for the Treatment of D-Gal-Induced Aging Mice. Int. J. Mol. Sci. 2024, 25, 3872. https://doi.org/10.3390/ijms25073872
Liang L, Chen S, Su W, Zhang H, Yu R. Integrated Transcriptomic and Proteomic Study of the Mechanism of Action of the Novel Small-Molecule Positive Allosteric Modulator 1 in Targeting PAC1-R for the Treatment of D-Gal-Induced Aging Mice. International Journal of Molecular Sciences. 2024; 25(7):3872. https://doi.org/10.3390/ijms25073872
Chicago/Turabian StyleLiang, Lili, Shang Chen, Wanlin Su, Huahua Zhang, and Rongjie Yu. 2024. "Integrated Transcriptomic and Proteomic Study of the Mechanism of Action of the Novel Small-Molecule Positive Allosteric Modulator 1 in Targeting PAC1-R for the Treatment of D-Gal-Induced Aging Mice" International Journal of Molecular Sciences 25, no. 7: 3872. https://doi.org/10.3390/ijms25073872
APA StyleLiang, L., Chen, S., Su, W., Zhang, H., & Yu, R. (2024). Integrated Transcriptomic and Proteomic Study of the Mechanism of Action of the Novel Small-Molecule Positive Allosteric Modulator 1 in Targeting PAC1-R for the Treatment of D-Gal-Induced Aging Mice. International Journal of Molecular Sciences, 25(7), 3872. https://doi.org/10.3390/ijms25073872